miR-7 Knockdown by Peptide Nucleic Acids in the Ascidian Ciona intestinalis

 ,
,  , ,
, , 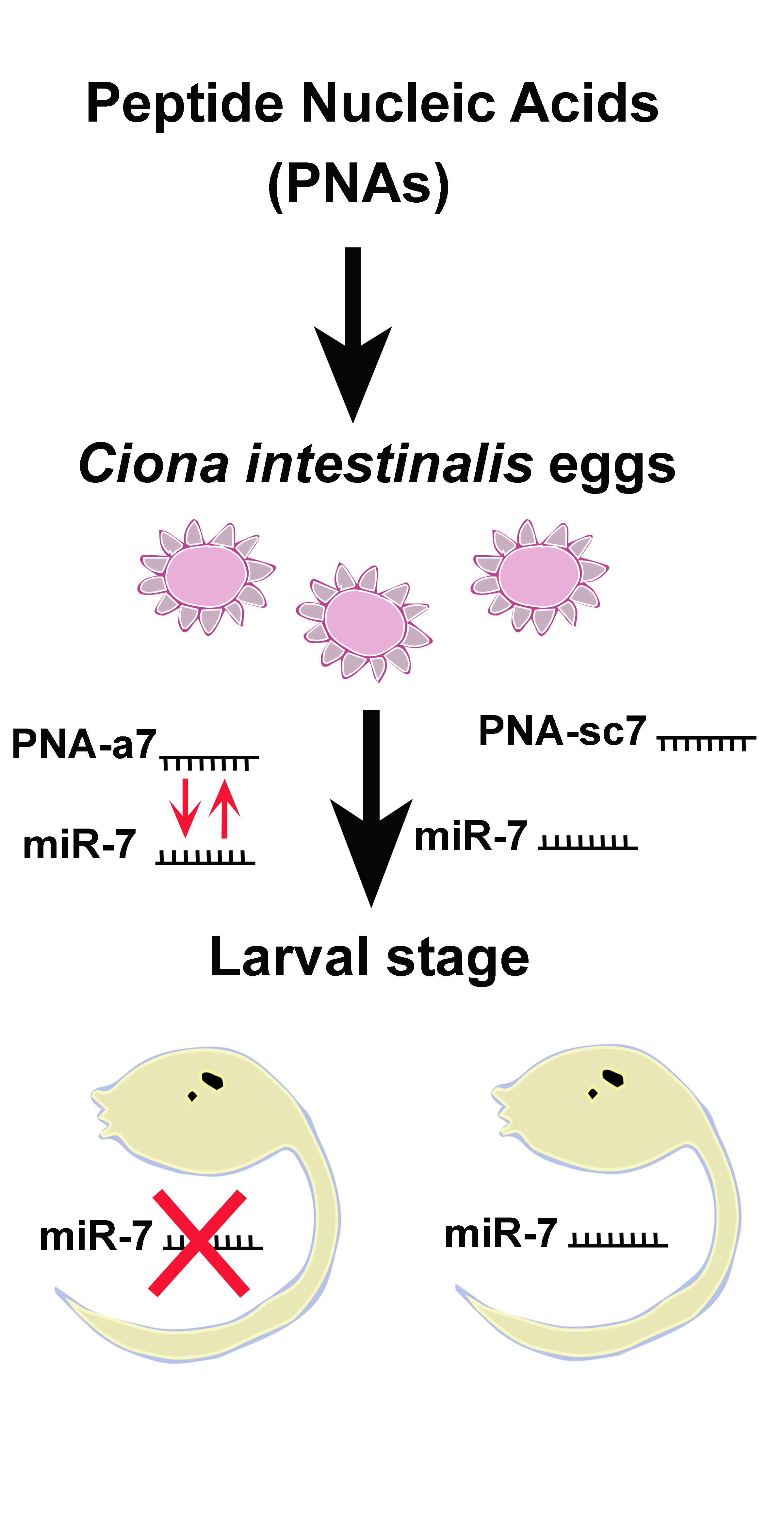{kind=link}
{kind=link}
{kind=link}
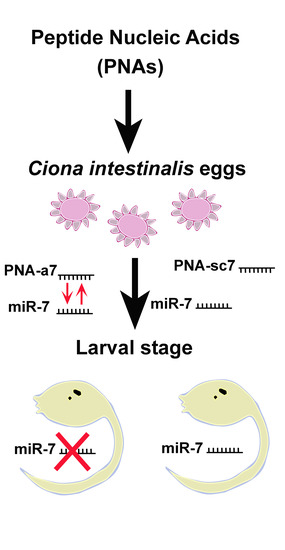{kind=link}
{kind=link}
{kind=link}
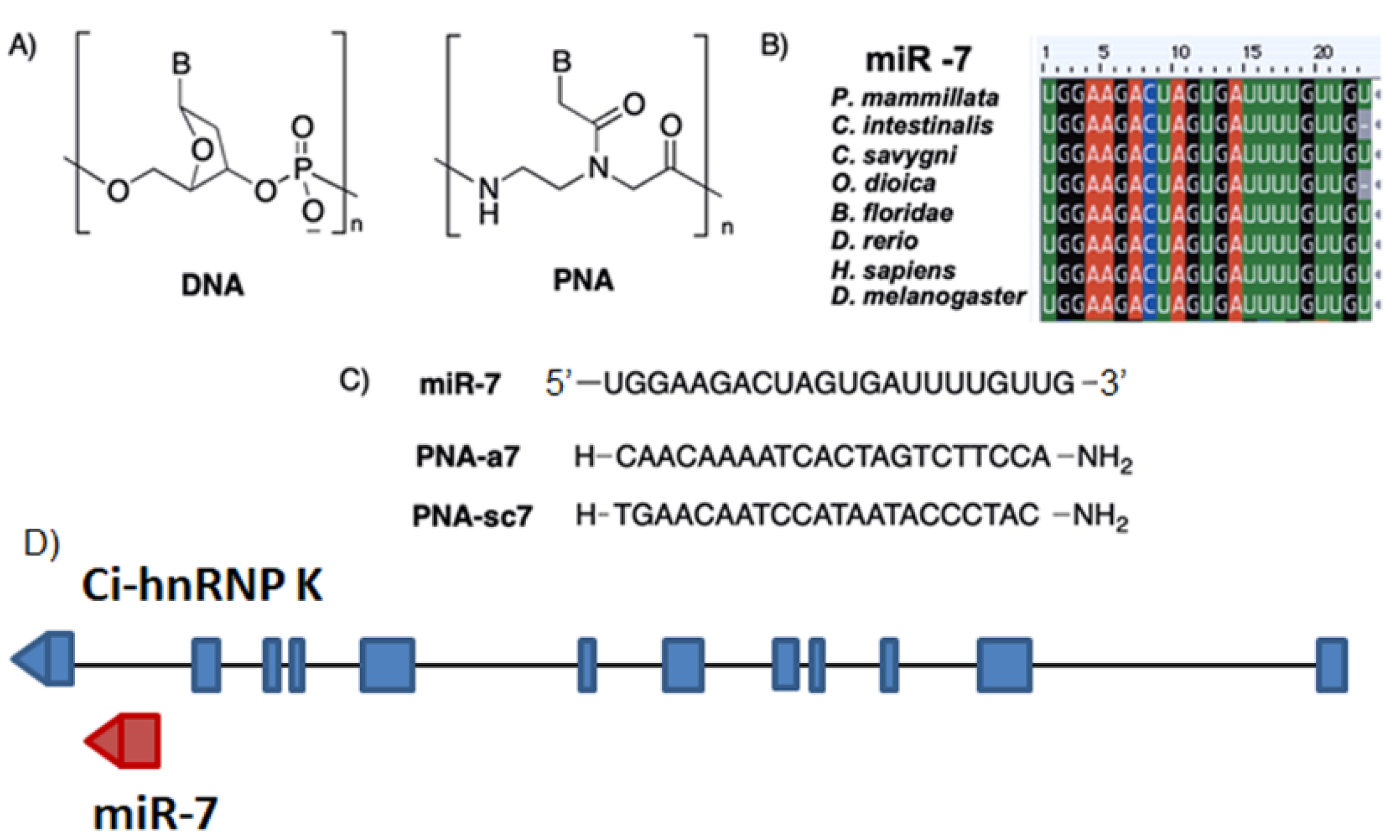{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
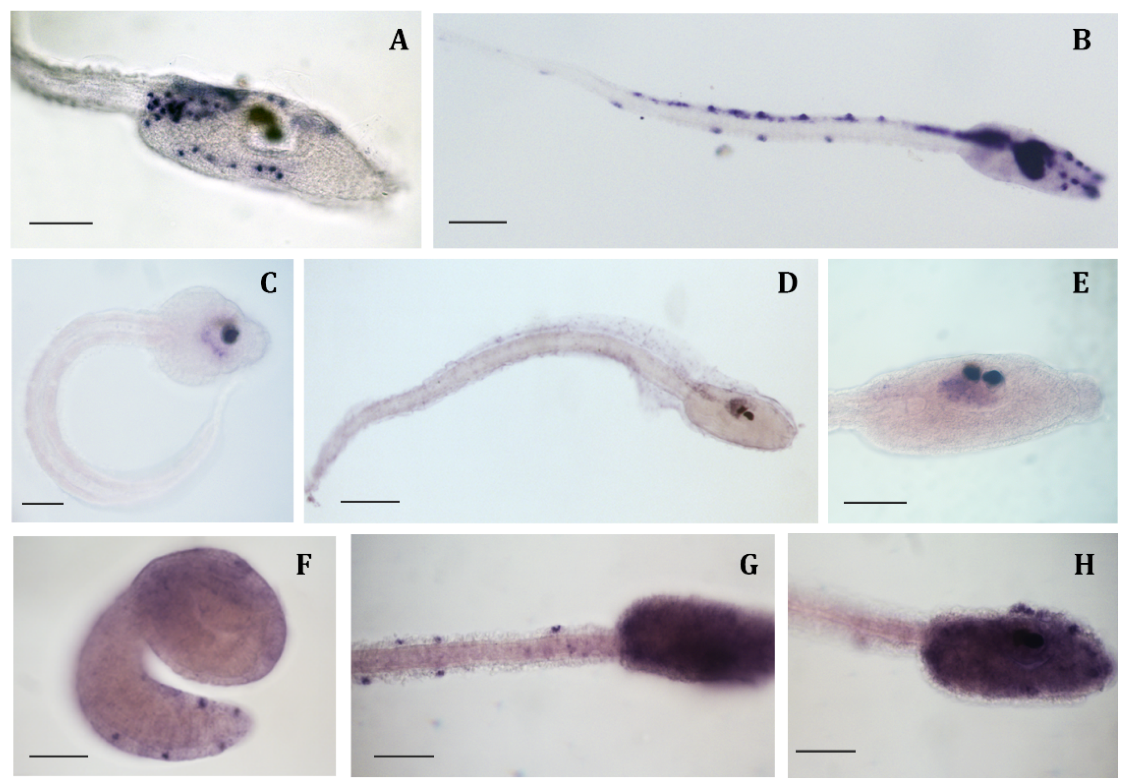
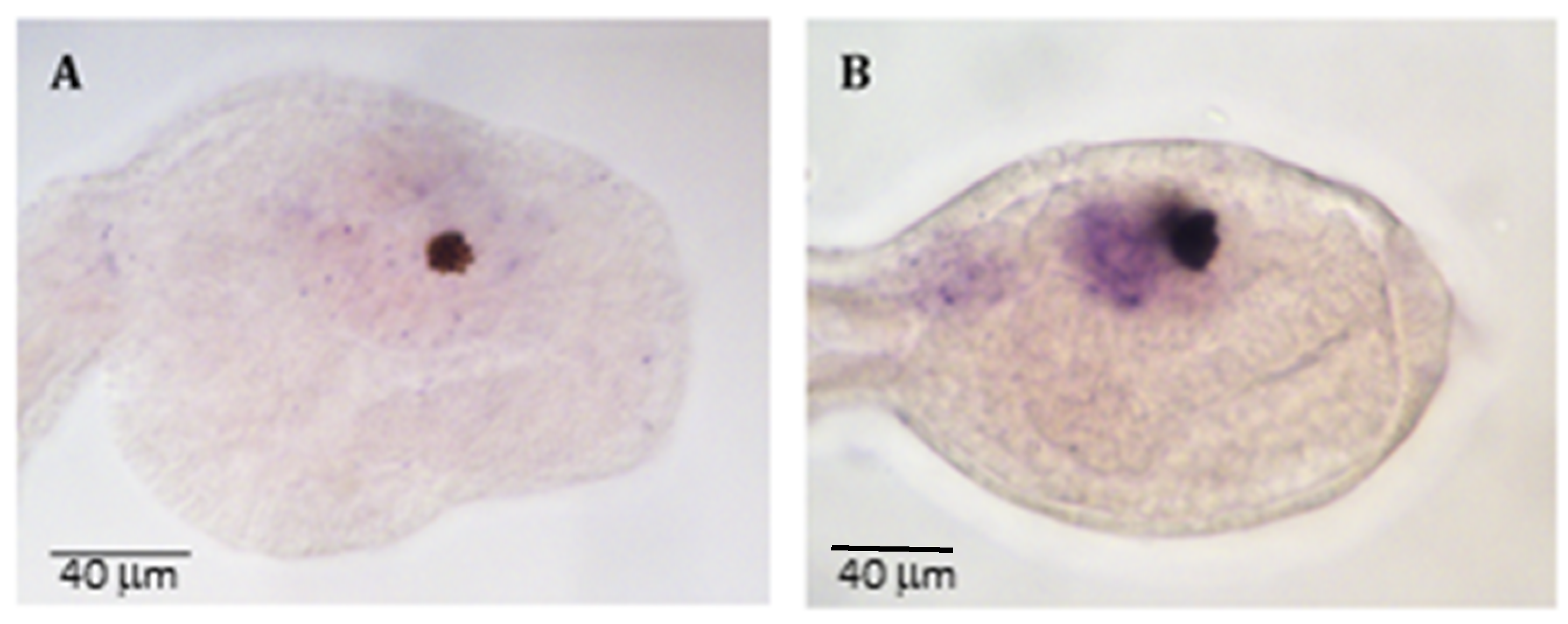
2.1. miR-7 in Ciona intestinalis
2.2. Genes Expression Profile
2.3. miR-7 Downregulation by PNAs
3. Discussion
4. Materials and Methods
4.1. Animals and Embryos Culture
4.2. Reagents
4.3. PNAs Synthesis and Characterization
4.4. Microinjections
4.5. Whole Mount In Situ Hybridization
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ASWH | Artificial Sea Water with HEPES |
| AmiR-7 | AntagomiR anti-miR-7 |
| Boc | tert-Butyloxycarbonyl |
| ESI-Q-TOF MS | Electrospray-ionisation quadrupole time-of-flight mass spectrometry |
| LNA | Locked Nucleic Acid |
| MBHA | 4-Methylbenzhydrylamine hydrochloride |
| PNA | Peptide Nucleic Acid |
| PNA-a7 | Peptide Nucleic Acid anti-miR-7 |
| PNA-sc7 | Peptide Nucleic Acid scrambled sequence |
| RP-HPLC | Reverse phase-high pressure liquid chromatography |
| Z | Benzyloxycarbonyl |
References
- Nielsen, P.E.; Egholm, M.; Berg, R.; Buchardt, O. Sequence-selective recognition of DNA by strand displacement with a thymine-substituted polyamide. Science 1991, 254, 1497–1500. [Google Scholar] [CrossRef] [PubMed]
- Peter, E.N. Gene Targeting and Expression Modulation by Peptide Nucleic Acids (PNA). Curr. Pharm. Des. 2010, 16, 3118–3123. [Google Scholar]
- Egholm, M.; Buchardt, O.; Christensen, L.; Behrens, C.; Freier, S.M.; Driver, D.A.; Berg, R.H.; Kim, S.K.; Norden, B.; Nielsen, P.E. PNA hybridizes to complementary oligonucleotides obeying the Watson–Crick hydrogen-bonding rules. Nature 1993, 365, 566–568. [Google Scholar] [CrossRef] [PubMed]
