Age- and BMI-Associated Expression of Angiogenic Factors in White Adipose Tissue of Children

 ,
,
Abstract
:1. Introduction
2. Results
2.1. General Characteristics of the Ulm Childhood Adipose Tissue Collective
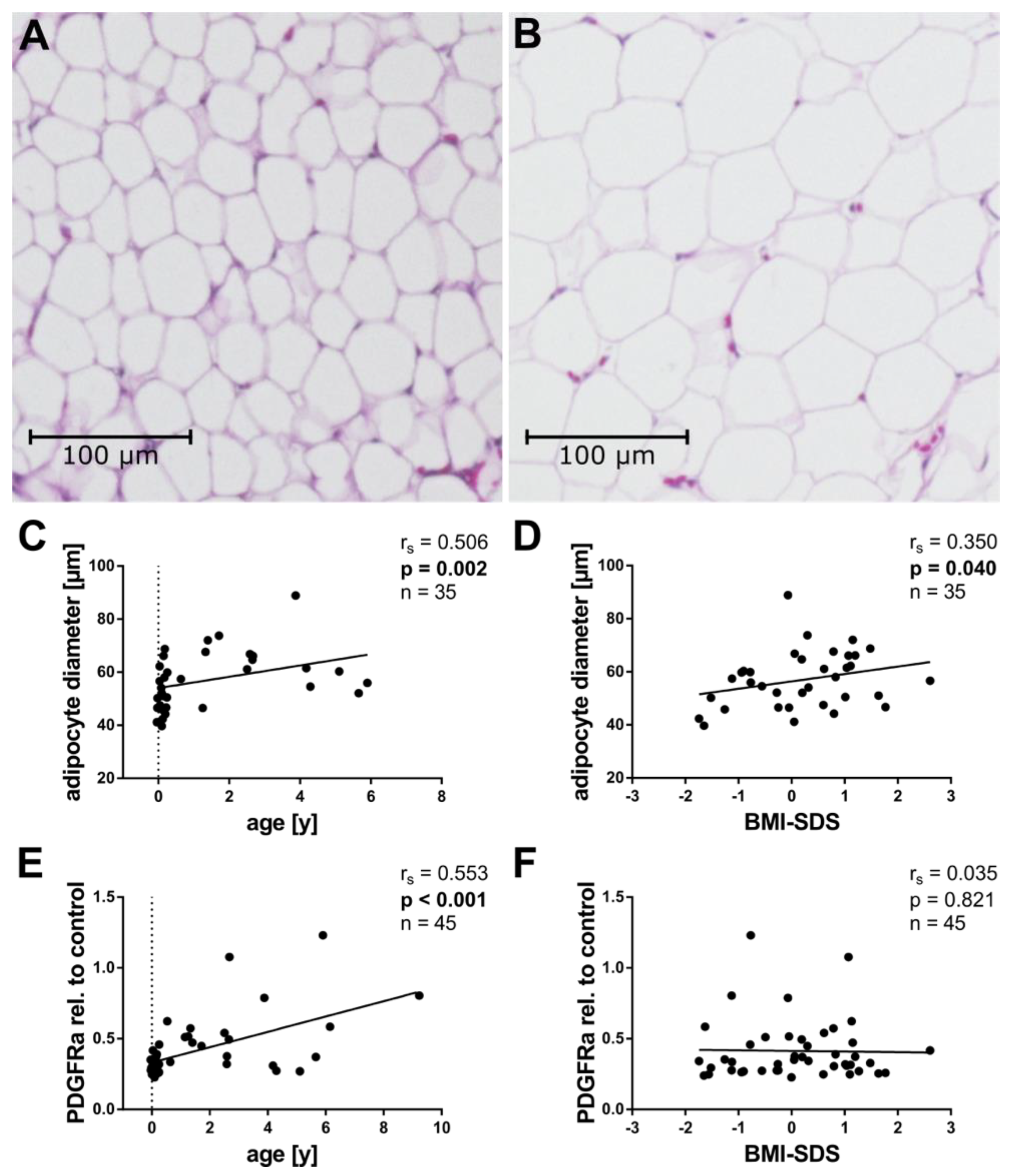
2.2. Tissue Cellularity
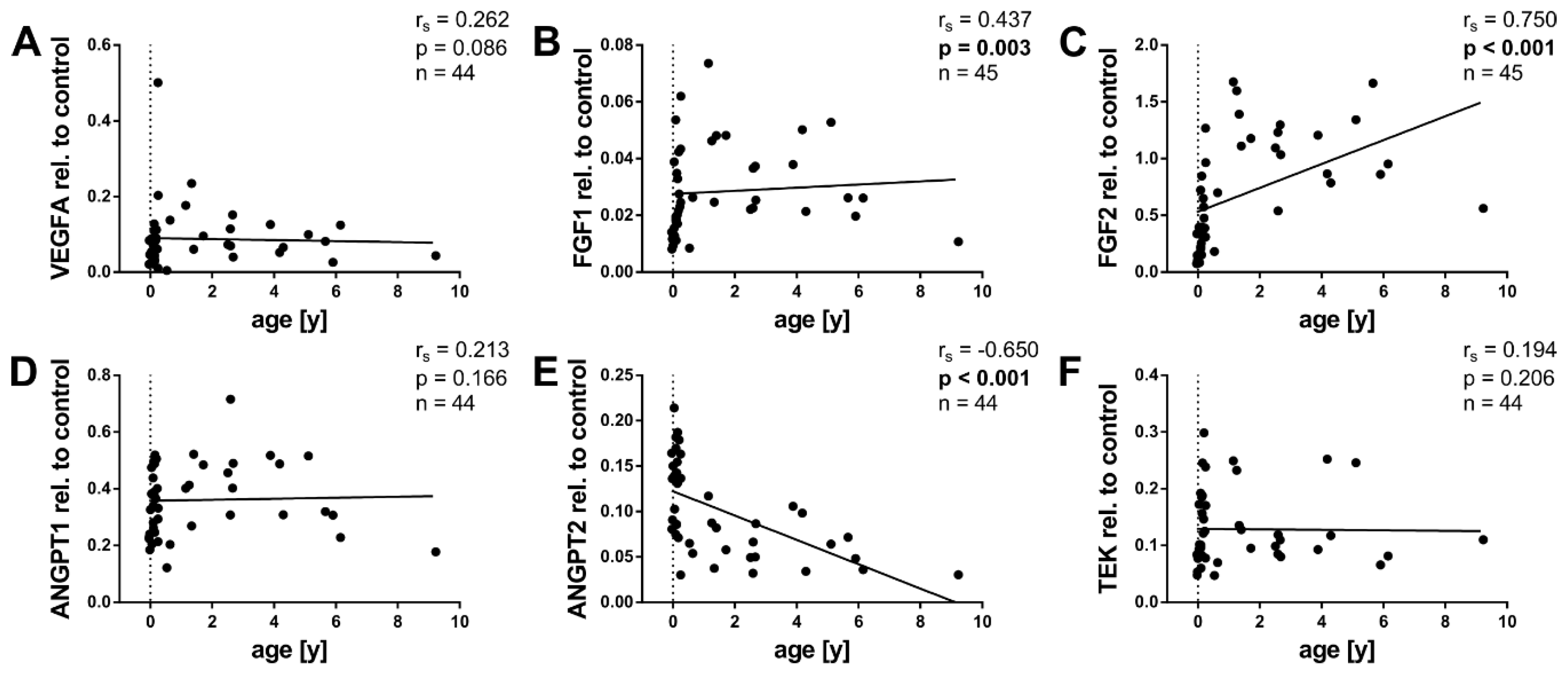
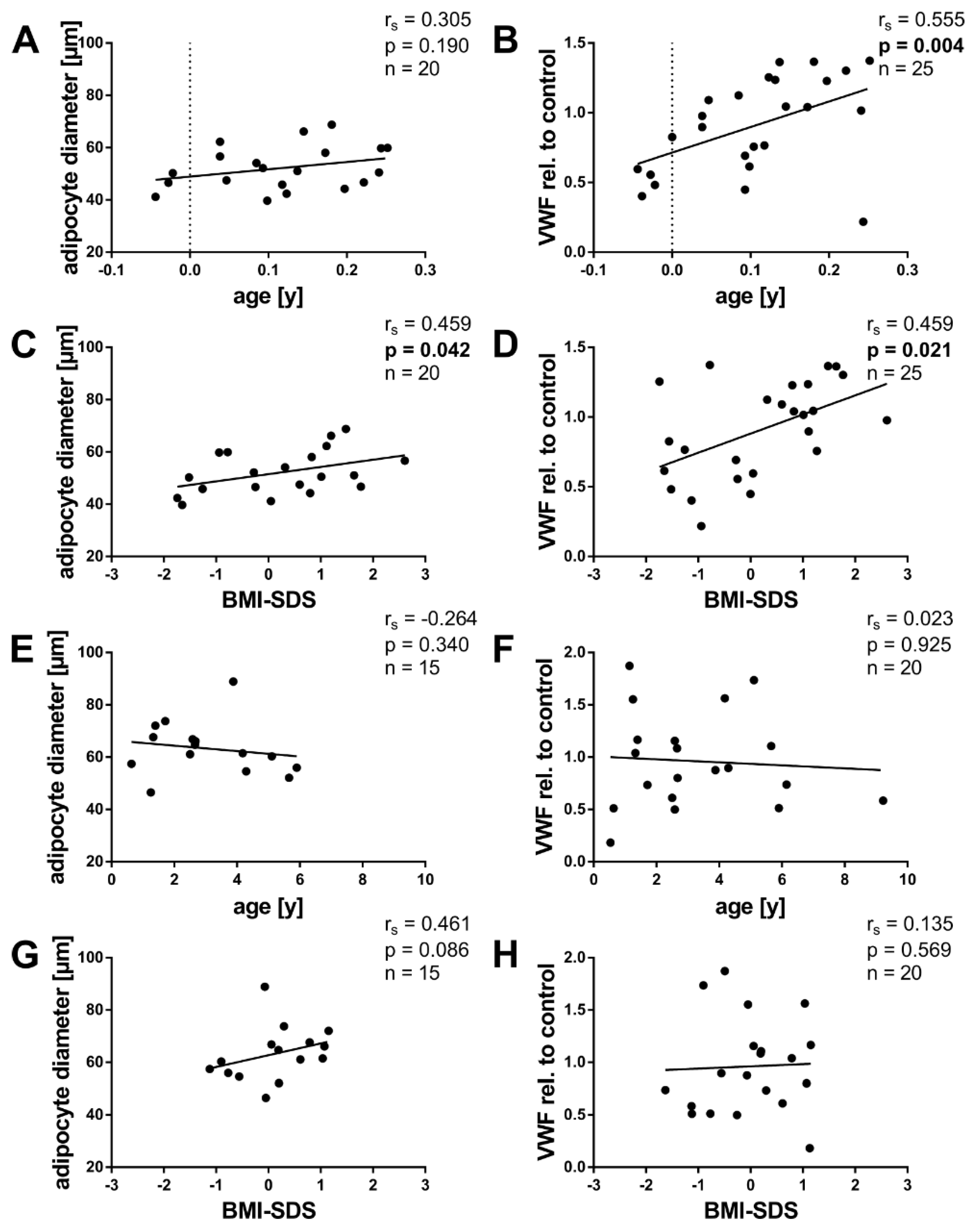
2.3. Angiogenic Factors and Age
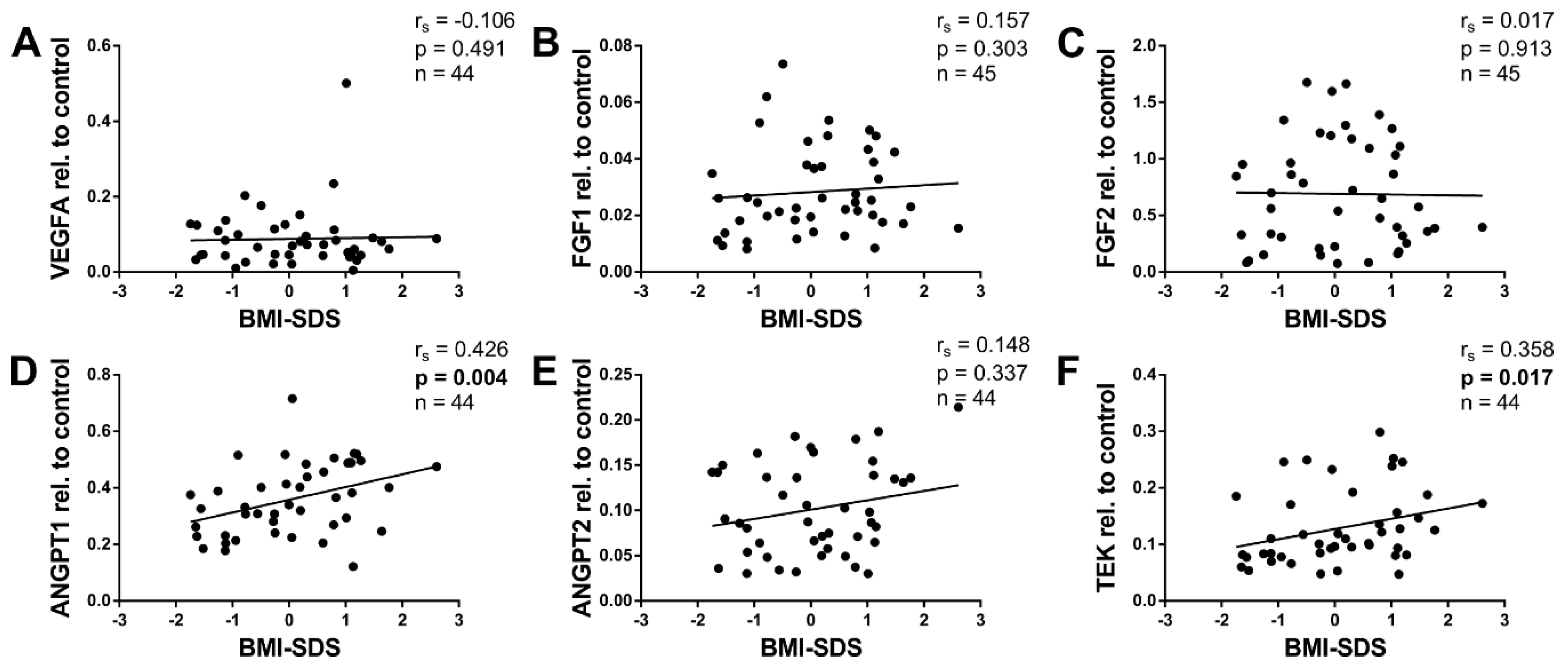
2.4. Angiogenic Factors and BMI-SDS
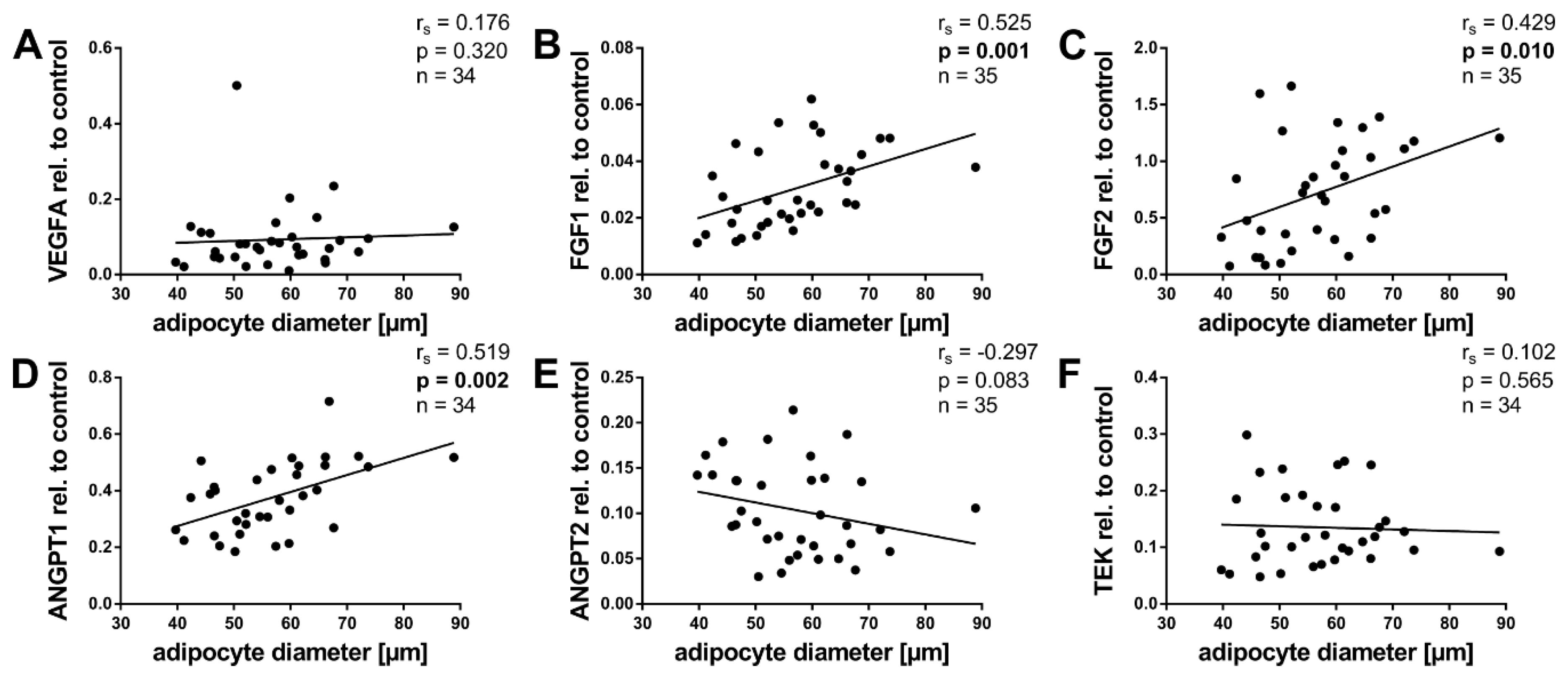
2.5. Angiogenic Factors and Adipocyte Size
