Citrus Genetic Engineering for Disease Resistance: Past, Present and Future


1
Institute of Citrus Research, Zhejiang Academy of Agricultural Sciences, Taizhou 318026, China
2
National Center for Citrus Variety Improvement, Zhejiang Branch, Taizhou 318026, China
3
Department of Plant Biology and Ecology, College of Life Sciences, Nankai University, Tianjin 300071, China
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2019, 20(21), 5256; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20215256
Submission received: 18 September 2019
/
Revised: 20 October 2019
/
Accepted: 21 October 2019
/
Published: 23 October 2019
(This article belongs to the Special Issue Plant Elicitors of Resistance and the Future of Plant Protection)
Abstract
:Worldwide, citrus is one of the most important fruit crops and is grown in more than 130 countries, predominantly in tropical and subtropical areas. The healthy progress of the citrus industry has been seriously affected by biotic and abiotic stresses. Several diseases, such as canker and huanglongbing, etc., rigorously affect citrus plant growth, fruit quality, and yield. Genetic engineering technologies, such as genetic transformation and genome editing, represent successful and attractive approaches for developing disease-resistant crops. These genetic engineering technologies have been widely used to develop citrus disease-resistant varieties against canker, huanglongbing, and many other fungal and viral diseases. Recently, clustered regularly interspaced short palindromic repeats (CRISPR)-based systems have made genome editing an indispensable genetic manipulation tool that has been applied to many crops, including citrus. The improved CRISPR systems, such as CRISPR/CRISPR-associated protein (Cas)9 and CRISPR/Cpf1 systems, can provide a promising new corridor for generating citrus varieties that are resistant to different pathogens. The advances in biotechnological tools and the complete genome sequence of several citrus species will undoubtedly improve the breeding for citrus disease resistance with a much greater degree of precision. Here, we attempt to summarize the recent successful progress that has been achieved in the effective application of genetic engineering and genome editing technologies to obtain citrus disease-resistant (bacterial, fungal, and virus) crops. Furthermore, we also discuss the opportunities and challenges of genetic engineering and genome editing technologies for citrus disease resistance.
1. Introduction
The genus Citrus and related genera (Fortunella, Poncirus, Eremocitrus, and Microcitrus), representing one of the most widely grown fruits, belong to the angiosperm subfamily Aurantioideae of the Rutaceae family [1]. The fruits of citrus are rich in many nutrients, especially vitamin C, and hence constitute important parts of the daily diet and world fruit crops. Disease, drought, cold, and soil salinity are the main factors that can limit citrus production, among which, disease is especially significant, such as citrus canker, huanglongbing (HLB, citrus greening), and other fungal or viral diseases [2,3]. Diseases (bacterial, fungal, or viral) can appear in a region, and within a few years, can spread and have a major economic impact. Various approaches, such as the use of chemical pesticides and other synthetic molecules, have been used to control diseases in crop plants, including citrus [4,5]. However, the side effects of these chemicals should not be ignored, such as an increased pesticide resistance in the pathogens, resurgence, pesticide residues in agricultural products, environmental pollution, and ecological balance issues [6]. Therefore, durable disease resistance is an important aim in each breeding program, and developing disease-resistant cultivars is a prime objective of breeders. Traditional breeding methods have been successfully used to improve citrus cultivars and develop new varieties in the past, but this has been done with difficulty and limitations due to the large plant size and long juvenility of this crop, incompatibility, polyembryony, heterozygosity, and parthenocarpy, etc. [7,8]. Hence, in traditional breeding, it is difficult to improve the desired traits of citrus in the short term. Additionally, traditional breeding is mainly restricted to the traits related to fruit quality, such as the fruit ripening time, flesh color, and seed number [9].
Genetic engineering is the deliberate modification of the characteristics of an organism by the manipulation of its genetic material, which has been described as a new technological paradigm [10]. The main genetic engineering technology is based on transgenesis that followed the discovery of the recombinant DNA technique, which allows plant breeders to cross crop species and introduce genes from non-related plants and other organisms into crop plants [11,12]. The fundamental strategy in genetic engineering is to modify plants so that they are productive in adverse conditions caused by biotic and abiotic stress, for instance, pathogens, pests, drought, saline, and unfertile environments [8]. Generally, genetic engineering via transformation is an alternative method to incorporate desirable traits in plants in a short cycle, with a high efficiency and easy control. Since the first reports of transgenic plants appeared in 1984 [13], there has been very rapid progress directed at using this new technology for the practical end of crop improvement. In recent years, genetic manipulation has been employed as a new route to overcome the intrinsic barriers of traditional techniques, and genetic engineering methods based on the introduction of transgenes and development of transgenic plants have been successfully adopted to improve crops [14]. Transgenic crops have gained attention worldwide since they have emerged. By introducing bioengineering technology in crop-breeding issues, genetically modified crops with an improved quality, enhanced resistance to biotic or abiotic stresses, increased yield, or reduced harmful components have been generated [15,16,17].