- Karkare, S.; Bhatnagar, D. Promising nucleic acid analogs and mimics: Characteristic features and applications of PNA, LNA, and morpholino. Appl. Microbiol. Biotechnol. 2006, 71, 575–586. [Google Scholar] [CrossRef]
- Hyrup, B.; Nielsen, P.E. Peptide nucleic acids (PNA): Synthesis, properties and potential applications. Bioorg. Med. Chem. 1996, 4, 5–23. [Google Scholar] [CrossRef]
- Shakeel, S.; Karim, S.; Ali, A. Peptide nucleic acid (PNA)—A review. J. Chem. Technol. Biotechnol. 2006, 81, 892–899. [Google Scholar] [CrossRef]
- Hatamoto, M.; Ohashi, A.; Imachi, H. Peptide nucleic acids (PNAs) antisense effect to bacterial growth and their application potentiality in biotechnology. Appl. Microbiol. Biotechnol. 2010, 86, 397–402. [Google Scholar] [CrossRef]
- Fabani, M.M.; Abreu-Goodger, C.; Williams, D.; Lyons, P.A.; Torres, A.G.; Smith, K.G.C.; Enright, A.J.; Gait, M.J.; Vigorito, E. Efficient inhibition of miR-155 function in vivo by peptide nucleic acids. Nucleic Acids Res. 2010, 38, 4466–4475. [Google Scholar] [CrossRef] [Green Version]
- Yan, L.X.; Wu, Q.N.; Zhang, Y.; Li, Y.Y.; Liao, D.Z.; Hou, J.H.; Fu, J.; Zeng, M.S.; Yun, J.P.; Wu, Q.L. Knockdown of miR-21 in human breast cancer cell lines inhibits proliferation, in vitro migration and in vivo tumor growth. Breast Cancer Res. 2011, 13, R2. [Google Scholar] [CrossRef]
- Nielsen, P.E. Addressing the challenges of cellular delivery and bioavailability of peptide nucleic acids (PNA). Q. Rev. Biophys. 2006, 38, 345–350. [Google Scholar] [CrossRef]
- Rozners, E. Recent Advances in Chemical Modification of Peptide Nucleic Acids. J. Nucleic acids 2012, 2012, 518162. [Google Scholar] [CrossRef] [PubMed]