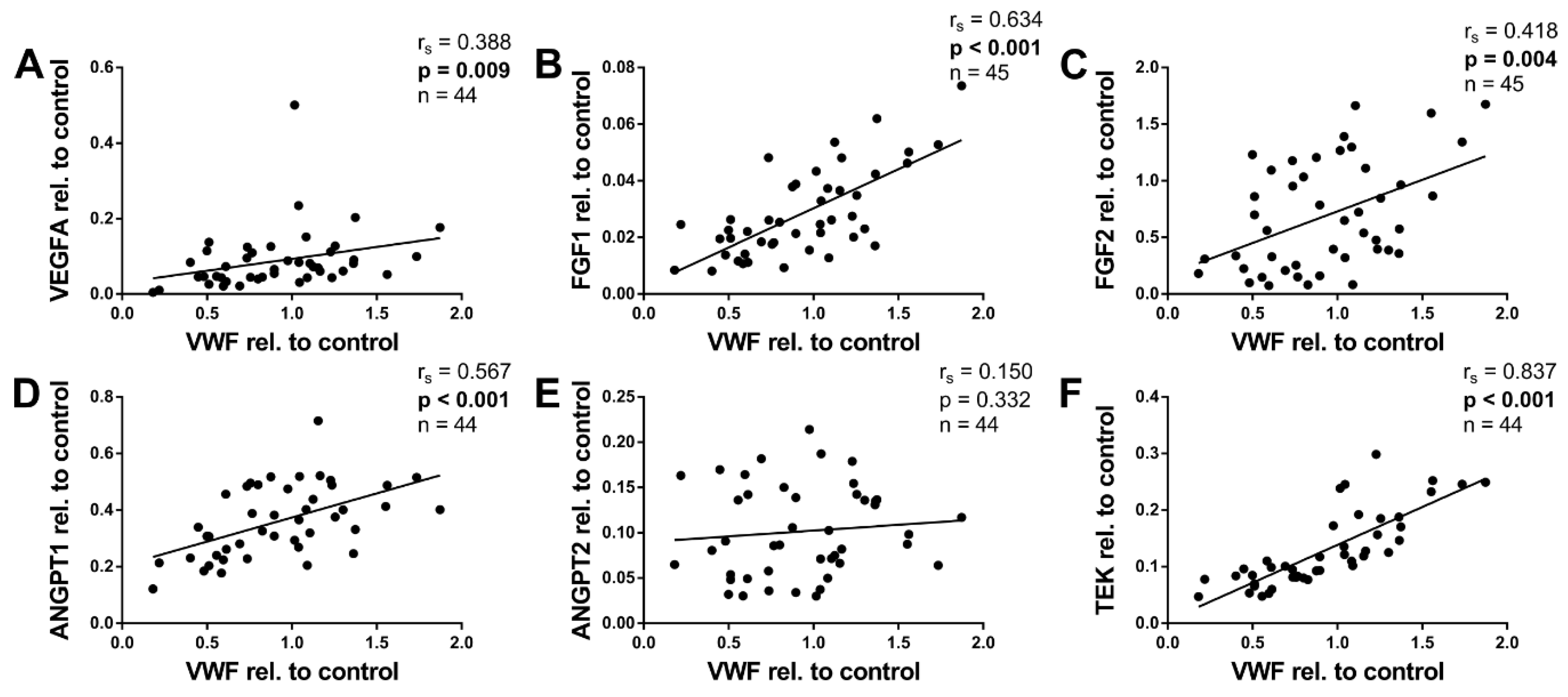
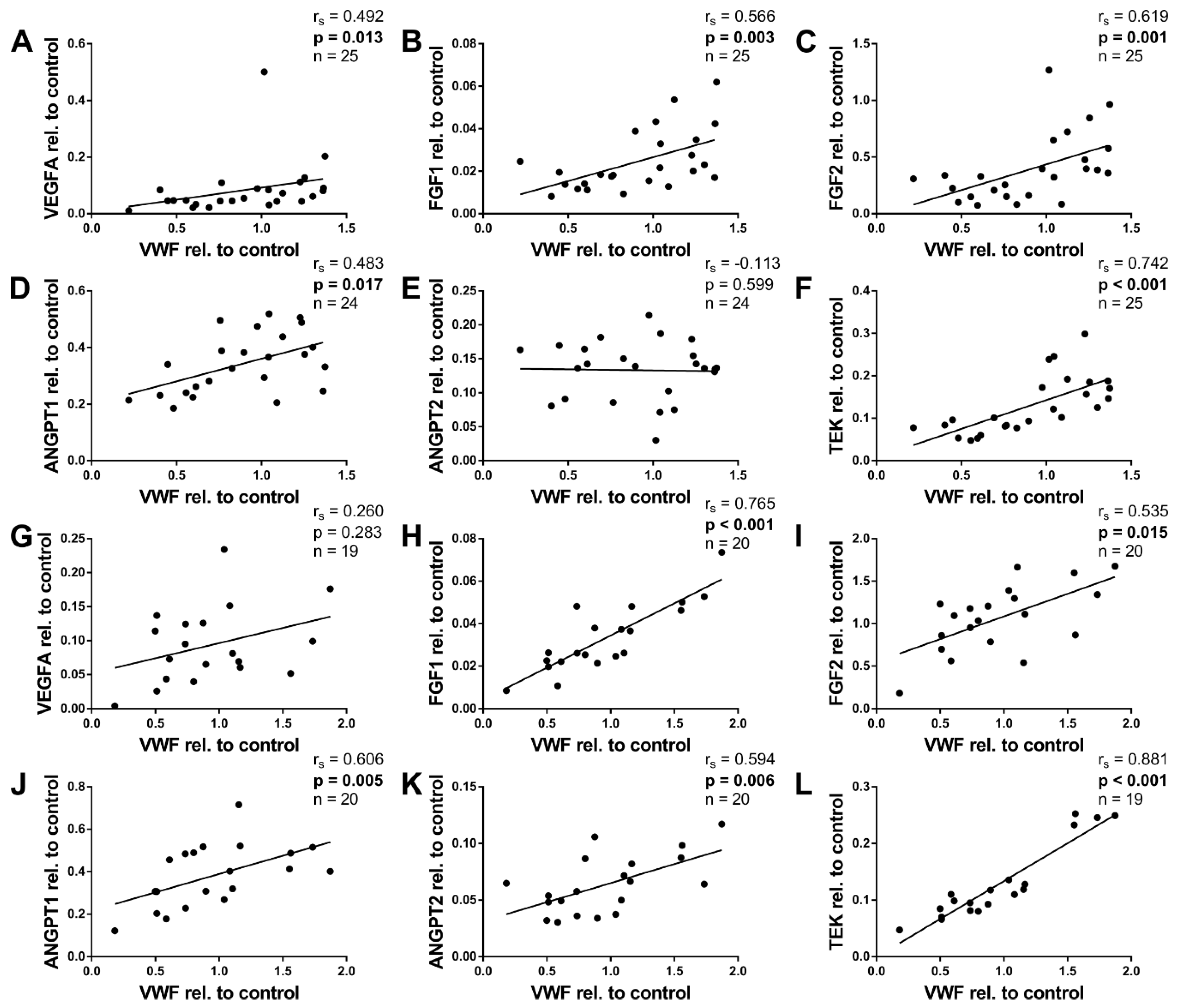
2.6. Angiogenic Factors and von Willebrand Factor
2.7. Angiogenic Factors in Infants <6 Months
3. Discussion
4. Materials and Methods
4.1. The Ulm Childhood Adipose Tissue Collective
4.2. Processing of the Adipose Tissue Samples
4.3. Histological Sample Preparation
4.4. Determination of the Adipocyte Size
4.5. RNA Isolation and cDNA Synthesis
4.6. Quantitative Real-Time Polymerase Chain Reaction
- 36B4 forw 5′-TGC ATC AGT ACC CCA TTC TAT CAT-3′;
