Protective Effects of Flavone from Tamarix aphylla against CCl4-Induced Liver Injury in Mice Mediated by Suppression of Oxidative Stress, Apoptosis and Angiogenesis



Abstract
:1. Introduction
2. Results
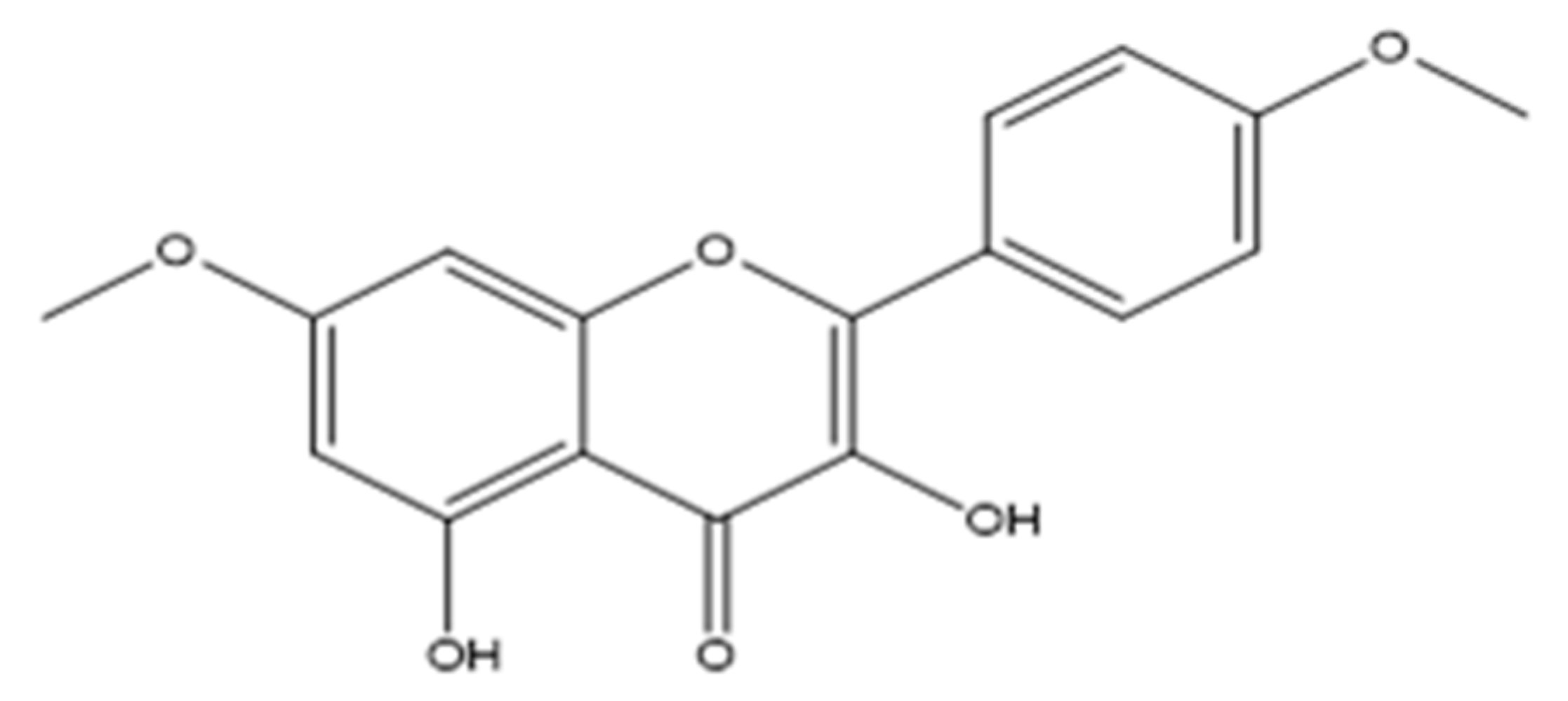
2.1. Chemical Elucidation of the Flavone
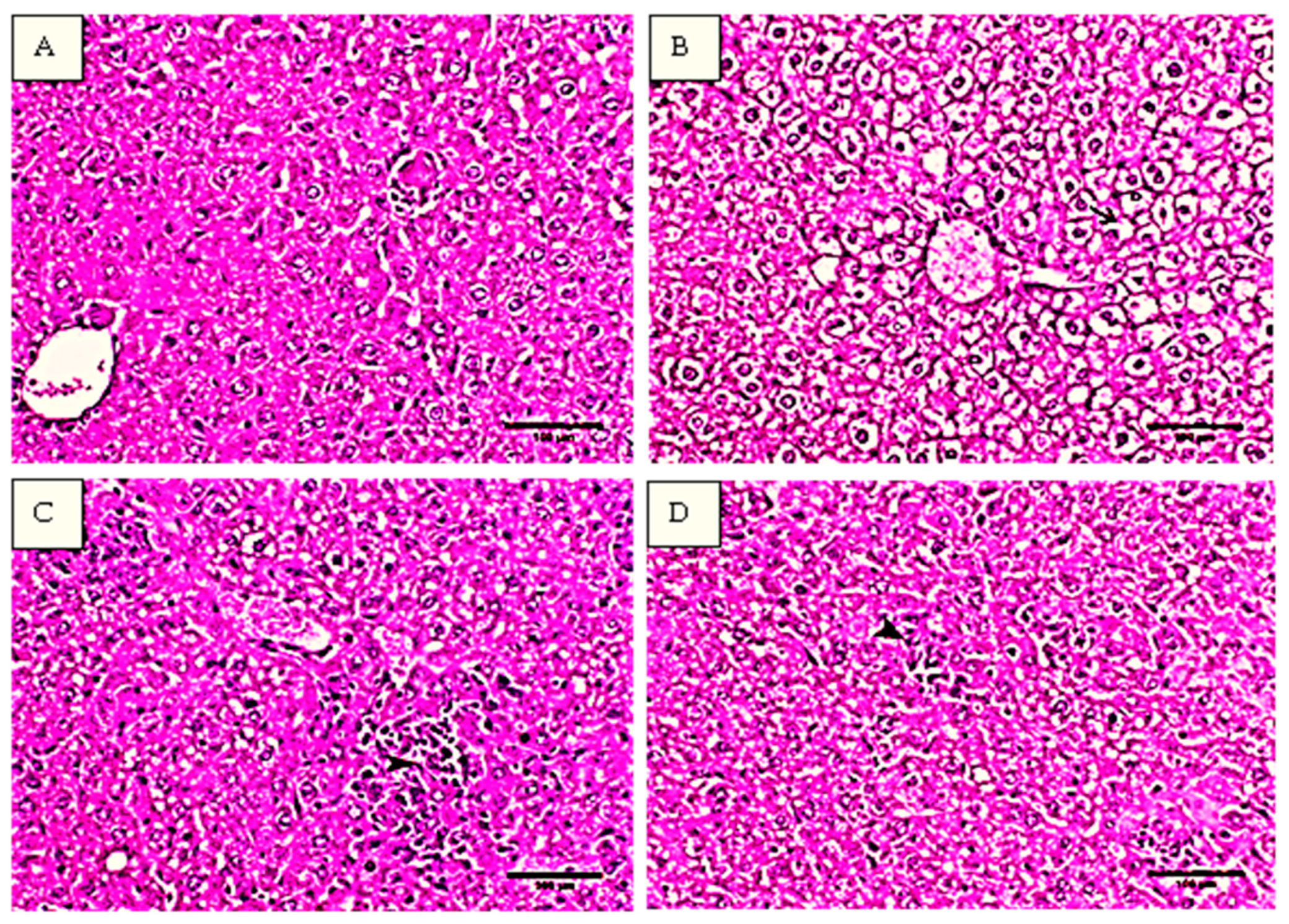
2.2. Histopathological Analysis of the Liver Tissues
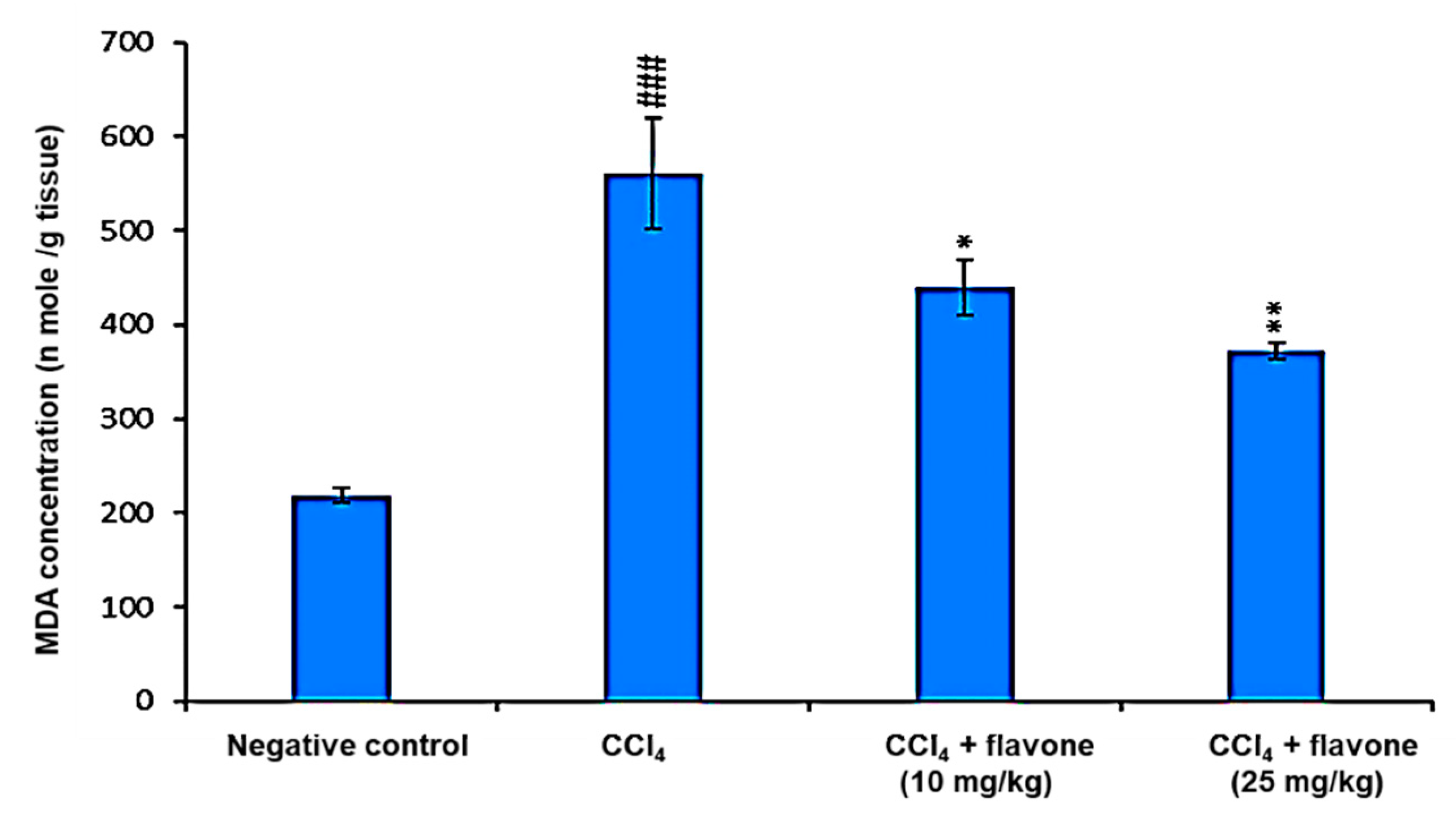
2.3. Markers of Oxidative Stress in Liver Tissue Homogenates
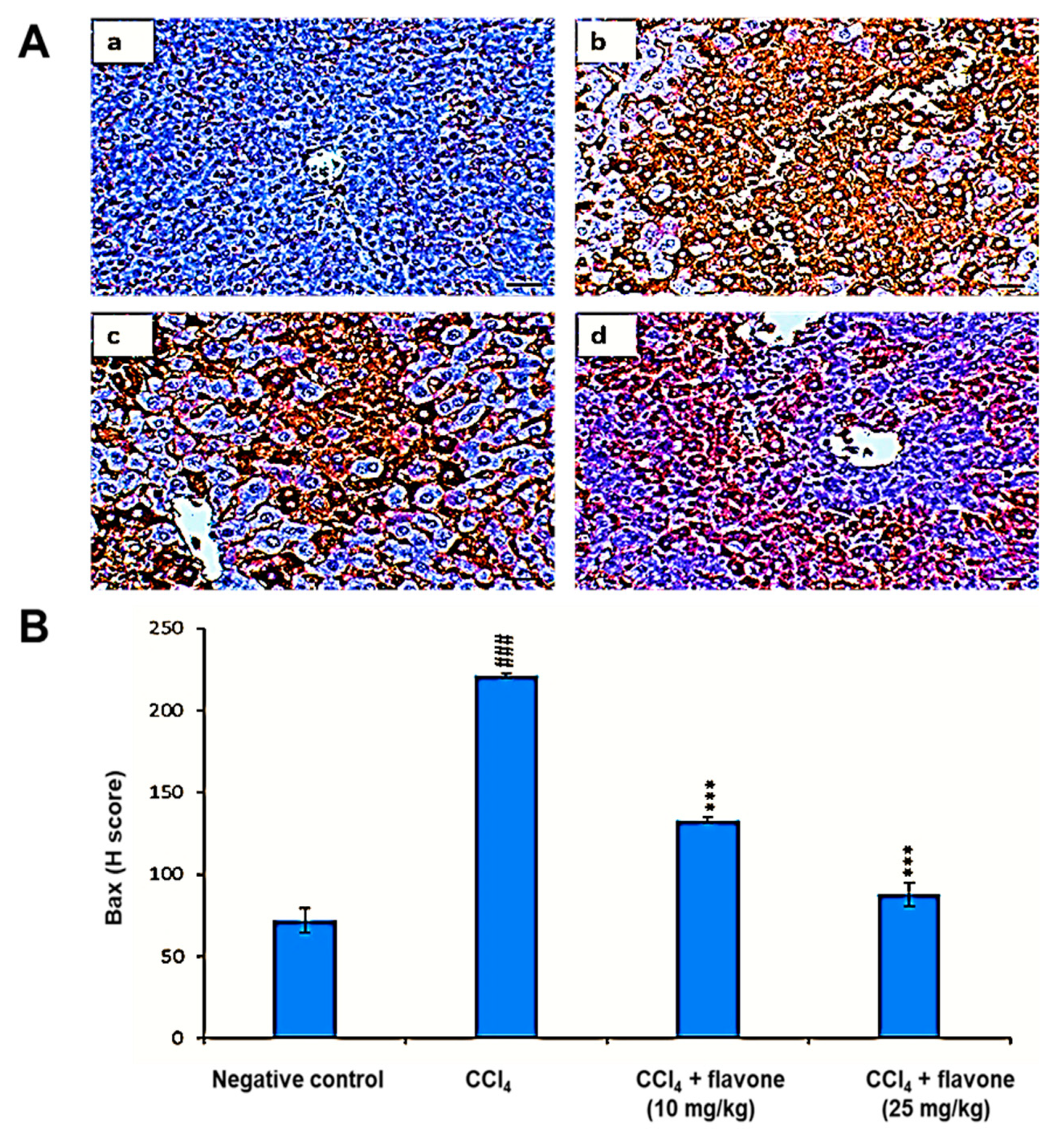
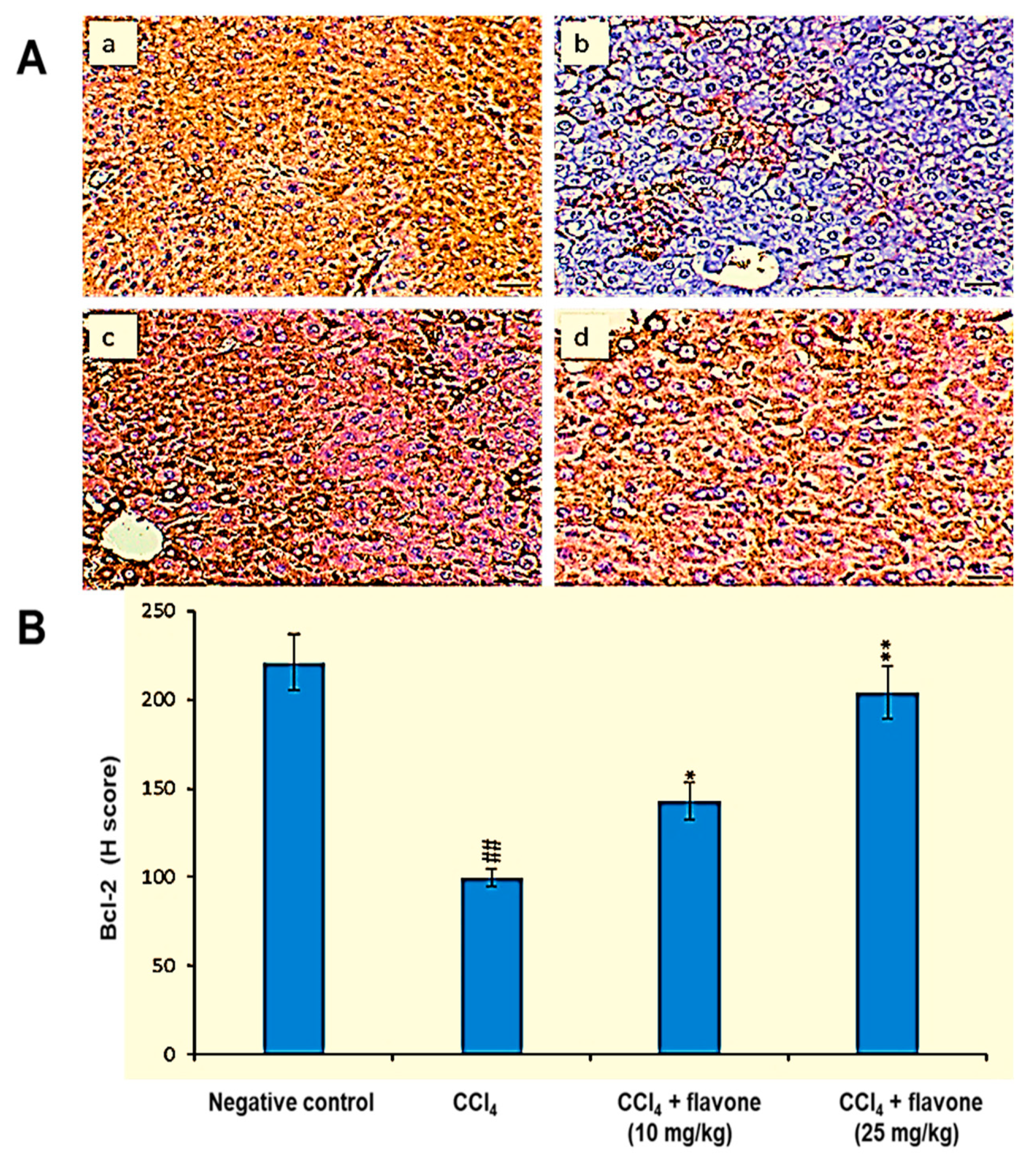
2.4. Apoptosis Regulators in Liver Tissue
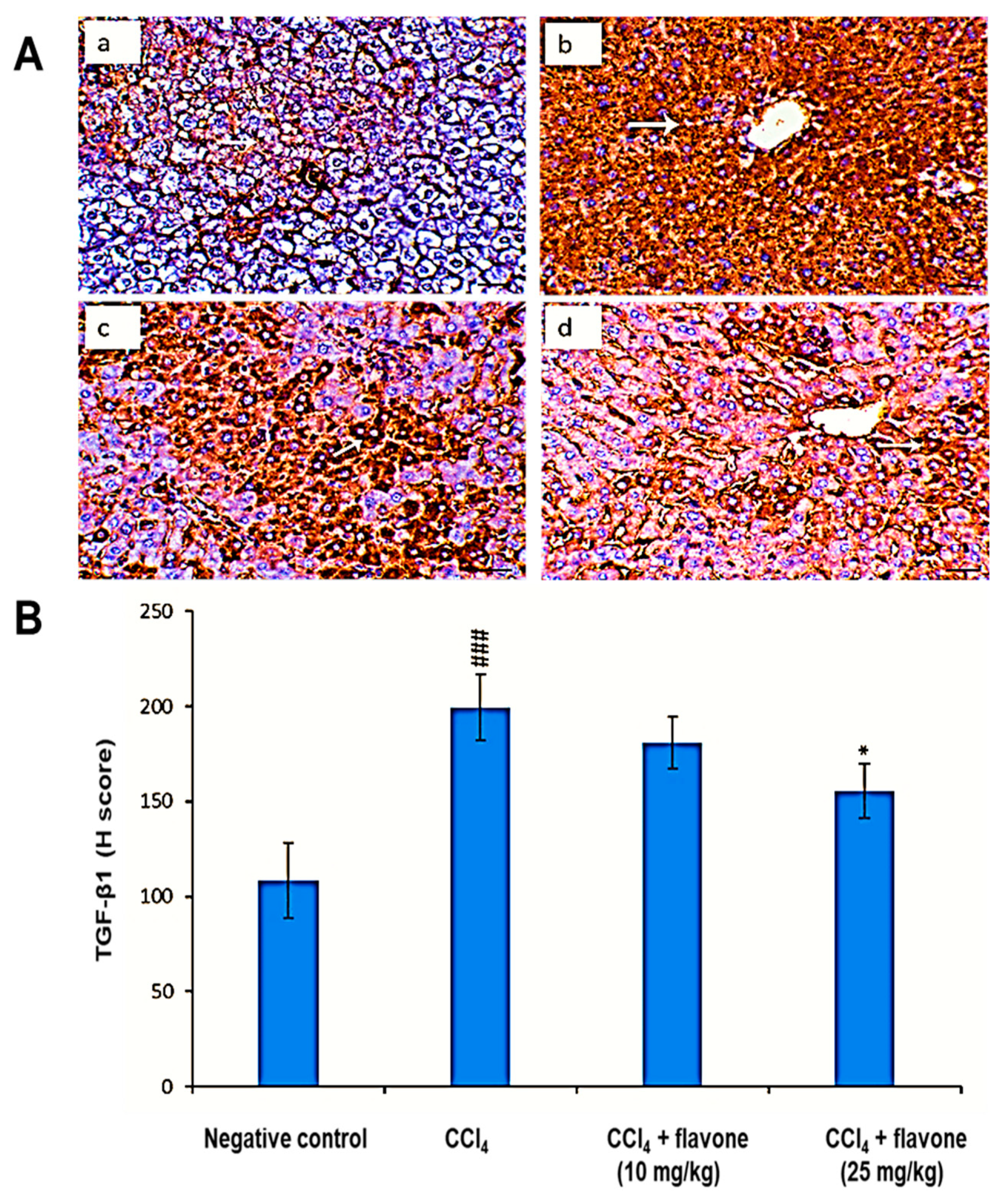
2.5. TGF-β1 Protein Levels in the Liver Tissue
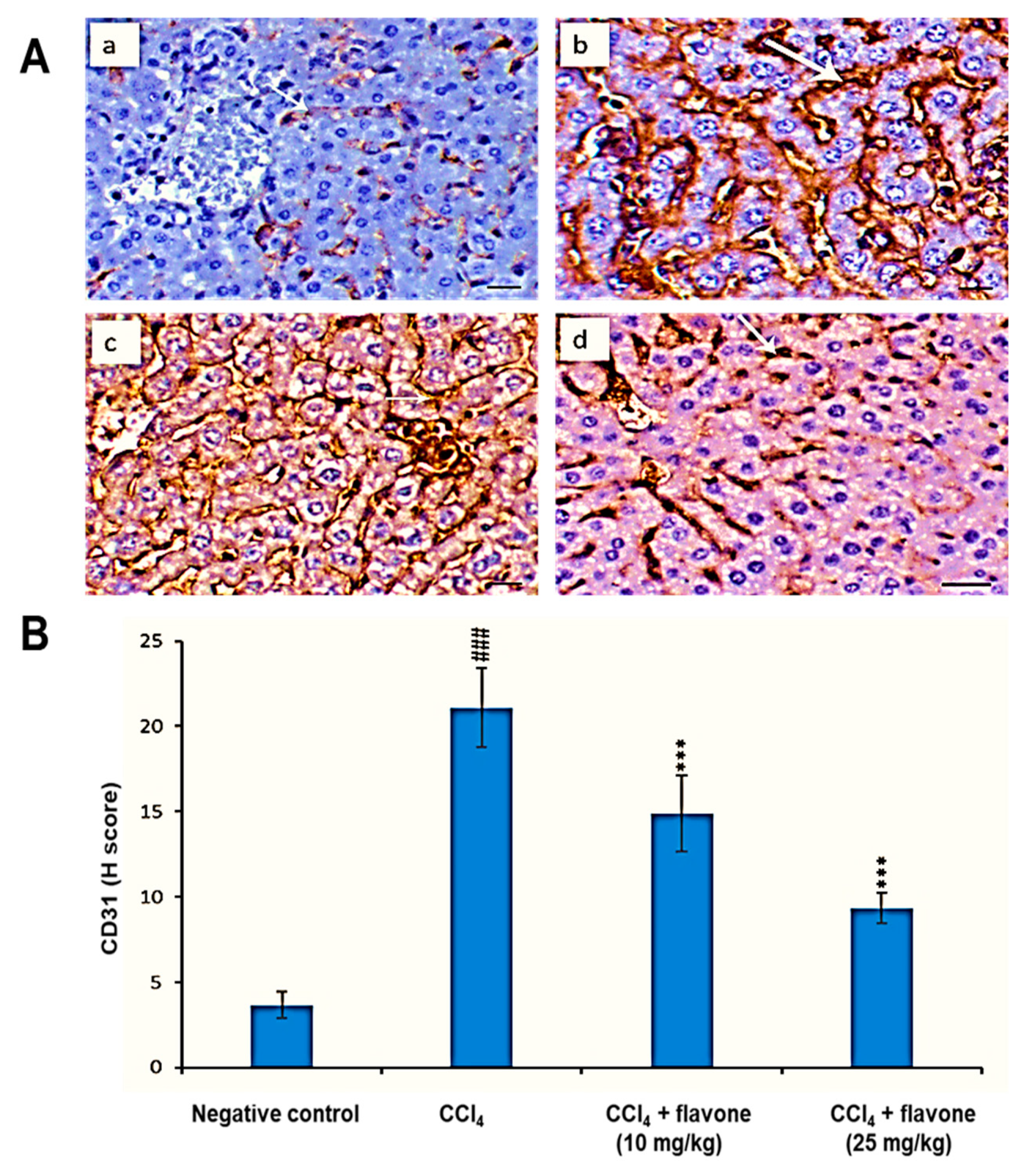
2.6. CD31 Protein Levels in the Liver Tissue
3. Discussion
4. Materials and Methods
4.1. Extraction, Isolation and Elucidation of Flavone
4.2. Animals
4.3. Liver Injury Induction and Experimental Design
4.4. Histological Examination
4.5. Estimation of Malondialdehyde (MDA) Level in Liver Homogenates
4.6. Determination of Anti-Oxidant Enzymes Levels in Liver Homogenates
4.7. Immunohistochemical Examination of Apoptosis and Angiogenesis-Related Protein Markers
4.8. Statistical Analysis
5. Conclusions, Limitations and Future Research
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lu, Y.; Hu, D.; Ma, S.; Zhao, X.; Wang, S.; Wei, G.; Wang, J. Protective effect of wedelolactone against CCl4-induced acute liver injury in mice. Int. Immunopharmacol. 2016, 34, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.Y.; Zhang, X.Y.; Guan, S.W.; Hua, Z.C. Protective effect of procyanidin B2 against CCl4-induced acute liver injury in mice. Molecules 2015, 20, 12250–12265. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.H.; Park, S.W.; Kim, Y.S.; Kang, S.S.; Kim, J.A.; Lee, S.H.; Lee, S.M. Protective mechanism of glycyrrhizin on acute liver injury induced by carbon tetrachloride in mice. Biol. Pharm. Bull. 2007, 30, 1898–1904. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.M. Drug-induced hepatotoxicity. N. Engl. J. Med. 2003, 349, 474–485. [Google Scholar] [CrossRef]
- Das, P.; Holt, S. Liver disease and renal dysfunction. Medicine 2011, 39, 492–496. [Google Scholar] [CrossRef]
- Weber, L.W.; Boll, M.; Stampfl, A. Hepatotoxicity and mechanism of action of haloalkanes: Carbon tetrachloride as a toxicological model. Crit. Rev. Toxicol. 2003, 33, 105–136. [Google Scholar] [CrossRef]
