Caenorhabditis Elegans and Probiotics Interactions from a Prolongevity Perspective

 , and
, and
Abstract
:1. Caenorhabditis Elegans as a Model System to Study Prolongevity
2. Probiotics: Characteristics and Relevance to Ageing
3. Review Methodology
4. Mechanisms Involved in C. elegans Lifespan Extension Induced by Probiotics
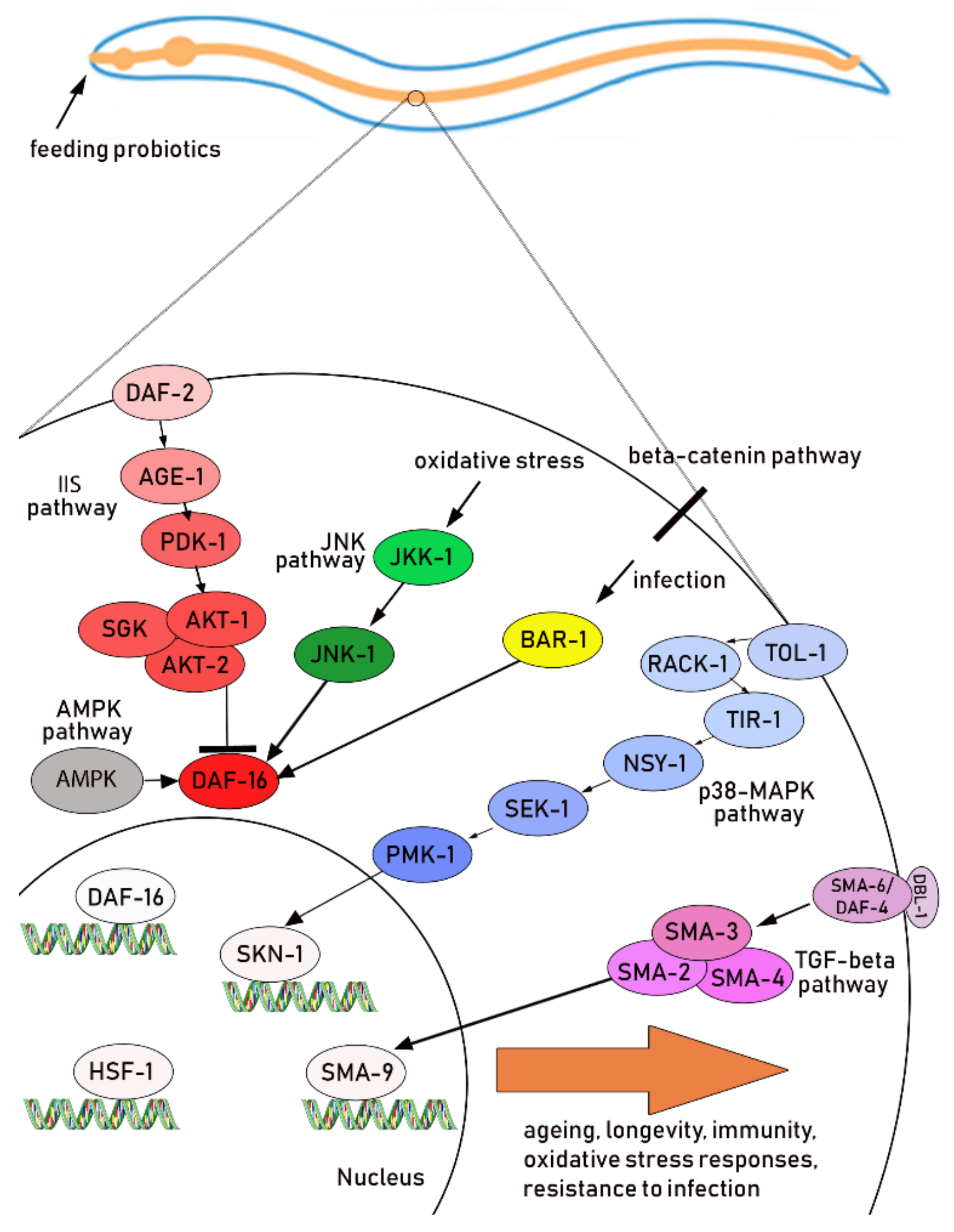
4.1. Description of the Main Pathways
4.2. Oxidative Stress Response
4.3. Immune Response and Pathogen Protection
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AMPK | 5′-AMP-activated protein kinase |
| CLEC | C-type lectins |
| DAF | Dauer Formation |
| ETEC | Enterotoxigenic E. coli |
| FOXO | Forkhead box O |
| GST | Glutathione S-transferase |
| HSF-1 | Heat-Shock transcription Factor-1 |
| IIS | Insulin/insulin-like growth factor-1 |
| JNK | c-Jun N-terminal kinase |
| LYS | Lysozyme |
| MAMP | Microbial associated molecular pattern |
| p38 MAPK | p38 mitogen-activated protein kinase |
| PMK-1 | Mitogen-activated protein kinase-1 |
| ROS | Reactive oxygen species; |
| SKN-1 | Skinhead family member-1 |
| SOD | Superoxide dismutase |
| TGF-beta | Transforming growth factor-beta |
| TIR-1 | Toll interleukin-1 receptor-1 |
| TLR | Toll-like receptor |
References
- Brenner, S. The genetics of Caenorhabditis elegans. Genetics 1974, 77, 71–94. [Google Scholar] [PubMed]
- Kenyon, C. A conserved regulatory system for aging. Cell 2001, 105, 165–168. [Google Scholar] [CrossRef]
- O’Kane, C.J. Modelling human diseases in Drosophila and Caenorhabditis. Semin. Cell Dev. Biol. 2003, 14, 3–10. [Google Scholar] [CrossRef]
- Fontana, L.; Partridge, L.; Longo, V.D. Extending healthy life span—From yeast to humans. Science 2010, 328, 321–326. [Google Scholar] [CrossRef] [PubMed]
- Murphy, C.T.; McCarroll, S.A.; Bargmann, C.I.; Fraser, A.; Kamath, R.S.; Ahringer, J.; Li, H.; Kenyon, C. Genes that act downstream of DAF-16 to influence the lifespan of Caenorhabditis elegans. Nature 2003, 424, 277–283. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.H.; Feinbaum, R.; Alloing, G.; Emerson, F.E.; Garsin, D.A.; Inoue, H.; Tanaka-Hino, M.; Hisamoto, N.; Matsumoto, K.; Tan, M.W.; et al. A conserved p38 MAP kinase pathway in Caenorhabditis elegans innate immunity. Science 2002, 297, 623–626. [Google Scholar] [CrossRef] [PubMed]