- 36B4 rev 5′-AGG CAG ATG GAT CAG CCA AGA-3′;
- ANGPT1 forw 5′-TGA GAC CCA GGT ACT AAA TCA AAC TTC TCG AC-3′;
- ANGPT1 rev 5′-TGA AGA AGT TGC TTC TCT AGC TTG TAG GTG-3′;
- ANGPT2 forw 5′- ACA GCA GAA TGC AGT ACA GAA CCA GAC G -3′;
- ANGPT2 rev 5′-CAA GTC TCG TGG TCT GAT TTA ATA CTT GGG CT-3′;
- FGF1 forw 5′-GAC CAG CAC ATT CAG CTG CAG CTC AGT G-3′;
- FGF1 rev 5′-ACA AAC ATT CCT CAT TTG GTG TCT GTG AGC CG-3′;
- FGF2 forw 5′-CGA CCC TCA CAT CAA GCT ACA ACT TCA AGC AG-3′;
- FGF2 rev 5′-AGC CAG TAA TCT TCC ATC TTC CTT CAT AGC CA-3′;
- HPRT forw 5′-GAG ATG GGA GGC CAT CAC ATT GTA GCC CTC-3′;
- HPRT rev 5′-CTC CAC CAA TTA CTT TTA TGT CCC CTG TTG ACT GGT C-3′;
- PDGFRa forw 5′-GAA TAA CAT CGG AGG AGA AGT TTC CCA GAG-3′;
- PDGFRa rev 5′-CAT TTG GAA GGA TAG AGG GTA ATG AAA GCT GG-3′;
- TEK forw 5′-CTA GAA GTA CAC CTG CCT CAT GCT CAG CC-3′;
- TEK rev 5′-CAG TTC ACA AGC CTT CTC ACA CGT CCT TCC-3′;
- VEGFA forw 5′-CTT GCC TTG CTG CTC TAC CT-3′;
- VEGFA rev 5′-AGC TGC GCT GAT AGA CAT CC-3′;
- VWF forw 5′-GAC CAA AGA GTC TCC ATG CCC TAT GCC T-3′;
- VWF rev 5′-AGA TGC CCG TTC ACA CCA CTG TTC TCC A-3′.
