Analysis of Interacting Proteins of Aluminum Toxicity Response Factor ALS3 and CAD in Citrus

 and
and
Abstract
:1. Introduction
2. Results
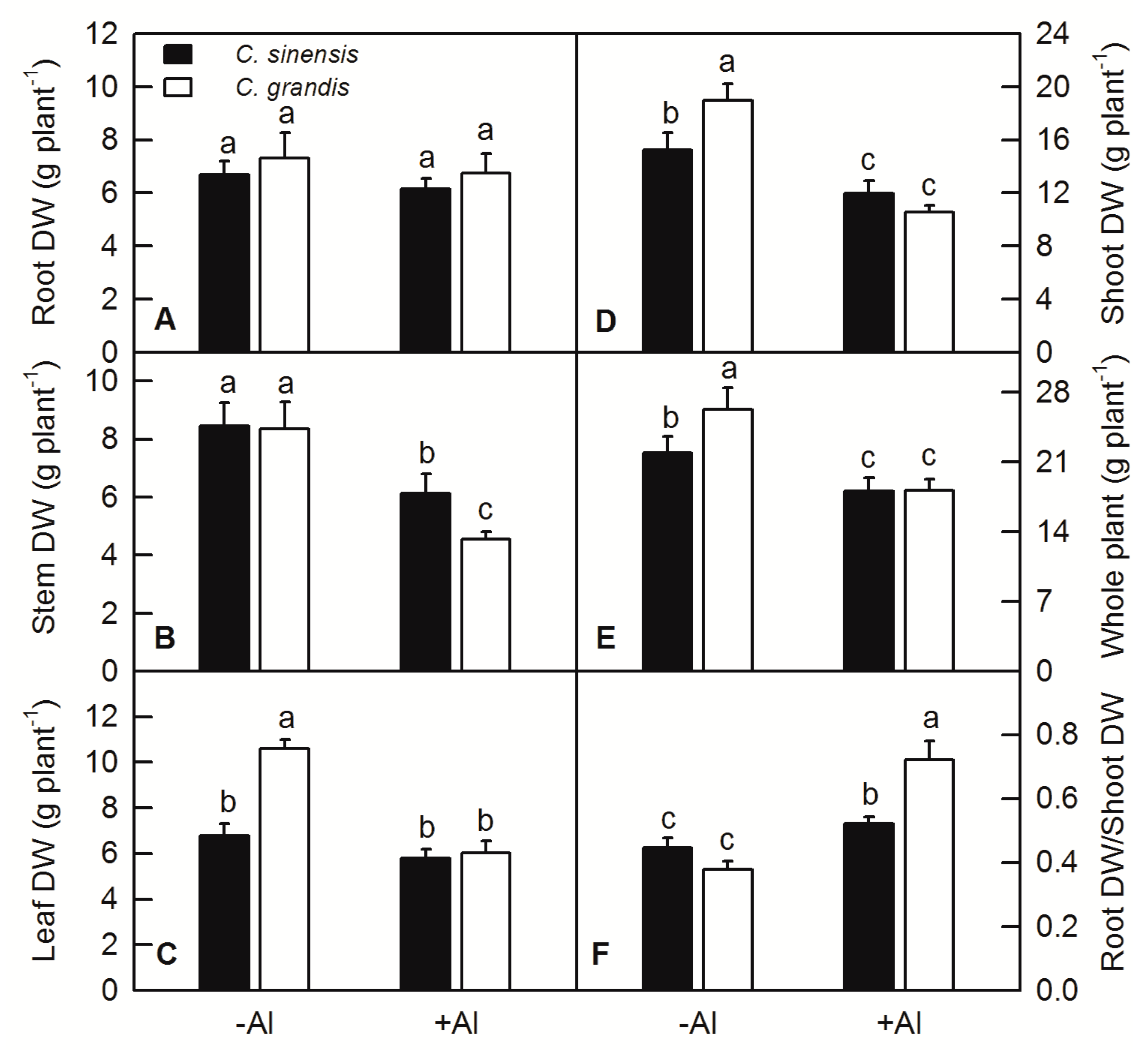
2.1. Effects of Al Treatment on the Plant DW and Root/Shoot in C. sinensis and C. grandis
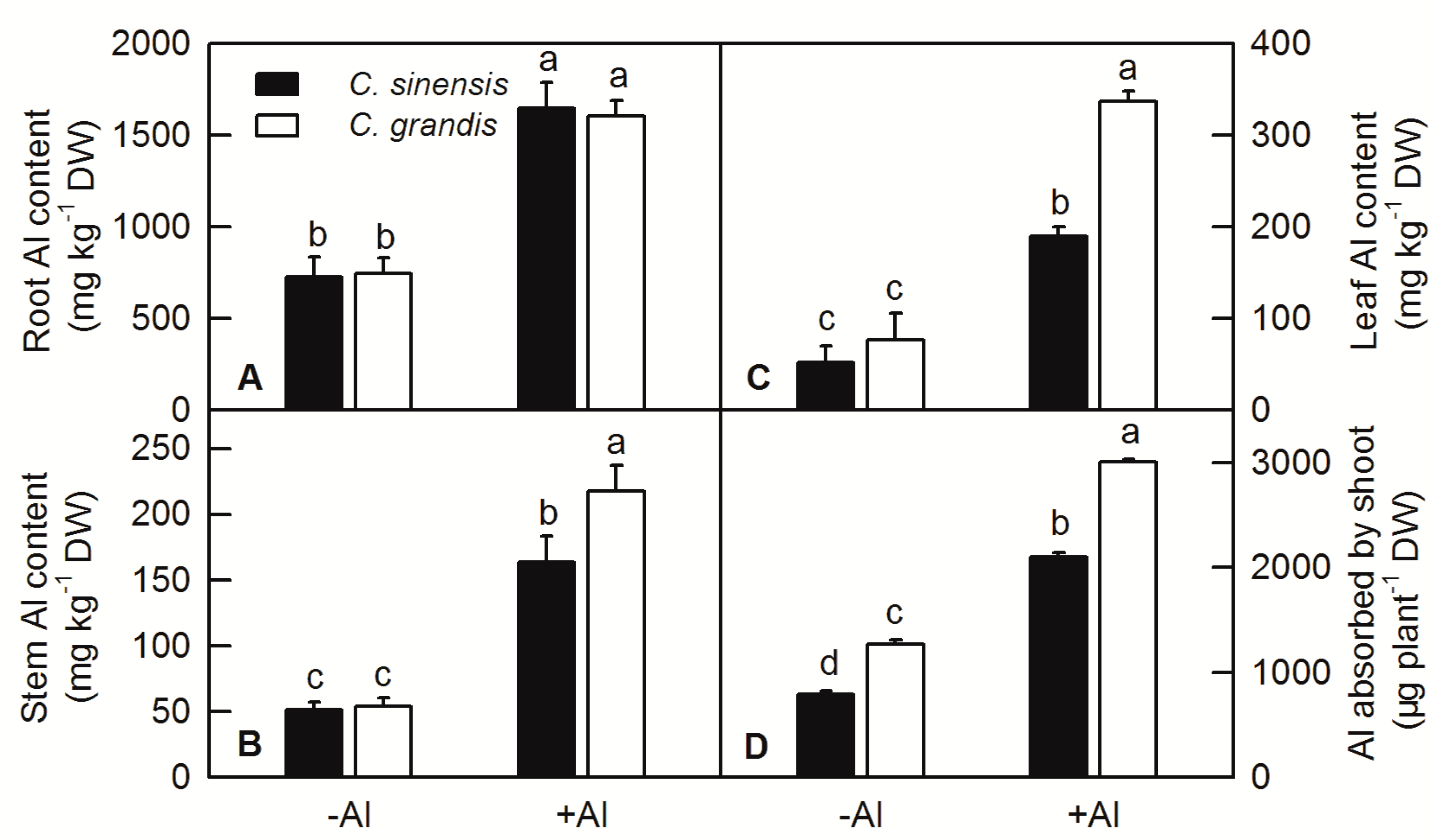
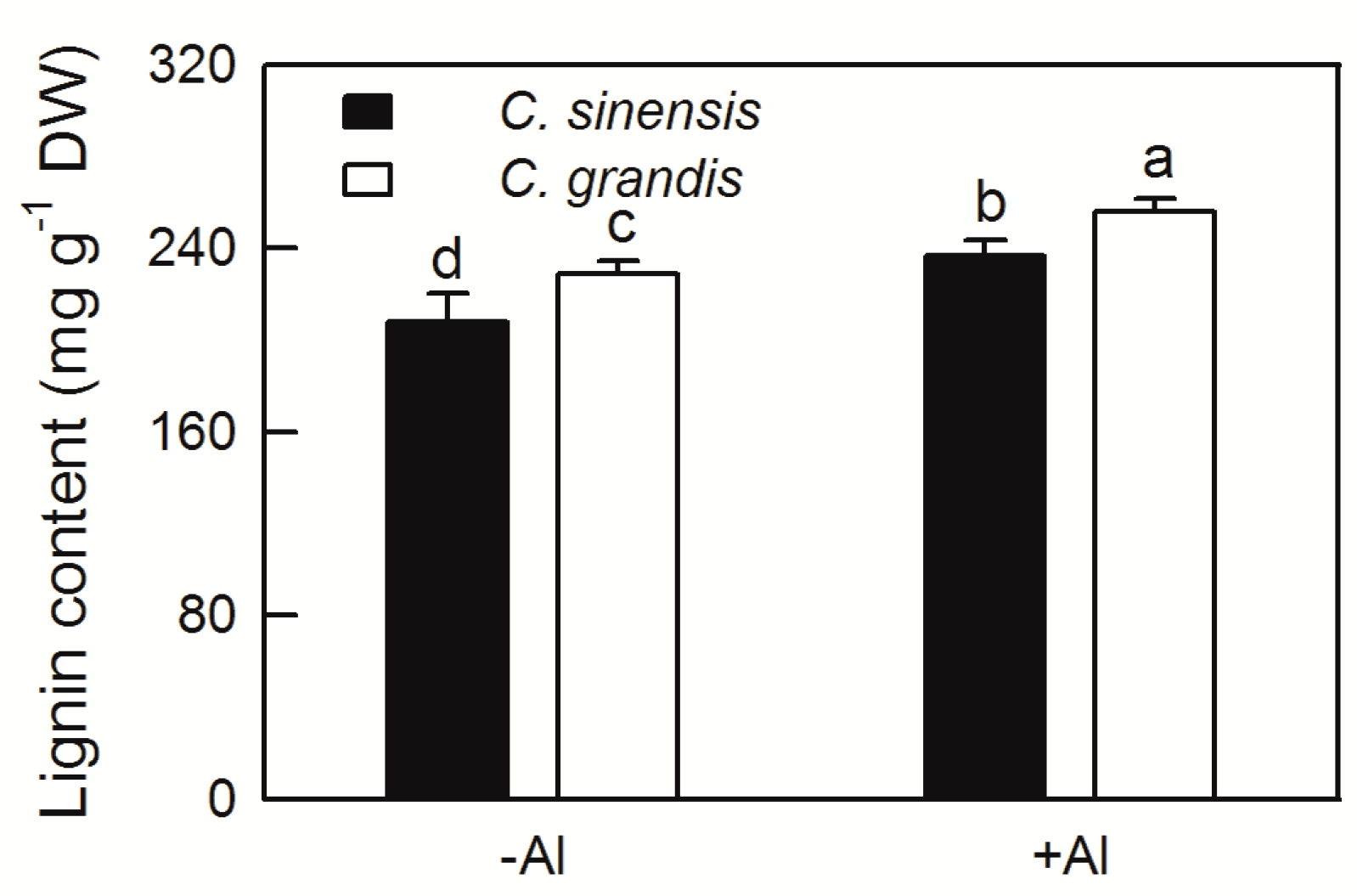
2.2. Effects of Al Treatment on Al Content and Root Lignin Content in Citrus
2.3. Expression of ALS3 and CAD Genes in Citrus under Al Treatment
2.4. Yeast Mating and Screening of Positive Colony
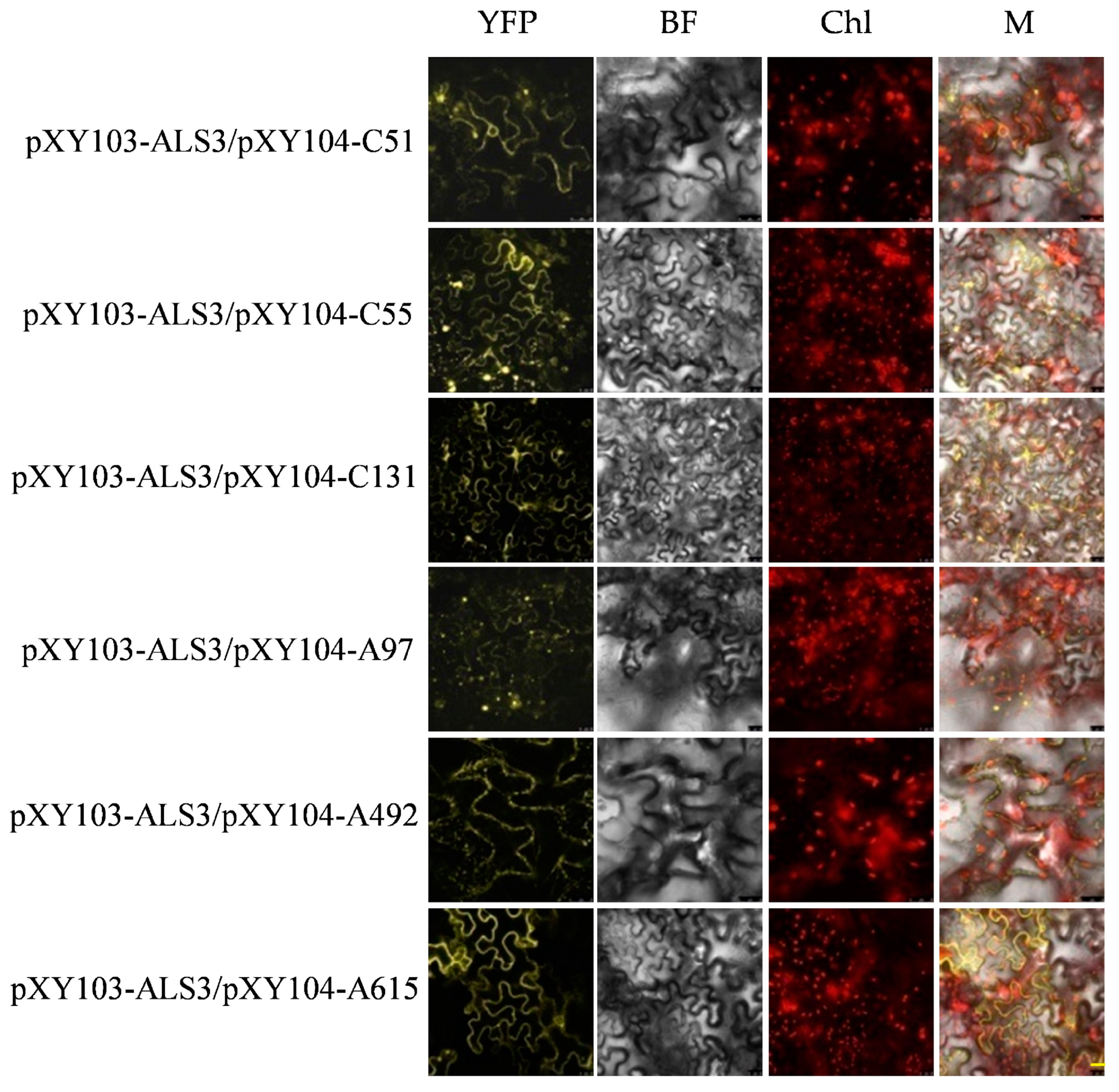
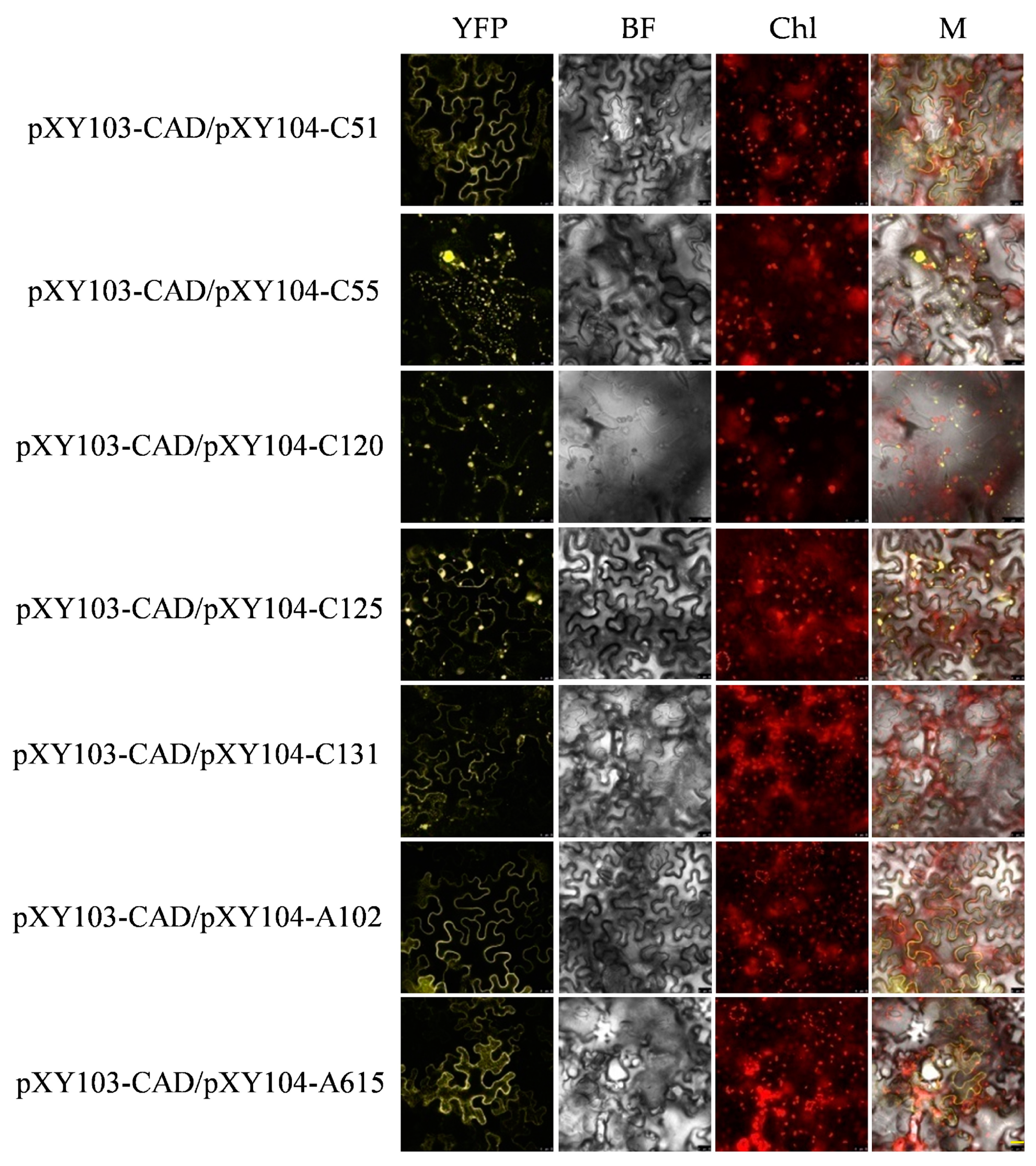
2.5. Verification of Candidate Proteins of ALS3 and CAD by BiFC
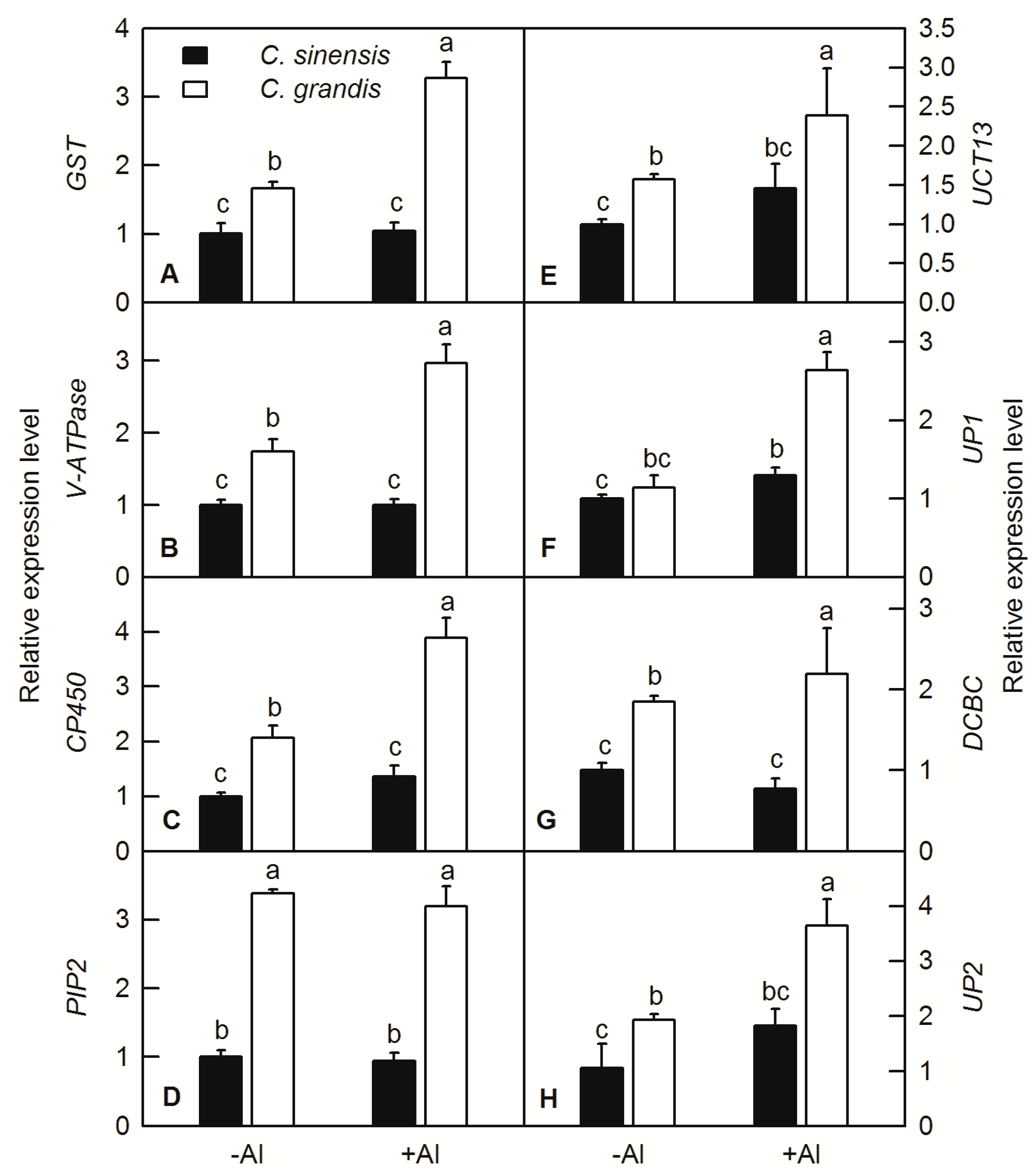
2.6. RT-q PCR Analysis of ALS3 and CAD Interacting Protein Genes under Al Treatment
3. Discussion
3.1. Effects of Al Treatment on the Growth and Al Distribution of C. sinensis and C. grandis
3.2. Interacting Proteins of ALS3 and CAD in C. sinensis and C. grandis
4. Materials and Methods
4.1. Seedling Culture and Aluminum Treatments
4.2. Plant Dry Weight (DW), Al and Lignin Content in Roots of Citrus
4.3. RT-qPCR Analysis of ALS3 and CAD
4.4. Y2H cDNA Library Construction and Screening of Interacting Proteins of ALS3 and CAD
4.5. Verification of Candidate Interacting Proteins of ALS3 and CAD by BiFC Assay
4.6. RT-qPCR Analysis the Interacting Proteins of ALS3 and CAD
4.7. Experimental Design and Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Kinraide, T.B. Identity of the rhizotoxic aluminium species. Plant Soil 1991, 134, 167–178. [Google Scholar] [CrossRef]
- Delhaize, E.; Ryan, P.R. Aluminum toxicity and tolerance in plants. Plant Physiol. 1995, 107, 315. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.F.; Shen, R.; Nagao, S.; Tanimoto, E. Aluminum targets elongating cells by reducing cell wall extensibility in wheat roots. Plant Cell Physiol. 2004, 45, 583. [Google Scholar] [CrossRef] [PubMed]
- Kochian, L.V. Cellular mechanisms of aluminum toxicity and resistance in plants. Ann. Rev. Plant Physiol. Plant Mol. Biol. 1995, 46, 237–260. [Google Scholar] [CrossRef]
