Hydrogen Sulfide in Bone Tissue Regeneration and Repair: State of the Art and New Perspectives


Laboratorio RAMSES: IRCCS Istituto Ortopedico Rizzoli, Via di Barbiano 1/10, 40136 Bologna, Italy
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2019, 20(20), 5231; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20205231
Submission received: 10 September 2019
/
Revised: 17 October 2019
/
Accepted: 18 October 2019
/
Published: 22 October 2019
(This article belongs to the Special Issue Hydrogen Sulfide in the Diseases and Tissue Regeneration)
Abstract
:The importance of hydrogen sulfide (H2S) in the regulation of multiple physiological functions has been clearly recognized in the over 20 years since it was first identified as a novel gasotransmitter. In bone tissue H2S exerts a cytoprotective effect and promotes bone formation. Just recently, the scientific community has begun to appreciate its role as a therapeutic agent in bone pathologies. Pharmacological administration of H2S achieved encouraging results in preclinical studies in the treatment of systemic bone diseases, such as osteoporosis; however, a local delivery of H2S at sites of bone damage may provide additional opportunities of treatment. Here, we highlight how H2S stimulates multiple signaling pathways involved in various stages of the processes of bone repair. Moreover, we discuss how material science and chemistry have recently developed biomaterials and H2S-donors with improved features, laying the ground for the development of H2S-releasing devices for bone regenerative medicine. This review is intended to give a state-of-the-art description of the pro-regenerative properties of H2S, with a focus on bone tissue, and to discuss the potential of H2S-releasing scaffolds as a support for bone repair.
1. Introduction
Tissue development and repair has long been associated with sulfur metabolism: The first evidence dates back to the study by Williamson and Fromm (1955) which found an association between the activation of sulfur-containing amino acids metabolism and the wound healing process [1]. However, more than six decades have passed before the first studies linking biological functions of sulfurated amino acids to hydrogen sulfide (H2S) came to light. Notably, the identification (1996) of a physiological role of H2S in the brain tissue [2] opened the way to several studies unravelling its multiple biological functions. H2S is a gaseous molecule produced endogenously by the enzymes cystathionine-β-synthase (CBS), cystathionine-γ-lyase (CSE) and, to a lower extent, 3-mercaptopyruvate sulfurtransferase (MPST) [3], within the transsulfuration pathway. A broad range of studies further clarified that H2S is not only a “secondary reaction product” of this pathway but is a critical mediator of physiological functions in many tissues.
Over the past years, several therapeutic applications of sulfur-containing compounds or H2S-releasing donors have emerged and a few clinical trials are ongoing [4]. However, while the studies on the systemic use of H2S-donors are at an advanced stage, the need for obtaining an organ-specific delivery of H2S to exploit the tissue-specific cytoprotective and regenerative properties of H2S has only recently emerged [5]. One field where H2S-releasing biomaterials hold a great potential interest is bone repair and regeneration.
Bone is a vascularized, dynamic tissue which provides the body with structural and movement support functions, organ protection, mineral reserve, and endocrine system function [6,7]. It is continuously remodeled by the coordination and balance of two key processes, bone formation and bone resorption [8,9], and is capable of repairing and reshaping without leaving scars [10]. However, bone lesions caused by trauma and pathological bone resorption (infectious diseases, biochemical disorders, congenital disorders, or abnormal skeletal development) have limited capacity to heal and require large amounts of autologous/allogenic bone or bone substitutes for reconstruction [11,12]. In recent years, research approaches have moved from a focus on inert biomaterials toward the complex design of biomaterials mimicking the structure of the native bone extracellular matrix and being able to provide a controlled release of biomimetic, pro-osteogenic factors [13].
Over the past few years, it has become increasingly clear that H2S regulates bone homeostasis and that systemic administration of H2S holds good therapeutic potential, as it prevented or reversed pathological bone loss [14,15,16]. However, whether H2S affects the processes of bone tissue regeneration and repair and whether it could be used as a biomimetic factor in regenerative medicine approaches has not been fully elucidated. This review is intended to first summarize the emerging evidence linking H2S and bone tissue repair and regeneration. Then, the state-of-the-art of the development of H2S-releasing scaffolds and biomaterials for promoting tissue regeneration will be described, with a focus on bone repair. Finally, implications and future challenges in the field of H2S-releasing scaffold for bone tissue engineering (BTE) will be discussed.
2. Bone Tissue Repair: H2S as a Suitable Biological Cue for Scaffold Functionalization
Bone remodeling is an orchestrated process constantly occurring to let the skeleton adapt to biomechanical loading, microfractures, environmental stress [17], and to control calcium and other ions levels in the circulation. Similarly, bone repair occurs in response to injury by activating complex and orchestrated regenerative processes to restore bone tissue integrity, structure, and homeostasis [18,19]. Despite the high innate regenerative capacity of bone, critical-size defects and non-unions fail to heal, thus making them a global health burden and a major challenge in regenerative medicine [12,20,21,22]. Autograft is currently the gold standard therapy and achieve an 80% success rate [23]. However, the percentage of non-union remains significantly high and the procedure involves two surgical operations and donor site morbidity for patients, thus conditioning their quality of life. A wide variety of bone substitutes have been tested in clinics to improve bone repair, among which are allogenic bone and natural or synthetic osteoconductive matrices [24,25], obtaining promising results. In this context, BTE offers the opportunity to develop innovative alternative treatment options that will ideally eliminate the limitations of current treatments. BTE is based on various combinations of three principal components: biomaterials as scaffolds, regulatory signals such as growth factors (GF), and cells. Recently, the design of functionalized cell-free scaffolds has emerged as a useful tool to overcome current drawbacks linked to cell-based approaches (limited autologous cells, time/cost-intensive cell expansion procedures, relative low cell survival rate, and high risk of immune-rejection) [26]. Strategies of scaffold functionalization involve GF, chemokines, and peptides that are able to activate bone resident cells and promote bone regeneration [26]. These biological cues are usually incorporated into scaffolds via physical adsorption, chemical covalent coupling, and encapsulation [26], and their controlled and site specific release is crucial for the success of tissue engineering strategies. Over the past 50 years, several studies aimed at understanding the biological processes which takes place in in vivo models of bone fractures during bone repair [27], particularly in in the presence of bone fractures [28,29], and allowed the identification of several biological cues. Accordingly, bioinspired and biomimetic cell-free scaffolds were recently developed to mimic the natural self-healing events of bone healing by using biological signals able to modulate inflammation and induce cell recruitment, vascularization, and osteogenic differentiation [26]. Figure 1 summarizes the biological processes involved in bone fracture repair, highlighting which of them may be regulated by H2S (discussed more in detail in the next sections).
2.1. H2S Regulates Inflammation in Bone Tissue
Recent investigations in the field of osteoimmunology have demonstrated extensive cross-talk between immune and skeletal systems [30]. In particular, upon bone injury, hematoma formation is closely followed by acute inflammation and release of inflammatory cytokines which activate a cascade of processes to promote bone healing. A tightly regulated balance between pro-inflammatory macrophages (M1) and pro-healing, anti-inflammatory macrophages (M2) results in inflammatory and mesenchymal stromal cells (MSCs) recruitment, promotion of neovascularization, and induction of MSCs towards osteogenesis [30]. Notably, their absence has been shown to completely abolish callus formation [31]. Treg also promotes tissue healing by inducing M2 polarization, inhibiting neutrophil infiltration, activity, and inducing their apoptosis [32], supporting MSCs differentiation [33] and inhibiting osteoclasts (OCs) differentiation [32]. The ablation of δγ-T-cells promotes bone healing since mice deficient in this cellular type have a shorter time to fracture union and show improved biomechanical strength compared to control mice [34]. Conversely high presence of T-cells promotes OCs differentiation and inhibit MSCs [35] causing additional “by-stander” tissue damage [30]. Therefore, scaffold functionalization strategies are currently aimed at modulating pro-inflammatory and anti-inflammatory activity, thus preventing chronic inflammation and unbalanced inflammatory responses responsible for the formation of fibrous capsules around scaffolds, which make scaffolds bioinert [36].
H2S plays an important role during inflammation in several tissues [37] and the increased H2S levels occurring during inflammation have been proposed as a stress-response mechanism to the injury caused by oxidative stress [38]. In OCs, inhibition of reactive oxygen species (ROS) signaling by H2S occurs through direct and indirect mechanisms by triggering NRF-2-dependent antioxidant response and thus inhibiting osteoclast differentiation (Figure 2) [39]. Interestingly, H2S was shown to be a key mediator in the immediate anti-inflammatory and antioxidant function of N-acetyl-cysteine, a broadly used mucolytic medicine [40]. H2S was shown to reduce edema formation [41], promote neutrophil apoptosis, suppress the expression of some leukocyte and endothelial adhesion molecules [42], stimulate macrophages polarization to M2 [43,44,45], and activate the AnxA1 pro-resolutive pathway [46]. Concerning bone tissue, in the animal model of hyperhomocysteinemia (CBS−/− mice), the autosomal recessive disease involving CBS, higher levels of inflammation has been correlated to lower bone tissue [16]. Particularly, these mice had increased acetylation of NF-kB p65, resulting in increased levels of IL-6 and TNF-α in the circulation and inhibition of Runx-2 and Ocn gene expression [16]. Administration of NaHS suppressed histone acetylation dependent NF-kB p65 signaling activation and inflammation and rescued osteogenic genes expression [16]. Moreover, H2S exogenous administration (by using the H2S-releasing moiety of ATB-346 and ATB-352 molecules) has been used to inhibit inflammation and inflammatory bone loss in experimental periodontitis [47,48]. A therapeutic platform has been developed to deliver H2S and inhibit P-selectin expression in human platelets, which play an essential role in the initial recruitment of leukocytes to the site of injury during inflammation [49]. Moreover, H2S has been shown to mediate the inhibition of inflammatory responses during bacterial infections in an ex vivo model of cells infected with mycoplasma [50]. The molecular pathways imply inhibition and activation of nuclear translocation of NF-κB, reducing the transcription of pro-inflammatory genes [50]. Finally, H2S plays a role in mediating both innate and adaptive immunity, since it was also found to be essential for Treg cell differentiation and function and enhances T-cell proliferation, activation, and cell death [51,52]. Moreover, CBS−/− mice developed autoimmune disease [53].
In tissues other than bone, H2S has been shown to improve wound healing by attenuating inflammation and increasing angiogenesis. H2S has been hypothesized to play a role in severe burns, which are associated with processes that causes increased permeability and edema [54], and has been proven to improve diabetic wound healing [55,56].
Taken together these data indicates that future design of H2S-releasing scaffolds for BTE may be tuned to temporally modulate pro-inflammatory and anti-inflammatory activities, thus promoting the formation of regenerated bone tissue.
2.2. H2S Regulates Migration and Survival of Cells Involved in Bone Repair
The recruitment of autogenous endothelial progenitor cells (EPCs) and MSCs to the site of bone defect is critical to induce bone tissue repair [57]. More in general, EPCs play a crucial role in endothelial restoration after arterial injury [58], and the migratory potential of MSCs has been demonstrated to be one of the most important factors for neovascularization and regeneration [59,60]. H2S has been found to induce migratory properties in both cell types. In particular, NaHS, an H2S donor, increases the mobilization of EPCs after vascular injury, enhancing their adhesion and colony formation capacities [61] and improving re-endothelization in nude mice with carotid artery injury [62]. CSE-mediated H2S deficiency in high glucose-treated MSCs impairs their migratory capacity, which was further restored by NaHS treatments [63]. Furthermore, H2S has been found to mediate the migratory potential of other cells types. In particular, NaHS accelerates skin fibroblasts and human keratinocytes in migration assays [64], thus enhancing wound healing. H2S promotes macrophage migration to an infarcted area, by accelerating internalization of integrin β1 and activating the downstream Src-FAK/Pyk2-Rac pathway, thus promoting infarction healing [65].