4.7. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| 36B4 | Acidic ribosomal phosphoprotein P0 |
| ANGPT1 | Angiopoietin-1 |
| ANGPT2 | Angiopoietin-2 |
| BMI | Body mass index |
| FGFs | Fibroblast growth factors |
| H&E | Haematoxylin and eosin |
| HPRT | Hypoxanthine-guanine phosphoribosyltransferase |
| mRNA | Messenger ribonucleic acid |
| PDGFRa | Platelet-derived growth factor receptor alpha |
| qPCR | Quantitative polymerase chain reaction |
| RNA | Ribonucleic acid |
| SD | Standard deviation |
| SDS | Standard deviation score |
| SVCs | Stromal vascular cells |
| TEK (TIE-2) | Tunica interna endothelial cell kinase, an angiopoietin receptor |
| VEGFA | Vascular endothelial growth factor A |
| VWF | Von Willebrand factor |
| WAT | White adipose tissue |
| WHO | World Health Organization |
Appendix A





References
- WHO. Childhood Overweight and Obesity. Available online: https://www.who.int/dietphysicalactivity/childhood/en/ (accessed on 22 March 2019).
- Ward, Z.J.; Long, M.W.; Resch, S.C.; Giles, C.M.; Cradock, A.L.; Gortmaker, S.L. Simulation of Growth Trajectories of Childhood Obesity into Adulthood. N. Engl. J. Med. 2017, 377, 2145–2153. [Google Scholar] [CrossRef] [PubMed]
- Biro, F.M.; Wien, M. Childhood Obesity and Adult Morbidities. Am. J. Clin. Nutr. 2010, 91, 1499S–1505S. [Google Scholar] [CrossRef] [PubMed]
- Ghaben, A.L.; Scherer, P.E. Adipogenesis and Metabolic Health. Nat. Rev. Mol. Cell Biol. 2019, 20, 242–258. [Google Scholar] [CrossRef] [PubMed]
- Wassermann, F. The development of adipose tissue. In Handbook of Physiology. Section 5: Adipose Tissue; Renold, A.E., Cahill, G., Jr., Eds.; American Physiological Society: Washington, DC, USA, 1965; pp. 87–100. [Google Scholar]
- Poissonnet, C.M.; Burdi, A.R.; Bookstein, F.L. Growth and Development of Human Adipose Tissue during Early Gestation. Early Hum. Dev. 1983, 8, 1–11. [Google Scholar] [CrossRef]
- Fischer-Posovszky, P.; Roos, J.; Zoller, V.; Wabitsch, M. White Adipose Tissue Development and Function in Children and Adolescents: Preclinical Models. In Pediatric Obesity: Etiology, Pathogenesis and Treatment; Freemark, M.S., Ed.; Springer: Cham, Switzerland, 2018; pp. 81–93. [Google Scholar]
- Fomon, S.J.; Haschke, F.; Ziegler, E.E.; Nelson, S.E. Body Composition of Reference Children from Birth to Age 10 Years. Am. J. Clin. Nutr. 1982, 35, 1169–1175. [Google Scholar] [CrossRef] [PubMed]
- Butte, N.F.; Hopkinson, J.M.; Wong, W.W.; Smith, E.O.; Ellis, K.J. Body Composition during the First 2 Years of Life: An Updated Reference. Pediatr. Res. 2000, 47, 578–585. [Google Scholar] [CrossRef] [Green Version]
- Hager, A.; Sjostrm, L.; Arvidsson, B.; Bjorntorp, P.; Smith, U. Body Fat and Adipose Tissue Cellularity in Infants: A Longitudinal Study. Metabolism 1977, 26, 607–614. [Google Scholar] [CrossRef]
- Hausman, G.J.; Campion, D.R.; Martin, R.J. Search for the Adipocyte Precursor Cell and Factors that Promote its Differentiation. J. Lipid Res. 1980, 21, 657–670. [Google Scholar]
- Poissonnet, C.M.; Burdi, A.R.; Garn, S.M. The Chronology of Adipose Tissue Appearance and Distribution in the Human Fetus. Early Hum. Dev. 1984, 10, 1–11. [Google Scholar] [CrossRef]
- Tang, W.; Zeve, D.; Suh, J.M.; Bosnakovski, D.; Kyba, M.; Hammer, R.E.; Tallquist, M.D.; Graff, J.M. White Fat Progenitor Cells Reside in the Adipose Vasculature. Science 2008, 322, 583–586. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y. Angiogenesis Modulates Adipogenesis and Obesity. J. Clin. Investig. 2007, 117, 2362–2368. [Google Scholar] [CrossRef] [PubMed]
- Korner, A.; Kratzsch, J.; Gausche, R.; Schaab, M.; Erbs, S.; Kiess, W. New Predictors of the Metabolic Syndrome in Children—Role of Adipocytokines. Pediatr. Res. 2007, 61, 640–645. [Google Scholar] [CrossRef] [PubMed]
- Landgraf, K.; Rockstroh, D.; Wagner, I.V.; Weise, S.; Tauscher, R.; Schwartze, J.T.; Loffler, D.; Buhligen, U.; Wojan, M.; Till, H.; et al. Evidence of Early Alterations in Adipose Tissue Biology and Function and its Association with Obesity-Related Inflammation and Insulin Resistance in Children. Diabetes 2015, 64, 1249–1261. [Google Scholar] [CrossRef] [PubMed]
- Hughes, K.; Horwood, J.F.; Clements, C.; Leyland, D.; Corbett, H.J. Complications of Inguinal Herniotomy are Comparable in Term and Premature Infants. Hernia 2016, 20, 565–569. [Google Scholar] [CrossRef] [PubMed]
- Tam, C.S.; Tordjman, J.; Divoux, A.; Baur, L.A.; Clement, K. Adipose Tissue Remodeling in Children: The Link between Collagen Deposition and Age-Related Adipocyte Growth. J. Clin. Endocrinol. Metab. 2012, 97, 1320–1327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niedzielski, J.K.; Oszukowska, E.; Slowikowska-Hilczer, J. Undescended Testis—Current Trends and Guidelines: A Review of the Literature. Arch. Med. Sci. 2016, 12, 667–677. [Google Scholar] [CrossRef] [PubMed]