- Li, Y.Y.; Yang, J.L.; Zhang, Y.J.; Zheng, S.J. Disorganized distribution of homogalacturonan epitopes in cell walls as one possible mechanism for aluminium-induced root growth inhibition in maize. Ann. Bot. 2009, 104, 235–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kochian, L.V.; Piñeros, M.A.; Hoekenga, O.A. The physiology, genetics and molecular biology of plant aluminum resistance and toxicity. Plant Soil 2005, 274, 175–195. [Google Scholar] [CrossRef]
- Kochian, L.V.; Piñeros, M.A.; Liu, J.; Magalhaes, J.V. Plant adaptation to acid soils: the molecular basis for crop aluminum resistance. Annu. Rev. Plant Biol. 2015, 66, 571–598. [Google Scholar] [CrossRef]
- Huang, C.F.; Yamaji, N.; Mitani, N.; Yano, M.; Nagamura, Y.; Ma, J.F. A bacterial-type ABC transporter is involved in aluminum tolerance in rice. Plant Cell 2009, 21, 655–667. [Google Scholar] [CrossRef] [PubMed]
- Larsen, P.B.; Geisler, M.J.; Jones, C.A.; Williams, K.M.; Cancel, J.D. ALS3 encodes a phloem-localized ABC transporter-like protein that is required for aluminum tolerance in Arabidopsis. Plant J. 2005, 41, 353–363. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.F.; Yamaji, N.; Chen, Z.; Ma, J.F. A tonoplast-localized half-size ABC transporter is required for internal detoxification of aluminum in rice. Plant J. 2012, 69, 857–867. [Google Scholar] [CrossRef]
- Liu, J.; Piñeros, M.A.; Kochian, L.V. The role of aluminum sensing and signaling in plant aluminum resistance. J. Int. Plant Biol. 2014, 56, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Zhou, D.; Yang, Y.; Zhang, J.; Jiang, F.; Craft, E.; Thannhauser, T.W.; Kochian, L.V.; Liu, J. Quantitative iTRAQ proteomics revealed possible roles for antioxidant proteins in sorghum aluminum tolerance. Front. Plant Sci. 2017, 7, 2043. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.T.; Liu, J.W.; Wu, Y.M.; Qi, Y.P.; Wang, J.L.; Lai, N.W.; Ye, X.; Chen, L.S. Proteome profile analysis of boron-induced alleviation of aluminum-toxicity in Citrus grandis roots. Ecotox. Environ. Safe 2018, 162, 488–498. [Google Scholar] [CrossRef] [PubMed]
- Yabuta, Y.; Osada, R.; Morishita, T.; Nishizawa-Yokoi, A.; Tamoi, M.; Maruta, T.; Shigeoka, S. Involvement of Arabidopsis NAC transcription factor in the regulation of 20S and 26S proteasomes. Plant Sci. 2011, 181, 421–427. [Google Scholar] [CrossRef] [PubMed]
- Drewes, G.; Bouwmeester, T. Global approaches to protein-protein interactions. Curr. Opin. Cell Biol. 2003, 15, 199–205. [Google Scholar] [CrossRef]