During the last two or three decades, genetic engineering methods based on the use of transgenes have been successfully adopted to improve fruit plants and have been mainly focused on an enhanced tolerance to biotic and abiotic stresses, increased fruit yield, improved post-harvest shelf life, reduced generation time, and production of fruit with a higher nutritional value [18]. Many successful transformation events have been achieved and different genes have been introduced into citrus that comprise antibiotic and reporter genes, genes that shorten the juvenile phase, genes that confer stress tolerance and disease resistance, and fruit quality-related genes [19,20,21,22,23].
Among all the techniques, genetic engineering has been successfully used to obtain enhanced disease-resistant citrus, which suggests that genetic engineering is an efficient approach for the development of disease-resistant citrus cultivars. In this review article, we attempt to summarize the recent successful progress that has been achieved in the effective application of genetic engineering and genome-editing technologies to obtain citrus disease-resistant (bacterial, fungal, and virus) crops. Furthermore, the opportunities and challenges of genetic engineering and genome editing technology for citrus disease resistance are also discussed here.
2. Genetic Engineering History of Citrus Crops
Tremendous progress has been made in both our scientific understanding and technological capabilities since the first report of a genetically engineered crop conferring resistance to disease, which was a virus-resistant tobacco expressing a viral coat protein gene [24]. For a better understanding of citrus genetic engineering history, we have divided it into three phases, based on major advances (Figure 1).
2.1. Phase I: Development of Transformation Protocols (1989–1999)
After the development of the polymerase chain reaction (PCR) in 1983 by Kary Mullis, a revolutionary method, the development of genetic transformation protocols, has been an attractive field of research. For citrus genetic transformation, several protocols have been developed. Kobayashi and Uchimiya made the first attempt to use a citrus protoplast to develop genetically modified plants; however, Vardi et al. reported the first successful attempt by generating a transgenic citrus plant through a polyethylene glycol (PEG) mediated direct DNA transfer method [25,26]. Subsequently, Agrobacterium-mediated transformation was developed by Hidaka et al. and has become the most widely used gene transfer method in citrus since [27]. Besides the indirect gene-transfer methods, some successful studies have been performed by direct gene-transfer methods using electroporation or particle bombardment [28]. Yao et al. reported the first successful transformation of citrus embryogenic cells using particle bombardment, which was reported in tangelo (C. reticulata × C. paradisi) [29]. To optimize the efficiency of these transformation protocols/methods, many studies have continually been published for different citrus species.
2.2. Phase II: Genetic Transformation for Stress Tolerance (2000–2013)
To develop stress-tolerant citrus for both biotic and abiotic stresses, Agrobacterium-mediated transformation has been the most widely used gene transfer method in citrus. In order to obtain a high degree of genetic transformation precision, different types of citrus explants have been used, such as internodal segments, embryogenic suspension cultures, embryo callus, axillary buds, leaf segments, cotyledon, epicotyls, and shoot segments [9,30,31,32,33,34,35,36,37]. Most of the achievements made through Agrobacterium-mediated transformation have been cited in later parts of the paper. Recently, RNA interference (RNAi) protocols have been used to develop transgenic resistance to citrus tristeza virus (CTV) by silencing the expression of critical genes in CTV-infected cells [38,39,40].
2.3. Phase III: Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR)/CAS (CRISPR-Associated) Systems (2014–Present)
Genome editing technology, a way to make precise changes to the genomic DNA of a cell or organism, have become a powerful tool for the precise manipulation of targeted genome sequences in crops [41,42,43]. Currently, there are three major types of sequence-specific nucleases for genome editing: zinc finger nucleases (ZFNs) and transcription activators such as effector nucleases (TALENs) and the clustered regularly interspaced short palindromic repeats (CRISPR)/Cas (CRISPR-associated protein) system [43]. In comparison to ZFNs and TALENs, the CRISPR/Cas9 system is widely used in laboratories because of its simplicity, design flexibility, and high efficiency. Recently, the CRISPR/Cas system has been successfully reported for citrus, with some good results for enhancing disease resistance [44,45,46,47,48,49]. More details will be discussed later in the paper in Section 5.