- So, S.; Tokumaru, T.; Miyahara, K.; Ohshima, Y. Control of lifespan by food bacteria, nutrient limitation and pathogenicity of food in C. elegans. Mech. Ageing Dev. 2011, 132, 210–212. [Google Scholar] [CrossRef] [PubMed]
- Troemel, E.R.; Chu, S.W.; Reinke, V.; Lee, S.S.; Ausubel, F.M.; Kim, D.H. p38 MAPK regulates expression of immune response genes and contributes to longevity in C. elegans. PLoS Genet. 2006, 2, e183. [Google Scholar] [CrossRef]
- Nakagawa, H.; Shiozaki, T.; Kobatake, E.; Hosoya, T.; Moriya, T.; Sakai, F.; Taru, H.; Miyazaki, T. Effects and mechanisms of prolongevity induced by Lactobacillus gasseri SBT2055 in Caenorhabditis elegans. Aging Cell 2016, 15, 227–236. [Google Scholar] [CrossRef]
- Johnson, T.E.; Henderson, S.; Murakami, S.; de Castro, E.; de Castro, S.H.; Cypser, J.; Rikke, B.; Tedesco, P.; Link, C. Longevity genes in the nematode Caenorhabditis elegans also mediate increased resistance to stress and prevent disease. J. Inherit. Metab. Dis. 2002, 25, 197–206. [Google Scholar] [CrossRef]
- Pincus, Z.; Mazer, T.C.; Slack, F.J. Autofluorescence as a measure of senescence in C. elegans: Look to red, not blue or green. Aging 2016, 8, 889–898. [Google Scholar] [CrossRef] [PubMed]
- Son, H.G.; Altintas, O.; Kim, E.J.E.; Kwon, S.; Lee, S.V. Age-dependent changes and biomarkers of aging in Caenorhabditis elegans. Aging Cell 2019, 18, e12853. [Google Scholar] [CrossRef]
- Clark, L.C.; Hodgkin, J. Commensals, probiotics and pathogens in the Caenorhabditis elegans model. Cell Microbiol. 2014, 16, 27–38. [Google Scholar] [CrossRef] [PubMed]
- FAO/WHO. Probiotics in Food. Health and nutritional properties and guidelines for evaluation. Available online: http://www.fao.org/3/a-a0512e.pdf (accessed on 7 February 2019).