- Kim, Y. Bimolecular Fluorescence Complementation (BiFC) analysis: Advances and recent applications for genome-wide interaction studies. J. Mol. Biol. 2015, 427, 2039–2055. [Google Scholar]
- Li, Y.; Han, M.Q.; Lin, F.; Ten, Y.; Lin, J.; Zhu, D.H.; Guo, P.; Weng, Y.B.; Chen, L.S. Soil chemical properties, ’Guanximiyou’ pummelo leaf mineral nutrient status and fruit quality in the southern region of Fujian province, China. J. Soil Sci. Plant Nut. 2015, 15, 263–269. [Google Scholar] [CrossRef]
- Diaz, J.V.; Anthon, G.E.; Barrett, D.M. Nonenzymatic degradation of citrus pectin and pectate during prolonged heating: effects of pH, temperature, and degree of methyl esterification. J. Agric. Food Chem. 2007, 55, 5131–5136. [Google Scholar] [CrossRef]
- Yang, L.T.; Jiang, H.X.; Tang, N.; Chen, L.S. Mechanisms of aluminum-tolerance in two species of citrus: secretion of organic acid anions and immobilization of aluminum by phosphorus in roots. Plant Sci. 2011, 180, 521–530. [Google Scholar] [CrossRef]
- Guo, P.; Qi, Y.P.; Yang, L.T.; Lai, N.W.; Ye, X.; Yang, Y.; Chen, L.S. Root adaptive responses to aluminum-treatment revealed by RNA-seq in two citrus species with different aluminum-tolerance. Front. Plant Sci. 2017, 8, 330. [Google Scholar] [CrossRef]
- Jiang, H.X.; Yang, L.T.; Qi, Y.P.; Lu, Y.B.; Huang, Z.R.; Chen, L.S. Root iTRAQ protein profile analysis of two Citrus species differing in aluminum-tolerance in response to long-term aluminum-toxicity. BMC Genom. 2015, 16, 949. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.Y.; Shen, R.F.; Wang, C.; Wang, W. Root protein profile changes induced by Al exposure in two rice cultivars differing in Al tolerance. J. Proteomics 2013, 78, 281–293. [Google Scholar] [CrossRef] [PubMed]
- He, G.; Zhang, J.; Hu, X.; Wu, J. Effect of aluminum toxicity and phosphorus deficiency on the growth and photosynthesis of oil tea (Camellia oleifera Abel.) seedlings in acidic red soils. Acta Physiol. Plant 2011, 33, 1285–1292. [Google Scholar] [CrossRef]
- Kumari, M.; Taylor, G.J.; Deyholos, M.K. Transcriptomic responses to aluminum stress in roots of Arabidopsis thaliana. Mol. Genet. Genomics 2008, 279, 339–357. [Google Scholar] [CrossRef] [PubMed]
- Oh, M.W.; Roy, S.K.; Kamal, A.H.; Cho, K.; Cho, S.W.; Park, C.S.; Choi, J.S.; Komatsu, S.; Woo, S.H. Proteome analysis of roots of wheat seedlings under aluminum stress. Mol. Biol. Rep. 2014, 41, 671–681. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, M.T. Phosphorus alleviates aluminum toxicity in Al-sensitive wheat seedlings. Commun. Soil Sci. Plan 2014, 45, 437–450. [Google Scholar] [CrossRef]