- Kromeyer-Hauschild, K.; Wabitsch, M.; Kunze, D.; Geller, F.; Geiß, H.C.; Hesse, V.; Von Hippel, A.; Jaeger, U.; Johnsen, D.; Korte, W.; et al. Perzentile Für Den Body-Mass-Index Für Das Kindes- Und Jugendalter Unter Heranziehung Verschiedener Deutscher Stichproben. Monatsschr Kinderheilkd. 2001, 149, 807–818. [Google Scholar] [CrossRef]
- Malina, R.M.; Bouchard, C.; Bar-Or, O. Adipose Tissue. In Growth, Maturation, and Physical Activity, 2nd ed.; Malina, R.M., Bouchard, C., Bar-Or, O., Eds.; Human Kinetics: Champaign, IL, USA, 2004; pp. 159–177. [Google Scholar]
- Hepler, C.; Vishvanath, L.; Gupta, R.K. Sorting Out Adipocyte Precursors and their Role in Physiology and Disease. Genes Dev. 2017, 31, 127–140. [Google Scholar] [CrossRef]
- Wabitsch, M.; Fischer-Posovsky, P. Entwicklung und Regulation des Fettgewebes. In Adipositas Bei Kindern Und Jugendlichen; Wabitsch, M., Ed.; Springer: Berlin, Heidelberg, Germany, 2005; pp. 73–80. [Google Scholar]
- Adya, R.; Homer-Vanniasinkam, S. Adipokines and Adipose Tissue Angiogenesis in Obesity. Immunoendocrinology 2015, 2, e918. [Google Scholar]
- Christiaens, V.; Lijnen, H.R. Angiogenesis and Development of Adipose Tissue. Mol. Cell. Endocrinol. 2010, 318, 2–9. [Google Scholar] [CrossRef]
- Lenting, P.J.; Christophe, O.D.; Denis, C.V. Von Willebrand Factor Biosynthesis, Secretion, and Clearance: Connecting the Far Ends. Blood 2015, 125, 2019–2028. [Google Scholar] [CrossRef] [PubMed]
- Randi, A.M.; Laffan, M.A. Von Willebrand Factor and Angiogenesis: Basic and Applied Issues. J. Thromb. Haemost. 2017, 15, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Thomas, M.; Augustin, H.G. The Role of the Angiopoietins in Vascular Morphogenesis. Angiogenesis 2009, 12, 125–137. [Google Scholar] [CrossRef] [PubMed]
- Knittle, J.L.; Timmers, K.; Ginsberg-Fellner, F.; Brown, R.E.; Katz, D.P. The Growth of Adipose Tissue in Children and Adolescents. Cross-Sectional and Longitudinal Studies of Adipose Cell Number and Size. J. Clin. Investig. 1979, 63, 239–246. [Google Scholar] [CrossRef] [PubMed]
- Crandall, D.L.; Hausman, G.J.; Kral, J.G. A Review of the Microcirculation of Adipose Tissue: Anatomic, Metabolic, and Angiogenic Perspectives. Microcirculation 1997, 4, 211–232. [Google Scholar] [CrossRef] [PubMed]
- Hausman, G.J.; Kauffman, R.G. The Histology of Developing Porcine Adipose Tissue. J. Anim. Sci. 1986, 63, 642–658. [Google Scholar] [CrossRef]
- Zhang, J.F.; Yao, G.Y.; Wu, Y.H. Expression Profiling Based on Coexpressed Modules in Obese Prepubertal Children. Genet. Mol. Res. 2012, 11, 3077–3085. [Google Scholar] [CrossRef]
- Li, L.; Wang, G.; Li, N.; Yu, H.; Si, J.; Wang, J. Identification of Key Genes and Pathways Associated with Obesity in Children. Exp. Ther. Med. 2017, 14, 1065–1073. [Google Scholar] [CrossRef]
- Garcia, A.G.; Nunez, G.G.; Sandoval, M.E.V.; Castellanos, S.G.; Aguilar, C.A. Factors Associated with Early Platelet Activation in Obese Children. Clin. Med. Res. 2014, 12, 21–26. [Google Scholar] [CrossRef] [Green Version]
- Singh, A.; Foster, G.D.; Gunawardana, J.; McCoy, T.A.; Nguyen, T.; Veur, S.V.; Komaroff, E.; Rao, A.K. Elevated Circulating Tissue Factor Procoagulant Activity, Factor VII, and Plasminogen Activator Inhibitor-1 in Childhood Obesity: Evidence of a Procoagulant State. Br. J. Haematol. 2012, 158, 523–527. [Google Scholar] [CrossRef]
- Breier, G.; Clauss, M.; Risau, W. Coordinate Expression of Vascular Endothelial Growth Factor Receptor-1 (Flt-1) and its Ligand Suggests a Paracrine Regulation of Murine Vascular Development. Dev. Dyn. 1995, 204, 228–239. [Google Scholar] [CrossRef] [PubMed]
- Ferrara, N.; Henzel, W.J. Pituitary Follicular Cells Secrete a Novel Heparin-Binding Growth Factor Specific for Vascular Endothelial Cells. Biochem. Biophys. Res. Commun. 1989, 161, 851–858. [Google Scholar] [CrossRef]
- Beck, B.; Driessens, G.; Goossens, S.; Youssef, K.K.; Kuchnio, A.; Caauwe, A.; Sotiropoulou, P.A.; Loges, S.; Lapouge, G.; Candi, A.; et al. A Vascular Niche and a VEGF-Nrp1 Loop Regulate the Initiation and Stemness of Skin Tumours. Nature 2011, 478, 399–403. [Google Scholar] [CrossRef] [PubMed]
- Ferrara, N.; Carver-Moore, K.; Chen, H.; Dowd, M.; Lu, L.; O’Shea, K.S.; Powell-Braxton, L.; Hillan, K.J.; Moore, M.W. Heterozygous Embryonic Lethality Induced by Targeted Inactivation of the VEGF Gene. Nature 1996, 380, 439–442. [Google Scholar] [CrossRef]
- Carmeliet, P.; Ferreira, V.; Breier, G.; Pollefeyt, S.; Kieckens, L.; Gertsenstein, M.; Fahrig, M.; Vandenhoeck, A.; Harpal, K.; Eberhardt, C.; et al. Abnormal Blood Vessel Development and Lethality in Embryos Lacking a Single VEGF Allele. Nature 1996, 380, 435–439. [Google Scholar] [CrossRef]
- Tinahones, F.J.; Coin-Araguez, L.; Mayas, M.D.; Garcia-Fuentes, E.; Hurtado-Del-Pozo, C.; Vendrell, J.; Cardona, F.; Calvo, R.M.; Obregon, M.J.; El Bekay, R. Obesity-Associated Insulin Resistance is Correlated to Adipose Tissue Vascular Endothelial Growth Factors and Metalloproteinase Levels. BMC Physiol. 2012, 12, 4. [Google Scholar] [CrossRef]