- Ingawale, D.K.; Mandlik, S.K.; Naik, S.R. Models of hepatotoxicity and the underlying cellular, biochemical and immunological mechanism(s): A critical discussion. Environ. Toxicol. Pharmacol. 2014, 37, 118–133. [Google Scholar] [CrossRef]
- Lettéron, P.; Labbe, G.; Degott, C.; Berson, A.; Fromenty, B.; Delaforge, M.; Larrey, D.; Pessayre, D. Mechanism for the protective effects of silymarin against carbon tetrachloride-induced lipid peroxidation and hepatotoxicity in mice. Evidence that silymarin acts both as an inhibitor of metabolic activation and as a chain-breaking antioxidant. Biochem. Pharmacol. 1990, 39, 2027–2034. [Google Scholar] [CrossRef]
- Amirshahrokhi, K.; Bohlooli, S. Effect of Methylsulfonylmethane on Paraquat-Induced Acute Lung and Liver Injury in Mice. Inflammation 2013, 36, 1111–1121. [Google Scholar] [CrossRef]
- Amirshahrokhi, K.; Bohlooli, S.; Chinifroush, M.M. The effect of methylsulfonylmethane on the experimental colitis in the rat. Toxicol. Appl. Pharmacol. 2011, 253, 197–202. [Google Scholar] [CrossRef] [Green Version]
- Lei, X.G.; Cheng, W.H.; McClung, J.P. Metabolic regulation and function of glutathione peroxidase-1. Annu. Rev. Nutr. 2007, 27, 41–61. [Google Scholar] [CrossRef] [PubMed]
- Recknagel, R.O.; Glende, E.A., Jr.; Dolak, J.A.; Waller, R.L. Mechanisms of carbon tetrachloride toxicity. Pharmacol. Ther. 1989, 43, 139–154. [Google Scholar] [CrossRef]
- Chen, Q.; Zhan, Q.; Li, Y.; Sun, S.; Zhao, L.; Zhang, H.; Zhang, G. Schisandra lignan extract protects against carbon tetrachloride-induced liver injury in mice by inhibiting oxidative stress and regulating the NF-κB and JNK signaling pathways. Evid.-Based Complement. Altern. Med. 2017, 2017, 5140297. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.F.; Safhi, M.M.; Anwer, T.; Siddiqui, R.; Khan, G.; Moni, S.S. Therapeutic potential of Vanillylacetone against CCl4 induced hepatotoxicity by suppressing the serum marker, oxidative stress, inflammatory cytokines and apoptosis in Swiss albino mice. Exp. Mol. Pathol. 2018, 105, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Tan, H.Y.; Wang, N.; Zhang, Z.J.; Lao, L.; Wong, C.W.; Feng, Y. The role of oxidative stress and antioxidants in liver diseases. Int. J. Mol. Sci. 2015, 16, 26087–26124. [Google Scholar] [CrossRef] [PubMed]