Moreover, it is well known that H2S promotes proliferation and survival of MSCs and of other cell types during several stresses such as hypoxia, oxidative damage, and serum deprivation. Particularly, it inhibits cellular death by apoptosis [66,67]. Therefore, H2S may act similarly to other GF, which initiate the repair process by facilitating proliferation and differentiation of the stem cells that give rise to the fracture callus [68].
Whether H2S modulates cells migration, survival, and proliferation to the bone defect to induce bone tissue repair is still an open question.
2.3. H2S Promotes the Expression of Angiogenic Factors in Bone Tissue
Vascularization is a primary driving force behind both intramembranous and endochondral ossification. To achieve a highly functional new regenerated tissue, the tight connection of new vasculature formation and osteogenesis is of utmost importance. New vessels are fundamental both in the initial and in the final step of bone repair. In the first phase, new blood vessels are essential to provide nutrients and facilitate cells migration toward the damaged site and in the final phases of repair, vascular remodeling occurs and vessels regresses to the original state [69]. Importantly, failure of angiogenesis can lead to nonunion [70]. Deletion of vascular endothelial growth factor (VEGF) in osteoblasts (OBs) interrupts the coupling of osteogenesis and angiogenesis, and delays the intramembranous ossification-mediated repair during cortical bone defect healing [71]. To date, there is no information linking H2S promotion of neo-angiogenesis during bone regeneration and repair. However, several evidences may suggest a role. H2S has been shown to stimulate in vitro the three processes crucial for the neo-angiogenesis: proliferation, migration, and tube-like network formation by endothelial cells [72]. Evidence of neo-vascularization of Matrigel implants in vivo further confirmed the ability of H2S in promoting new blood vessel formation [73,74]. These features are at least partly mediated by H2S-dependent activation of two factors critical for bone vascularization: hypoxia-inducible factor-1 (HIF-1α) and VEGF. Indeed, a number of studies have established extensive crosstalk between H2S and VEGF. Exogenously administered H2S has been shown to upregulate VEGF expression in several tissues and cells [75], among which skeletal muscle cells and bone cells in vitro and in vivo [76,77]. H2S also significantly upregulated HIF-1alpha protein levels and increased HIF-1alpha binding activity under hypoxic conditions [78]. Finally, H2S improved vascular density and blood flow in ischemic skeletal muscles of CBS+/− mice or diabetic type-2FAL mice models with hindlimb femoral artery ligation [63,79].
2.4. H2S Stimulates Bone Formation
Over the past few years, it has become increasingly clear that H2S affects bone tissue regeneration by acting at several levels such as regulation of bone cells activity, reduction of oxidative stress, regulation of calcium intake by bone cells, and promotion of angiogenesis. CBS and CSE, the H2S-producing enzymes, are expressed both in MSCs [15,80] and in OBs [80,81,82]. In particular, CSE is the predominant source of H2S in OBs [81]. H2S plays a cytoprotective role in bone cells; it protects OBs against homocysteine-induced mitochondrial toxicity [83]; and OBs against hydrogen peroxide (H2O2)-induced cell death and apoptosis [84].
H2S was shown to play an important role in sustaining osteogenic differentiation of human MSCs (hMSCs) [14,15] and in modulating OCs differentiation [39,85,86,87]. CBS and CSE levels increased during osteogenic differentiation and play an essential role in osteogenic differentiation in vitro [15,80]. Notably, H2S-deficiency due to CBS knock-down was linked to impaired osteogenesis and bone loss in mice [15]. Several pathways are implicated in H2S-dependent induction of osteogenesis, summarized in Figure 2: the WNT pathway [14,88]; BMP pathway [77]; and p38-MAPK and ERK signaling pathways [84,89]. Moreover, S-sulfhydration of cysteine residues in critical proteins involved in osteogenesis, such as Runx-2 [81] and Ca2+ TRP channels [15], has been shown to be a mechanism of H2S-dependent induction of osteogenesis. Furthermore, overexpression of CSE in OBs increased Runx-2 S-sulfhydration and enhanced OBs biologic function (increased ALP activity, Alizarin red-positive calcification nodules, and osteogenic gene expression). Interestingly, mechanical loading in periodontal tissues up-regulated CSE expression and OCs formation inducing bone remodeling and accelerating orthodontic tooth movement (OTM) speed. Similarly, OTM and bone remodeling are stimulated by the up-regulation of osteoclastogenesis and osteoblastogenesis induced by H2S exogenous administration [90].
An imbalance in H2S metabolism is associated with defective bone homeostasis. Evidence from several preclinical models showed that the depletion of H2S levels is implicated in bone loss; data were reported in ovariectomized mice [14,91], in H2S-deficient CBS+/− mice [15], in a model of H2S-deficiency linked to diet-induced hyperhomocysteinemia [16,92], and in methionine-restricted fed rats [93]. Interestingly, when the treatment with H2S donors in these models was investigated, finalized to normalize the plasma level of H2S, it was found that H2S was able to prevent or even reverse the bone loss. Moreover, asides from these conditions, H2S exogenous administration has been shown to have therapeutic potential against bone loss induced by modelled microgravity [94] and other conditions such as distraction osteogenesis [95,96]. Overall, these data demonstrate that H2S regulates osteogenesis and bone formation in both healthy and pathological conditions.
In the context of bone repair, new bone formation occurs by direct and indirect mechanisms, involving intramembranous and endochondral ossification, as extensively reviewed elsewhere [97,98,99,100,101]. During these processes MSCs proliferate and differentiate into the osteogenic or chondrogenic lineages and increase the production of blood vessels from pre-existing vessels.
In models of fracture healing, treatment with H2S by various means affected both direct and indirect mechanisms of bone healing. In a mouse model of distraction osteogenesis, H2S injection was observed to accelerate the formation of mature microstructures of trabeculae and mineralization substituting the fibrous-connective tissue and improve the mechanical properties of the callus healing within the distraction gaps [95]. Moreover, in a rat model of intramedullary fixed fractural bone, Zheng et al. investigated the delivery of CSE adenovirus through absorbable gelatin. CSE overexpression resulted in improved bone fracture healing: less inflammatory infiltration, greater collagen expression, fibrocartilage, and osteocyte deposition. Interestingly, the authors showed increased endochondral ossification at the tissue repair site, and a full bridge between fracture sites in rat femurs [81].
A summary of the main studies investigating the effects of H2S-releasing donors on bone cells are reported in Table 1.
3. H2S-Releasing Scaffolds for Regenerative Medicine
Besides the efficacy of systemic administration of H2S, several reasons provide a rationale for the development of H2S-releasing scaffolds for bone regeneration.
First, cell-free scaffolds have undergone intense development in the recent years due to the drawbacks linked to the employment of cells in regenerative medicine [26]. Many physical, chemical, or biological functionalizations have led to tremendous progresses in the ability to support the physiological healing process of the bone [26]. For example, it was demonstrated that topographically defined implants are able to recruit osteoprogenitor cells at the site of bone injury and promote a more efficient repair in vivo [102,103]. Moreover, chemical modifications of the surface can endow biomaterials with increased osteoinductive capacity; for example, doping dense biomaterials with inorganic ions such as magnesium (Mg), Cobalt (Co), silicon (Sr), and Strontium (Sr) was shown to promote osteogenic differentiation in vitro and in vivo [104,105,106,107].
Second, the field of H2S-releasing materials is readily expanding. Several reports over the past few years showed promising results from in vitro and in vivo models and demonstrated that the biological properties of H2S can be successfully recapitulated in H2S-releasing devices for biomedical applications. In a recent report, Wu et al. successfully doped phosporodithioate donors (JK-1), a kind of H2S donors triggered by hydrolysis and releasing H2S in a pH-dependent fashion, with polycaprolactone (PCL) and the resulting fibers showing a slower kinetic of H2S-release compared to the native molecules in solution. When applied in an animal model of full-thickness skin damage, the doped PCL-JK1 fiber showed overall improved wound healing times as compared to non-doped fibers [108], thus confirming that H2S released by the fibers retained the previously evidenced properties of promoting wound healing [64,109,110]. The same authors developed a biomimetic hyaluronic acid (HY) hydrogel doped with JK1, to overcome the low permeability to oxygen of PCL fibers. This scaffold accelerated the wound repair process by decreasing inflammation, driving macrophages polarization toward M2, and increasing angiogenesis, and the effect was higher compared to adding JK1 to the wound microenvironment without being embedded in HY [45].
Nano-structured patches obtained by electrospun fibers are a valuable tool to promote tissue healing and to achieve local release of bioactive compounds [111]. Cacciotti et al. recently developed H2S-releasing poly(lactic) acid fibrous membranes functionalized with H2S donors of natural origin (from garlic oil-soluble extracts and diallyl disulfide) [112]. These donors where entrapped within the fibers and the H2S release by the membranes was prolonged and gradual and induces MSCs proliferation and anti-microbial activity. Following a similar approach, Feng et al. added the H2S donors (NSHd1) to PCL fiber spun by electrospinning to produce controlled, thiol-triggered H2S release [113]. Interestingly, by tuning the diameter of the fibers the authors achieved a controlled rate of H2S release. The authors showed that H2S increased gene expression of collagen type I and type III in fibroblasts while protecting them from the oxidative stress produced by H2O2, suggesting that these microfibers may support wound dressing. The group of Mauretti et al. developed a photopolymerizable PEG-fibrinogen hydrogel incorporating albumin microbubbles functionalized with thiosulfate cyanide sulfurtransferase, one of the enzymes able to catalyze the H2S-production [114]. H2S produced by the scaffold increased cardiac progenitor cell proliferation, thus retaining the feature previously described by others [115]. The group of Liang et al. loaded a partially oxidized alginate (ALG-CHO) with the H2S donor 2-aminopyridine-5-thiocarboxamide and tetraaniline (a conductive oligomer), and adipose-derived stem cells (ADSCs) [116]. The hydrogel, which mimics the slow and continuous release of endogenous H2S, increased the ejection fraction value and reduced the infarction size in rats with myocardial infarction, thus evidencing that H2S released retained the H2S-dependent protective role on myocardial infarction and heart failure previously described [115] and offering a promising therapeutic strategy for treating infarction.
Overall, the strategies to add H2S to the biomaterials described above were based on physical adsorption, chemical covalent coupling, and encapsulation. Beside these strategies, chemical research is also growing to develop methods for obtaining H2S-releasing donors with improved physical and chemical features. Of interest, an H2S-releasing gel was developed by the group of JB Matson, by using an H2S-releasing peptide that self-assembles into a robust hydrogel in water, obtained by chemical synthesis of an aromatic peptide amphiphile and the H2S moiety, S-aroylthiooxime—SATO—functional group. This H2S-releasing gel, showed a slow kinetic of H2S release and absence of cytotoxicity [117]. In a recent report, this H2S-releasing hydrogel substantially reduced intimal hyperplasia in human veins, showing efficacy at a dose five times lower than NaHS, a fast release H2S donor [118].
