Lectin Sequence Distribution in QTLs from Rice (Oryza sativa) Suggest a Role in Morphological Traits and Stress Responses



Abstract
:
1. Introduction
2. Results
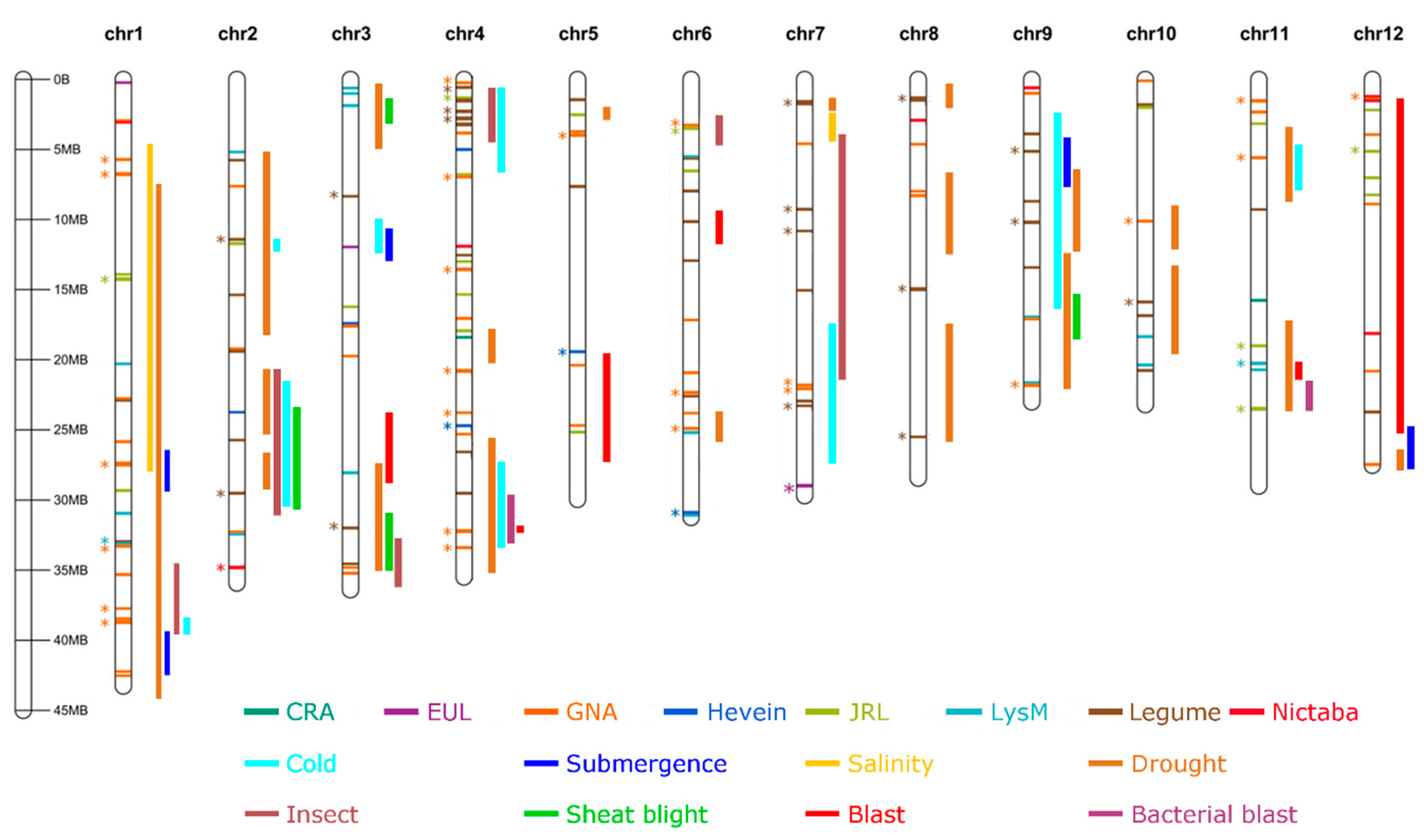
2.1. Lectin and QTL Distribution in Rice Genomes
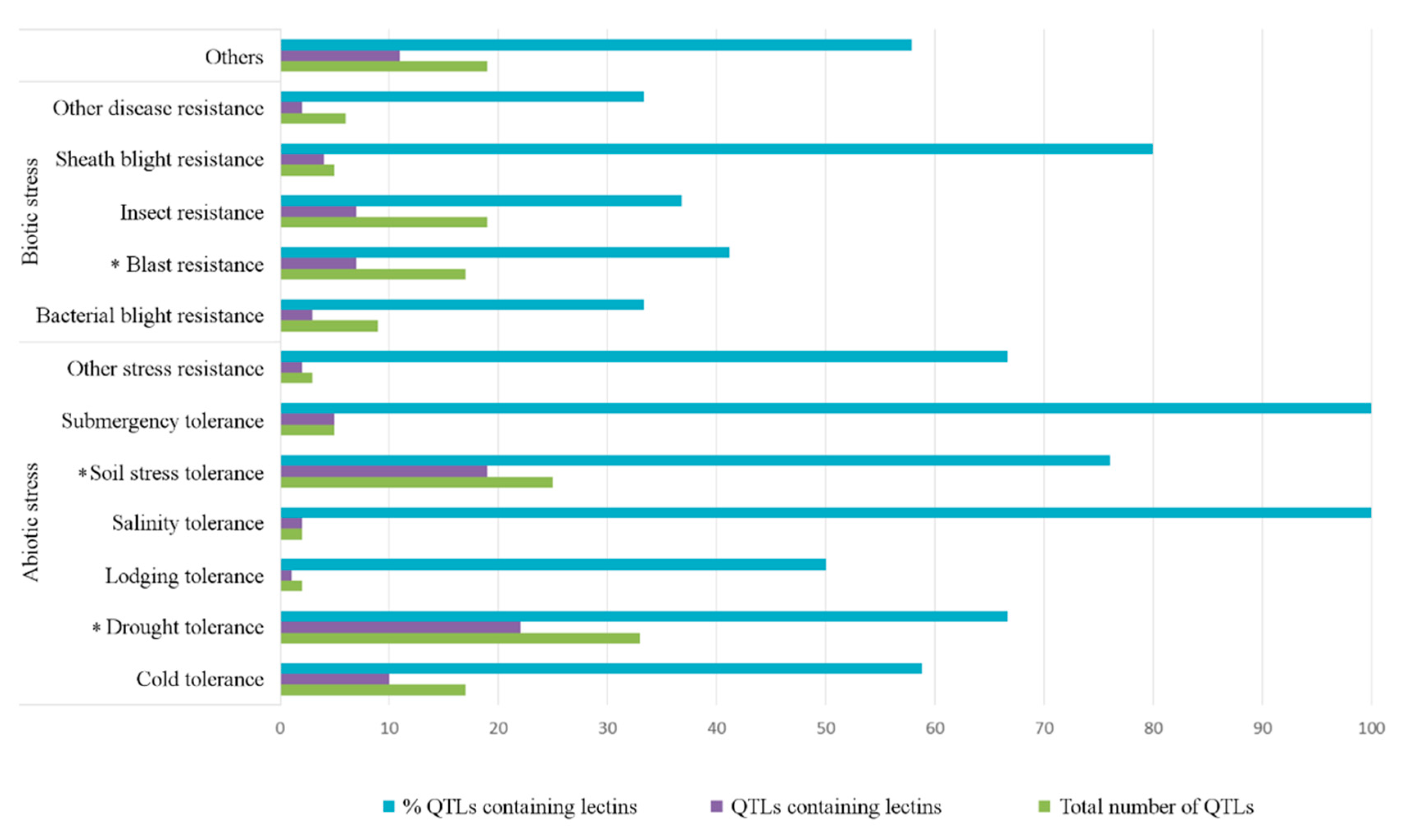
2.2. Lectin Sequences Are Widely Distributed in QTLs for Resistance or Tolerance
2.2.1. Abiotic Stress
2.2.2. Biotic Stress
2.3. Lectins Are Widely Distributed in QTLs for Morphological and Physiological Traits
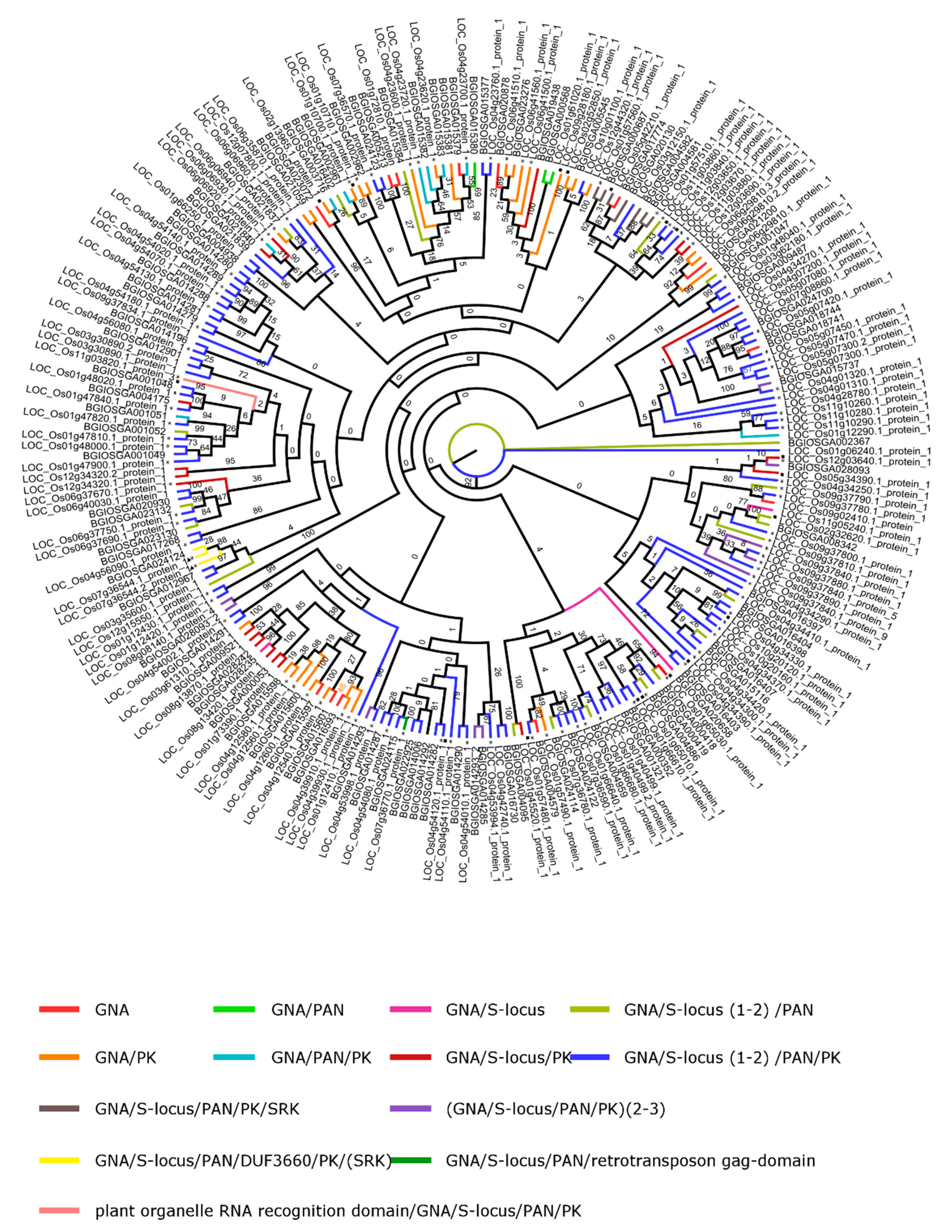
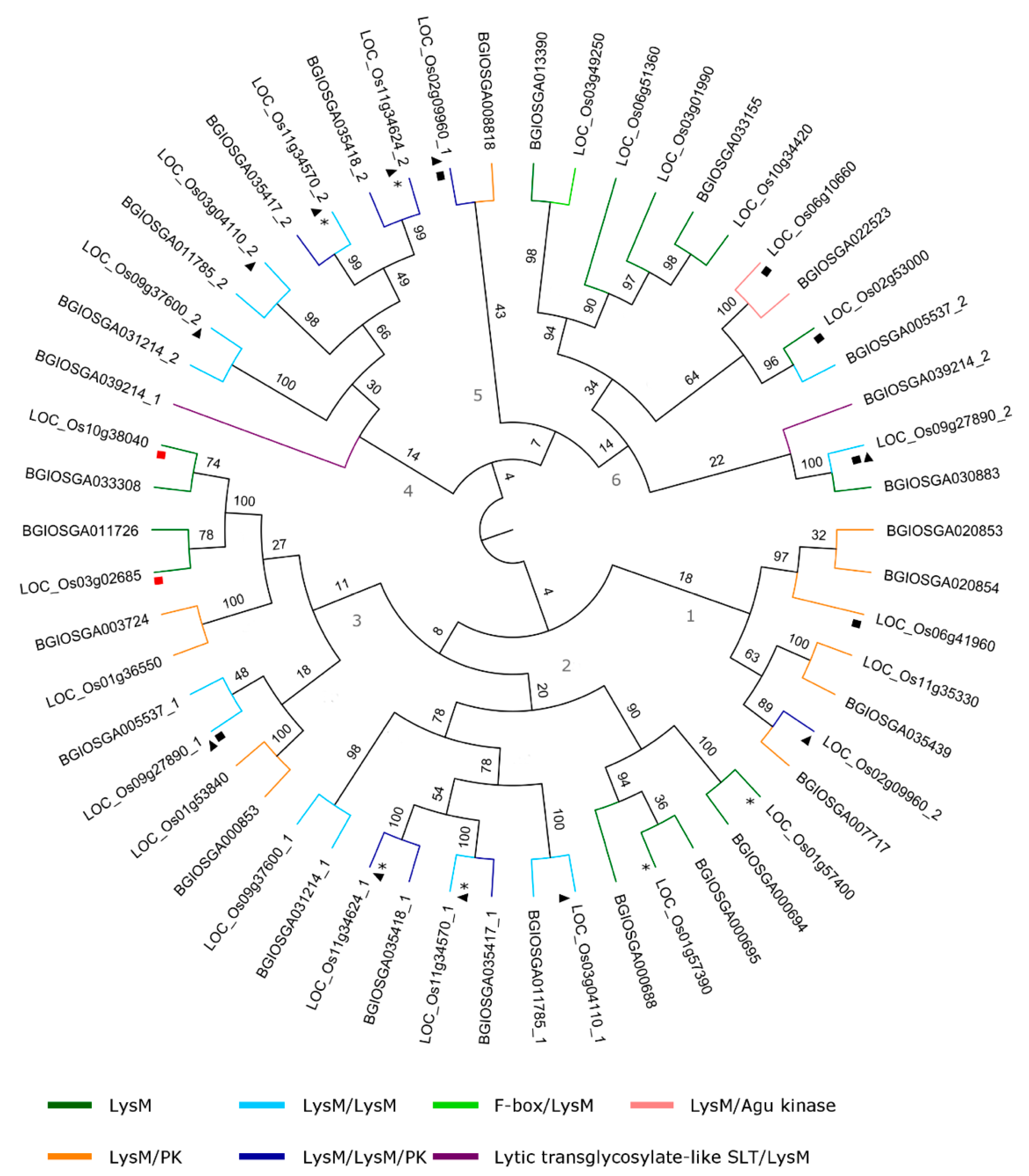
2.4. Lectin Domain Structure and Phylogenetic Analysis in O. sativa spp.
2.4.1. GNA
2.4.2. Jacalin-Related Lectins
2.4.3. LysM Domain
3. Discussion
4. Materials and Methods
4.1. Identification of Putative Lectin Genes in the Genome of O. sativa spp. Indica and Japonica
4.2. Analysis of Segmental and Tandem Duplications in O. sativa spp. Japonica
4.3. Mapping of the Lectin Genes on the Rice Chromosomes and in QTL Regions
4.4. Statistical Analysis
4.5. Phylogenetic Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Liu, F.; Xu, W.; Wei, Q.; Zhang, Z.; Xing, Z.; Tan, L.; Di, C.; Yao, D.; Wang, C.; Tan, Y.; et al. Gene expression profiles deciphering rice phenotypic variation between Nipponbare (Japonica) and 93-11 (Indica) during oxidative stress. PLoS ONE 2010, 5, e8632. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Zhu, K.; Xia, H.; Chen, L.; Chen, K. Comparative proteomic analysis of indica and japonica rice varieties. Genet. Mol. Biol. 2014, 37, 652–661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dixit, S.; Huang, B.E.; Cruz, M.T.S.; Maturan, P.T.; Ontoy, J.C.E.; Kumar, A. QTLs for tolerance of drought and breeding for tolerance of abiotic and biotic stress: An integrated approach. PLoS ONE 2014, 9, e109574. [Google Scholar] [CrossRef] [PubMed]