3. Advances in Transgenic Research for Bacterial Disease Resistance in Citrus
Genetic transformation is one of the important methods of choice for protecting susceptible citrus cultivars against canker or HLB caused by related bacterial pathogens in a shorter time. Therefore, transgenic approaches introducing exogenous genes, such as plant resistance genes, key positive regulators of SAR genes, antimicrobial peptide genes, plant metabolic genes, pathogenic genes, and kinase genes, have been applied to generate transgenic citrus crops resistant to canker or HLB infections through Agrobacterium-mediated transformation. All the successful transformation events for improving bacterial disease resistance in citrus are discussed and listed below.
3.1. Transgenic Research Related to Canker Resistance
Citrus canker, caused by the bacterial pathogen Xanthomonas citri ssp. citri (Xcc), is one of the most destructive citrus cultivar diseases reported all over the world [50]. Citrus canker affects citrus production, leading to yield losses, a poor fruit quality, and trade barriers [51]. Strategies like eradication and pathogen exclusion have been mainly used to manage the disease [5]. At present, cultural practices and chemical controls are the main methods used to manage citrus canker. Among the chemicals, copper-based chemicals have been expressed an adequate control of Xcc due to prolonged residual activity compared to other contact bactericides [52,53], such as copper oxychloride, copper hydroxide, copper sulphate, and ammonia-copper carbonate, which have been found to be highly effective against Xcc [54]. However, a continuous reliance on these compounds can cause mutations and the emergence of aggressive races of Xcc [52,53]. In addition, these methods are expensive and harmful to the environment. Hence, the use of resistant cultivars would be a better method to control citrus canker and resistance might be introduced through cross breeding. However, there have been no reports about the development of resistant citrus cultivars via conventional breeding. Genetic engineering may be a better method of improving disease resistance to citrus canker [55].
To improve the resistance to canker in citrus by transgenic approaches, different strategies have been proposed, including the over-expression of genes that code for antibacterial peptides, disease-resistance proteins, the kinase gene, transcription factors, and other exogenous genes of a plant/non-plant origin that enhance natural plant defenses (Table 1). Antimicrobial peptides are important components of innate immune defense against microbial pathogens in a wide range of organisms [56,57]. To obtain resistant cultivars, different antimicrobial peptide genes, such as attacin A, Shiva A, Cecropin B, Stx IA, D2A21, and the dermaseptin gene, have been introduced into citrus cultivars and rootstock to enhance canker resistance [35,58,59,60,61,62,63], the results of which indicate that the over-expression of these exogenous peptide genes significantly enhances their canker disease resistance. The Arabidopsis NPR1 gene (AtNPR1) has been well-established as a key positive regulator of systemic acquired resistance (SAR), which acts downstream of the signal molecule SA. Over-expression of the AtNPR1 gene or its orthologs also enhances disease resistance in many crop plants including rice, wheat, rapeseed, tomato, and apple [64,65,66,67,68], which makes AtNPR1 a workable target for the genetic engineering of non-specific resistance in plants. This kind of broad-spectrum disease resistance gene was also introduced into citrus to enhance canker resistance. Two previous reports showed that transgenic grapefruit and sweet orange that over-express the positive regulator of SAR, the AtNPR1 gene, or its homologous gene CtNH1 from Citrus maxima, were less susceptible to Xcc [55,69]. The introduction of resistance genes (R-genes) is one of the strategies used to improve the plant’s resistance to pathogens [70]. Disease-resistant R-genes are frequently used in breeding for crop protection. The Bs2 gene is a member of the nucleotide binding site-leucine-rich repeat (NBS-LRR) class of R genes, which has been shown to confer resistance against pathogenic strains of Xanthomonas campestris pv. vesicatoria (Xcv) in susceptible pepper, tomato, and tobacco plants [71]. It has been reported that the over-expression of Bs2 from pepper leads to a decreased susceptibility to Xcc [72,73]. Another R gene, Xa21, first cloned from the wild rice Oryza longistaminata, encodes a receptor kinase-like protein that consists of LRR [74]. Likewise, transgenic citrus with the rice Xa21 gene showed less susceptibility to Xcc [75,76,77,78]. Reactive oxygen species (ROSs) have emerged as important regulators of plant stress responses, and were observed in a wide range of plant–pathogen interactions involving bacteria, fungi, and viruses [79]. The accumulation of ROSs was proposed as the earliest event induced during plant–pathogen interaction, which controls and inhibits pathogen growth. Transient elevations in ROS levels can enhance stress tolerance by activating the defense mechanisms, including kinases and components of the signaling network [80]. Over-expression of the pathogen-associated molecular pattern (PAMP) receptor NbFLS2 can increase ROS production and activate PAMP-triggered immunity and defense-associated gene expression in citrus, the results of which showed that the integration and expression of the NbFLS2 gene can increase canker resistance [81]; the mitogen-activated protein kinase gene CsMAPK1 functions in the citrus canker defense response through the activation of defense-related gene expression and ROS production during infection, leading to a reduction in canker symptoms and a decrease in bacterial growth [82].