Altogether, these data evidence how H2S can be used to functionalize biomaterials inducing cell differentiation and function, and thus can be considered as a regulatory signal in the “tissue engineering triad” (Figure 3).
The only reported H2S-releasing biomaterial targeting bone regeneration was developed by our group [77]. Based on previous preclinical studies showing efficient stimulation of osteogenesis by H2S donors, we postulated that a local release of physiologically relevant H2S concentrations may trigger osteogenic differentiation of h-MSCs. An H2S-releasing silk fibroin (SF) sponge was developed by first applying a salt leaching approach to add appropriate porosity to the SF fibers and by then doping the H2S donor (GYY4137) to the sponges. The resulting SF-GYY scaffolds showed unaltered mechanical properties and no cytotoxicity compared to the native SF scaffolds. When tested in a model of 3D culture of h-MSCs within a perfusion bioreactor, H2S induced a significant increase in the differentiation to mature OBs, revealed by increased deposition of the mineral matrix (as shown in Figure 4) and increased expression of osteogenic genes after three weeks in culture. This study first provided a proof-of-principle that exposing h-MSCs to H2S released by a scaffold induced their differentiation toward OBs, and demonstrated that loading osteoconductive biomaterials with an H2S donor is a suitable strategy to promote OBs differentiation at sites of bone regeneration.
Overall, the evidences reported in the present review set H2S as an important emerging candidate for promoting regenerative medicine and particularly for BTE.
4. Perspectives for H2S-Loaded Scaffolds for BTE
Although research on the biological activities of H2S was first triggered by investigations in the field of neurology and circulation, a substantial body of evidence, summarized in this review, has recently shown that H2S also plays an important role in bone physiology and can stimulate the process of bone repair at multiple levels. The field of H2S-releasing scaffolds for BTE holds the promise to open new opportunities to treat bone tissue damages. However, it is still on its infancy and a number of challenges are still ahead of us, which will require joint efforts from material scientists, chemists, and biologists.
First, the range of biomaterials used for the combination with H2S is limited. Interestingly, all the H2S-releasing scaffold before mentioned (PCL, polylactic, PEG, alginate, and peptide amphiphilic) are made by biomaterials previously tested for bone regeneration alone or composite [119,120,121,122,123], making them of interest also in the field of BTE. However, further combination with other biomaterials, especially those which better mimics the composition and the biomechanical properties of bone, is necessary to take a step forward in this field.
Second, the design of new formulations of H2S donors should address the need for improved stability (e.g., in aqueous solution during scaffold development) and a “controlled” release of H2S (for avoiding the burst–release phenomenon). Notably, several formulations of H2S donors has been developed to release H2S triggered by a change in pH [124,125] or amounts of thiols, reactive oxygen species, or enzymes [5,126,127,128,129] within the microenvironment. These formulations partially addressed this need and may provide further opportunities to tune the H2S release in bone microenvironments according to different phases of bone healing to better exploit its biological properties. However, changes in the technologies for scaffolds development require a steady advancement in the development of H2S-donors. As an example, H2S donors are able to endure the high temperature required for gel printing while preventing premature H2S release would be of great interest given the rapid expansion of 3D printing technology for the development of critical-size bone defect customized products.
Third, new formulations of H2S donors should be tailored for applications in BTE. As an example, H2S donors able to bind with high affinity bone or bone substitutes, such as hydroxyapatite, would be useful for optimizing the interaction of the H2S donor within the scaffold matrices. The use of H2S-donors hybridized with bisphosphonates, molecules able to bind avidly bone, such as the recently developed H2S-releasing DM-22 [130], could represent an interesting approach to this challenge.
The knowledge generated by studies addressing these points will be instrumental for the development of H2S-releasing scaffolds tailored for bone regeneration and would greatly impact the advancement in the field of BTE. Furthermore, the direct correlation between bone healing and local H2S release by bone scaffolds will be fundamental to corroborate the rationale of H2S’s role in mediating bone repair.
5. Concluding Remarks
This review provides a concise and up-to-date overview of the role of H2S in bone tissue repair and of the ongoing research and future challenges for the development of H2S-releasing scaffolds for regenerative purposes. The research of new formulations of H2S-releasing scaffolds for BTE may provide patients with skeletal injuries with novel opportunities of treatment.
Author Contributions
Manuscript drafted and revised by F.G., L.G. and B.G.
Funding
This study was supported by grant “Ricerca Finalizzata” from the Italian Ministry of Health (RF PE-2011-02348395, “Novel approach for bone regeneration and repair using sulfur donor-based therapy”).
Conflicts of Interest
The authors declare no conflict of interest.
The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Williamson, M.B.; Fromm, H.J. The Incorporation of Sulfur Amino Acids into the Proteins of Regenerating Wound Tissue. J. Biol. Chem. 1955, 212, 705–712. [Google Scholar]
- Abe, K.; Kimura, H. The Possible Role of Hydrogen Sulfide as an Endogenous Neuromodulator. J. Neurosci. 1996, 16, 1066–1071. [Google Scholar] [CrossRef] [PubMed]
- Sbodio, J.I.; Snyder, S.H.; Paul, B.D. Regulators of the Transsulfuration Pathway. Br. J. Pharmacol. 2019, 176, 583–593. [Google Scholar] [CrossRef] [PubMed]
- Szabo, C.; Papapetropoulos, A. International Union of Basic and Clinical Pharmacology. CII: Pharmacological Modulation of H2S Levels: H2S Donors and H2S Biosynthesis Inhibitors. Pharmacol. Rev. 2017, 69, 497–564. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.T.; Chen, L.; Xu, S.; Day, J.J.; Li, X.; Xian, M. Recent Development of Hydrogen Sulfide Releasing/Stimulating Reagents and Their Potential Applications in Cancer and Glycometabolic Disorders. Front. Pharmacol. 2017, 8, 664. [Google Scholar] [CrossRef] [Green Version]
- Grabowski, P. Physiology of Bone. Endocr. Dev. 2015, 28, 33–55. [Google Scholar] [PubMed]
- Guntur, A.R.; Rosen, C.J. Bone as an Endocrine Organ. Endocr. Pract. 2012, 18, 758–762. [Google Scholar] [CrossRef] [PubMed]
- Hadjidakis, D.J.; Androulakis, I.I. Bone Remodeling. Ann. N. Y. Acad. Sci. 2006, 1092, 385–396. [Google Scholar] [CrossRef] [PubMed]
- Martin, T.J.; Seeman, E. Bone Remodelling: Its Local Regulation and the Emergence of Bone Fragility. Best Pract. Res. Clin. Endocrinol. Metab. 2008, 22, 701–722. [Google Scholar] [CrossRef]
- Sommerfeldt, D.W.; Rubin, C.T. Biology of Bone and How It Orchestrates the Form and Function of the Skeleton. Eur. Spine J. 2001, 10 (Suppl. 2), S86–S95. [Google Scholar]
- Winkler, T.; Sass, F.A.; Duda, G.N.; Schmidt-Bleek, K. A Review of Biomaterials in Bone Defect Healing, Remaining Shortcomings and Future Opportunities for Bone Tissue Engineering: The Unsolved Challenge. Bone Jt. Res. 2018, 7, 232–243. [Google Scholar] [CrossRef] [PubMed]
- Nauth, A.; Schemitsch, E.; Norris, B.; Nollin, Z.; Watson, J.T. Critical-Size Bone Defects: Is There a Consensus for Diagnosis and Treatment? J. Orthop. Trauma 2018, 32 (Suppl. 1), S7–S11. [Google Scholar] [CrossRef] [PubMed]
- Curry, A.S.; Pensa, N.W.; Barlow, A.M.; Bellis, S.L. Taking Cues from the Extracellular Matrix to Design Bone-Mimetic Regenerative Scaffolds. Matrix Biol. 2016, 52–54, 397–412. [Google Scholar] [CrossRef] [PubMed]
- Grassi, F.; Tyagi, A.M.; Calvert, J.W.; Gambari, L.; Walker, L.D.; Yu, M.; Robinson, J.; Li, J.Y.; Lisignoli, G.; Vaccaro, C.; et al. Hydrogen Sulfide Is a Novel Regulator of Bone Formation Implicated in the Bone Loss Induced by Estrogen Deficiency. J. Bone Miner. Res. 2016, 31, 949–963. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Yang, R.; Liu, X.; Zhou, Y.; Qu, C.; Kikuiri, T.; Wang, S.; Zandi, E.; Du, J.; Ambudkar, I.S.; et al. Hydrogen Sulfide Maintains Mesenchymal Stem Cell Function and Bone Homeostasis Via Regulation of Ca(2+) Channel Sulfhydration. Cell Stem Cell 2014, 15, 66–78. [Google Scholar] [CrossRef] [PubMed]
- Behera, J.; Kelly, K.E.; Voor, M.J.; Metreveli, N.; Tyagi, S.C.; Tyagi, N. Hydrogen Sulfide Promotes Bone Homeostasis by Balancing Inflammatory Cytokine Signaling in Cbs-Deficient Mice through an Epigenetic Mechanism. Sci. Rep. 2018, 8, 15226. [Google Scholar] [CrossRef] [PubMed]
- Ozcivici, E.; Luu, Y.K.; Adler, B.; Qin, Y.X.; Rubin, J.; Judex, S.; Rubin, C.T. Mechanical Signals as Anabolic Agents in Bone. Nat. Rev. Rheumatol. 2010, 6, 50–59. [Google Scholar] [CrossRef]
- Marsell, R.; Einhorn, T.A. The Biology of Fracture Healing. Injury 2011, 42, 551–555. [Google Scholar] [CrossRef]