- Muthayya, S.; Sugimoto, J.D.; Montgomery, S.; Maberly, G.F. An overview of global rice production, supply, trade, and consumption. Ann. N. Y. Acad. Sci. 2014, 1324, 7–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, G.; Rao, G.J.N. Molecular marker assisted gene stacking for biotic and abiotic stress resistance genes in an elite rice cultivar. Front. Plant Sci. 2015, 6, 698. [Google Scholar] [CrossRef]
- Fahad, S.; Bajwa, A.A.; Nazir, U.; Anjum, S.A.; Farooq, A.; Zohaib, A.; Sadia, S.; Nasim, W.; Adkins, S.; Saud, S.; et al. Crop production under drought and heat stress: Plant responses and management options. Front. Plant Sci. 2017, 8, 1147. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.H.; Zheng, X.M.; Xu, Q.; Yuan, X.P.; Huang, L.; Zhou, H.F.; Wei, X.H.; Ge, S. Genetic diversity and classification of Oryza sativa with emphasis on Chinese rice germplasm. Heredity 2014, 112, 489–496. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.; Shi, J.; Quan, S.; Cui, B.; Kleessen, S.; Nikoloski, Z.; Tohge, T.; Alexander, D.; Guo, L.; Lin, H.; et al. Metabolic variation between japonica and indica rice cultivars as revealed by non-targeted metabolomics. Sci. Rep. 2014, 4, 5067. [Google Scholar] [CrossRef] [PubMed]
- Miura, K.; Ashikari, M.; Matsuoka, M. The role of QTLs in the breeding of high-yielding rice. Trends Plant Sci. 2011, 16, 319–326. [Google Scholar] [CrossRef] [PubMed]
- Tello-Ruiz, M.K.; Naithani, S.; Stein, J.C.; Gupta, P.; Campbell, M.; Olson, A.; Wei, S.; Preece, J.; Geniza, M.J.; Jiao, Y.; et al. Gramene 2018: Unifying comparative genomics and pathway resources for plant research. Nucleic Acids Res. 2017, 46, D1181–D1189. [Google Scholar] [CrossRef]
- Yonemaru, J.I.; Yamamoto, T.; Fukuoka, S.; Uga, Y.; Hori, K.; Yano, M. Q-TARO: QTL annotation rice online database. Rice 2010, 3, 194–203. [Google Scholar] [CrossRef]
- Fukao, T.; Xiong, L. Genetic mechanisms conferring adaptation to submergence and drought in rice: Simple or complex? Curr. Opin. Plant Biol. 2013, 16, 196–204. [Google Scholar] [CrossRef] [PubMed]