- Zhen, Y.; Qi, J.L.; Wang, S.S.; Su, J.; Xu, G.H.; Zhang, M.S.; Miao, L.; Peng, X.X.; Tian, D.C.; Yang, Y.H. Comparative proteome analysis of differentially expressed proteins induced by Al toxicity in soybean. Physiol. Plant. 2007, 131, 542–554. [Google Scholar] [CrossRef]
- Guo, P.; Li, Q.; Qi, Y.P.; Yang, L.T.; Ye, X.; Chen, H.H.; Chen, L.S. Sulfur-mediated-alleviation of aluminum-toxicity in Citrus grandis seedlings. Int. J. Mol. Sci. 2017, 18, 2570. [Google Scholar] [CrossRef]
- Janhunen, S.; Palomäki, V.; Holopainen, T. Aluminium causes nutrient imbalance and structural changes in the needles of Scots pine without inducing clear root injuries. Trees - Struct. Funct. 1995, 9, 134. [Google Scholar] [CrossRef]
- Wang, J.W.; Kao, C.H. Aluminum-inhibited root growth of rice seedlings is mediated through putrescine accumulation. Plant Soil 2006, 288, 373–381. [Google Scholar] [CrossRef]
- Passaia, G.; Margis-Pinheiro, M. Glutathione peroxidases as redox sensor proteins in plant cells. Plant Sci. 2015, 234, 22–26. [Google Scholar] [CrossRef] [PubMed]
- Nianiou-Obeidat, I.; Madesis, P.; Kissoudis, C.; Voulgari, G.; Chronopoulou, E.; Tsaftaris, A.; Labrou, N.E. Plant glutathione transferase-mediated stress tolerance: functions and biotechnological applications. Plant Cell Rep. 2017, 36, 791–805. [Google Scholar] [CrossRef] [PubMed]
- Cicero, L.L.; Madesis, P.; Tsaftaris, A.; Lo Piero, A.R. Tobacco plants over-expressing the sweet orange tau glutathione transferases (CsGSTUs) acquire tolerance to the diphenyl ether herbicide fluorodifen and to salt and drought stresses. Phytochemistry 2015, 116, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Roxas, V.P.; Lodhi, S.A.; Garrett, D.K.; Mahan, J.R.; Allen, R.D. Stress tolerance in transgenic tobacco seedlings that overexpress glutathione s-transferase/glutathione peroxidase. Plant Cell Physiol. 2000, 41, 1229. [Google Scholar] [CrossRef] [PubMed]
- Ratajczak, R. Structure, function and regulation of the plant vacuolar H(+)-translocating ATPase. Biochim. Biophys. Acta 2000, 1465, 17–36. [Google Scholar] [CrossRef]