- Colnot, C.; Thompson, Z.; Miclau, T.; Werb, Z.; Helms, J.A. Altered Fracture Repair in the Absence of Mmp9. Development 2003, 130, 4123–4133. [Google Scholar] [CrossRef]
- Perez, J.R.; Kouroupis, D.; Li, D.J.; Best, T.M.; Kaplan, L.; Correa, D. Tissue Engineering and Cell-Based Therapies for Fractures and Bone Defects. Front. Bioeng. Biotechnol. 2018, 6, 105. [Google Scholar] [CrossRef] [Green Version]
- Hak, D.J.; Fitzpatrick, D.; Bishop, J.A.; Marsh, J.L.; Tilp, S.; Schnettler, R.; Simpson, H.; Alt, V. Delayed Union and Nonunions: Epidemiology, Clinical Issues, and Financial Aspects. Injury 2014, 45 (Suppl. 2), S3–S7. [Google Scholar] [CrossRef] [PubMed]
- Verrier, S.; Alini, M.; Alsberg, E.; Buchman, S.R.; Kelly, D.; Laschke, M.W.; Menger, M.D.; Murphy, W.L.; Stegemann, J.P.; Schutz, M.; et al. Tissue Engineering and Regenerative Approaches to Improving the Healing of Large Bone Defects. Eur. Cell Mater. 2016, 32, 87–110. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, G.; Moghaddam, A. Allograft Bone Matrix Versus Synthetic Bone Graft Substitutes. Injury 2011, 42 (Suppl. 2), S16–S21. [Google Scholar] [CrossRef]
- Baldwin, P.; Li, D.J.; Auston, D.A.; Mir, H.S.; Yoon, R.S.; Koval, K.J. Autograft, Allograft, and Bone Graft Substitutes: Clinical Evidence and Indications for Use in the Setting of Orthopaedic Trauma Surgery. J. Orthop. Trauma 2019, 33, 203–213. [Google Scholar] [CrossRef] [PubMed]
- Calcei, J.G.; Rodeo, S.A. Orthobiologics for Bone Healing. Clin. Sports Med. 2019, 38, 79–95. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Lu, H.; Zhao, Y.; Luo, J.; Yang, L.; Liu, W.; He, Q. Functionalized Cell-Free Scaffolds for Bone Defect Repair Inspired by Self-Healing of Bone Fractures: A Review and New Perspectives. Mater. Sci. Eng. C Mater. Biol. Appl. 2019, 98, 1241–1251. [Google Scholar] [CrossRef]
- Lopes, D.; Martins-Cruz, C.; Oliveira, M.B.; Mano, J.F. Bone Physiology as Inspiration for Tissue Regenerative Therapies. Biomaterials 2018, 185, 240–275. [Google Scholar] [CrossRef]
- Bigham-Sadegh, A.; Oryan, A. Selection of Animal Models for Pre-Clinical Strategies in Evaluating the Fracture Healing, Bone Graft Substitutes and Bone Tissue Regeneration and Engineering. Connect. Tissue Res. 2015, 56, 175–194. [Google Scholar] [CrossRef]
- Peric, M.; Dumic-Cule, I.; Grcevic, D.; Matijasic, M.; Verbanac, D.; Paul, R.; Grgurevic, L.; Trkulja, V.; Bagi, C.M.; Vukicevic, S. The Rational Use of Animal Models in the Evaluation of Novel Bone Regenerative Therapies. Bone 2015, 70, 73–86. [Google Scholar] [CrossRef]
- Loi, F.; Cordova, L.A.; Pajarinen, J.; Lin, T.H.; Yao, Z.; Goodman, S.B. Inflammation, Fracture and Bone Repair. Bone 2016, 86, 119–130. [Google Scholar] [CrossRef]
- Raggatt, L.J.; Wullschleger, M.E.; Alexander, K.A.; Wu, A.C.; Millard, S.M.; Kaur, S.; Maugham, M.L.; Gregory, L.S.; Steck, R.; Pettit, A.R. Fracture Healing Via Periosteal Callus Formation Requires Macrophages for Both Initiation and Progression of Early Endochondral Ossification. Am. J. Pathol. 2014, 184, 3192–3204. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Tan, J.; Martino, M.M.; Lui, K.O. Regulatory T-Cells: Potential Regulator of Tissue Repair and Regeneration. Front. Immunol. 2018, 9, 585. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Yang, R.; Shi, S. Systemic Infusion of Mesenchymal Stem Cells Improves Cell-Based Bone Regeneration Via Upregulation of Regulatory T Cells. Tissue Eng. Part A 2015, 21, 498–509. [Google Scholar] [CrossRef] [PubMed]
- Colburn, N.T.; Zaal, K.J.; Wang, F.; Tuan, R.S. A Role for Gamma/Delta T Cells in a Mouse Model of Fracture Healing. Arthritis Rheum. 2009, 60, 1694–1703. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, S.; Shi, S. The Role of Recipient T Cells in Mesenchymal Stem Cell-Based Tissue Regeneration. Int. J. Biochem. Cell Biol. 2012, 44, 2044–2050. [Google Scholar] [CrossRef]
- Sadtler, K.; Allen, B.W.; Estrellas, K.; Housseau, F.; Pardoll, D.M.; Elisseeff, J.H. The Scaffold Immune Microenvironment: Biomaterial-Mediated Immune Polarization in Traumatic and Nontraumatic Applications. Tissue Eng. Part A 2017, 23, 1044–1053. [Google Scholar] [CrossRef]
- Hegde, A.; Bhatia, M. Hydrogen Sulfide in Inflammation: Friend or Foe? Inflamm. Allergy Drug Targets 2011, 10, 118–122. [Google Scholar] [CrossRef]
- Fox, B.; Schantz, J.T.; Haigh, R.; Wood, M.E.; Moore, P.K.; Viner, N.; Spencer, J.P.; Winyard, P.G.; Whiteman, M. Inducible Hydrogen Sulfide Synthesis in Chondrocytes and Mesenchymal Progenitor Cells: Is H2S a Novel Cytoprotective Mediator in the Inflamed Joint? J. Cell Mol. Med. 2012, 16, 896–910. [Google Scholar] [CrossRef]
- Gambari, L.; Lisignoli, G.; Cattini, L.; Manferdini, C.; Facchini, A.; Grassi, F. Sodium Hydrosulfide Inhibits the Differentiation of Osteoclast Progenitor Cells Via Nrf2-Dependent Mechanism. Pharmacol. Res. 2014, 87, 99–112. [Google Scholar] [CrossRef]
- Ezeriņa, D.; Takano, Y.; Hanaoka, K.; Urano, Y.; Dick, T.P. N-Acetyl Cysteine Functions as a Fast-Acting Antioxidant by Triggering Intracellular H2S and Sulfane Sulfur Production. Cell Chem. Biol. 2018, 25, 447–459. [Google Scholar] [CrossRef]
- Cao, S.; Zhu, P.; Yu, X.; Chen, J.; Li, J.; Yan, F.; Wang, L.; Yu, J.; Chen, G. Hydrogen Sulfide Attenuates Brain Edema in Early Brain Injury after Subarachnoid Hemorrhage in Rats: Possible Involvement of Mmp-9 Induced Blood-Brain Barrier Disruption and Aqp4 Expression. Neurosci. Lett. 2016, 621, 88–97. [Google Scholar] [CrossRef] [PubMed]
- Zanardo, R.C.; Brancaleone, V.; Distrutti, E.; Fiorucci, S.; Cirino, G.; Wallace, J.L. Hydrogen Sulfide Is an Endogenous Modulator of Leukocyte-Mediated Inflammation. FASEB J. 2006, 20, 2118–2120. [Google Scholar] [CrossRef] [PubMed]
- Miao, L.; Shen, X.; Whiteman, M.; Xin, H.; Shen, Y.; Xin, X.; Moore, P.K.; Zhu, Y.Z. Hydrogen Sulfide Mitigates Myocardial Infarction Via Promotion of Mitochondrial Biogenesis-Dependent M2 Polarization of Macrophages. Antioxid. Redox Signal. 2016, 25, 268–281. [Google Scholar] [CrossRef]
- Zhou, X.; Chu, X.; Xin, D.; Li, T.; Bai, X.; Qiu, J.; Yuan, H.; Liu, D.; Wang, D.; Wang, Z. L-Cysteine-Derived H2S Promotes Microglia M2 Polarization Via Activation of the Ampk Pathway in Hypoxia-Ischemic Neonatal Mice. Front. Mol. Neurosci. 2019, 12, 58. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Chen, A.; Zhou, Y.; Zheng, S.; Yang, Y.; An, Y.; Xu, K.; Wang, Q. Novel H2S-Releasing Hydrogel for Wound Repair Via in Situ Polarization of M2 Macrophages. Biomaterials 2019, 222, 119398. [Google Scholar] [CrossRef]
- Brancaleone, V.; Mitidieri, E.; Flower, R.J.; Cirino, G.; Perretti, M. Annexin A1 Mediates Hydrogen Sulfide Properties in the Control of Inflammation. J. Pharmacol. Exp. Ther. 2014, 351, 96–104. [Google Scholar] [CrossRef] [Green Version]
- Herrera, B.S.; Coimbra, L.S.; da Silva, A.R.; Teixeira, S.A.; Costa, S.K.; Wallace, J.L.; Spolidorio, L.C.; Muscara, M.N. The H2S-Releasing Naproxen Derivative, Atb-346, Inhibits Alveolar Bone Loss and Inflammation in Rats with Ligature-Induced Periodontitis. Med. Gas. Res. 2015, 5, 4. [Google Scholar] [CrossRef]
- Gugliandolo, E.; Fusco, R.; D’Amico, R.; Militi, A.; Oteri, G.; Wallace, J.L.; Di Paola, R.; Cuzzocrea, S. Anti-Inflammatory Effect of Atb-352, a H2S -Releasing Ketoprofen Derivative, on Lipopolysaccharide-Induced Periodontitis in Rats. Pharmacol. Res. 2018, 132, 220–231. [Google Scholar] [CrossRef]
- Xiao, Z.; Bonnard, T.; Shakouri-Motlagh, A.; Wylie, R.A.L.; Collins, J.; White, J.; Heath, D.E.; Hagemeyer, C.E.; Connal, L.A. Triggered and Tunable Hydrogen Sulfide Release from Photogenerated Thiobenzaldehydes. Chemistry 2017, 23, 11294–11300. [Google Scholar] [CrossRef]
- Benedetti, F.; Curreli, S.; Krishnan, S.; Davinelli, S.; Cocchi, F.; Scapagnini, G.; Gallo, R.C.; Zella, D. Anti-Inflammatory Effects of H2S During Acute Bacterial Infection: A Review. J. Transl. Med. 2017, 15, 100. [Google Scholar] [CrossRef]
- Miller, T.W.; Wang, E.A.; Gould, S.; Stein, E.V.; Kaur, S.; Lim, L.; Amarnath, S.; Fowler, D.H.; Roberts, D.D. Hydrogen Sulfide Is an Endogenous Potentiator of T Cell Activation. J. Biol. Chem. 2012, 287, 4211–4221. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Liu, Y.; Shi, S. Hydrogen Sulfide Regulates Homeostasis of Mesenchymal Stem Cells and Regulatory T Cells. J. Dent. Res. 2016, 95, 1445–1451. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Qu, C.; Zhou, Y.; Konkel, J.E.; Shi, S.; Liu, Y.; Chen, C.; Liu, S.; Liu, D.; Chen, Y.; et al. Hydrogen Sulfide Promotes Tet1- and Tet2-Mediated Foxp3 Demethylation to Drive Regulatory T Cell Differentiation and Maintain Immune Homeostasis. Immunity 2015, 43, 251–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akter, F. The Role of Hydrogen Sulfide in Burns. Burns 2016, 42, 519–525. [Google Scholar] [CrossRef]
- Zhao, H.; Lu, S.; Chai, J.; Zhang, Y.; Ma, X.; Chen, J.; Guan, Q.; Wan, M.; Liu, Y. Hydrogen Sulfide Improves Diabetic Wound Healing in Ob/Ob Mice Via Attenuating Inflammation. J. Diabetes Complicat. 2017, 31, 1363–1369. [Google Scholar] [CrossRef]
- Wang, G.; Li, W.; Chen, Q.; Jiang, Y.; Lu, X.; Zhao, X. Hydrogen Sulfide Accelerates Wound Healing in Diabetic Rats. Int. J. Clin. Exp. Pathol. 2015, 8, 5097–5104. [Google Scholar]