- Negrão, S.; Almadanim, M.C.; Pires, I.S.; Abreu, I.A.; Maroco, J.; Courtois, B.; Gregorio, G.B.; McNally, K.L.; Oliveira, M.M. New allelic variants found in key rice salt-tolerance genes: An association study. J. Plant Biotechnol. 2013, 11, 87–100. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Shang, J.; Chen, D.; Lei, C.; Zou, Y.; Zhai, W.; Liu, G.; Xu, J.; Ling, Z.; Cao, G.; et al. AB-lectin receptor kinase gene conferring rice blast resistance. Plant J. 2006, 46, 794–804. [Google Scholar] [CrossRef] [PubMed]
- Dimkpa, S.O.; Lahari, Z.; Shrestha, R.; Douglas, A.; Gheysen, G.; Price, A.H. A genome-wide association study of a global rice panel reveals resistance in Oryza sativa to root-knot nematodes. J. Exp. Bot. 2015, 67, 1191–1200. [Google Scholar] [CrossRef] [PubMed]
- Van Damme, E.J.M.; Lannoo, N.; Peumans, W.J. Plant lectins. Adv. Bot. Res. 2008, 48, 107–209. [Google Scholar]
- Van Holle, S.; Van Damme, E.J.M. Signaling through plant lectins: Modulation of plant immunity and beyond. Biochem. Soc. Trans. 2018, 46, 217–233. [Google Scholar] [CrossRef]
- Lannoo, N.; Van Damme, E.J.M. Lectin domains at the frontiers of plant defense. Front. Plant Sci. 2014, 5, 397. [Google Scholar] [CrossRef]
- De Schutter, K.; Van Damme, E.J.M. Protein-carbohydrate interactions as part of plant defense and animal immunity. Molecules 2015, 20, 9029–9053. [Google Scholar] [CrossRef]
- Van Holle, S.; De Schutter, K.; Eggermont, L.; Tsaneva, M.; Dang, L.; Van Damme, E.J.M. Comparative study of lectin domains in model species: New insights into evolutionary dynamics. Int. J. Mol. Sci. 2017, 18, 1136. [Google Scholar] [CrossRef]
- Eggermont, L.; Verstraeten, B.; Van Damme, E.J.M. Genome-wide screening for lectin motifs in Arabidopsis thaliana. Plant Genome 2017, 10. [Google Scholar] [CrossRef]