- Briskin, D.P.; Hanson, J.B. How does the plant plasma membrane H+-ATPase pump protons? J. Exp. Bot. 1992, 43, 269–289. [Google Scholar] [CrossRef]
- Padmanaban, S.; Lin, X.; Perera, I.; Kawamura, Y.; Sze, H. Differential expression of vacuolar H+-ATPase subunit c genes in tissues active in membrane trafficking and their roles in plant growth as revealed by RNAi. Plant Physiol. 2004, 134, 1514–1526. [Google Scholar] [CrossRef] [PubMed]
- Herman, E.M.; Li, X.; Su, R.T.; Larsen, P.; Hsu, H.; Sze, H. Vacuolar-type H+-ATPases are associated with the endoplasmic reticulum and provacuoles of root tip cells. Plant Physiol. 1994, 106, 1313–1324. [Google Scholar] [CrossRef]
- Sze, H.; Schumacher, K.; Müller, M.L.; Padmanaban, S.; Taiz, L. A simple nomenclature for a complex proton pump: VHA genes encode the vacuolar H+-ATPase. Trends Plant Sci. 2002, 7, 157–161. [Google Scholar] [CrossRef]
- Chen, Q.; Kan, Q.; Wang, P.; Yu, W.; Yu, Y.Z.; Zhao, Y.; Yu, Y.X.; Li, K.; Chen, L. Phosphorylation and interaction with the 14-3-3 protein of the plasma membrane H+-ATPase are involved in the regulation of magnesium-mediated increases in aluminum-induced citrate exudation in broad bean (Vicia faba. L). Plant Cell Physiol. 2015, 56, 1144–1153. [Google Scholar] [CrossRef]
- Duressa, D.; Soliman, K.M.; Taylor, R.W.; Chen, D. Gene expression profiling in soybean under aluminum stress: genes differentially expressed between Al-tolerant and Al-sensitive genotypes. Am. J. Mol. Biol. 2011, 1, 156–173. [Google Scholar] [CrossRef] [Green Version]
- Zhou, S.; Sauvé, R.; Thannhauser, T.W. Proteome changes induced by aluminium stress in tomato roots. J. Exp. Bot. 2009, 60, 1849–1857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marín, M.; Ott, T. Phosphorylation of intrinsically disordered regions in remorin proteins. Front. Plant Sci. 2012, 3, 86. [Google Scholar] [CrossRef] [PubMed]
- Diédhiou, C.J.; Popova, O.V.; Dietz, K.J.; Golldack, D. The SNF1-type serine-threonine protein kinase SAPK4 regulates stress-responsive gene expression in rice. BMC Plant Biol. 2008, 8, 49. [Google Scholar] [CrossRef] [PubMed]
- Carbrey, J.M.; Agre, P. Discovery of the aquaporins and development of the field. Handb Exp. Pharmacol. 2009, 190, 3. [Google Scholar]
- Quigley, F.; Rosenberg, J.M.; Shachar-Hill, Y.; Bohnert, H.J. From genome to function: the Arabidopsis aquaporins. Genome Biol. 2002, 3, research0001.1–research0001.17. [Google Scholar] [PubMed]