- O’Hara, A.M.; Shanahan, F. Mechanisms of action of probiotics in intestinal diseases. Sci. World J. 2007, 7, 31–46. [Google Scholar] [CrossRef] [PubMed]
- Bermudez-Brito, M.; Plaza-Diaz, J.; Munoz-Quezada, S.; Gomez-Llorente, C.; Gil, A. Probiotic mechanisms of action. Ann. Nutr. Metab. 2012, 61, 160–174. [Google Scholar] [CrossRef]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. Expert consensus document. The International scientific association for probiotics and prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef]
- Calder, P.C.; Bosco, N.; Bourdet-Sicard, R.; Capuron, L.; Delzenne, N.; Dore, J.; Franceschi, C.; Lehtinen, M.J.; Recker, T.; Salvioli, S.; et al. Health relevance of the modification of low grade inflammation in ageing (inflammageing) and the role of nutrition. Ageing Res. Rev. 2017, 40, 95–119. [Google Scholar] [CrossRef]
- Ferrucci, L.; Fabbri, E. Inflammageing: Chronic inflammation in ageing, cardiovascular disease, and frailty. Nat. Rev. Cardiol. 2018, 15, 505–522. [Google Scholar] [CrossRef]
- Biagi, E.; Candela, M.; Turroni, S.; Garagnani, P.; Franceschi, C.; Brigidi, P. Ageing and gut microbes: Perspectives for health maintenance and longevity. Pharm. Res. 2013, 69, 11–20. [Google Scholar] [CrossRef]
- Neish, A.S. Microbes in gastrointestinal health and disease. Gastroenterology 2009, 136, 65–80. [Google Scholar] [CrossRef]
- Hopkins, M.J.; Macfarlane, G.T. Changes in predominant bacterial populations in human faeces with age and with Clostridium difficile infection. J. Med. Microbiol. 2002, 51, 448–454. [Google Scholar] [CrossRef]
- Claesson, M.J.; Jeffery, I.B.; Conde, S.; Power, S.E.; O’Connor, E.M.; Cusack, S.; Harris, H.M.; Coakley, M.; Lakshminarayanan, B.; O’Sullivan, O.; et al. Gut microbiota composition correlates with diet and health in the elderly. Nature 2012, 488, 178–184. [Google Scholar] [CrossRef] [PubMed]
- Aureli, P.; Capurso, L.; Castellazzi, A.M.; Clerici, M.; Giovannini, M.; Morelli, L.; Poli, A.; Pregliasco, F.; Salvini, F.; Zuccotti, G.V. Probiotics and health: An evidence-based review. Pharm. Res. 2011, 63, 366–376. [Google Scholar] [CrossRef] [PubMed]
- Finamore, A.; Roselli, M.; Donini, L.; Brasili, D.E.; Rami, R.; Carnevali, P.; Mistura, L.; Pinto, A.; Giusti, A.; Mengheri, E. Supplementation with Bifidobacterium longum Bar33 and Lactobacillus helveticus Bar13 mixture improves immunity in elderly humans (over 75 years) and aged mice. Nutrition 2019, 63, 184–192. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Hou, A. Host-Microbe Interactions in Caenorhabditis elegans. ISRN Microbiol. 2013, 2013. [Google Scholar] [CrossRef] [PubMed]
- Ventura, M.T.; Casciaro, M.; Gangemi, S.; Buquicchio, R. Immunosenescence in aging: Between immune cells depletion and cytokines up-regulation. Clin. Mol. Allergy 2017, 15, 21. [Google Scholar] [CrossRef]
- Fulop, T.; Larbi, A.; Dupuis, G.; Le Page, A.; Frost, E.H.; Cohen, A.A.; Witkowski, J.M.; Franceschi, C. Immunosenescence and inflamm-aging as two sides of the same coin: Friends or foes? Front. Immunol. 2017, 8, 1960. [Google Scholar] [CrossRef]
- Kim, Y.; Mylonakis, E. Caenorhabditis elegans immune conditioning with the probiotic bacterium Lactobacillus acidophilus strain NCFM enhances Gram-positive immune responses. Infect. Immun. 2012, 80, 2500–2508. [Google Scholar] [CrossRef]
- Chelliah, R.; Choi, J.G.; Hwang, S.B.; Park, B.J.; Daliri, E.B.; Kim, S.H.; Wei, S.; Ramakrishnan, S.R.; Oh, D.H. In vitro and in vivo defensive effect of probiotic LAB against Pseudomonas aeruginosa using Caenorhabditis elegans model. Virulence 2018, 9, 1489–1507. [Google Scholar] [CrossRef]