- Delsuc, F.; Brinkmann, H.; Chourrout, D.; Philippe, H. Tunicates and not cephalochordates are the closest living relatives of vertebrates. Nature 2006, 439, 965–968. [Google Scholar] [CrossRef] [PubMed]
- Passamaneck, Y.J.; Di Gregorio, A. Ciona intestinalis: Chordate development made simple. Dev. Dyn. 2005, 233, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Satoh, N. The ascidian tadpole larva: Comparative molecular development and genomics. Nature Rev. Genet. 2003, 4, 285–295. [Google Scholar] [CrossRef] [PubMed]
- Kari, W.; Zeng, F.; Zitzelsberger, L.; Will, J.; Rothbächer, U. Embryo Microinjection and Electroporation in the Chordate Ciona intestinalis. J. Vis. Exp. 2016, 116, e54313. [Google Scholar] [CrossRef] [PubMed]
- Stolfi, A.; Gandhi, S.; Salek, F.; Christiaen, L. Tissue-specific genome editing in Ciona embryos by CRISPR/Cas9. Development 2014, 141, 4115–4120. [Google Scholar] [CrossRef] [PubMed]
- Prochnik, S.E.; Rokhsar, D.S.; Aboobaker, A.A. Evidence for a microRNA expansion in the bilaterian ancestor. Dev. Genes Evol. 2007, 217, 73–77. [Google Scholar] [CrossRef] [PubMed]
- Alberti, C.; Cochella, L. A framework for understanding the roles of miRNAs in animal development. Development 2017, 144, 2548–2559. [Google Scholar] [CrossRef] [Green Version]
- Paul, P.; Chakraborty, A.; Sarkar, D.; Langthasa, M.; Rahman, M.; Bari, M.; Singha, R.S.; Malakar, A.K.; Chakraborty, S. Interplay between miRNAs and human diseases. J. Cell. Physiol. 2018, 233, 2007–2018. [Google Scholar] [CrossRef]
- Ambros, V. The functions of animal microRNAs. Nature 2004, 431, 350–355. [Google Scholar] [CrossRef]