- Del Campo, J.A.; Gallego, P.; Grande, L. Role of inflammatory response in liver diseases: Therapeutic strategies. World J. Hepatol. 2018, 10, 1–7. [Google Scholar] [CrossRef]
- Ravishankar, D.; Rajora, A.K.; Greco, F.; Osborn, H.M.I. Flavonoids as prospective compounds for anti-cancer therapy. Int. J. Biochem. Cell Biol. 2013, 45, 2821–2831. [Google Scholar] [CrossRef]
- Ma, J.Q.; Ding, J.; Zhao, H.; Liu, C.M. Puerarin attenuates carbon tetrachloride-induced liver oxidative stress and hyperlipidaemia in mouse by JNK/c-Jun/CYP7A1 pathway. Basic Clin. Pharmacol. Toxicol. 2014, 115, 389–395. [Google Scholar] [CrossRef]
- Ma, J.Q.; Li, Z.; Xie, W.R.; Liu, C.M.; Liu, S.S. Quercetin protects mouse liver against CCl4-induced inflammation by the TLR2/4 and MAPK/NF-κB pathway. Int. Immunopharmacol. 2015, 28, 531–539. [Google Scholar] [CrossRef]
- Kang, M.-C.; Kang, S.-M.; Ahn, G.; Kim, K.-N.; Kang, N.; Samarakoon, K.W.; Oh, M.-C.; Lee, J.-S.; Jeon, Y.-J. Protective effect of a marine polyphenol, dieckol against carbon tetrachlorideinduced acute liver damage in mouse. Environ. Toxicol. Pharmacol. 2013, 35, 517–523. [Google Scholar] [CrossRef]
- Jiang, H.; Wu, D.; Xu, D.; Yu, H.; Zhao, Z.; Ma, D.; Jin, J. Eupafolin exhibits potent anti-angiogenic and antitumor activity in hepatocellular carcinoma. Int. J. Biol. Sci. 2017, 13, 701–711. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Wang, L.; Sun, Z.; Li, H.; Wang, Q.; Yi, C.; Wang, X. Morusin shows potent antitumor activity for human hepatocellular carcinoma in vitro and in vivo through apoptosis induction and angiogenesis inhibition. Drug Des. Devel. Ther. 2017, 11, 1789–1802. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.B.; Zhang, H.F.; Zhang, X.; Li, D.Y.; Xue, H.Z.; Pan, C.E.; Zhao, S.H. Resveratrol inhibits VEGF expression of human hepatocellular carcinoma cells through a NF-kappa B-mediated mechanism. Hepatogastroenterology 2010, 57, 1241–1246. [Google Scholar] [PubMed]
- Hickman, J.C. The Jepson Manual: Higher Plants of California; University of California Press: Los Angeles, CA, USA, 1993; p. 1400. [Google Scholar]
- USDA (United States Department of Agriculture); NRCS (Natural Resource Conservation Service). The PLANTS Database; National Plant Data Center: Baton Rouge, LA, USA; pp. 70874–74490. Available online: http://plants.usda.gov (accessed on 6 March 2006).