- Wang, C.; Wang, J.; Gong, J.; Yu, H.; Pacan, J.C.; Niu, Z.; Si, W.; Sabour, P.M. Use of Caenorhabditis elegans for preselecting Lactobacillus isolates to control Salmonella Typhimurium. J. Food Prot. 2011, 74, 86–93. [Google Scholar] [CrossRef]
- Kamaladevi, A.; Balamurugan, K. Lactobacillus casei triggers a TLR mediated RACK-1 dependent p38 MAPK pathway in Caenorhabditis elegans to resist Klebsiella pneumoniae infection. Food Funct. 2016, 7, 3211–3223. [Google Scholar] [CrossRef] [PubMed]
- Kamaladevi, A.; Ganguli, A.; Balamurugan, K. Lactobacillus casei stimulates phase-II detoxification system and rescues malathion-induced physiological impairments in Caenorhabditis elegans. Comp. Biochem. Physiol. C Toxicol. Pharm. 2016, 179, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Guantario, B.; Zinno, P.; Schifano, E.; Roselli, M.; Perozzi, G.; Palleschi, C.; Uccelletti, D.; Devirgiliis, C. In vitro and in vivo selection of potentially probiotic lactobacilli from nocellara del belice table olives. Front. Microbiol. 2018, 9, 595. [Google Scholar] [CrossRef] [PubMed]
- Zanni, E.; Schifano, E.; Motta, S.; Sciubba, F.; Palleschi, C.; Mauri, P.; Perozzi, G.; Uccelletti, D.; Devirgiliis, C.; Miccheli, A. Combination of metabolomic and proteomic analysis revealed different features among Lactobacillus delbrueckii subspecies bulgaricus and lactis strains while in vivo testing in the model organism Caenorhabditis elegans highlighted probiotic properties. Front. Microbiol. 2017, 8, 1206. [Google Scholar] [CrossRef] [PubMed]
- Schifano, E.; Zinno, P.; Guantario, B.; Roselli, M.; Marcoccia, S.; Devirgiliis, C.; Uccelletti, D. The foodborne strain Lactobacillus fermentum MBC2 triggers pept-1-dependent pro-longevity effects in Caenorhabditis elegans. Microorganisms 2019, 7, 45. [Google Scholar] [CrossRef] [PubMed]
- Park, M.R.; Ryu, S.; Maburutse, B.E.; Oh, N.S.; Kim, S.H.; Oh, S.; Jeong, S.Y.; Jeong, D.Y.; Kim, Y. Probiotic Lactobacillus fermentum strain JDFM216 stimulates the longevity and immune response of Caenorhabditis elegans through a nuclear hormone receptor. Sci. Rep. 2018, 8, 7441. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Yun, H.S.; Cho, K.W.; Oh, S.; Kim, S.H.; Chun, T.; Kim, B.; Whang, K.Y. Evaluation of probiotic characteristics of newly isolated Lactobacillus spp.: Immune modulation and longevity. Int. J. Food Microbiol. 2011, 148, 80–86. [Google Scholar] [CrossRef]
- Lee, J.; Choe, J.; Kim, J.; Oh, S.; Park, S.; Kim, S.; Kim, Y. Heat-killed Lactobacillus spp. cells enhance survivals of Caenorhabditis elegans against Salmonella and Yersinia infections. Lett. Appl. Microbiol. 2015, 61, 523–530. [Google Scholar] [CrossRef]
- Ikeda, T.; Yasui, C.; Hoshino, K.; Arikawa, K.; Nishikawa, Y. Influence of lactic acid bacteria on longevity of Caenorhabditis elegans and host defense against Salmonella enterica serovar enteritidis. Appl. Environ. Microbiol. 2007, 73, 6404–6409. [Google Scholar] [CrossRef]
- Pan, F.; Zhang, L.; Li, M.; Hu, Y.; Zeng, B.; Yuan, H.; Zhao, L.; Zhang, C. Predominant gut Lactobacillus murinus strain mediates anti-inflammaging effects in calorie-restricted mice. Microbiome 2018, 6, 54. [Google Scholar] [CrossRef]
- De Barros, P.P.; Scorzoni, L.; Ribeiro, F.C.; Fugisaki, L.R.O.; Fuchs, B.B.; Mylonakis, E.; Jorge, A.O.C.; Junqueira, J.C.; Rossoni, R.D. Lactobacillus paracasei 28.4 reduces in vitro hyphae formation of Candida albicans and prevents the filamentation in an experimental model of Caenorhabditis elegans. Microb. Pathog. 2018, 117, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.K.; Choi, S.H.; Lee, C.R.; Lee, S.H.; Park, M.R.; Kim, Y.; Lee, M.K.; Kim, G.B. Screening and characterization of lactic acid bacteria strains with anti-inflammatory activities through in vitro and Caenorhabditis elegans model testing. Korean J. Food Sci. Anim. Resour. 2015, 35, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Park, M.R.; Yun, H.S.; Son, S.J.; Oh, S.; Kim, Y. Short communication: Development of a direct in vivo screening model to identify potential probiotic bacteria using Caenorhabditis elegans. J. Dairy Sci. 2014, 97, 6828–6834. [Google Scholar] [CrossRef] [PubMed]