- Smibert, P.; Lai, E.C. Lessons from microRNA mutants in worms, flies and mice. Cell Cycle 2008, 7, 2500–2508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bravo-Egana, V.; Rosero, S.; Molano, R.D.; Pileggi, A.; Ricordi, C.; Domínguez-Bendala, J.; Pastori, R.L. Quantitative differential expression analysis reveals miR-7 as major islet microRNA. Biochem. Biophys. Res. Commun. 2008, 366, 922–926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanek, N.A.; Young, W.S. Investigating the In Vivo Expression Patterns of miR-7 microRNA Family Members in the Adult Mouse Brain. MicroRNA 2012, 1, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Tessmar-Raible, K.; Raible, F.; Christodoulou, F.; Guy, K.; Rembold, M.; Hausen, H.; Arendt, D. Conserved Sensory-Neurosecretory Cell Types in Annelid and Fish Forebrain: Insights into Hypothalamus Evolution. Cell 2007, 129, 1389–1400. [Google Scholar] [CrossRef] [Green Version]
- Huang, K.M.; Dentchev, T.; Stambolian, D. MiRNA expression in the eye. Mamm. Genome 2008, 19, 510–516. [Google Scholar] [CrossRef]
- Candiani, S.; Moronti, L.; De Pietri Tonelli, D.; Garbarino, G.; Pestarino, M. A study of neural-related microRNAs in the developing amphioxus. EvoDevo 2011, 2, 15. [Google Scholar] [CrossRef]
- Li, X.; Cassidy, J.J.; Reinke, C.A.; Fischboeck, S.; Carthew, R.W. A MicroRNA Imparts Robustness against Environmental Fluctuation during Development. Cell 2009, 137, 273–282. [Google Scholar] [CrossRef] [Green Version]
- Mattes, J.; Yang, M.; Foster, P.S. Regulation of MicroRNA by Antagomirs. Am. J. Respir.Cell Mol. Biol. 2007, 36, 8–12. [Google Scholar] [CrossRef]
- Messinetti, S.; Mercurio, S.; Pennati, R. Bisphenol A affects neural development of the ascidian Ciona robusta. J. Exp. Zool. 2019, 331, 5–16. [Google Scholar] [CrossRef]
- Chen, J.S.; Pedro, M.S.; Zeller, R.W. miR-124 function during Ciona intestinalis neuronal development includes extensive interaction with the Notch signaling pathway. Development 2011, 138, 4943–4953. [Google Scholar] [CrossRef]