- Marwat, S.K.; Salvadora, P. Tamarix aphylla and Zizyphus mauritiana: Thouree Woody Plant Species Mentioned in Holy Quran and Ahadith, and their Ethnobotanical Uses in North Western Part (DI Khan) of Pakistan. Ethnobot. Leafl. 2008, 2008, 135. [Google Scholar]
- Auribie, M.A. Antioxidant activity of tannin fromTamarix aphylla L. leaves. Basra J. Agric. Sci. 2011, 24, 33–37. [Google Scholar]
- El Ansari, M.A.; Nawwar, M.A.M.; El Dein, A.; El Sherbeiny, A.; El Sissi, H.I. A sulphated kaempferol 7,4′-dimethyl ether and a quercetin isoferulylglucuronide from the flowers of Tamarix aphylla. Phytochemistry 1976, 15, 231–232. [Google Scholar] [CrossRef]
- Souliman, A.M.A.; Barakat, H.H.; El-Mousallamy, A.M.D.; Marzouk, M.S.A.; Nawwar, M.A.M. Phenolics fromthe bark of Tamarix aphylla. Phytochemistry 1991, 30, 3763–3766. [Google Scholar] [CrossRef]
- Mahfoudhi, A.; Grosso, C.; Gonc, R.F.; Khelifi, E.; Hammami, S.; Achour, S.; Trabelsi-Ayadi, M.; Valentão, P.; Andrade, P.B.; Mighri, Z. Evaluation of antioxidant, anti-cholinesterase, and antidiabetic potential of dry leaves and stems in Tamarix aphylla growing wild in Tunisia. Chem. Biodivers. 2016, 12, 1747–1755. [Google Scholar] [CrossRef]
- Yusufoglu, H.S.; Alqasoumi, S.I. Anti-inflammatory and wound healing activities of herbal gel containing an antioxidant tamarix aphylla leaf extract. Int. J. Pharmacol. 2011, 7, 829–835. [Google Scholar]
- Qadir, M.I.; Abbas, K.; Hamayun, R.; Ali, M. Analgesic, anti-inflammatory and anti-pyretic activities of aqueous ethanolic extract of Tamarix aphylla L. (Saltcedar) in mice. Pak. J. Pharm. Sci. 2014, 27, 1985–1988. [Google Scholar]
- El-Aarag, B.; Hussein, W.; Ibrahim, W.; Zahran, M. Thymoquinone Improves Anti-Diabetic Activity of Metformin in Streptozotocin-Induced Diabetic Male Rats. J. Diabetes Metab. 2017, 8, 780. [Google Scholar]
- El-Aarag, B.; Magdy, M.; AlAjmi, M.F.; Khalifa, S.A.; El-Seedi, H.R. Melittin Exerts Beneficial Effects on Paraquat-Induced Lung Injuries in Mice by Modifying Oxidative Stress and Apoptosis. Molecules 2019, 24, 1498. [Google Scholar] [CrossRef] [PubMed]
- Zahra, M.H.; Salem, T.A.; El-Aarag, B.; Yosri, N.; EL-Ghlban, S.; Zaki, K.; Marei, A.H.; Abd El-Wahed, A.; Saeed, A.; Khatib, A.; et al. Alpinia zerumbet (Pers.): Food and Medicinal Plant with Potential In Vitro and In Vivo Anti-Cancer Activities. Molecules 2019, 24, 2495. [Google Scholar] [CrossRef] [PubMed]