Other exogenous genes, such as hairpin gene hrpN, spermidine synthase gene MdSPDS1, transcription factor terf1, pathogenesis gene pthA-nls, and cysteine-rich peptide theonin, have also been introduced into citrus, and all the results showed that the transgenic lines of citrus with these genes were less susceptible to canker disease [83,84,85,86,87]. Therefore, the over-expression of exogenous genes in citrus is a promising approach for the development of cultivars that are more resistant to citrus canker.
3.2. Transgenic Research for Huanglongbing Resistance
Huanglongbing (HLB), another devastating citrus disease worldwide, is also known as citrus greening. HLB is mainly associated with phloem-limited bacteria that belong to the Candidatus Liberibacter genus, including Candidatus Liberibacter asiaticus (CaLas), Candidatus Liberibacter americanus (CaLam), and Candidatus Liberibacter africanus (CaLaf) [88,89,90]. The insect vector responsible for the transmission of these phloem bacteria can be either the Asian citrus psyllid, Diaphorina citri, or the African citrus psyllid, Trioza erytreae [91]. HLB infection often leads to a drastic reduction in the quantity and quality of citrus fruits and eventually renders the infected trees useless [91,92]. The major strategies for managing and controlling HLB disease have been summarized in previous literature, including the vector control of psyllid populations (chemical and biological control), use of antimicrobials, thermotherapy, use of disease-free planting materials, and nutrient enhancement of the trees [90,93]. Among these options, chemical control to reduce psyllid and Ca. Liberibacter populations has been used broadly. From the diverse set of compounds tested, ampicillin, carbenicillin, penicillin, cefalexin, oxytetracycline, streptomycin sulfate, rifampicin, and sulfadimethoxine have been shown to be highly effective in suppressing HLB [94]. However, the usage of antibiotics needs to continue over time to control HLB over the long term, which also introduces problems associated with operation costs and potentially adverse environmental effects. Furthermore, almost all commercial citrus cultivars are susceptible to HLB. Therefore, the best solution for the management of HLB is to develop resistant or tolerant cultivars of important citrus species through genetic engineering, which remains the fastest method for the improvement of existing citrus cultivars [95].
In a previous report, it was found that transgenic citrus lines expressing the broad-spectrum disease-resistance gene AtNPR1 in citrus increase resistance to citrus canker [55]. Recent results from two reports show that over-expressing AtNPR1 can also provide resistance to HLB. Dutt et al. showed that some transgenic Hamlin and Valencia sweet orange lines that over-express the AtNPR1 gene, under a constitutive CaMV35S promoter and a phloem-specific AtSUC2 promoter, exhibited reduced disease severity and a few lines remained disease-free, even after 36 months of planting in a high-disease pressure field site [22]. Results from Robertson et al. also indicate that the expression of high levels of AtNPR1 in citrus can provide tolerance to HLB under strong disease pressure in the greenhouse [89]. These results are also in agreement with the findings reported by Wang et al. [96], who showed that four AtNPR1-like genes were expressed to significantly higher levels in the HLB-tolerant genotype than in the HLB-susceptible genotype, by evaluating transcriptome differences between two closely related cultivars (HLB-tolerant Jackson grapefruit-like hybrid and HLB-susceptible Marsh grapefruit trees) after HLB infection. All these studies indicate that AtNPR1-like gene-mediated defense signaling may contribute to the HLB-tolerant phenotype [96]. In addition, three other exogenous-resistant genes were separately introduced into the citrus, all of which improved the resistance to HLB. The expression of antimicrobial gene attacin A (attA) in sweet orange Hamlin and Pêra showed significantly fewer symptoms of HLB compared to non-transgenic plants of this cultivar [97,98]. A modified thion was introduced into citrus and the transgenic citrus showed a stronger resistance to HLB and citrus canker [87]. Cecropin B, an antimicrobial peptide from Chinese oak silk worm, was expressed under the phloem-specific promoter GRP1.8 from the French bean to reduce the susceptibility of Tarocco blood orange to HLB [99].