- Saeed, B.; Baranwal, V.K.; Khurana, P. Identification and expression profiling of the lectin gene superfamily in mulberry. Plant Genome 2016, 9. [Google Scholar] [CrossRef] [PubMed]
- Van Holle, S.; Van Damme, E.J.M. Distribution and evolution of the lectin family in soybean (Glycine max). Molecules 2015, 20, 2868–2891. [Google Scholar] [CrossRef]
- Dang, L.; Van Damme, E.J.M. Genome-wide identification and domain organization of lectin domains in cucumber. Plant Physiol. Biochem. 2016, 108, 165–176. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.Y.; Ma, Z.; Ramachandran, S. Evolutionary history and stress regulation of the lectin superfamily in higher plants. BMC Evol. Biol. 2010, 10, 79. [Google Scholar] [CrossRef]
- De Schutter, K.; Tsaneva, M.; Kulkarni, S.R.; Rougé, P.; Vandepoele, K.; Van Damme, E.J.M. Evolutionary relationships and expression analysis of EUL domain proteins in rice (Oryza sativa). Rice 2017, 10, 26. [Google Scholar] [CrossRef]
- Han, Y.J.; Zhong, Z.H.; Song, L.L.; Stefan, O.; Wang, Z.H.; Lu, G.D. Evolutionary analysis of plant Jacalin-related lectins (JRLs) family and expression of rice JRLs in response to Magnaporthe oryzae. J. Integr. Agric. 2018, 17, 1252–1266. [Google Scholar] [CrossRef]
- Esch, L.; Schaffrath, U. An update on Jacalin-like lectins and their role in plant defense. Int. J. Mol. Sci. 2017, 18, 1592. [Google Scholar] [CrossRef]
- Al Atalah, B.; Smagghe, G.; Van Damme, E.J.M. Orysata, a Jacalin-related lectin from rice, could protect plants against biting-chewing and piercing-sucking insects. Plant Sci. 2014, 221, 21–28. [Google Scholar] [CrossRef]
- Zou, X.; Qin, Z.; Zhang, C.; Liu, B.; Liu, J.; Zhang, C.; Lin, C.; Li, H.; Zhao, T. Over-expression of an S-domain receptor-like kinase extracellular domain improves panicle architecture and grain yield in rice. J. Exp. Bot. 2015, 66, 7197–7209. [Google Scholar] [CrossRef] [Green Version]