- Seebach, C.; Henrich, D.; Wilhelm, K.; Barker, J.H.; Marzi, I. Endothelial Progenitor Cells Improve Directly and Indirectly Early Vascularization of Mesenchymal Stem Cell-Driven Bone Regeneration in a Critical Bone Defect in Rats. Cell Transplant. 2012, 21, 1667–1677. [Google Scholar] [CrossRef] [Green Version]
- Briasoulis, A.; Tousoulis, D.; Antoniades, C.; Papageorgiou, N.; Stefanadis, C. The Role of Endothelial Progenitor Cells in Vascular Repair after Arterial Injury and Atherosclerotic Plaque Development. Cardiovasc. Ther. 2011, 29, 125–139. [Google Scholar] [CrossRef]
- Ullah, M.; Liu, D.D.; Thakor, A.S. Mesenchymal Stromal Cell Homing: Mechanisms and Strategies for Improvement. iScience 2019, 15, 421–438. [Google Scholar] [CrossRef] [Green Version]
- Pankajakshan, D.; Agrawal, D.K. Mesenchymal Stem Cell Paracrine Factors in Vascular Repair and Regeneration. J. Biomed. Technol. Res. 2014, 1. [Google Scholar] [CrossRef] [Green Version]
- Hu, Q.; Ke, X.; Zhang, T.; Chen, Y.; Huang, Q.; Deng, B.; Xie, S.; Wang, J.; Nie, R. Hydrogen Sulfide Improves Vascular Repair by Promoting Endothelial Nitric Oxide Synthase-Dependent Mobilization of Endothelial Progenitor Cells. J. Hypertens. 2019, 37, 972–984. [Google Scholar] [CrossRef] [PubMed]
- Ke, X.; Zou, J.; Hu, Q.; Wang, X.; Hu, C.; Yang, R.; Liang, J.; Shu, X.; Nie, R.; Peng, C. Hydrogen Sulfide-Preconditioning of Human Endothelial Progenitor Cells Transplantation Improves Re-Endothelialization in Nude Mice with Carotid Artery Injury. Cell Physiol. Biochem. 2017, 43, 308–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, Z.; Garikipati, V.N.; Nickoloff, E.; Wang, C.; Polhemus, D.J.; Zhou, J.; Benedict, C.; Khan, M.; Verma, S.K.; Rabinowitz, J.E.; et al. Restoration of Hydrogen Sulfide Production in Diabetic Mice Improves Reparative Function of Bone Marrow Cells. Circulation 2016, 134, 1467–1483. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Hua, Y.; Qi, Y.; Meng, G.; Yang, S. Exogenous Hydrogen Sulphide Supplement Accelerates Skin Wound Healing Via Oxidative Stress Inhibition and Vascular Endothelial Growth Factor Enhancement. Exp. Dermatol. 2019, 28, 776–785. [Google Scholar] [CrossRef] [PubMed]
- Miao, L.; Xin, X.; Xin, H.; Shen, X.; Zhu, Y.Z. Hydrogen Sulfide Recruits Macrophage Migration by Integrin Beta1-Src-Fak/Pyk2-Rac Pathway in Myocardial Infarction. Sci. Rep. 2016, 6, 22363. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Li, C.S.; Wang, C.M.; Xie, Y.J.; Wang, A.L. Cse/H2S System Protects Mesenchymal Stem Cells from Hypoxia and Serum Deprivationinduced Apoptosis Via Mitochondrial Injury, Endoplasmic Reticulum Stress and Pi3k/Akt Activation Pathways. Mol. Med. Rep. 2015, 12, 2128–2134. [Google Scholar] [CrossRef]
- Zhao, Y.; Wei, H.; Kong, G.; Shim, W.; Zhang, G. Hydrogen Sulfide Augments the Proliferation and Survival of Human Induced Pluripotent Stem Cell-Derived Mesenchymal Stromal Cells through Inhibition of Bkca. Cytotherapy 2013, 15, 1395–1405. [Google Scholar] [CrossRef]
- Bahney, C.S.; Zondervan, R.L.; Allison, P.; Theologis, A.; Ashley, J.W.; Ahn, J.; Miclau, T.; Marcucio, R.S.; Hankenson, K.D. Cellular Biology of Fracture Healing. J. Orthop. Res. 2019, 37, 35–50. [Google Scholar] [CrossRef]
- Einhorn, T.A.; Gerstenfeld, L.C. Fracture Healing: Mechanisms and Interventions. Nat. Rev. Rheumatol. 2015, 11, 45–54. [Google Scholar] [CrossRef]
- Utvag, S.E.; Grundnes, O.; Rindal, D.B.; Reikeras, O. Influence of Extensive Muscle Injury on Fracture Healing in Rat Tibia. J. Orthop. Trauma 2003, 17, 430–435. [Google Scholar] [CrossRef]
- Hu, K.; Olsen, B.R. Osteoblast-Derived Vegf Regulates Osteoblast Differentiation and Bone Formation During Bone Repair. J. Clin. Investig. 2016, 126, 509–526. [Google Scholar] [CrossRef]
- Papapetropoulos, A.; Pyriochou, A.; Altaany, Z.; Yang, G.; Marazioti, A.; Zhou, Z.; Jeschke, M.G.; Branski, L.K.; Herndon, D.N.; Wang, R.; et al. Hydrogen Sulfide Is an Endogenous Stimulator of Angiogenesis. Proc. Natl. Acad. Sci. USA 2009, 106, 21972–21977. [Google Scholar] [CrossRef] [PubMed]
- Cai, W.J.; Wang, M.J.; Moore, P.K.; Jin, H.M.; Yao, T.; Zhu, Y.C. The Novel Proangiogenic Effect of Hydrogen Sulfide Is Dependent on Akt Phosphorylation. Cardiovasc. Res. 2007, 76, 29–40. [Google Scholar] [CrossRef] [PubMed]
- Katsouda, A.; Bibli, S.I.; Pyriochou, A.; Szabo, C.; Papapetropoulos, A. Regulation and Role of Endogenously Produced Hydrogen Sulfide in Angiogenesis. Pharmacol. Res. 2016, 113, 175–185. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.J.; Cai, W.J.; Zhu, Y.C. Mechanisms of Angiogenesis: Role of Hydrogen Sulphide. Clin. Exp. Pharmacol. Physiol. 2010, 37, 764–771. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.J.; Cai, W.J.; Li, N.; Ding, Y.J.; Chen, Y.; Zhu, Y.C. The Hydrogen Sulfide Donor Nahs Promotes Angiogenesis in a Rat Model of Hind Limb Ischemia. Antioxid. Redox Signal. 2010, 12, 1065–1077. [Google Scholar] [CrossRef]
- Gambari, L.; Amore, E.; Raggio, R.; Bonani, W.; Barone, M.; Lisignoli, G.; Grigolo, B.; Motta, A.; Grassi, F. Hydrogen Sulfide-Releasing Silk Fibroin Scaffold for Bone Tissue Engineering. Mater. Sci. Eng. C Mater. Biol. Appl. 2019, 102, 471–482. [Google Scholar] [CrossRef]
- Liu, X.; Pan, L.; Zhuo, Y.; Gong, Q.; Rose, P.; Zhu, Y. Hypoxia-Inducible Factor-1alpha Is Involved in the Pro-Angiogenic Effect of Hydrogen Sulfide under Hypoxic Stress. Biol. Pharm. Bull. 2010, 33, 1550–1554. [Google Scholar] [CrossRef]
- Majumder, A.; Singh, M.; George, A.K.; Behera, J.; Tyagi, N.; Tyagi, S.C. Hydrogen Sulfide Improves Postischemic Neoangiogenesis in the Hind Limb of Cystathionine-Beta-Synthase Mutant Mice Via Ppar-Gamma/Vegf Axis. Physiol. Rep. 2018, 6, e13858. [Google Scholar] [CrossRef]
- Gambari, L.; Lisignoli, G.; Gabusi, E.; Manferdini, C.; Paolella, F.; Piacentini, A.; Grassi, F. Distinctive Expression Pattern of Cystathionine-Beta-Synthase and Cystathionine-Gamma-Lyase Identifies Mesenchymal Stromal Cells Transition to Mineralizing Osteoblasts. J. Cell. Physiol. 2017, 232, 3574–3585. [Google Scholar] [CrossRef]
- Zheng, Y.; Liao, F.; Lin, X.; Zheng, F.; Fan, J.; Cui, Q.; Yang, J.; Geng, B.; Cai, J. Cystathionine Gamma-Lyase-Hydrogen Sulfide Induces Runt-Related Transcription Factor 2 Sulfhydration, Thereby Increasing Osteoblast Activity to Promote Bone Fracture Healing. Antioxid. Redox Signal. 2017, 27, 742–753. [Google Scholar] [CrossRef] [PubMed]
- Lambertini, E.; Penolazzi, L.; Angelozzi, M.; Grassi, F.; Gambari, L.; Lisignoli, G.; de Bonis, P.; Cavallo, M.; Piva, R. The Expression of Cystathionine Gamma-Lyase Is Regulated by Estrogen Receptor Alpha in Human Osteoblasts. Oncotarget 2017, 8, 101686–101696. [Google Scholar] [CrossRef] [PubMed]
- Zhai, Y.; Behera, J.; Tyagi, S.C.; Tyagi, N. Hydrogen Sulfide Attenuates Homocysteine-Induced Osteoblast Dysfunction by Inhibiting Mitochondrial Toxicity. J. Cell. Physiol. 2019, 234, 18602–18614. [Google Scholar] [CrossRef] [PubMed]
- Lv, M.; Liu, Y.; Xiao, T.H.; Jiang, W.; Lin, B.W.; Zhang, X.M.; Lin, Y.M.; Xu, Z.S. Gyy4137 Stimulates Osteoblastic Cell Proliferation and Differentiation Via an Erk1/2-Dependent Anti-Oxidant Mechanism. Am. J. Transl. Res. 2017, 9, 1183–1192. [Google Scholar] [PubMed]
- Mo, S.; Hua, Y. Cystathionine Gamma Lyase-H2S Contributes to Osteoclastogenesis During Bone Remodeling Induced by Mechanical Loading. Biochem. Biophys. Res. Commun. 2018, 501, 471–477. [Google Scholar] [CrossRef]
- Liu, F.; Wen, F.; He, D.; Liu, D.; Yang, R.; Wang, X.; Yan, Y.; Liu, Y.; Kou, X.; Zhou, Y. Force-Induced H2S by Pdlscs Modifies Osteoclastic Activity During Tooth Movement. J. Dent. Res. 2017, 96, 694–702. [Google Scholar] [CrossRef]
- Lee, S.K.; Chung, J.H.; Choi, S.C.; Auh, Q.S.; Lee, Y.M.; Lee, S.I.; Kim, E.C. Sodium Hydrogen Sulfide Inhibits Nicotine and Lipopolysaccharide-Induced Osteoclastic Differentiation and Reversed Osteoblastic Differentiation in Human Periodontal Ligament Cells. J. Cell Biochem. 2013, 114, 1183–1193. [Google Scholar] [CrossRef]
- Cen, S.D.; Yu, W.B.; Ren, M.M.; Chen, L.J.; Sun, C.F.; Ye, Z.L.; Deng, H.; Hu, R.D. Endogenous Hydrogen Sulfide Is Involved in Osteogenic Differentiation in Human Periodontal Ligament Cells. Arch. Oral. Biol. 2016, 68, 1–8. [Google Scholar] [CrossRef]
- Jiang, Z.; Hua, Y. Hydrogen Sulfide Promotes Osteogenic Differentiation of Human Periodontal Ligament Cells Via P38-Mapk Signaling Pathway under Proper Tension Stimulation. Arch. Oral. Biol. 2016, 72, 8–13. [Google Scholar] [CrossRef]
- Pu, H.; Hua, Y. Hydrogen Sulfide Regulates Bone Remodeling and Promotes Orthodontic Tooth Movement. Mol. Med. Rep. 2017, 16, 9415–9422. [Google Scholar] [CrossRef]
- Xu, Z.S.; Dai, F.; Chen, J.; Lv, M.; Cheng, J.W.; Zhang, X.M.; Lin, B.W. Experimental Research into the Potential Therapeutic Effect of Gyy4137 on Ovariectomy-Induced Osteoporosis. Cell. Mol. Biol. Lett. 2018, 23, 47. [Google Scholar] [CrossRef] [PubMed]
- Behera, J.; George, A.K.; Voor, M.J.; Tyagi, S.C.; Tyagi, N. Hydrogen Sulfide Epigenetically Mitigates Bone Loss through Opg/Rankl Regulation During Hyperhomocysteinemia in Mice. Bone 2018, 114, 90–108. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.H.; Lewis, J.L.; Lin, H.S.; Kuo, L.T.; Mao, S.W.; Tai, Y.S.; Chang, M.S.; Ables, G.P.; Perrone, C.E.; Yang, R.S. A Methionine-Restricted Diet and Endurance Exercise Decrease Bone Mass and Extrinsic Strength but Increase Intrinsic Strength in Growing Male Rats. J. Nutr. 2014, 144, 621–630. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Hao, D.; Zhang, H.; Liu, B.; Yang, M.; He, B. Treatment with Hydrogen Sulfide Attenuates Sublesional Skeletal Deterioration Following Motor Complete Spinal Cord Injury in Rats. Osteoporos. Int. 2017, 28, 687–695. [Google Scholar] [CrossRef]