4. Advances in Transgenic Research for Fungal and Viral Disease Resistance in Citrus
New approaches and research in genetic engineering have provided novel opportunities for the generation of plants for resistance against funguses and viruses outside of conventional breeding methods. In addition to canker and HLB diseases, attention is now also being given to other fungal and viral diseases in citrus, such as citrus tristeza virus (CTV), citrus psorosis (CP), root rot and gummosis, mal secco, gray mold, black spot, and citrus scab. Resistance to these fungal and viral diseases has also been enhanced by transgenic technology (Table 2).
CTV, one of the most important viral diseases affecting citrus, causes a quick decline of most citrus species, as well as a reduction in the fruit yield and quality of some cultivars [100,101]. Due to the agronomic characteristics of citrus cultivars, genetic transformation appears to be the most promising technique for developing CTV resistance, and the possibility of creating transgenic plants with an enhanced resistance to CTV has been evaluated using different gene constructs and citrus genotypes [101]. The pathogen-derived resistance (PDR) strategy has been successfully used in developing CTV tolerance or resistance materials by introducing genes or selected segments of the CTV genome in citrus plants. Transgenic Mexican limes that express p25 and p23 were obtained by genetic engineering and found to show a significant delay in virus accumulation and no CTV symptoms, so were capable of conferring resistance to CTV [102,103]. Ananthakrishnan et al. found that grapefruit expressing p23 and 3′-UTR exhibited reduced CTV replication in protoplasts [104]. Muniz et al. transformed two sweet oranges with p25, intron-spliced hairpin p25 and a 559-nt-long 3′-terminal conserved region, respectively, and found that some lines might have partial repressing effects on virus replication [105]. In some cases, expression of the CTV coat-protein gene led to an increase in CTV resistance. Protoplasts isolated from ten sweet orange callus lines genetically transformed with the CTV-392/393 sequence from the CTV genome exhibited different CTV replication levels [106]. Additionally, Cervera et al. reported the first transgenic Mexican lime by the ectopic expression of single-chain variable fragment (scFv) recombinant antibodies, and most transgenic lines displayed resistance or tolerance after being challenged by CTV inoculation [107]. RNA silencing or RNA interference (RNAi) has been found to be the most important mechanism that plants use to combat viral infections [108]. The main strategy of RNAi for obtaining viral resistance is to express viral sequences that will form self-complementary hairpin RNA (hpRNA) in the host when expressed [109]. In citrus, nine lines of transformed Mexican lime with sense, antisense, and intro-hairpin versions of the 549-nt-long 3′-terminal of CTV were found to show CTV resistance [38]. It was reported that three transgenic Mexican lime lines expressing an untranslatable version of the three CTV-silencing suppressor genes in the intron-hairpin version showed complete resistance to CTV-T36 infection under laboratory conditions [39]. Similarly, some lines of transformed sour orange with a hairpin-structured p20 conservative region showed resistance or tolerance to severe CTV strains [109,110].
Similar work has also been done to enhance the resistance of other citrus pathogens by researchers. Two transgenic sweet orange lines containing different genes of citrus psorosis virus (CPsV) were obtained separately, both of which showed a delay in CPsV symptoms [111,112]. Furthermore, intron-hairpin RNA transcripts corresponding to CP genes (ihpCP) of CPsV were introduced into sweet orange, leading to the regeneration of transgenic plants expressing ihpRNA with enhanced CPsV resistance [113]. Various endochitinase genes, such as chit42 from Trichoderma harzianum, have also been successfully transformed and expressed to impart an increased fungal tolerance in lemon. Transgenic lemon with chit42 showed significantly less lesion development after inoculation with Phoma tracheiphila and Botrytis cinerea, the causal agents of mal secco and gray mold in citrus [114,115]. A gene for the PR-5 protein from tomato has been expressed in transgenic sweet orange and regenerants, and all the transgenic lines exhibited significant protection against Phytophthora citrophthora, which causes root rot and gummosis [116]. Several transgenic Duncan grapefruit lines expressing the antimicrobial Attacin E gene had a significantly lower susceptibility to Elsinoë fawcettii compared to the non-transformed control, and significant activity against citrus scab was unexpectedly found [117]. Additionally, regulation of the level of D-limonene in citrus fruit by a transgenic approach was attempted to enhance the resistance against pathogens such as black spot [118].