- Sharma, K.; Pooranachithra, M.; Balamurugan, K.; Goel, G. Probiotic mediated colonization resistance against E. coli infection in experimentally challenged Caenorhabditis elegans. Microb. Pathog. 2019, 127, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Zhu, J.; Yu, H.; Yin, X.; Sabour, P.M.; Zhao, L.; Chen, W.; Gong, J. Investigation into in vitro and in vivo models using intestinal epithelial IPEC-J2 cells and Caenorhabditis elegans for selecting probiotic candidates to control porcine enterotoxigenic Escherichia coli. J. Appl. Microbiol. 2014, 117, 217–226. [Google Scholar] [CrossRef] [PubMed]
- Fasseas, M.K.; Fasseas, C.; Mountzouris, K.C.; Syntichaki, P. Effects of Lactobacillus salivarius, Lactobacillus reuteri, and Pediococcus acidilactici on the nematode Caenorhabditis elegans include possible antitumor activity. Appl. Microbiol. Biotechnol. 2013, 97, 2109–2118. [Google Scholar] [CrossRef]
- Azat, R.; Liu, Y.; Li, W.; Kayir, A.; Lin, D.B.; Zhou, W.W.; Zheng, X.D. Probiotic properties of lactic acid bacteria isolated from traditionally fermented Xinjiang cheese. J. Zhejiang Univ. Sci. B 2016, 17, 597–609. [Google Scholar] [CrossRef] [Green Version]
- Grompone, G.; Martorell, P.; Llopis, S.; Gonzalez, N.; Genoves, S.; Mulet, A.P.; Fernandez-Calero, T.; Tiscornia, I.; Bollati-Fogolin, M.; Chambaud, I.; et al. Anti-inflammatory Lactobacillus rhamnosus CNCM I-3690 strain protects against oxidative stress and increases lifespan in Caenorhabditis elegans. PLoS ONE 2012, 7, e52493. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhao, L.; Zheng, X.; Fu, T.; Guo, H.; Ren, F. Lactobacillus salivarius strain FDB89 induced longevity in Caenorhabditis elegans by dietary restriction. J. Microbiol. 2013, 51, 183–188. [Google Scholar] [CrossRef]
- Zhou, M.; Yu, H.; Yin, X.; Sabour, P.M.; Chen, W.; Gong, J. Lactobacillus zeae protects Caenorhabditis elegans from enterotoxigenic Escherichia coli-caused death by inhibiting enterotoxin gene expression of the pathogen. PLoS ONE 2014, 9, e89004. [Google Scholar] [CrossRef]
- Zhou, M.; Liu, X.; Yu, H.; Yin, X.; Nie, S.P.; Xie, M.Y.; Chen, W.; Gong, J. Cell signaling of Caenorhabditis elegans in response to enterotoxigenic Escherichia coli infection and Lactobacillus zeae protection. Front. Immunol. 2018, 9, 1745. [Google Scholar] [CrossRef] [PubMed]
- Martorell, P.; Llopis, S.; Gonzalez, N.; Chenoll, E.; Lopez-Carreras, N.; Aleixandre, A.; Chen, Y.; Karoly, E.D.; Ramon, D.; Genoves, S. Probiotic strain Bifidobacterium animalis subsp. lactis CECT 8145 reduces fat content and modulates lipid metabolism and antioxidant response in Caenorhabditis elegans. J. Agric. Food Chem. 2016, 64, 3462–3472. [Google Scholar] [CrossRef] [PubMed]
- Christiaen, S.E.; O’Connell Motherway, M.; Bottacini, F.; Lanigan, N.; Casey, P.G.; Huys, G.; Nelis, H.J.; van Sinderen, D.; Coenye, T. Autoinducer-2 plays a crucial role in gut colonization and probiotic functionality of Bifidobacterium breve UCC2003. PLoS ONE 2014, 9, e98111. [Google Scholar] [CrossRef] [PubMed]
- Komura, T.; Yasui, C.; Miyamoto, H.; Nishikawa, Y. Caenorhabditis elegans as an alternative model host for Legionella pneumophila, and protective effects of Bifidobacterium infantis. Appl. Environ. Microbiol. 2010, 76, 4105–4108. [Google Scholar] [CrossRef] [PubMed]