- El-Saied, F.; El-Aarag, B.; Salem, T.; Said, G.; Khalifa, S.A.M.; El-Seedi, H.R. Synthesis, Characterization, and In Vivo Anti-Cancer Activity of New Metal Complexes Derived from Isatin-N(4)antipyrinethiosemicarbazone Ligand Against Ehrlich Ascites Carcinoma Cells. Molecules 2019, 24, 3313. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.; Dai, C.; Liu, Q.; Li, J.; Qiu, J. Curcumin attenuates on carbon tetrachloride-induced acute liver injury in mice via modulation of the Nrf2/HO-1 and TGF-_1/Smad3 pathway. Molecules 2018, 23, 215. [Google Scholar] [CrossRef]
- Wang, R.; Yang, Z.; Zhang, J.; Mu, J.; Zhou, X.; Zhao, X. Liver Injury Induced by Carbon Tetrachloride in Mice Is Prevented by the Antioxidant Capacity of Anji White Tea Polyphenols. Antioxidants 2019, 8, 64. [Google Scholar] [CrossRef]
- Zhanga, Y.; Miaoa, H.; Yana, H.; Shengb, Y.; Jia, L. Hepatoprotective effect of Forsythiae Fructus water extract against carbon tetrachloride-induced liver fibrosis in mice. J. Ethnopharmacol. 2018, 218, 27–34. [Google Scholar] [CrossRef]
- Wang, Y.; Tang, C.; Zhang, H. Hepatoprotective effects of kaempferol 3-O-rutinoside and kaempferol 3-O-glucoside from Carthamus tinctorius L. on CCl4-induced oxidative liver injury in mice. J. Food Drug Anal. 2015, 23, 310–317. [Google Scholar] [CrossRef]
- Mansour, M.A. Protective effects of thymoquinone and desferrioxamine against hepatotoxicity of carbon tetrachloride in mice. Life Sci. 2000, 66, 2583–2591. [Google Scholar] [CrossRef]
- Dreissgacker, U.; Suchy, M.T.; Maassen, N.; Tsikas, D. Human plasma concentrations of malondialdehyde (MDA) and the F2-isoprostane 15(S)-8-iso-PGF2a may be markedly compromised by hemolysis: Evidence by GC-MS/MS and potential analytical and biological ramifications. Clin. Biochem. 2010, 43, 159–167. [Google Scholar] [CrossRef]
- Zou, J.; Qi, F.; Ye, L.; Yao, S. Protective Role of grape seed proanthocyanidins against CCl4 induced acute liver injury in mice. Med. Sci. Monit. 2016, 22, 880–889. [Google Scholar] [CrossRef]
- Lee, Y.-S.; Cho, J.; Wan Kim, J.; Lee, M.-K.; Ku, S.K.; Choi, J.-S.; Lee, H.-J. Hepatoprotective effects of blue honeysuckle on CCl4-induced acute liver damaged mice. Food Sci. Nutr. 2019, 7, 322–338. [Google Scholar] [CrossRef]
- Lu, Y.; Chen, J.; Ren, D.; Yang, X.; Zhao, Y. Hepatoprotective effects of phloretin against CCl4-induced liver injury in mice. Food Agric. Immunol. 2017, 28, 211–222. [Google Scholar] [CrossRef]