- Kumar, K.; Mosa, K.A.; Chhikara, S.; Musante, C.; White, J.C.; Dhankher, O.P. Two rice plasma membrane intrinsic proteins, OsPIP2;4 and OsPIP2;7, are involved in transport and providing tolerance to boron toxicity. Planta 2014, 239, 187–198. [Google Scholar] [CrossRef] [PubMed]
- Fitzpatrick, K.L.; Reid, R.J. The involvement of aquaglyceroporins in transport of boron in barley roots. Plant Cell Environ. 2009, 32, 1357–1365. [Google Scholar] [CrossRef] [PubMed]
- Negishi, T.; Oshima, K.; Hattori, M.; Kanai, M.; Mano, S.; Nishimura, M.; Yoshida, K. Tonoplast- and plasma membrane-localized aquaporin-family transporters in blue hydrangea sepals of aluminum hyperaccumulating plant. PLoS ONE 2012, 7, e43189. [Google Scholar] [CrossRef]
- Negishi, T.; Oshima, K.; Hattori, M.; Yoshida, K. Plasma membrane-localized Al-transporter from blue hydrangea sepals is a member of the anion permease family. Genes Cells 2013, 18, 341–352. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.T.; Qi, Y.P.; Lu, Y.B.; Guo, P.; Sang, W.; Feng, H.; Zhang, H.X.; Chen, L.S. iTRAQ protein profile analysis of Citrus sinensis roots in response to long-term boron-deficiency. J. Proteimics 2013, 93, 179–206. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.T.; Lu, Y.B.; Zhang, Y.; Guo, P.; Chen, L.S. Proteomic profile of Citrus grandis roots under long-term boron-deficiency revealed by iTRAQ. Trees - Struct. Funct. 2016, 30, 1057–1071. [Google Scholar] [CrossRef]
- Kosová, K.; Vítámvás, P.; Urban, M.O.; Prášil, I.T.; Renaut, J. Plant abiotic stress proteomics: the major factors determining alterations in cellular proteome. Front. Plant Sci. 2018, 9, 122. [Google Scholar] [CrossRef] [PubMed]
- Morant, M.; Bak, S.; Møller, B.L.; Werck-Reichhart, D. Plant cytochromes P450: tools for pharmacology, plant protection and phytoremediation. Curr. Opin. Biotech. 2003, 14, 151–162. [Google Scholar] [CrossRef]
- Schuler, M.A.; Werck-Reichhart, D. Functional genomics of P450s. Annu. Rev. Plant Biol. 2003, 54, 629–667. [Google Scholar] [CrossRef] [PubMed]
- Ruegger, M.; Meyer, K.; Chapple, C.C. Regulation of ferulate-5-hydroxylase expression in Arabidopsis in the context of sinapate ester biosynthesis. Plant Physiol. 1999, 119, 101–110. [Google Scholar] [CrossRef]