- Komura, T.; Ikeda, T.; Yasui, C.; Saeki, S.; Nishikawa, Y. Mechanism underlying prolongevity induced by bifidobacteria in Caenorhabditis elegans. Biogerontology 2013, 14, 73–87. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Mizuno, Y.; Komura, T.; Nishikawa, Y.; Kage-Nakadai, E. Toll-like receptor homolog TOL-1 regulates Bifidobacterium infantis-elicited longevity and behavior in Caenorhabditis elegans. Biosci. Microbiota. Food Health 2019, 38, 105–110. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Zhao, Y.; Liu, R.; Zheng, X.; Zhang, M.; Guo, H.; Zhang, H.; Ren, F. The transcription factor DAF-16 is essential for increased longevity in C. elegans exposed to Bifidobacterium longum BB68. Sci. Rep. 2017, 7, 7408. [Google Scholar] [CrossRef]
- Sugawara, T.; Sakamoto, K. Killed Bifidobacterium longum enhanced stress tolerance and prolonged life span of Caenorhabditis elegans via DAF-16. Br. J. Nutr. 2018, 120, 872–880. [Google Scholar] [CrossRef]
- Park, M.R.; Oh, S.; Son, S.J.; Park, D.J.; Kim, S.H.; Jeong, D.Y.; Oh, N.S.; Lee, Y.; Song, M.; Kim, Y. Bacillus licheniformis isolated from traditional Korean food resources enhances the longevity of Caenorhabditis elegans through serotonin signaling. J. Agric. Food Chem. 2015, 63, 10227–10233. [Google Scholar] [CrossRef]
- Kwon, G.; Lee, J.; Koh, J.H.; Lim, Y.H. Lifespan Extension of Caenorhabditis elegans by Butyricicoccus pullicaecorum and Megasphaera elsdenii with probiotic potential. Curr. Microbiol. 2018, 75, 557–564. [Google Scholar] [CrossRef]
- Kato, M.; Hamazaki, Y.; Sun, S.; Nishikawa, Y.; Kage-Nakadai, E. Clostridium butyricum MIYAIRI 588 increases the lifespan and multiple-stress resistance of Caenorhabditis elegans. Nutrients 2018, 10, 1921. [Google Scholar] [CrossRef] [PubMed]
- Yuen, G.J.; Ausubel, F.M. Both live and dead enterococci activate Caenorhabditis elegans host defense via immune and stress pathways. Virulence 2018, 9, 683–699. [Google Scholar] [CrossRef] [PubMed]
- Neuhaus, K.; Lamparter, M.C.; Zolch, B.; Landstorfer, R.; Simon, S.; Spanier, B.; Ehrmann, M.A.; Vogel, R.F. Probiotic Enterococcus faecalis Symbioflor((R)) down regulates virulence genes of EHEC in vitro and decrease pathogenicity in a Caenorhabditis elegans model. Arch. Microbiol. 2017, 199, 203–213. [Google Scholar] [CrossRef] [PubMed]
- Sim, I.; Park, K.T.; Kwon, G.; Koh, J.H.; Lim, Y.H. Probiotic potential of Enterococcus faecium isolated from chicken cecum with immunomodulating activity and promoting longevity in Caenorhabditis elegans. J. Microbiol. Biotechnol. 2018, 28, 883–892. [Google Scholar] [CrossRef] [PubMed]
- Hwang, I.Y.; Koh, E.; Wong, A.; March, J.C.; Bentley, W.E.; Lee, Y.S.; Chang, M.W. Engineered probiotic Escherichia coli can eliminate and prevent Pseudomonas aeruginosa gut infection in animal models. Nat. Commun. 2017, 8, 15028. [Google Scholar] [CrossRef] [PubMed]
- Kwon, G.; Lee, J.; Lim, Y.H. Dairy Propionibacterium extends the mean lifespan of Caenorhabditis elegans via activation of the innate immune system. Sci. Rep. 2016, 6, 31713. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Kwon, G.; Lim, Y.H. Elucidating the Mechanism of Weissella-dependent lifespan extension in Caenorhabditis elegans. Sci. Rep. 2015, 5, 17128. [Google Scholar] [CrossRef]
- Romanin, D.E.; Llopis, S.; Genoves, S.; Martorell, P.; Ramon, V.D.; Garrote, G.L.; Rumbo, M. Probiotic yeast Kluyveromyces marxianus CIDCA 8154 shows anti-inflammatory and anti-oxidative stress properties in in vivo models. Benef. Microbes. 2016, 7, 83–93. [Google Scholar] [CrossRef]