- Benavente-Garcia, O.; Castillo, J. Update on uses and properties of citrus flavonoids: New findings in anticancer, cardiovascular, and anti-inflammatory activity. J. Agric. Food Chem. 2008, 56, 6185–6205. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.Y.; Li, Y.; Wang, F.; Wu, C.F. Hepatoprotective effects of apple polyphenols on CCl4-induced acute liver damage in mice. J. Agric. Food Chem. 2010, 58, 6525–6531. [Google Scholar] [CrossRef] [PubMed]
- Sahreen, S.; Khan, M.R.; Khan, R.A. Hepatoprotective effects of methanol extract of Carissa opaca leaves on CCl4-induced damage in rat. BMC Complement. Altern. Med. 2011, 11, 48–56. [Google Scholar] [CrossRef]
- Al-Sayed, E.; El-Lakkany, N.M.; Seif El-Din, S.H.; Sabra, A.N.; Hammam, O.A. Hepatoprotective and antioxidant activity of Melaleuca styphelioides on carbon tetrachloride-induced hepatotoxicity in mice. Pharm. Biol. 2014, 52, 1581–1590. [Google Scholar] [CrossRef]
- Cui, Y.M.; Yang, X.B.; Lu, X.S.; Chen, J.W.; Zhao, Y. Protective effects of polyphenolsenriched extract from Huangshan Maofeng green tea against CCl4-induced liver injury in mice. Chem. Biol. Interact. 2014, 220, 75–83. [Google Scholar] [CrossRef]
- Sun, Y.F.; Yang, X.B.; Lu, X.S.; Wang, D.Y.; Zhao, Y. Protective effects of Keemun black tea polysaccharides on acute carbon tetrachloride-caused oxidative hepatotoxicity in mice. Food Chem. Toxicol. 2013, 58, 184–192. [Google Scholar] [CrossRef]
- Yang, X.B.; Yang, S.; Guo, Y.R.; Jiao, Y.D.; Zhao, Y. Compositional characterisation of soluble apple polysaccharides, and their antioxidant and hepatoprotective effects on acute CCl4- caused liver damage in mice. Food Chem. 2013, 138, 1256–1264. [Google Scholar] [CrossRef]
- Lopez, J.; Tait, S.W.G. Mitochondrial apoptosis: Killing cancer using the enemy within. Br. J. Cancer 2015, 112, 957–962. [Google Scholar] [CrossRef] [PubMed]
- Zaman, S.; Wang, R.; Gandhi, V. Targeting the apoptosis pathway in hematologic malignancies. Leuk. Lymphoma 2014, 55, 1980–1992. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Song, F.; Li, S.; Wu, B.; Gu, Y.; Yuan, Y. Salvianolic acid A attenuates CCl4-induced liver fibrosis by regulating the PI3K/AKT/mTOR, Bcl-2/Bax and caspase-3/cleaved caspase-3 signaling pathways. Drug Des. Dev. Ther. 2019, 13, 1889–1900. [Google Scholar] [CrossRef]