- Yamada, T.; Kawakami, S.; Nakanishi, N. Fat Depot-Specific Differences in Angiogenic Growth Factor Gene Expression and its Relation to Adipocyte Size in Cattle. J. Vet. Med. Sci. 2010, 72, 991–997. [Google Scholar] [CrossRef]
- Beenken, A.; Mohammadi, M. The FGF Family: Biology, Pathophysiology and Therapy. Nat. Rev. Drug Discov. 2009, 8, 235–253. [Google Scholar] [CrossRef]
- Maciag, T.; Mehlman, T.; Friesel, R.; Schreiber, A.B. Heparin Binds Endothelial Cell Growth Factor, the Principal Endothelial Cell Mitogen in Bovine Brain. Science 1984, 225, 932–935. [Google Scholar] [CrossRef]
- Shing, Y.; Folkman, J.; Sullivan, R.; Butterfield, C.; Murray, J.; Klagsbrun, M. Heparin Affinity: Purification of a Tumor-Derived Capillary Endothelial Cell Growth Factor. Science 1984, 223, 1296–1299. [Google Scholar] [CrossRef]
- Yu, P.; Wilhelm, K.; Dubrac, A.; Tung, J.K.; Alves, T.C.; Fang, J.S.; Xie, Y.; Zhu, J.; Chen, Z.; De Smet, F.; et al. FGF-Dependent Metabolic Control of Vascular Development. Nature 2017, 545, 224–228. [Google Scholar] [CrossRef] [PubMed]
- Hutley, L.; Shurety, W.; Newell, F.; McGeary, R.; Pelton, N.; Grant, J.; Herington, A.; Cameron, D.; Whitehead, J.; Prins, J. Fibroblast Growth Factor 1: A Key Regulator of Human Adipogenesis. Diabetes 2004, 53, 3097–3106. [Google Scholar] [CrossRef] [PubMed]
- Jonker, J.W.; Suh, J.M.; Atkins, A.R.; Ahmadian, M.; Li, P.; Whyte, J.; He, M.; Juguilon, H.; Yin, Y.Q.; Phillips, C.T.; et al. A PPARgamma-FGF1 Axis is Required for Adaptive Adipose Remodelling and Metabolic Homeostasis. Nature 2012, 485, 391–394. [Google Scholar] [CrossRef] [PubMed]
- Kakudo, N.; Shimotsuma, A.; Kusumoto, K. Fibroblast Growth Factor-2 Stimulates Adipogenic Differentiation of Human Adipose-Derived Stem Cells. Biochem. Biophys. Res. Commun. 2007, 359, 239–244. [Google Scholar] [CrossRef]
- Kim, S.; Ahn, C.; Bong, N.; Choe, S.; Lee, D.K. Biphasic Effects of FGF2 on Adipogenesis. PLoS ONE 2015, 10, e0120073. [Google Scholar] [CrossRef]
- Maisonpierre, P.C.; Suri, C.; Jones, P.F.; Bartunkova, S.; Wiegand, S.J.; Radziejewski, C.; Compton, D.; McClain, J.; Aldrich, T.H.; Papadopoulos, N.; et al. Angiopoietin-2, a Natural Antagonist for Tie2 that Disrupts in Vivo Angiogenesis. Science 1997, 277, 55–60. [Google Scholar] [CrossRef]
- Kim, I.; Kim, J.H.; Moon, S.O.; Kwak, H.J.; Kim, N.G.; Koh, G.Y. Angiopoietin-2 at High Concentration can Enhance Endothelial Cell Survival through the Phosphatidylinositol 3’-kinase/Akt Signal Transduction Pathway. Oncogene 2000, 19, 4549–4552. [Google Scholar] [CrossRef]
- Kim, M.; Allen, B.; Korhonen, E.A.; Nitschke, M.; Yang, H.W.; Baluk, P.; Saharinen, P.; Alitalo, K.; Daly, C.; Thurston, G.; et al. Opposing Actions of Angiopoietin-2 on Tie2 Signaling and FOXO1 Activation. J. Clin. Investig. 2016, 126, 3511–3525. [Google Scholar] [CrossRef]
- Yuan, H.T.; Khankin, E.V.; Karumanchi, S.A.; Parikh, S.M. Angiopoietin 2 is a Partial agonist/antagonist of Tie2 Signaling in the Endothelium. Mol. Cell. Biol. 2009, 29, 2011–2022. [Google Scholar] [CrossRef]
- Holash, J.; Maisonpierre, P.C.; Compton, D.; Boland, P.; Alexander, C.R.; Zagzag, D.; Yancopoulos, G.D.; Wiegand, S.J. Vessel Cooption, Regression, and Growth in Tumors Mediated by Angiopoietins and VEGF. Science 1999, 284, 1994–1998. [Google Scholar] [CrossRef] [Green Version]
- Lobov, I.B.; Brooks, P.C.; Lang, R.A. Angiopoietin-2 Displays VEGF-Dependent Modulation of Capillary Structure and Endothelial Cell Survival in Vivo. Proc. Natl. Acad. Sci. USA 2002, 99, 11205–11210. [Google Scholar] [CrossRef] [PubMed]
- Hakanpaa, L.; Sipila, T.; Leppanen, V.M.; Gautam, P.; Nurmi, H.; Jacquemet, G.; Eklund, L.; Ivaska, J.; Alitalo, K.; Saharinen, P. Endothelial Destabilization by Angiopoietin-2 Via Integrin beta1 Activation. Nat. Commun. 2015, 6, 5962. [Google Scholar] [CrossRef] [PubMed]
- Scharpfenecker, M.; Fiedler, U.; Reiss, Y.; Augustin, H.G. The Tie-2 Ligand Angiopoietin-2 Destabilizes Quiescent Endothelium through an Internal Autocrine Loop Mechanism. J. Cell Sci. 2005, 118, 771–780. [Google Scholar] [CrossRef] [PubMed]
- Hammes, H.P.; Lin, J.; Wagner, P.; Feng, Y.; Vom Hagen, F.; Krzizok, T.; Renner, O.; Breier, G.; Brownlee, M.; Deutsch, U. Angiopoietin-2 Causes Pericyte Dropout in the Normal Retina: Evidence for Involvement in Diabetic Retinopathy. Diabetes 2004, 53, 1104–1110. [Google Scholar] [CrossRef]
- Park, S.W.; Yun, J.H.; Kim, J.H.; Kim, K.W.; Cho, C.H.; Kim, J.H. Angiopoietin 2 Induces Pericyte Apoptosis Via alpha3beta1 Integrin Signaling in Diabetic Retinopathy. Diabetes 2014, 63, 3057–3068. [Google Scholar] [CrossRef]
- Fiedler, U.; Scharpfenecker, M.; Koidl, S.; Hegen, A.; Grunow, V.; Schmidt, J.M.; Kriz, W.; Thurston, G.; Augustin, H.G. The Tie-2 Ligand Angiopoietin-2 is Stored in and Rapidly Released upon Stimulation from Endothelial Cell Weibel-Palade Bodies. Blood 2004, 103, 4150–4156. [Google Scholar] [CrossRef]