- Mattarelli, P.; Bonaparte, C.; Pot, B.; Biavati, B. Proposal to reclassify the three biotypes of Bifidobacterium longum as three subspecies: Bifidobacterium longum subsp. longum subsp. nov., Bifidobacterium longum subsp. infantis comb. nov. and Bifidobacterium longum subsp. suis comb. Nov. Int. J. Syst. Evol. Microbiol. 2008, 58, 767–772. [Google Scholar] [CrossRef]
- Kurz, C.L.; Ewbank, J.J. Caenorhabditis elegans: An emerging genetic model for the study of innate immunity. Nat. Rev. Genet. 2003, 4, 380–390. [Google Scholar] [CrossRef]
- Sun, X.; Chen, W.D.; Wang, Y.D. DAF-16/FOXO transcription factor in aging and longevity. Front. Pharm. 2017, 8, 548. [Google Scholar] [CrossRef] [PubMed]
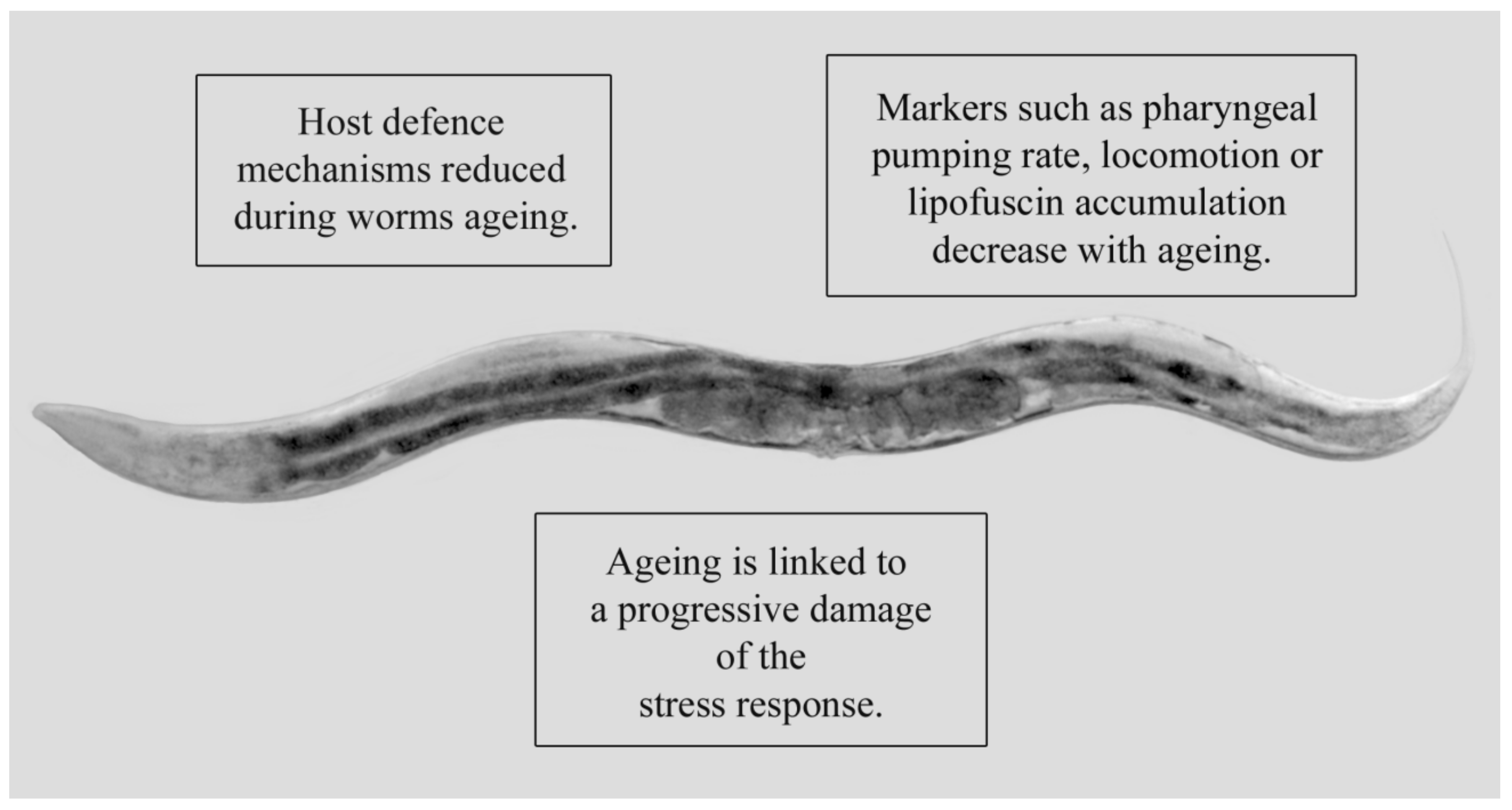
- Dues, D.J.; Andrews, E.K.; Schaar, C.E.; Bergsma, A.L.; Senchuk, M.M.; Van Raamsdonk, J.M. Aging causes decreased resistance to multiple stresses and a failure to activate specific stress response pathways. Aging 2016, 8, 777–795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Irazoqui, J.E.; Urbach, J.M.; Ausubel, F.M. Evolution of host innate defence: Insights from Caenorhabditis elegans and primitive invertebrates. Nat. Rev. Immunol. 2010, 10, 47–58. [Google Scholar] [CrossRef] [PubMed]
- Pan, H.; Finkel, T. Key proteins and pathways that regulate lifespan. J. Biol. Chem. 2017, 292, 6452–6460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jesko, H.; Stepien, A.; Lukiw, W.J.; Strosznajder, R.P. The Cross-talk between sphingolipids and insulin-like growth factor signaling: significance for aging and neurodegeneration. Mol. Neurobiol. 2019, 56, 3501–3521. [Google Scholar] [CrossRef] [PubMed]
- Ristow, M.; Schmeisser, S. Extending life span by increasing oxidative stress. Free Radic. Biol. Med. 2011, 51, 327–336. [Google Scholar] [CrossRef] [Green Version]
- Tullet, J.M.; Hertweck, M.; An, J.H.; Baker, J.; Hwang, J.Y.; Liu, S.; Oliveira, R.P.; Baumeister, R.; Blackwell, T.K. Direct inhibition of the longevity-promoting factor SKN-1 by insulin-like signaling in C. elegans. Cell 2008, 132, 1025–1038. [Google Scholar] [CrossRef]
- Oh, S.W.; Mukhopadhyay, A.; Svrzikapa, N.; Jiang, F.; Davis, R.J.; Tissenbaum, H.A. JNK regulates lifespan in Caenorhabditis elegans by modulating nuclear translocation of forkhead transcription factor/DAF-16. Proc. Natl. Acad. Sci. USA 2005, 102, 4494–4499. [Google Scholar] [CrossRef]
- Hu, Q.; D’Amora, D.R.; MacNeil, L.T.; Walhout, A.J.M.; Kubiseski, T.J. The Caenorhabditis elegans oxidative stress response requires the NHR-49 transcription factor. G3 2018, 8, 3857–3863. [Google Scholar] [CrossRef]
- Zugasti, O.; Ewbank, J.J. Neuroimmune regulation of antimicrobial peptide expression by a noncanonical TGF-beta signaling pathway in Caenorhabditis elegans epidermis. Nat. Immunol. 2009, 10, 249–256. [Google Scholar] [CrossRef]
- Irazoqui, J.E.; Ng, A.; Xavier, R.J.; Ausubel, F.M. Role for beta-catenin and HOX transcription factors in Caenorhabditis elegans and mammalian host epithelial-pathogen interactions. Proc. Natl. Acad. Sci. USA 2008, 105, 17469–17474. [Google Scholar] [CrossRef] [PubMed]
- Tenor, J.L.; Aballay, A. A conserved Toll-like receptor is required for Caenorhabditis elegans innate immunity. EMBO Rep. 2008, 9, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Lebeer, S.; Vanderleyden, J.; De Keersmaecker, S.C. Host interactions of probiotic bacterial surface molecules: Comparison with commensals and pathogens. Nat. Rev. Microbiol. 2010, 8, 171–184. [Google Scholar] [CrossRef] [PubMed]
- Alper, S.; McBride, S.J.; Lackford, B.; Freedman, J.H.; Schwartz, D.A. Specificity and complexity of the Caenorhabditis elegans innate immune response. Mol. Cell Biol. 2007, 27, 5544–5553. [Google Scholar] [CrossRef] [PubMed]