- Guo, X.-L.; Liang, B.; Wang, X.-W.; Fan, F.-G.; Jin, J.; Lan, R.; Yang, J.-H.; Wang, X.-C.; Jin, L.; Cao, Q. Glycyrrhizic acid attenuates CCl4-induced hepatocyte apoptosis in rats via a p53-mediated pathway. World J. Gastroenterol. 2013, 28, 3781–3791. [Google Scholar] [CrossRef] [PubMed]
- Dooley, S.; Dijke, P. TGF-β in progression of liver disease. Cell Tissue Res. 2012, 347, 245–256. [Google Scholar] [CrossRef] [PubMed]
- McMillin, M.; Galindo, C.; Pae, H.Y.; Frampton, G.; Di Patre, P.L.; Quinn, M.; Whittington, E.; de Morrow, S. Gli1 activation and protection against hepatic encephalopathy is suppressed by circulating transforming growth factor beta1 in mice. J. Hepatol. 2014, 61, 1260–1266. [Google Scholar] [CrossRef] [PubMed]
- Niu, L.; Cui, X.; Qi, Y.; Xie, D.; Wu, Q.; Chen, X.; Ge, J.; Liu, Z. Involvement of TGF-β1/Smad3 Signaling in Carbon Tetrachloride-Induced Acute Liver Injury in Mice. PLoS ONE 2016, 11, e0156090. [Google Scholar] [CrossRef]
- Jiang, J.; Li, X.; Pan, D.; Lin, Y. Protective effect of tripterine on carbon tetrachloride-induced hepatic fibrosis in immature rats. Int. J. Clin. Exp. Med. 2019, 12, 4161–4167. [Google Scholar]
- Zhou, Y.N.; Mu, Y.P.; Fu, W.W.; Ning, B.B.; Du, G.L.; Chen, J.M. Yiguanjian decoction and its ingredients inhibit angiogenesis in carbon tetrachloride-induced cirrhosis mice. BMC Complement. Altern. Med. 2015, 15, 342–351. [Google Scholar] [CrossRef]
- Woodfin, A.; Voisin, M.-B.; Nourshargh, S. PECAM-1: A Multi-Functional Molecule in Inflammation and Vascular Biology. Arterioscler. Thromb. Vasc. Biol. 2007, 27, 2514–2523. [Google Scholar] [CrossRef] [Green Version]
- Brown, S.; Heinisch, I.; Ross, E.; Shaw, K.; Buckley, C.D.; Savill, J. Apoptosis disables CD31-mediated cell detachment from phagocytes promoting binding and engulfment. Nature 2002, 418, 200–203. [Google Scholar] [CrossRef] [PubMed]
- Coulon, S.; Heindryckx, F.; Geerts, A.; Van Steenkiste, C.; Colle, I.; Van Vlierberghe, H. Angiogenesis in chronic liver disease and its complications. Liver Int. 2011, 31, 146–162. [Google Scholar] [CrossRef] [PubMed]
- Elpek, G.O. Angiogenesis and liver fibrosis. World J. Hepatol. 2015, 7, 377–391. [Google Scholar] [CrossRef]