5. Application of Genome-Editing Techniques in Disease-Resistance Breeding of Citrus
Genome editing, a specific and efficient tool for generating useful novel phenotypes, surely represents a great technical innovation in plant breeding. Generally, genome-editing technology employs three types of engineered endonucleases: zinc finger nucleases (ZFNs), transcription activator-like effector nucleases (TALENs), and clustered regularly interspaced short palindromic repeats and CRISPR-associated proteins (CRISPR/Cas) for site-specific cleavage and the emerging CRISPR/Cas9 is comparatively easy to prepare, affordable, and can be scaled up better than ZFNs and TALENs [119,120,121]. Though developed recently, CRISPR/Cas9 technology has already been established in several important plant species through gene mutation, repression, activation, and epigenome editing, such as rice [122], wheat [123], maize [124], and some horticultural crops, including tomato, petunia, citrus, grape, potato, carrot, and apple [125,126,127,128,129]. Using this technology, many agronomically important traits, such as heat/cold tolerance, disease resistance, herbicide tolerance, and yield improvement, have been introduced in plants [119,130]. CRISPR/Cas9 technology has been efficiently applied in developing disease resistance to many viruses [131,132]. In a previous report, CRISPR/Cas9 was successfully used to incorporate the mutagenesis insusceptible genes MLO-7 in grape cultivar Chardonnay and DIPM-1, DIPM-2, and DIPM-4 in apple cultivar Golden delicious, to increase their resistance to powdery mildew and fire blight disease, respectively [133].
In citrus, the CRISPR/Cas9 system was firstly used to target the CsPDS gene in sweet orange and Duncan grapefruit and successfully modified the CsPDS gene via Xcc-facilitated agro infiltration, an optimized transient expression method [44,134]. As expected, the modified CsPDS sequence expressing Cas9/sgRNA was not detected in sweet orange leaves [44], which indicated that CRISPR/Cas9 successfully induced mutations in the targeted gene in citrus. CsLOB1, a member of the lateral organ boundaries domain (LBD) family of transcription factors, was previously identified as a critical citrus disease susceptibility gene for citrus canker [135]. The strain of Xcc, bacteria that causes citrus canker, encodes transcription activator-like (TAL) effector PthA4 that binds to the effector-binding elements (EBE) in the promoter of CsLOB1 and activates the expression of this canker-susceptibility gene [135]. Interestingly, CRISPR/Cas9-mediated modification of the EBE of one single allele of the CsLOB1 gene in Duncan grapefruit alleviated the canker symptoms slightly [45]. However, mutation of the EBEs of both alleles of CsLOB1 promoters showed a high degree of resistance to citrus canker in Wanjincheng orange [47]. Furthermore, another study showed that editing the coding region of CsLOB1 via CRISPR/Cas9 also provided resistance to canker in Duncan grapefruit [46]. In a recent report, CsWRKY22, a marker gene for pathogen-triggered immunity in Wanjincheng orange, was knocked out by the CRISPR/Cas9 system, and the mutant plants showed a significantly decreased susceptibility to citrus canker [49]. In addition, CRISPR/Cas12a (Cpf1), another improved CRISPR/Cas system, has been employed to edit the Duncan grapefruit gene CsPDS with a higher efficiency and lower off-target effects, which will enhance the scope of citrus genome editing [48]. These studies indicate that CRISPR-mediated genome editing can be a promising pathway to generate disease-resistant citrus cultivars. Due to the vague virulence mechanisms of HLB and other diseases, the CRISPR/Cas system cannot achieve the goal of conferring resistance to citrus HLB and other diseases at present. However, researchers are now attempting to use the genome editing system to create citrus cultivars that are less susceptible to HLB [136,137].
Besides the recent successes, the CRISPR/Cas system faces several limitations and challenges in citrus. For instance, the most debated challenges in the CRISPR/Cas system are its potential off-target mutations and unintended on-target changes. In citrus, its polyploidy nature poses a challenge for the CRISPR/Cas system, because it has been proved that the gene-editing efficiency is usually lower in polyploidy plants than diploids, as multiple alleles must be edited simultaneously [45,47,138]. Therefore, further research is needed to increase the efficiency of tissue culture-based Agrobacterium-mediated delivery of the CRISPR/Cas system efficiency in different citrus species. However, in a recent report, Hongkong kumquat (Fortunella hindsii), a wild citrus species with a short juvenility, monoembryony, and applicability of CRISPR, is being exploited as a model species for citrus research [139].