- Satou, Y.; Takatori, N.; Yamada, L.; Mochizuki, Y.; Hamaguchi, M.; Ishikawa, H.; Chiba, S.; Imai, K.; Kano, S.; Murakami, S.D.; et al. Gene expression profiles in Ciona intestinalis tailbud embryos. Development 2001, 128, 2893–2904. [Google Scholar] [PubMed]
- Candiani, S.; Moronti, L.; Pennati, R.; De Bernardi, F.; Benfenati, F.; Pestarino, M. The synapsin gene family in basal chordates: Evolutionary perspectives in metazoans. BMC Evol. Biol. 2010, 10, 32. [Google Scholar] [CrossRef] [PubMed]
- Christodoulou, F.; Raible, F.; Tomer, R.; Simakov, O.; Trachana, K.; Klaus, S.; Snyman, H.; Hannon, G.J.; Bork, P.; Arendt, D. Ancient animal microRNAs and the evolution of tissue identity. Nature 2010, 463, 1084–1088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamada, M.; Shimozono, N.; Ohta, N.; Satou, Y.; Horie, T.; Kawada, T.; Satake, H.; Sasakura, Y.; Satoh, N. Expression of neuropeptide- and hormone-encoding genes in the Ciona intestinalis larval brain. Dev. Biol. 2011, 352, 202–214. [Google Scholar] [CrossRef] [Green Version]
- Moret, F.; Christiaen, L.; Deyts, C.; Blin, M.; Joly, J.-S.; Vernier, P. The dopamine-synthesizing cells in the swimming larva of the tunicate Ciona intestinalis are located only in the hypothalamus-related domain of the sensory vesicle. Eur. J. Neurosci. 2005, 21, 3043–3055. [Google Scholar] [CrossRef]
- Razy-Krajka, F.; Brown, E.R.; Horie, T.; Callebert, J.; Sasakura, Y.; Joly, J.-S.; Kusakabe, T.G.; Vernier, P. Monoaminergic modulation of photoreception in ascidian: Evidence for a proto-hypothalamo-retinal territory. BMC Biol. 2012, 10, 45. [Google Scholar] [CrossRef]