- Fernández, M.; Semela, D.; Bruix, J.; Colle, I.; Pinzani, M.; Bosch, J. Angiogenesis in liver disease. J. Hepatol. 2009, 50, 604–620. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Li, Z.; Xiu, A.-Y.; Meng, D.-X.; Wang, S.-N.; Zhang, C.-Q. Carvedilol attenuates carbon tetrachloride-induced liver fibrosis and hepatic sinusoidal capillarization in mice. Drug Des. Dev. Ther. 2019, 13, 2667–2676. [Google Scholar] [CrossRef]
- Cheng, G.-Y.; Jiang, Q.; Deng, A.-P.; Wang, Y.; Liu, J.; Zhou, Q.; Zheng, X.-H.; Li, Y.-Y. CD31 induces inflammatory response by promoting hepatic inflammatory response and cell apoptosis. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 7543–7550. [Google Scholar]
- Domitrović, R.; Jakovac, H.; Tomac, J.; Šain, I. Liver fibrosis in mice induced by carbon tetrachloride and its reversion by luteolin. Toxicol. Appl. Pharmacol. 2009, 241, 311–321. [Google Scholar] [CrossRef]
- Bancroft, J.D.; Stevens, A. Theory and Practice of Histological Techniques, 4th ed.; Elsevier: New York, NY, USA, 1996. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mice Group | SOD (U/g Tissue) | CAT (U/g Tissue) | Gpx (U/g Tissue) |
|---|---|---|---|
| Normal control | 1.23 ± 0.03 | 2.69 ± 0.01 | 38.74 ± 3.61 |
| CCl4 model group | 0.32 ± 0.07 ### | 1.98 ± 0.23 ### | 21.76 ± 4.01 ## |
| Flavone (10 mg/kg) | 0.58 ± 0.02* | 2.27 ± 0.03** | 27.65 ± 2.35* |
| Flavone (25 mg/kg) | 0.89 ± 0.07** | 2.48 ± 0.29** | 30 ± 1.63* |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
El-Aarag, B.; Khairy, A.; Khalifa, S.A.M.; El-Seedi, H.R. Protective Effects of Flavone from Tamarix aphylla against CCl4-Induced Liver Injury in Mice Mediated by Suppression of Oxidative Stress, Apoptosis and Angiogenesis. Int. J. Mol. Sci. 2019, 20, 5215. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20205215
El-Aarag B, Khairy A, Khalifa SAM, El-Seedi HR. Protective Effects of Flavone from Tamarix aphylla against CCl4-Induced Liver Injury in Mice Mediated by Suppression of Oxidative Stress, Apoptosis and Angiogenesis. International Journal of Molecular Sciences. 2019; 20(20):5215. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20205215
Chicago/Turabian StyleEl-Aarag, Bishoy, Asmaa Khairy, Shaden A. M. Khalifa, and Hesham R. El-Seedi. 2019. "Protective Effects of Flavone from Tamarix aphylla against CCl4-Induced Liver Injury in Mice Mediated by Suppression of Oxidative Stress, Apoptosis and Angiogenesis" International Journal of Molecular Sciences 20, no. 20: 5215. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20205215