- Cho, C.H.; Kim, K.E.; Byun, J.; Jang, H.S.; Kim, D.K.; Baluk, P.; Baffert, F.; Lee, G.M.; Mochizuki, N.; Kim, J.; et al. Long-Term and Sustained COMP-Ang1 Induces Long-Lasting Vascular Enlargement and Enhanced Blood Flow. Circ. Res. 2005, 97, 86–94. [Google Scholar] [CrossRef] [Green Version]
- Pasarica, M.; Sereda, O.R.; Redman, L.M.; Albarado, D.C.; Hymel, D.T.; Roan, L.E.; Rood, J.C.; Burk, D.H.; Smith, S.R. Reduced Adipose Tissue Oxygenation in Human Obesity: Evidence for Rarefaction, Macrophage Chemotaxis, and Inflammation without an Angiogenic Response. Diabetes 2009, 58, 718–725. [Google Scholar] [CrossRef]
- Dallabrida, S.M.; Zurakowski, D.; Shih, S.C.; Smith, L.E.; Folkman, J.; Moulton, K.S.; Rupnick, M.A. Adipose Tissue Growth and Regression are Regulated by Angiopoietin-1. Biochem. Biophys. Res. Commun. 2003, 311, 563–571. [Google Scholar] [CrossRef]
- Halberg, N.; Khan, T.; Trujillo, M.E.; Wernstedt-Asterholm, I.; Attie, A.D.; Sherwani, S.; Wang, Z.V.; Landskroner-Eiger, S.; Dineen, S.; Magalang, U.J.; et al. Hypoxia-Inducible Factor 1alpha Induces Fibrosis and Insulin Resistance in White Adipose Tissue. Mol. Cell. Biol. 2009, 29, 4467–4483. [Google Scholar] [CrossRef]
- Voros, G.; Maquoi, E.; Demeulemeester, D.; Clerx, N.; Collen, D.; Lijnen, H.R. Modulation of Angiogenesis during Adipose Tissue Development in Murine Models of Obesity. Endocrinology 2005, 146, 4545–4554. [Google Scholar] [CrossRef] [PubMed]
- Adya, R.; Tan, B.K.; Randeva, H.S. Differential Effects of Leptin and Adiponectin in Endothelial Angiogenesis. J. Diabetes Res. 2015, 2015, 648239. [Google Scholar] [CrossRef] [PubMed]
- Engle, W.A.; Blackmon, L.R.; Batton, D.G.; Bell, E.F.; Denson, S.E.; Kanto, W.P.; Martin, G.I.; Stark, A. Age Terminology during the Perinatal Period. Pediatrics 2004, 114, 1362–1364. [Google Scholar] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | n | Mean ± SD | Median | Range |
|---|---|---|---|---|
| Gender | 45 | 36 male, 9 female | ||
| Corrected age [y] | 45 | 1.51 ± 2.18 | 0.24 | −0.04–9.23 |
| Weight [kg] | 45 | 8.94 ± 6.30 | 5.62 | 2.07–25 |
| Height [cm] | 45 | 72.10 ± 24.83 | 61 | 42–131 |
| BMI [kg·m−2] | 45 | 15.16 ± 1.96 | 15.43 | 10.63–18.28 |
| BMI-SDS | 45 | 0.09 ± 1.08 | 0.06 | −1.74–2.61 |
| Adipocyte diameter [µm] | 35 | 56.94 ± 10.63 | 56.66 | 39.73–88.89 |
| Gender | 35 | 28 male, 7 female | ||
| Age | BMI-SDS | Adipocyte Diameter | VWF | ||||||
|---|---|---|---|---|---|---|---|---|---|
| rs | p | rs | p | rs | p | rs | p | ||
| VEGFA | all | 0,262 | 0.086 | −0.106 | 0.491 | 0.176 | 0.320 | 0.388 | 0.009 |
| <6 m | 0.321 | 0.118 | 0.091 | 0.666 | 0.021 | 0.930 | 0.492 | 0.013 | |
| ≥6 m | –0.270 | 0.263 | −0.282 | 0.241 | 0.218 | 0.454 | 0.260 | 0.283 | |
| FGF1 | all | 0.437 | 0.003 | 0.157 | 0.303 | 0.525 | 0.001 | 0.634 | <0.001 |
| <6 m | 0.687 | <0.001 | 0.296 | 0.151 | 0.547 | 0.012 | 0.566 | 0.003 | |
| ≥6 m | −0.189 | 0.424 | 0.078 | 0.743 | 0.307 | 0.265 | 0.765 | <0.001 | |
| FGF2 | all | 0.750 | <0.001 | 0.017 | 0.913 | 0.429 | 0.010 | 0.418 | 0.004 |
| <6 m | 0.665 | <0.001 | 0.259 | 0.211 | 0.257 | 0.274 | 0.619 | 0.001 | |
| ≥6 m | −0.134 | 0.574 | 0.080 | 0.738 | −0.050 | 0.863 | 0.535 | 0.015 | |
| ANGPT1 | all | 0.213 | 0.166 | 0.426 | 0.004 | 0.519 | 0.002 | 0.567 | <0.001 |
| <6 m | 0.353 | 0.090 | 0.494 | 0.014 | 0.226 | 0.352 | 0.483 | 0.017 | |
| ≥6 m | 0.015 | 0.950 | 0.373 | 0.105 | 0.536 | 0.042 | 0.606 | 0.005 | |
| ANGPT2 | all | −0.650 | <0.001 | 0.148 | 0.337 | −0.297 | 0.083 | 0.150 | 0.332 |
| <6 m | −0.038 | 0.861 | 0.067 | 0.756 | 0.003 | 0.990 | −0.113 | 0.599 | |
| ≥6 m | −0.325 | 0.162 | 0.429 | 0.059 | 0.214 | 0.442 | 0.594 | 0.006 | |
| TEK | all | 0.194 | 0.206 | 0.358 | 0.017 | 0.102 | 0.565 | 0.837 | <0.001 |
| <6 m | 0.572 | 0.003 | 0.480 | 0.015 | 0.281 | 0.230 | 0.742 | <0.001 | |
| ≥6 m | −0.007 | 0.977 | 0.100 | 0.684 | −0.064 | 0.832 | 0.881 | <0.001 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gaebler, N.; Haggenmüller, B.; Kapapa, M.; Serra, A.; Tews, D.; Funcke, J.-B.; Brandt, S.; Ioannidis, V.; Schön, M.; Möller, P.; et al. Age- and BMI-Associated Expression of Angiogenic Factors in White Adipose Tissue of Children. Int. J. Mol. Sci. 2019, 20, 5204. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20205204
Gaebler N, Haggenmüller B, Kapapa M, Serra A, Tews D, Funcke J-B, Brandt S, Ioannidis V, Schön M, Möller P, et al. Age- and BMI-Associated Expression of Angiogenic Factors in White Adipose Tissue of Children. International Journal of Molecular Sciences. 2019; 20(20):5204. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20205204
Chicago/Turabian StyleGaebler, Niklas, Benedikt Haggenmüller, Melanie Kapapa, Alexandre Serra, Daniel Tews, Jan-Bernd Funcke, Stephanie Brandt, Valentin Ioannidis, Michael Schön, Peter Möller, and et al. 2019. "Age- and BMI-Associated Expression of Angiogenic Factors in White Adipose Tissue of Children" International Journal of Molecular Sciences 20, no. 20: 5204. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20205204