- Jiang, X.; Chen, Y.; Lu, K.; Zhang, H.; Fan, X. Gyy4137 Promotes Bone Formation in a Rabbit Distraction Osteogenesis Model: A Preliminary Report. J. Oral Maxillofac. Surg. 2015, 73, 732. [Google Scholar] [CrossRef]
- Jiang, X.W.; Zhang, Y.; Cheng, Y.Z.; Fan, X.S.; Deng, X.; Peng, H.Y. The Expression of Endogenous Hydrogen Sulfide Signal During Distraction Osteogenesis in a Rabbit Model. Int. J. Oral Maxillofac. Surg. 2018, 47, 262–267. [Google Scholar] [CrossRef]
- Hu, K.; Olsen, B.R. The Roles of Vascular Endothelial Growth Factor in Bone Repair and Regeneration. Bone 2016, 91, 30–38. [Google Scholar] [CrossRef]
- Miclau, K.R.; Brazina, S.A.; Bahney, C.S.; Hankenson, K.D.; Hunt, T.K.; Marcucio, R.S.; Miclau, T. Stimulating Fracture Healing in Ischemic Environments: Does Oxygen Direct Stem Cell Fate During Fracture Healing? Front. Cell Dev. Biol. 2017, 5, 45. [Google Scholar] [CrossRef]
- Runyan, C.M.; Gabrick, K.S. Biology of Bone Formation, Fracture Healing, and Distraction Osteogenesis. J. Craniofac. Surg. 2017, 28, 1380–1389. [Google Scholar] [CrossRef]
- Coates, B.A.; McKenzie, J.A.; Buettmann, E.G.; Liu, X.; Gontarz, P.M.; Zhang, B.; Silva, M.J. Transcriptional Profiling of Intramembranous and Endochondral Ossification after Fracture in Mice. Bone 2019, 127, 577–591. [Google Scholar] [CrossRef]
- Gerstenfeld, L.C.; Cullinane, D.M.; Barnes, G.L.; Graves, D.T.; Einhorn, T.A. Fracture Healing as a Post-Natal Developmental Process: Molecular, Spatial, and Temporal Aspects of Its Regulation. J. Cell Biochem. 2003, 88, 873–884. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.K.; Kim, H.N.; Bhang, S.H.; Shin, J.Y.; Han, J.; La, W.G.; Jeong, G.J.; Kang, S.; Lee, J.R.; Oh, J.; et al. Enhanced Bone Repair by Guided Osteoblast Recruitment Using Topographically Defined Implant. Tissue Eng. Part A 2016, 22, 654–664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, M.S.; Lee, D.H.; Jeon, J.; Oh, S.H.; Yang, H.S. Topographically Defined, Biodegradable Nanopatterned Patches to Regulate Cell Fate and Acceleration of Bone Regeneration. ACS Appl. Mater. Interfaces 2018, 10, 38780–38790. [Google Scholar] [CrossRef] [PubMed]
- Marie, P.J.; Ammann, P.; Boivin, G.; Rey, C. Mechanisms of Action and Therapeutic Potential of Strontium in Bone. Calcified Tissue Int. 2001, 69, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Zreiqat, H.; Howlett, C.R.; Zannettino, A.; Evans, P.; Schulze-Tanzil, G.; Knabe, C.; Shakibaei, M. Mechanisms of Magnesium-Stimulated Adhesion of Osteoblastic Cells to Commonly Used Orthopaedic Implants. J. Biomed. Mater. Res. 2002, 62, 175–184. [Google Scholar] [CrossRef]
- Quinlan, E.; Partap, S.; Azevedo, M.M.; Jell, G.; Stevens, M.M.; O’Brien, F.J. Hypoxia-Mimicking Bioactive Glass/Collagen Glycosaminoglycan Composite Scaffolds to Enhance Angiogenesis and Bone Repair. Biomaterials 2015, 52, 358–366. [Google Scholar] [CrossRef] [PubMed]
- Carlisle, E.M. Silicon: A Requirement in Bone Formation Independent of Vitamin D1. Calcified Tissue Int. 1981, 33, 27–34. [Google Scholar] [CrossRef]
- Wu, J.; Li, Y.; He, C.; Kang, J.; Ye, J.; Xiao, Z.; Zhu, J.; Chen, A.; Feng, S.; Li, X.; et al. Novel H2S Releasing Nanofibrous Coating for In Vivo Dermal Wound Regeneration. ACS Appl. Mater. Interfaces 2016, 8, 27474–27481. [Google Scholar] [CrossRef]
- Goren, I.; Kohler, Y.; Aglan, A.; Pfeilschifter, J.; Beck, K.F.; Frank, S. Increase of Cystathionine-Gamma-Lyase (Cse) During Late Wound Repair: Hydrogen Sulfide Triggers Cytokeratin 10 Expression in Keratinocytes. Nitric Oxide 2019, 87, 31–42. [Google Scholar] [CrossRef]
- Magierowski, M.; Magierowska, K.; Hubalewska-Mazgaj, M.; Surmiak, M.; Sliwowski, Z.; Wierdak, M.; Kwiecien, S.; Chmura, A.; Brzozowski, T. Cross-Talk between Hydrogen Sulfide and Carbon Monoxide in the Mechanism of Experimental Gastric Ulcers Healing, Regulation of Gastric Blood Flow and Accompanying Inflammation. Biochem. Pharmacol. 2018, 149, 131–142. [Google Scholar] [CrossRef]
- Andreu, V.; Mendoza, G.; Arruebo, M.; Irusta, S. Smart Dressings Based on Nanostructured Fibers Containing Natural Origin Antimicrobial, Anti-Inflammatory, and Regenerative Compounds. Materials 2015, 8, 5154–5193. [Google Scholar] [CrossRef] [PubMed]
- Cacciotti, I.; Ciocci, M.; di Giovanni, E.; Nanni, F.; Melino, S. Hydrogen Sulfide-Releasing Fibrous Membranes: Potential Patches for Stimulating Human Stem Cells Proliferation and Viability under Oxidative Stress. Int. J. Mol. Sci. 2018, 19, 2368. [Google Scholar] [CrossRef] [PubMed]
- Feng, S.; Zhao, Y.; Xian, M.; Wang, Q. Biological Thiols-Triggered Hydrogen Sulfide Releasing Microfibers for Tissue Engineering Applications. Acta Biomater. 2015, 27, 205–213. [Google Scholar] [CrossRef]
- Mauretti, A.; Neri, A.; Kossover, O.; Seliktar, D.; Nardo, P.D.; Melino, S. Design of a Novel Composite H2S-Releasing Hydrogel for Cardiac Tissue Repair. Macromol. Biosci. 2016, 16, 847–858. [Google Scholar] [CrossRef] [PubMed]
- Abdelmonem, M.; Shahin, N.N.; Rashed, L.A.; Amin, H.A.A.; Shamaa, A.A.; Shaheen, A.A. Hydrogen Sulfide Enhances the Effectiveness of Mesenchymal Stem Cell Therapy in Rats with Heart Failure: In Vitro Preconditioning Versus In Vivo Co-Delivery. Biomed. Pharmacother. 2019, 112, 108584. [Google Scholar] [CrossRef]
- Liang, W.; Chen, J.; Li, L.; Li, M.; Wei, X.; Tan, B.; Shang, Y.; Fan, G.; Wang, W.; Liu, W. Conductive Hydrogen Sulfide-Releasing Hydrogel Encapsulating Adscs for Myocardial Infarction Treatment. ACS Appl. Mater. Interfaces 2019, 11, 14619–14629. [Google Scholar] [CrossRef]
- Carter, J.M.; Qian, Y.; Foster, J.C.; Matson, J.B. Peptide-Based Hydrogen Sulphide-Releasing Gels. Chem. Commun. 2015, 51, 13131–13134. [Google Scholar] [CrossRef]
- Longchamp, A.; Kaur, K.; Macabrey, D.; Dubuis, C.; Corpataux, J.S.; Deglise, M.; Matson, J.B.; Allagnat, F. Hydrogen Sulfide-Releasing Peptide Hydrogel Limits the Development of Intimal Hyperplasia in Human Vein Segments. Acta Biomater. 2019, 97, 374–384. [Google Scholar] [CrossRef]
- Shitole, A.A.; Raut, P.W.; Sharma, N.; Giram, P.; Khandwekar, A.P.; Garnaik, B. Electrospun Polycaprolactone/Hydroxyapatite/Zno Nanofibers as Potential Biomaterials for Bone Tissue Regeneration. J. Mater. Sci. Mater. Med. 2019, 30, 51. [Google Scholar] [CrossRef]
- Wu, Y.A.; Chiu, Y.C.; Lin, Y.H.; Ho, C.C.; Shie, M.Y.; Chen, Y.W. 3d-Printed Bioactive Calcium Silicate/Poly-Epsilon-Caprolactone Bioscaffolds Modified with Biomimetic Extracellular Matrices for Bone Regeneration. Int. J. Mol. Sci. 2019, 20, 942. [Google Scholar] [CrossRef]
- Fairag, R.; Rosenzweig, D.H.; Ramirez-Garcialuna, J.L.; Weber, M.H.; Haglund, L. Three-Dimensional Printed Polylactic Acid Scaffolds Promote Bone-Like Matrix Deposition In Vitro. ACS Appl. Mater. Interfaces 2019, 11, 15306–15315. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.Z.; You, M.L.; Ding, Z.Q.; Ye, W.B. A Review of Emerging Bone Tissue Engineering Via Peg Conjugated Biodegradable Amphiphilic Copolymers. Mater. Sci. Eng. C Mater. Biol. Appl. 2019, 97, 1021–1035. [Google Scholar] [CrossRef] [PubMed]
- Ma, D.; An, G.; Liang, M.; Liu, Y.; Zhang, B.; Wang, Y. A Composited Peg-Silk Hydrogel Combining with Polymeric Particles Delivering Rhbmp-2 for Bone Regeneration. Mater. Sci. Eng. C Mater. Biol. Appl. 2016, 65, 221–231. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.; Li, Z.; Organ, C.L.; Park, C.M.; Yang, C.T.; Pacheco, A.; Wang, D.; Lefer, D.J.; Xian, M. Ph-Controlled Hydrogen Sulfide Release for Myocardial Ischemia-Reperfusion Injury. J. Am. Chem. Soc. 2016, 138, 6336–6339. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.T.; Lai, Z.Z.; Zheng, Z.H.; Kang, J.M.; Xian, M.; Wang, R.Y.; Shi, K.; Meng, F.H.; Li, X.; Chen, L.; et al. A Novel Ph-Controlled Hydrogen Sulfide Donor Protects Gastric Mucosa from Aspirin-Induced Injury. J. Cell Mol. Med. 2017, 21, 2441–2451. [Google Scholar] [CrossRef]
- Zhao, Y.; Bhushan, S.; Yang, C.; Otsuka, H.; Stein, J.D.; Pacheco, A.; Peng, B.; Devarie-Baez, N.O.; Aguilar, H.C.; Lefer, D.J.; et al. Controllable Hydrogen Sulfide Donors and Their Activity against Myocardial Ischemia-Reperfusion Injury. ACS Chem. Biol. 2013, 8, 1283–1290. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Yang, C.; Organ, C.; Li, Z.; Bhushan, S.; Otsuka, H.; Pacheco, A.; Kang, J.; Aguilar, H.C.; Lefer, D.J.; et al. Design, Synthesis, and Cardioprotective Effects of N-Mercapto-Based Hydrogen Sulfide Donors. J. Med. Chem. 2015, 58, 7501–7511. [Google Scholar] [CrossRef]
- Zheng, Y.; Yu, B.; de la Cruz, L.K.; Choudhury, M.R.; Anifowose, A.; Wang, B. Toward Hydrogen Sulfide Based Therapeutics: Critical Drug Delivery and Developability Issues. Med. Res. Rev. 2018, 38, 57–100. [Google Scholar] [CrossRef]
- Zhao, Y.; Biggs, T.D.; Xian, M. Hydrogen Sulfide (H2S) Releasing Agents: Chemistry and Biological Applications. Chem. Commun. 2014, 50, 11788–11805. [Google Scholar] [CrossRef]
- Rapposelli, S.; Gambari, L.; Digiacomo, M.; Citi, V.; Lisignoli, G.; Manferdini, C.; Calderone, V.; Grassi, F. A Novel H2S-Releasing Amino-Bisphosphonate Which Combines Bone Anti-Catabolic and Anabolic Functions. Sci. Rep. 2017, 7, 11940. [Google Scholar] [CrossRef]
Figure 1.