- D’Aniello, S.; D’Aniello, E.; Locascio, A.; Memoli, A.; Corrado, M.; Russo, M.T.; Aniello, F.; Fucci, L.; Brown, E.R.; Branno, M. The ascidian homolog of the vertebrate homeobox gene Rx is essential for ocellus development and function. Differentiation 2006, 74, 222–234. [Google Scholar] [CrossRef]
- Choudhury, N.R.; de Lima Alves, F.; de Andrés-Aguayo, L.; Graf, T.; Cáceres, J.F.; Rappsilber, J.; Michlewski, G. Tissue-specific control of brain-enriched miR-7 biogenesis. Genes Dev. 2013, 27, 24–38. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Githinji, J.; Mclaughlin, B.; Wilczek, K.; Nolta, J. Role of miRNAs in Neuronal Differentiation from Human Embryonic Stem Cell—Derived Neural Stem Cells. Stem Cell. Rev. 2012, 8, 1129–1137. [Google Scholar] [CrossRef]
- Bomsztyk, K.; Denisenko, O.; Ostrowski, J. hnRNP K: One protein multiple processes. Bioessays 2004, 26, 629–638. [Google Scholar] [CrossRef]
- Liu, Y.; Gervasi, C.; Szaro, B.G. A crucial role for hnRNP K in axon development in Xenopus laevis. Development 2008, 135, 3125–3135. [Google Scholar] [CrossRef] [PubMed]
- Piva, R.; Spandidos, D.A.; Gambari, R. From microRNA functions to microRNA therapeutics: Novel targets and novel drugs in breast cancer research and treatment. Int. J. Oncol. 2013, 43, 985–994. [Google Scholar] [CrossRef] [PubMed]
- Prencipe, G.; Maiorana, S.; Verderio, P.; Colombo, M.; Fermo, P.; Caneva, E.; Prosperi, D.; Licandro, E. Magnetic peptide nucleic acids for DNA targeting. Chem. Commun. 2009, 40, 6017–6019. [Google Scholar] [CrossRef] [PubMed]
- Giesen, U.; Kleider, W.; Berding, C.; Geiger, A.; Orum, H.; Nielsen, P.E. A formula for thermal stability (Tm) prediction of PNA/DNA duplexes. Nucleic Acids Res. 1998, 26, 5004–5006. [Google Scholar] [CrossRef] [PubMed]
- SantaLucia, J.; Allawi, H.T.; Seneviratne, P.A. Improved nearest-neighbor parameters for predicting DNA duplex stability. Biochemistry 1996, 35, 3555–3562. [Google Scholar] [CrossRef]