- Cheng, X.; Wu, Y.; Guo, J.; Du, B.; Chen, R.; Zhu, L.; He, G. A Rice lectin receptor-like kinase that is involved in innate immune responses also contributes to seed germination. Plant J. 2013, 76, 687–698. [Google Scholar] [CrossRef] [PubMed]
- Li, C.H.; Wang, G.; Zhao, J.L.; Zhang, L.Q.; Ai, L.F.; Han, Y.F.; Sun, D.Y.; Zhang, S.W.; Sun, Y. The receptor-like kinase SIT1 mediates salt sensitivity by activating MAPK3/6 and regulating ethylene homeostasis in rice. Plant Cell 2014, 26, 2538–2553. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, E.; Yonemaru, J.I.; Yamamoto, T.; Yano, M. OGRO: The Overview of functionally characterized Genes in rice online database. Rice 2012, 5, 26. [Google Scholar] [CrossRef] [PubMed]
- Vij, S.; Giri, J.; Dansana, P.K.; Kapoor, S.; Tyagi, A.K. The receptor-like cytoplasmic kinase (OsRLCK) gene family in rice: Organization, phylogenetic relationship, and expression during development and stress. Mol. Plant 2008, 1, 732–750. [Google Scholar] [CrossRef] [PubMed]
- Todaka, D.; Nakashima, K.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Toward understanding transcriptional regulatory networks in abiotic stress responses and tolerance in rice. Rice 2012, 5, 6. [Google Scholar] [CrossRef] [Green Version]
- Salih, H.; Adelson, D.L. QTL global meta-analysis: Are trait determining genes clustered? BMC Gen. 2009, 10, 184. [Google Scholar] [CrossRef]
- Oono, Y.; Yazawa, T.; Kawahara, Y.; Kanamori, H.; Kobayashi, F.; Sasaki, H.; Mori, S.; Wu, J.; Handa, H.; Itoh, T.; et al. Genome-wide transcriptome analysis reveals that cadmium stress signaling controls the expression of genes in drought stress signal pathways in rice. PLoS ONE 2014, 9, e96946. [Google Scholar] [CrossRef]
- Shimizu, T.; Nakano, T.; Takamizawa, D.; Desaki, Y.; Ishii-Minami, N.; Nishizawa, Y.; Minami, E.; Okada, K.; Yamane, H.; Kaku, H.; et al. Two LysM receptor molecules, CEBiP and OsCERK1, cooperatively regulate chitin elicitor signaling in rice. Plant J. 2010, 64, 204–214. [Google Scholar] [CrossRef] [Green Version]