- Smolentseva, O.; Gusarov, I.; Gautier, L.; Shamovsky, I.; DeFrancesco, A.S.; Losick, R.; Nudler, E. Mechanism of biofilm-mediated stress resistance and lifespan extension in C. elegans. Sci. Rep. 2017, 7, 7137. [Google Scholar] [CrossRef] [PubMed]
- Yun, H.S.; Heo, J.H.; Son, S.J.; Park, M.R.; Oh, S.; Song, M.H.; Kim, J.N.; Go, G.W.; Cho, H.S.; Choi, N.J.; et al. Bacillus licheniformis isolated from Korean traditional food sources enhances the resistance of Caenorhabditis elegans to infection by Staphylococcus aureus. J. Microbiol. Biotechnol. 2014, 24, 1105–1108. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Genus | Species | Strain(s) | Nematode Signaling Pathway(s) Influenced | References |
|---|---|---|---|---|
| Lactobacillus | acidophilus | NCFM | p38 MAPK beta-catenin | [29] |
| brevis | SDL1411 | unknown | [30] | |
| casei | CL11 LAB9 | unknown p38 MAPK | [31] [32,33] | |
| coryniformis | H307.6 | unknown | [34] | |
| delbrueckii | bulgaricus ATCC11842; lactis LMG6401; lactis 23 | unknown | [35] | |
| fermentum | MBC2 JDFM216 LA12 LF21 | unknown p38 MAPK unknown IIS | [36] [37] [38] [39] | |
| gasseri | SBT2055 | p38 MAPK | [9] | |
| helveticus | NBRC15019 | unknown | [40] | |
| murinus | CR147 | unknown | [41] | |
| paracasei | 28.4 | unknown | [42] | |
| plantarum | CAU1054; CAU1055; CAU1064; CAU1106 JDFM60; JDFM440; JDFM970; JDFM1000 CJLP133 K90 NBRC15891 | unknown unknown IIS unknown unknown | [43] [44] [38,39] [45] [40] | |
| pentosus | D303.36 | unknown | [34] | |
| reuteri | CL9 S64 DSM 20016 | unknown unknown unknown | [46] [31] [47] | |
| rhamnosus | R4 CNCM I-3690 NBRC14710 | unknown IIS unknown | [48] [49] [40] | |
| salivarius | FDB89 DSM 20555 | unknown unknown | [50] [47] | |
| zeae | LB1 | p38 MAPK IIS | [51,52] | |
| Bifidobacterium | animalis subsp. lactis | CECT8145 | IIS | [53] |
| breve | UCC2003 | unknown | [54] | |
| Infantis1 | ATCC15697 | p38 MAPK IIS | [40,55,56,57] | |
| longum | ATCC15707 BB68 BR-108 | unknown JNK IIS | [29,40] [58] [59] | |
| Bacillus | licheniformis | 141 | unknown | [60] |
| Butyricicoccus | pullicaecorum | KCTC 15070 | TGF-beta | [61] |
| Clostridium | butyricum | MIYAIRI 588 (CBM 588) | IIS | [62] |
| Enterococcus | faecalis | MMH594 Symbioflor® | p38 MAPK beta-catenin unknown | [63] [64] |
| faecium | L11 E007 | TGF-beta p38 MAPK p38 MAPK beta-catenin | [65] [63] | |
| Escherichia | coli | Nissle 1917 | unknown | [66] |
| Megasphaera | elsdenii | KCTC 5187 | TGF-beta | [61] |
| Pediococcus | acidilactici | DSM 20284 DM-9 | unknown unknown | [47] [30] |
| pentosaceus | SDL1409 | unknown | [30] | |
| Propionibacterium | freudenreichii | KCTC 1063 | p38 MAPK | [67] |
| Weissella | cibaria | KACC11845 | JNK AMPK | [68] |
| koreensis | KACC11853 | JNK AMPK | ||
| Kluyveromyces | marxianus | CIDCA 8154 | p38 MAPK | [69] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roselli, M.; Schifano, E.; Guantario, B.; Zinno, P.; Uccelletti, D.; Devirgiliis, C. Caenorhabditis Elegans and Probiotics Interactions from a Prolongevity Perspective. Int. J. Mol. Sci. 2019, 20, 5020. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20205020
Roselli M, Schifano E, Guantario B, Zinno P, Uccelletti D, Devirgiliis C. Caenorhabditis Elegans and Probiotics Interactions from a Prolongevity Perspective. International Journal of Molecular Sciences. 2019; 20(20):5020. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20205020
Chicago/Turabian StyleRoselli, Marianna, Emily Schifano, Barbara Guantario, Paola Zinno, Daniela Uccelletti, and Chiara Devirgiliis. 2019. "Caenorhabditis Elegans and Probiotics Interactions from a Prolongevity Perspective" International Journal of Molecular Sciences 20, no. 20: 5020. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20205020