- Franke, R. Modified lignin in tobacco and poplar plants overexpressing the Arabidopsis gene encoding ferulate 5-hydroxylase. Plant J. 2000, 22, 223–234. [Google Scholar] [CrossRef]
- Bolwell, G.P.; Bozak, K.; Zimmerlin, A. Plant cytochrome P450. Phytochemistry 1994, 37, 1491–1506. [Google Scholar] [CrossRef]
- Li, L.; Liu, C.; Lian, X. Gene expression profiles in rice roots under low phosphorus stress. Plant Mol. Biol. 2010, 72, 423–432. [Google Scholar] [CrossRef]
- Jin, X.L.; Ma, C.L.; Yang, L.T.; Chen, L.S. Alterations of physiology and gene expression due to long-term magnesium-deficiency differ between leaves and roots of Citrus reticulata. J. Plant Physiol. 2016, 198, 103–115. [Google Scholar] [CrossRef]
- Yang, L.T.; Zhou, Y.F.; Wang, Y.Y.; Wu, Y.M.; Ye, X.; Guo, J.X.; Chen, L.S. Magnesium deficiency induced global transcriptome change in Citrus sinensis leaves revealed by RNA-Seq. Int. J. Mol. Sci. 2019, 20, 3129. [Google Scholar] [CrossRef] [PubMed]
- Scheible, W.R.; Morcuende, R.; Czechowski, T.; Fritz, C.; Osuna, D.; Palacios-Rojas, N.; Schindelasch, D.; Thimm, O.; Udvardi, M.K.; Stitt, M. Genome-wide reprogramming of primary and secondary metabolism, protein synthesis, cellular growth processes, and the regulatory infrastructure of Arabidopsis in response to nitrogen. Plant Physiol. 2004, 136, 2483–2499. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.B.; Qi, Y.P.; Lee, J.; Guo, P.; Ye, X.; Jia, M.Y.; Li, M.L.; Chen, L.S. Long-term boron-deficiency-responsive genes revealed by cDNA-AFLP differ between Citrus sinensis roots and leaves. Front. Plant Sci. 2015, 6, 271. [Google Scholar] [CrossRef] [PubMed]
- Hall, J.F.; Kanbi, L.D.; Harvey, I.; Murphy, L.M.; Hasnain, S.S. Modulating the redox potential and acid stability of rusticyanin by site-directed mutagenesis of Ser86. Biochemistry-US 1998, 37, 11451–11458. [Google Scholar] [CrossRef] [PubMed]
- Zeng, J.; Geng, M.; Liu, Y.; Xia, L.; Liu, J.; Qiu, G. The sulfhydryl group of Cys138 of rusticyanin from Acidithiobacillus ferrooxidans is crucial for copper binding. BBA-Proteins Proteom. 2007, 1774, 519–525. [Google Scholar] [CrossRef] [PubMed]
- Olson, G.J.; Brierley, J.A.; Brierley, C.L. Bioleaching review part B: progress in bioleaching: applications of microbial processes by the minerals industries. Appl. Microbiol. Biotechnol. 2003, 63, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Ezaki, B.; Sasaki, K.; Matsumoto, H.; Nakashima, S. Functions of two genes in aluminium (Al) stress resistance: repression of oxidative damage by the AtBCB gene and promotion of efflux of Al ions by the NtGDI1 gene. J. Exp. Bot. 2005, 56, 2661–2671. [Google Scholar] [CrossRef] [PubMed]