- Desaki, Y.; Kouzai, Y.; Ninomiya, Y.; Iwase, R.; Shimizu, Y.; Seko, K.; Molinaro, A.; Minami, E.; Shibuya, N.; Kaku, H.; et al. OsCERK1 plays a crucial role in the lipopolysaccharide-induced immune response of rice. New Phytol. 2018, 217, 1042–1049. [Google Scholar] [CrossRef]
- Carotenuto, G.; Chabaud, M.; Miyata, K.; Capozzi, M.; Takeda, N.; Kaku, H.; Shibuya, N.; Nakagawa, T.; Barker, D.G.; Genre, A. The Rice LysM receptor-like kinase OsCERK 1 is required for the perception of short-chain chitin oligomers in arbuscular mycorrhizal signaling. New Phytol. 2017, 214, 1440–1446. [Google Scholar] [CrossRef]
- Liu, B.; Li, J.F.; Ao, Y.; Qu, J.; Li, Z.; Su, J.; Zhang, Y.; Liu, J.; Feng, D.; Qi, K.; et al. Lysin motif–containing proteins LYP4 and LYP6 play dual roles in peptidoglycan and chitin perception in rice innate immunity. Plant Cell 2012, 24, 3406–3419. [Google Scholar] [CrossRef] [PubMed]
- Kittur, F.S.; Yu, H.Y.; Bevan, D.R.; Esen, A. Deletion of the N-terminal dirigent domain in maize β-glucosidase aggregating factor and its homolog sorghum lectin dramatically alters the sugar-specificities of their lectin domains. Plant Physiol. Biochem. 2010, 48, 731–734. [Google Scholar] [CrossRef]
- Fujita, M.; Fujita, Y.; Noutoshi, Y.; Takahashi, F.; Narusaka, Y.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Crosstalk between abiotic and biotic stress responses: A current view from the points of convergence in the stress signaling networks. Curr. Opin. Plant Biol. 2006, 9, 436–442. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.; Zhang, X.L.; Nadir, S.; DongChen, W.H.; Guo, X.Q.; Zhang, H.X.; Li, C.Y.; Chen, L.J.; Lee, D.S. A LysM Domain-Containing Gene OsEMSA1 Involved in Embryo sac Development in Rice (Oryza sativa L.). Front. Plant Sci. 2017, 8, 1596. [Google Scholar] [CrossRef]