- Hotta, K.; Mitsuhara, K.; Takahashi, H.; Inaba, K.; Oka, K.; Gojobori, T.; Ikeo, K. A web-based interactive developmental table for the ascidian Ciona intestinalis, including 3D real-image embryo reconstructions: I. From fertilized egg to hatching larva. Dev. Dyn. 2007, 236, 1790–1805. [Google Scholar] [CrossRef]
- Satou, Y.; Yamada, L.; Mochizuki, Y.; Takatori, N.; Kawashima, T.; Sasaki, A.; Hamaguchi, M.; Awazu, S.; Yagi, K.; Sasakura, Y.; et al. A cDNA resource from the basal chordate Ciona intestinalis. Genesis 2002, 33, 153–154. [Google Scholar] [CrossRef]
- Urtishak, K.A.; Choob, M.; Tian, X.; Sternheim, N.; Talbot, W.S.; Wickstrom, E.; Farber, S.A. Targeted gene knockdown in zebrafish using negatively charged peptide nucleic acid mimics. Dev. Dyn. 2003, 228, 405–413. [Google Scholar] [CrossRef]




© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mercurio, S.; Cauteruccio, S.; Manenti, R.; Candiani, S.; Scarì, G.; Licandro, E.; Pennati, R. miR-7 Knockdown by Peptide Nucleic Acids in the Ascidian Ciona intestinalis. Int. J. Mol. Sci. 2019, 20, 5127. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20205127
Mercurio S, Cauteruccio S, Manenti R, Candiani S, Scarì G, Licandro E, Pennati R. miR-7 Knockdown by Peptide Nucleic Acids in the Ascidian Ciona intestinalis. International Journal of Molecular Sciences. 2019; 20(20):5127. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20205127
Chicago/Turabian StyleMercurio, Silvia, Silvia Cauteruccio, Raoul Manenti, Simona Candiani, Giorgio Scarì, Emanuela Licandro, and Roberta Pennati. 2019. "miR-7 Knockdown by Peptide Nucleic Acids in the Ascidian Ciona intestinalis" International Journal of Molecular Sciences 20, no. 20: 5127. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20205127