Schematic representation of the process of bone fracture healing. Immediately after bone injury, the healing process is initiated by hematoma formation and inflammatory response accompanied by the release of pro-inflammatory cytokines. In direct bone healing (occurring at fractures that are perfectly aligned and with no vascular disruption) the inflammation activates a process of intramembranous ossification which directly produces a new lamellar bone structure without requiring any remodeling step. In indirect bone healing (occurring in fractures with large, hypoxic defects and interfragmentary strains), the intramembranous ossification response occurs subperiosteally directly adjacent to the ends of the fracture, generating a hard callus. Adjacent to the fracture line, endochondral ossification occurs: A cartilaginous callus form and several cells migrates and proliferate within it, building an intermediate cartilaginous template which subsequently undergoes calcification (primary bone formation); the cartilage and primary mineralized matrix are then resorbed and secondary bone formation occurs; ultimately bone remodeling by osteoclasts and osteoblasts take place and the callus is totally replaced by new bone indistinguishable from adjacent, uninjured tissue. The processes by which H2S may modulate during bone fracture healing are highlighted by a yellow arrow.
Figure 1.
Schematic representation of the process of bone fracture healing. Immediately after bone injury, the healing process is initiated by hematoma formation and inflammatory response accompanied by the release of pro-inflammatory cytokines. In direct bone healing (occurring at fractures that are perfectly aligned and with no vascular disruption) the inflammation activates a process of intramembranous ossification which directly produces a new lamellar bone structure without requiring any remodeling step. In indirect bone healing (occurring in fractures with large, hypoxic defects and interfragmentary strains), the intramembranous ossification response occurs subperiosteally directly adjacent to the ends of the fracture, generating a hard callus. Adjacent to the fracture line, endochondral ossification occurs: A cartilaginous callus form and several cells migrates and proliferate within it, building an intermediate cartilaginous template which subsequently undergoes calcification (primary bone formation); the cartilage and primary mineralized matrix are then resorbed and secondary bone formation occurs; ultimately bone remodeling by osteoclasts and osteoblasts take place and the callus is totally replaced by new bone indistinguishable from adjacent, uninjured tissue. The processes by which H2S may modulate during bone fracture healing are highlighted by a yellow arrow.

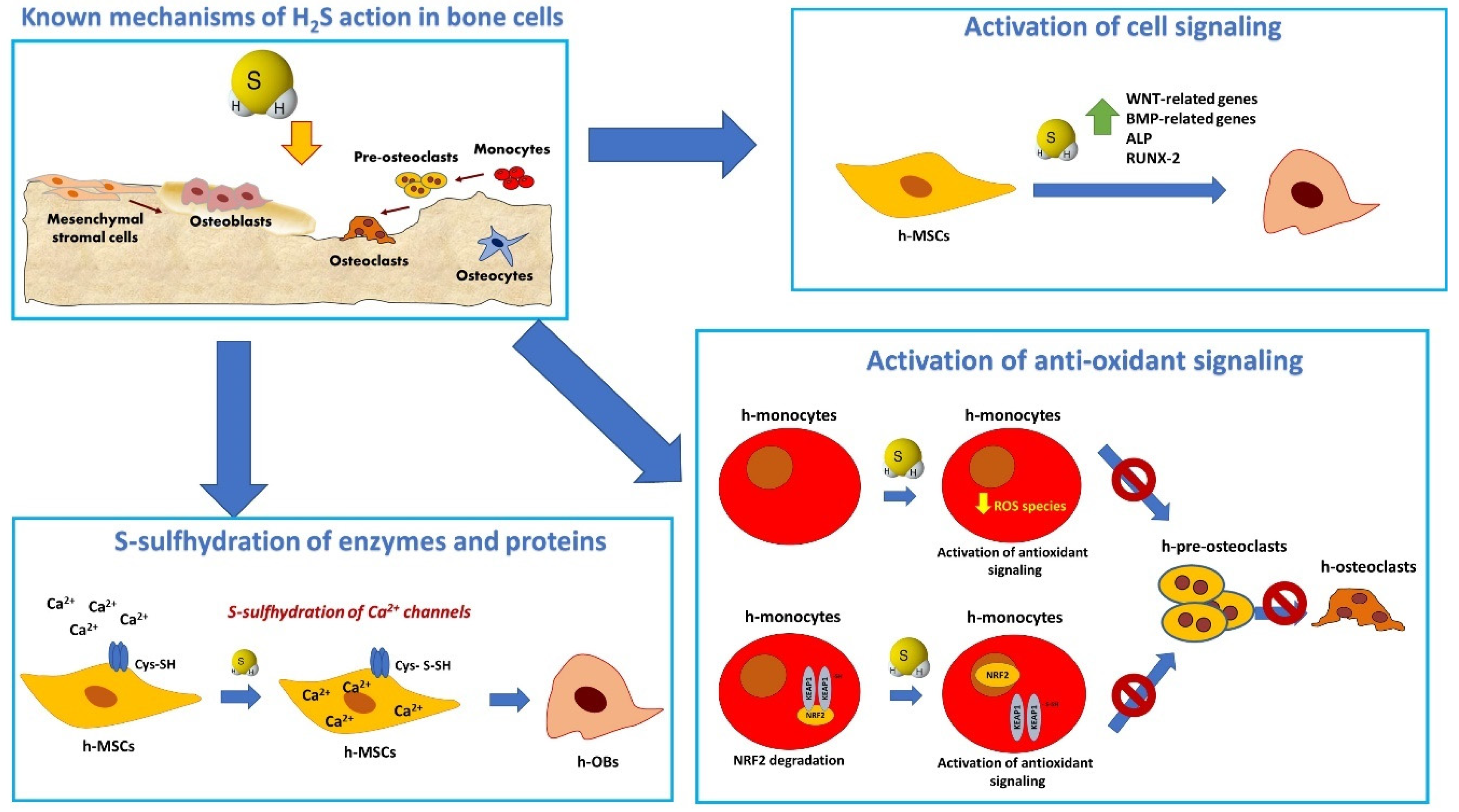
Figure 2.
Schematic representation of the main mechanisms of action of H2S in mesenchymal stromal cells, osteoblasts, and osteoclasts. H2S modulates osteogenic differentiation in osteoprogenitor cells by inducing S-sulfhydration in TRP channels leading to increased Ca2+ influx in MSC [15]; at the transcriptional level, H2S donors induce the expression of osteogenic signaling through stimulation of multiple signaling pathway [14,77,88]. In the context of inflammation, H2S triggers an antioxidant signaling in OC which involves the increased nuclear translocations of NRF-2 and results in the inhibition of OC differentiation [39,87].
Figure 2.
Schematic representation of the main mechanisms of action of H2S in mesenchymal stromal cells, osteoblasts, and osteoclasts. H2S modulates osteogenic differentiation in osteoprogenitor cells by inducing S-sulfhydration in TRP channels leading to increased Ca2+ influx in MSC [15]; at the transcriptional level, H2S donors induce the expression of osteogenic signaling through stimulation of multiple signaling pathway [14,77,88]. In the context of inflammation, H2S triggers an antioxidant signaling in OC which involves the increased nuclear translocations of NRF-2 and results in the inhibition of OC differentiation [39,87].