6. Genetic Approaches to Suppress Vector-Borne Bacterial/Viral Diseases in Citrus
Recently, several reports have shown the great potential of RNAi and CRISPR to develop novel management strategies of vector-borne viral/bacterial diseases in citrus [140,141]. For instance, HLB diseases have been successfully controlled by applying RNAi-mediated protection against the Candidatus Liberibacter asiaticus (CLas) bacteria vector African citrus psyllid (ACP) Diaphorina citri through bioassays using plant feeding systems [142,143,144,145,146,147,148], detached leaf bioassays [149,150], leaf disc, artificial diet, sugar solutions [151]; topical applications and soaking [152,153,154]. All the results showed that the ACP was very sensitive to ingested dsRNA, causing significant suppression of the targeted transcript and increased psyllid mortality. Additionally, the application of RNA suppression using antisense oligonucleotides, such as 2’F-ANA-ASO/ASO and cell-penetrating peptide morpholinos (PPMOs), was also reported to trigger RNAi degradation for suppressing pests and pathogens in infected citrus seedling trees, which significantly reduced ACP and CLas bacteria [140]. CRISPR-Cas9 was used to conduct a knockout of the thioredoxin gene in ACP, and the CRISPR-Cas9-treated psyllids resulted in psyllids with longer development times, shorter adult lifespans, and a reduced fecundity [140,155]. Interestingly, CRISPR/Cas system was ever found in the CLas genera [156], which could be used to develop effective strategies for controlling the HLB pathogen.
Besides HLB, CTV and citrus variegated chlorosis (CVC) diseases have also been reduced by RNAi-mediated protection strategies against Aphis (Toxoptera) citricidus (Kirkaldy) [157,158], and Homalodisca vitripennis [159,160] in citrus. For citrus canker, recently, two studies have shown that the action mode of the main TAL effector PthA4/PthA4AT from Xcc may provide new clues for controlling canker by interfering with the bacteria Xcc [161,162]. Therefore, RNAi and CRISPR could definitely be exploited to develop novel management strategies to suppress the vectors and pathogens, thereby bringing benefits to both growers and consumers without gene-editing plants.
7. Conclusions and Future Perspectives
Due to some of the biological characteristics of citrus, conventional breeding methods have demonstrated limitations to producing new citrus cultivars with an improved resistance to diseases. Therefore, genetic engineering, including transgenic or genome-editing technologies, can be a method of choice to overcome the limitations posed by traditional breeding and have made it possible to protect susceptible commercial cultivars against pathogens, which allows the release of improved cultivars with desirable characteristics in a shorter time. Exogenous genes, such as plant-resistance genes, key positive SAR regulator genes, insect antimicrobial peptide genes, plant metabolic genes, pathogenic genes, and kinase genes, have been introduced in citrus through Agrobacterium-mediated genetic transformation, and transgenic lines with an excellent resistance to canker have been obtained (Table 1). Furthermore, the broad-spectrum disease-resistance gene AtNPR1, a modified plant thionin gene, and antimicrobial peptide genes Cecropin B and attacin A, have been respectively introduced into citrus, and all effectively improved the resistance to HLB. These genes of a plant or non-plant origin encoding antibacterial proteins, such as plant resistance genes (R-genes), regulators of SAR genes, antimicrobial peptide genes, etc., have been introduced in citrus to inhibit bacterial pathogenicity factors though various genetic techniques. Transgenic plants that constitutively express proteins with potential antibacterial or antifungal activity can reduce the activities of specific soil-borne plant pathogens in the rhizosphere, and affect specific plant-beneficial components of the rhizosphere microflora [163,164], resulting in changes of citrus disease resistance. Antimicrobial peptides (AMPs), especially, are important components of the innate immune defense system against microbial pathogens [165], for which the modes of action in transgenic plants may involve interactions between the peptides and bacterial membrane, leading to bacterial membrane disruption, cytoplasmic leakage, and interference with intracellular macromolecule synthesis to rapidly kill the bacteria [165,166]. Besides the main bacterial diseases, resistance to other fungal and viral diseases in citrus, including anthracnose, gray mold, scab, black spot, root rot and gummosis, tristeza virus, and psorosis, were also enhanced by transgenic technology (Table 2). However, conventional biomolecule delivery methods in plants, including electroporation, biolistics, Agrobacterium-mediated delivery, and cationic delivery, have critical drawbacks, such as a low efficiency, narrow species range, limited cargo types, and tissue damage [167]. Therefore, improving the existing delivery systems and developing new systems will be key to reducing barriers to the inexpensive application of genetic engineering in plants, especially genome editing. Nanomaterials have unique and tunable physical and chemical properties, which can interact with biological matter with exquisite control and precision [168]. Nanoparticles can be used as a carrier system to deliver the genetic materials, such as plasmid DNA, RNA, and oligonucleotides into cells efficiently and rapidly, which reduce the drawbacks and limitations associated with current Agrobacterium-mediated transgene delivery systems [167,168,169]. A few successful examples show promise for nanoparticle-mediated passive delivery to plants in vitro [170,171,172] and in vivo [173,174], indicating the potential for passive nanoparticle-mediated delivery with a high efficiency and low toxicity. In the future, nanotechnology-mediated delivery system can overcome the delivery challenges and enhance the efficacy of citrus genetic engineering.