- Jiang, J.F.; Xu, Y.Y.; Chong, K. Overexpression of OsJAC1, a lectin gene, suppresses the coleoptile and stem elongation in rice. J. Integr. Plant Biol. 2007, 49, 230–237. [Google Scholar] [CrossRef]
- Xiao, J.; Li, C.; Xu, S.; Xing, L.; Xu, Y.; Chong, K. JACALIN-LECTIN LIKE1 regulates the nuclear accumulation of GLYCINE-RICH RNA-BINDING PROTEIN7, influencing the RNA processing of FLOWERING LOCUS C antisense transcripts and flowering time in Arabidopsis. Plant Physiol. 2015, 169, 2102–2117. [Google Scholar]
- Lee, T.H.; Tang, H.; Wang, X.; Paterson, A.H. PGDD: A database of gene and genome duplication in plants. Nucleic Acids Res. 2012, 41, D1152–D1158. [Google Scholar] [CrossRef]
- Voorrips, R.E. MapChart: Software for the graphical presentation of linkage maps and QTLs. J. Hered. 2002, 93, 77–78. [Google Scholar] [CrossRef]
- Quinlan, A.R.; Hall, I.M. BEDTools: a flexible suite of utilities for comparing genomic features. Bioinformatics 2010, 26, 841–842. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- Capella-Gutiérrez, S.; Silla-Martínez, J.M.; Gabaldón, T. trimAl: A tool for automated alignment trimming in large-scale phylogenetic analyses. Bioinformatics 2009, 25, 1972–1973. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lectin and Domain Architecture | Schematic Representation | Spp. Japonica | Spp. Indica |
|---|---|---|---|
| CRA |  | 2 | 3 |
| EUL |  | 2 | 2 |
| (EUL)2 |  | 3 | 3 |
| Hevein |  | 1 | 1 |
| (Hevein)4 |  | 1 | 1 |
| Hevein/GH19 |  | 8 | 8 |
| Nictaba |  | 4 | 5 |
| F-box/Nictaba |  | 13 | 14 |
| Fbox/(Nictaba)2 |  | 1 | 2 |
| Protein kinase/Nictaba |  | 1 | 1 |
| Jacalin |  | 16 | 21 |
| (Jacalin)2 |  | 1 | 1 |
| (Jacalin)3 |  | 1 | 1 |
| Protein kinase/Jacalin |  | 1 | 0 |
| Protein kinase/(Jacalin)2 |  | 1 | 0 |
| Protein kinase/(Jacalin)3 |  | 2 | 1 |
| Dirigent/Jacalin |  | 4 | 4 |
| NB-ARC/Jacalin |  | 2 | 2 |
| NB-ARC/(Jacalin)3 |  | 2 | 2 |
| LysM |  | 8 | 8 |
| (LysM)2 |  | 4 | 3 |
| LysM/Protein kinase |  | 4 | 7 |