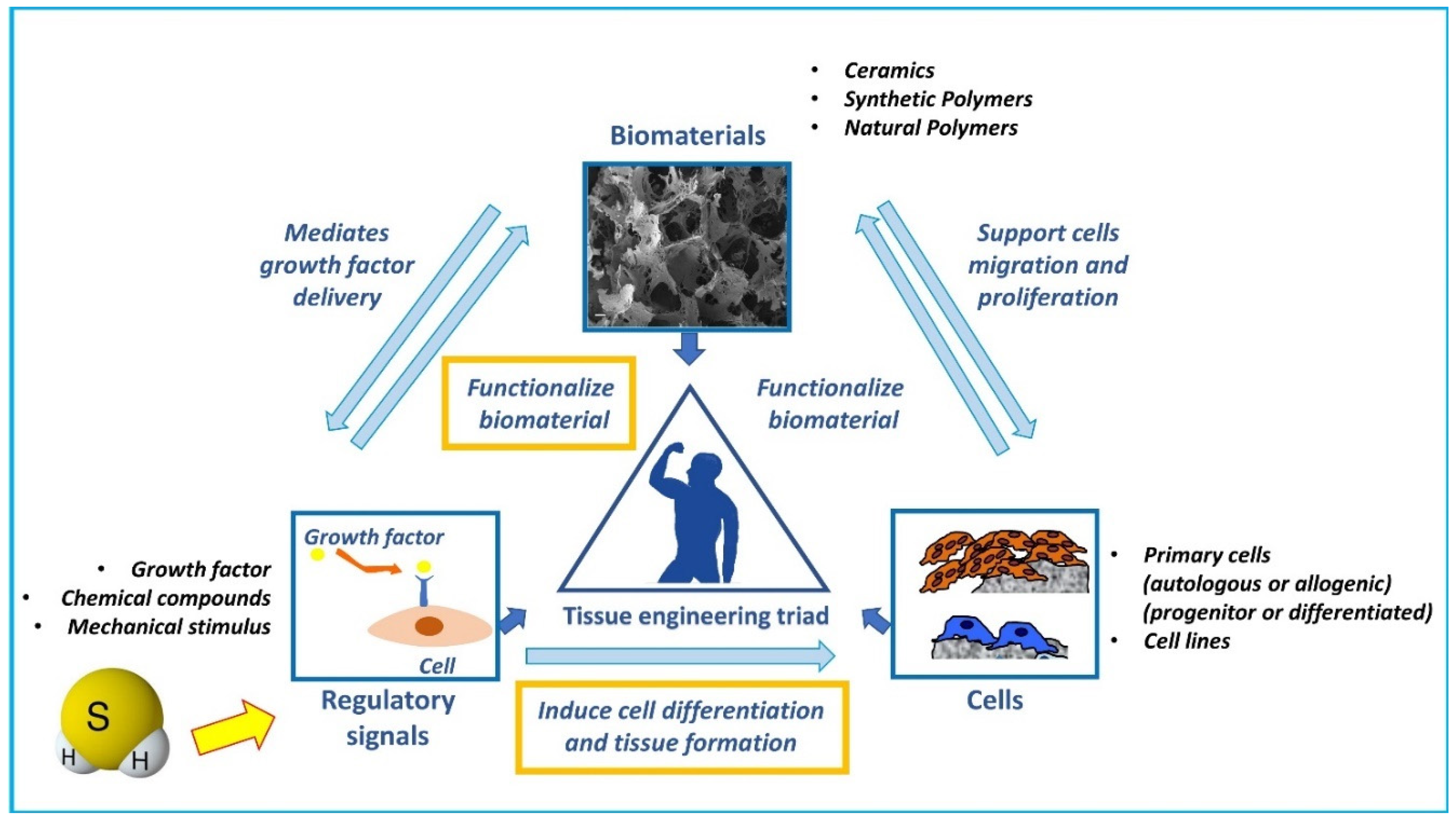
Figure 3.
Schematic representation of potential H2S role in the “tissue engineering triad”. Three main factors are the key players of tissue engineering strategies and compose the so-called “tissue engineering triad”: 1) a scaffold that provides structure and support cells migration, colonization, proliferation, and differentiation, and is the substrate of loading of regulatory signals such as growth factors; 2) a source of cells to promote the required tissue formation (endogenous cells, primary cells, or cell lines pre-cultured or loaded within the scaffold); and 3) regulatory signals (growth factors, biophysical stimuli, etc.) to induce cell differentiation and tissue formation. We postulate that H2S may be considered as a regulatory signal able to functionalize biomaterials and induce endogenous or loaded cells differentiation and promote bone tissue formation.
Figure 3.
Schematic representation of potential H2S role in the “tissue engineering triad”. Three main factors are the key players of tissue engineering strategies and compose the so-called “tissue engineering triad”: 1) a scaffold that provides structure and support cells migration, colonization, proliferation, and differentiation, and is the substrate of loading of regulatory signals such as growth factors; 2) a source of cells to promote the required tissue formation (endogenous cells, primary cells, or cell lines pre-cultured or loaded within the scaffold); and 3) regulatory signals (growth factors, biophysical stimuli, etc.) to induce cell differentiation and tissue formation. We postulate that H2S may be considered as a regulatory signal able to functionalize biomaterials and induce endogenous or loaded cells differentiation and promote bone tissue formation.

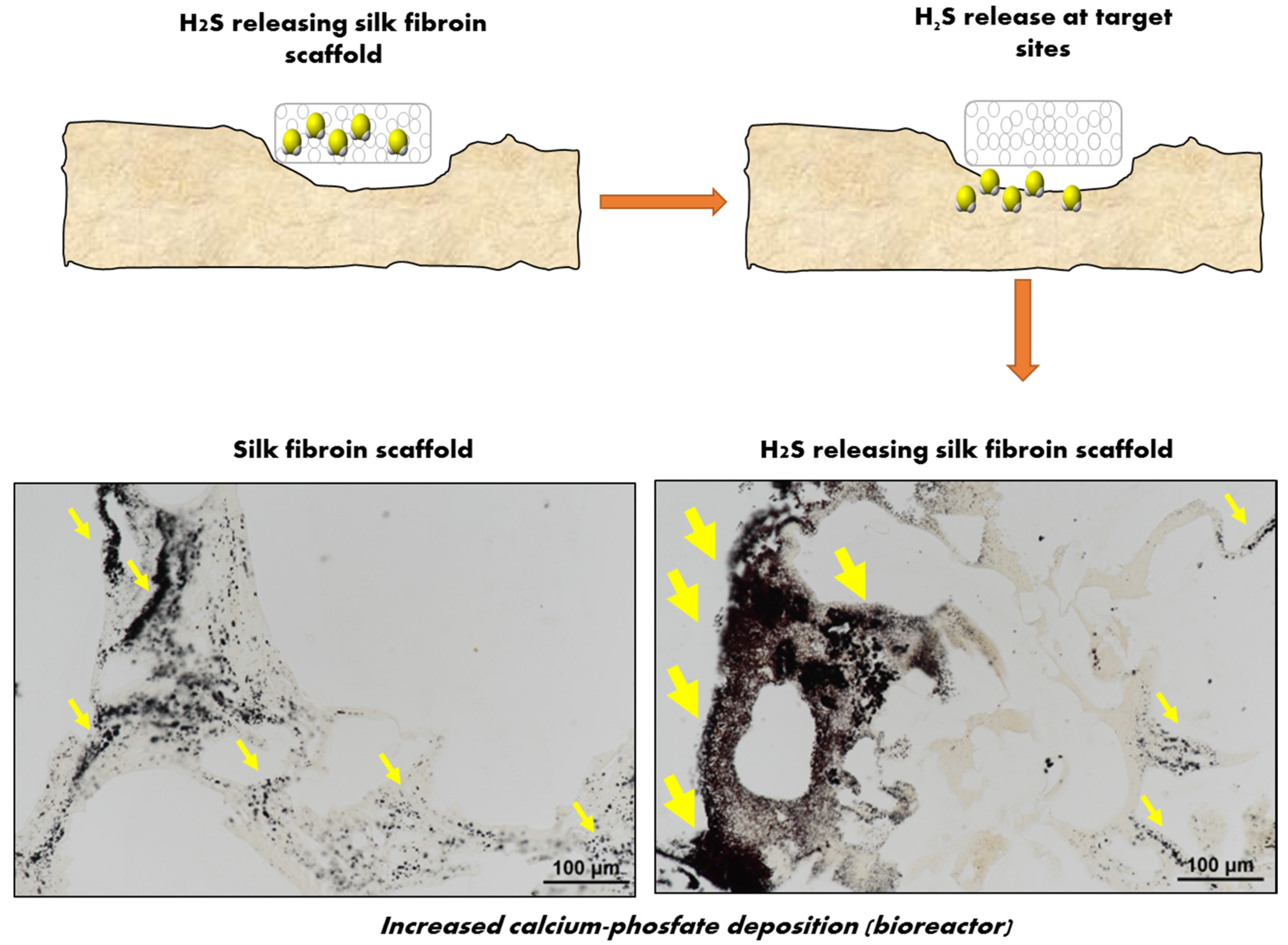
Figure 4.
Schematic representation of the rationale for developing H2S releasing scaffolds. A silk fibroin (SF) porous scaffold loaded with the H2S donor GYY4137 (GYY) was developed with the rationale of providing the anabolic function of H2S to the damaged bone tissue. The combination of SF with H2S potentiated the osteoconductive properties of SF when h-MSCs where cultured in osteogenic media on a perfusion bioreactor. Cells remained viable and colonized the scaffolds. H2S activated genes and proteins involved in ossification, OBs differentiation, bone mineral metabolism, and angiogenesis, allowing a high and early mineralization (as shown by Von Kossa staining in the figure, yellow arrows indicate areas with more intense staining for mineralized matrix).
Figure 4.
Schematic representation of the rationale for developing H2S releasing scaffolds. A silk fibroin (SF) porous scaffold loaded with the H2S donor GYY4137 (GYY) was developed with the rationale of providing the anabolic function of H2S to the damaged bone tissue. The combination of SF with H2S potentiated the osteoconductive properties of SF when h-MSCs where cultured in osteogenic media on a perfusion bioreactor. Cells remained viable and colonized the scaffolds. H2S activated genes and proteins involved in ossification, OBs differentiation, bone mineral metabolism, and angiogenesis, allowing a high and early mineralization (as shown by Von Kossa staining in the figure, yellow arrows indicate areas with more intense staining for mineralized matrix).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Summary of the main studies and key findings on the regulation of bone cell function and bone metabolism by H2S-releasing agents.
Table 1.
Summary of the main studies and key findings on the regulation of bone cell function and bone metabolism by H2S-releasing agents.
| Ref. | Authors | H2S-Donor | In Vitro/In Vivo | Species | Main Findings |
|---|---|---|---|---|---|
| [14] | Grassi et al. |
| In vivo In vitro | Mice |
|
| [15] | Liu et al. |
| In vivo | Mice |
|
| [16] | Behera et al. |
| In vivo | Mice |
|
| [75] | Gambari et al. | GYY4137 loaded in a SF scaffold | In vitro | Human |
|
| [81] | Zhai et al. | NaHS | In vitro | Murine cell line |
|
| [82] | Lv et al. | GYY4137 | In vitro | Murine cell line |
|
| [83] | Mo et al. | GYY4137 | In vivo | Mice |
|
| [86] | Lee et al. | NaHS | In vitro | Human Mice |
|
| [88] | Jiang et al. | NaHS | In vitro | Human |
|
| [89] | Pu et al. | NaHS | In vivo | Mice |
|
| [90] | Xu et al. | GYY4137 | In vivo | Rat |
|
| [91] | Behera et al. | NaHS | In vivo In vitro | Mice |
|
| [93] | Yang et al. | NaHS | In vivo | Rats |
|
| [94] | Jiang et al. | GYY4137 | In vivo | Rabbits |
|
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Gambari, L.; Grigolo, B.; Grassi, F. Hydrogen Sulfide in Bone Tissue Regeneration and Repair: State of the Art and New Perspectives. Int. J. Mol. Sci. 2019, 20, 5231. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20205231
AMA Style
Gambari L, Grigolo B, Grassi F. Hydrogen Sulfide in Bone Tissue Regeneration and Repair: State of the Art and New Perspectives. International Journal of Molecular Sciences. 2019; 20(20):5231. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20205231
Chicago/Turabian StyleGambari, Laura, Brunella Grigolo, and Francesco Grassi. 2019. "Hydrogen Sulfide in Bone Tissue Regeneration and Repair: State of the Art and New Perspectives" International Journal of Molecular Sciences 20, no. 20: 5231. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20205231
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.