- Hsu, P.H. Effect of initial pH, phosphate, and silicate on the determination of aluminum with aluminon. Soil Sci. 1963, 96, 230–238. [Google Scholar] [CrossRef]
- Morrison, I.M. A semi-micro method for the determination of lignin and its use in predicting the digestibility of forage crops. J. Sci. Food Agric. 1972, 23, 455–463. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Y2H Code | Gene ID Number | Gene Annotation |
|---|---|---|
| C51 | Cs5g32800.1 | Glutathione S-transferase, GST |
| C55 | Cs3g25710.1 | Vacuolar-type proton ATPase, V-ATPase |
| C120 | Cs6g03670.1 | Aluminum Sensitive 3, ALS3 |
| C125 | Cs1g17390.1 | Cytochrome P450 71A1, CP450 |
| C131 | Cs8g16640.1 | Aquaporin PIP2, PIP2 |
| A58 | orange1.1t03320 | Uncharacterized protein |
| A62 | Cs6g13410.1 | Fasciclin-like arabinogalactan protein 17, FAP17 |
| A69 | Cs4g04340.1 | Auxin efflux carrier component 2, AECC2 |
| A97 | orange1.1t04179.1 | Ubiquitin carboxyl-terminal hydrolase 13, UCT13 |
| A102 | Cs8g03780.1 | Uncharacterized protein1, UP1 |
| A492 | Cs4g18120.1 | Putative dicyanin blue copper protein, DCBC |
| A615 | Cs7g02860.1 | Uncharacterized protein2, UP2 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Y.-M.; Wang, Y.-Y.; Zhou, Y.-F.; Meng, X.; Huang, Z.-R.; Chen, L.-S.; Yang, L.-T. Analysis of Interacting Proteins of Aluminum Toxicity Response Factor ALS3 and CAD in Citrus. Int. J. Mol. Sci. 2019, 20, 4846. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20194846
Wu Y-M, Wang Y-Y, Zhou Y-F, Meng X, Huang Z-R, Chen L-S, Yang L-T. Analysis of Interacting Proteins of Aluminum Toxicity Response Factor ALS3 and CAD in Citrus. International Journal of Molecular Sciences. 2019; 20(19):4846. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20194846
Chicago/Turabian StyleWu, Yan-Mei, Yan-Yu Wang, Yang-Fei Zhou, Xin Meng, Zeng-Rong Huang, Li-Song Chen, and Lin-Tong Yang. 2019. "Analysis of Interacting Proteins of Aluminum Toxicity Response Factor ALS3 and CAD in Citrus" International Journal of Molecular Sciences 20, no. 19: 4846. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20194846