| (LysM)2/Protein kinase |  | 1 | 1 |
| Aspartate/glutamate/uridylate kinase/LysM |  | 2 | 2 |
| Legume lectin |  | 14 | 12 |
| (Legume lectin)2 |  | 1 | 0 |
| Legume lectin/Protein kinase |  | 61 | 73 |
| (Legume lectin)2/Protein kinase |  | 1 | 2 |
| GNA |  | 2 | 11 |
| GNA/(S-locus glycoprotein)1–2/(PAN)/(protein kinase)1–2/(SRK) * |  | 119 | 82 ** |
| GNA/S-locus glycoprotein/PAN/DUF ***/protein kinase/(SRK) |  | 2 | 1 |
| Legend | |||
 | CRA (PF00704) |  | GH19 (PF00182) |
 | EUL (PF14200) |  | F-box (PF00646) |
 | Hevein (PF00187) |  | Protein kinase domains (PF00069, PF50011, PF07714) |
 | Nictaba (PF14299) |  | Dirigent (PF03018) |
 | Jacalins (PF01419) |  | NB-ARC (PF00931) |
 | LysM (PF01476) |  | Aspartate/glutamate/uridylate kinase (PF00696) |
 | Legume lectin (PF00139) |  | S-locus glycoprotein (PF00954) |
 | GNA (PF01453) |  | PAN domains (PF00024, PF08276, PF14295) |
 | SRK (PF11883) | ||
 | DUF domains (PF12398, PF11884) | ||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsaneva, M.; De Schutter, K.; Verstraeten, B.; Van Damme, E.J.M. Lectin Sequence Distribution in QTLs from Rice (Oryza sativa) Suggest a Role in Morphological Traits and Stress Responses. Int. J. Mol. Sci. 2019, 20, 437. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20020437
Tsaneva M, De Schutter K, Verstraeten B, Van Damme EJM. Lectin Sequence Distribution in QTLs from Rice (Oryza sativa) Suggest a Role in Morphological Traits and Stress Responses. International Journal of Molecular Sciences. 2019; 20(2):437. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20020437
Chicago/Turabian StyleTsaneva, Mariya, Kristof De Schutter, Bruno Verstraeten, and Els J.M. Van Damme. 2019. "Lectin Sequence Distribution in QTLs from Rice (Oryza sativa) Suggest a Role in Morphological Traits and Stress Responses" International Journal of Molecular Sciences 20, no. 2: 437. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20020437