Novel genome-editing systems help introduce stably inherited point modifications in the plant genome and allow the creation of non-transgenic plants [175,176], which may meet the challenges faced by citrus breeders, such as the quest to develop productive, disease-resistant varieties with tasty, high-quality, and nutrient-packed fruit. In contrast to ZFNs and TALENs, second-generation genome editing techniques like CRISPR-Cas9 and CRISPR/Cpf1 involve easier design and execution methodologies that are also more time- and cost-effective [120], both of which are currently the best studied and most widely used CRISPR systems in plants. In citrus, the canker susceptibility gene CsLOB1 and canker immunity response gene CsWRKY22 have been modified at the promoter region or coding region by CRISPR/Cas9 [45,46,47,49] and the citrus CsPDS gene was also effectively modified via a new improved CRISPR/Cpf1 system [48]. The genomes of an increasing number of citrus and disease-related bacterial species have already been sequenced or are in the process of being sequenced, which will solve the problem of a lack of genomic information and thus permit the target gene/site (susceptibility gene) to be modified using genome-editing technology. Furthermore, continuously improved genome-editing technologies with a nanotechnology-mediated delivery system will play a significant role in developing new disease-resistant citrus cultivars in the future.
Worldwide, the human population is projected to reach 9.8 billion in 2050, and it is estimated that the global food supply will need to increase by 70% till 2050 to meet the rapidly mounting population nutritional demands [177,178]. Various abiotic stresses, biotic stresses, and recent climate change waves worsen the task that the world face in terms of meeting the nutritional necessities. Genetic engineering, which refers to the direct alteration of an organism’s genetic material using biotechnology, could be an integrated and diversified approach against these challenges [179]. Genetically engineered crops in general increase crop productivity through a reduction in the cost of cultivation and crop loss caused by biotic and abiotic stresses, so provide more affordable food [180]. In addition to economic gains, genetically engineered crops reduce the number of insecticide applications, importantly contributing to a more sustainable environment. The products of genetic engineering are often referred to as GE or GMO (genetically modified organisms) [181]. Despite transgenic crops, such as soybeans, maize, cotton, and canola, having enjoyed wide commercial success around the world [182], their commercial utilization is still hampered by some legal, ethical, and experimental issues, which mostly limit their use to research. In contrast, CRISPR/Cas-mediated genome editing seems to be the most promising strategy to improve crop cultivars without introducing foreign genes, and the United States Department of Agriculture (USDA) has stated that CRISPR/Cas9-edited crops, which have the potential to be called non-GMO, can be cultivated and sold without regulatory monitoring [183,184]. However, confidence in applying genome-editing tools in agriculture remains limited. The biggest potential obstacles for the use of genome-editing technologies in agriculture are public acceptance and government regulation [185]. There is still no internationally accepted regulatory framework for gene-editing products, and different countries/agencies have different takes on the use of GMOs [185,186]. For instance, the European regulatory agencies emphasize how the plants were produced, and have recently ruled that gene-edited products/crops should be treated like traditional GMOs, which are under very strict regulation in the European Union [187,188]. By contrast, the USDA tends to focus on the end product, and they have determined that gene-edited crops are exempt from GMO regulations [184]. Furthermore, the USDA has outlined that genome-edited crops can be considered as products from biological mutagenesis, such as chemical and radiation mutagenesis widely used in conventional plant breeding. Overcoming regulatory obstacles requires public awareness and political willingness in order to strive for some form of consistency among countries to establish a clear position on genome editing technologies. The regulatory processes are often unpredictable and challenging. However, continued dialog among regulatory authorities, as well as positive actions taken by progress-oriented countries, will help to bring about improvements in the GMO product regulations.
Author Contributions
L.S. drafted the manuscript. N., J.X. and F.K. contributed to editing of the manuscript. Z.N. and P.W. reviewed and supervised. All authors have read and approved the final manuscript to be published.
Funding
This work was supported by the earmarked fund for China Agriculture Research System (CARS-26), Sub-project of “Breeding New Fruit Cultivars Major Project of Zhejiang Province” (grant no. 2016C12904-1), Citrus Breeding Supporting Discipline Project of Zhejiang Academy of Agricultural Sciences and Advantaged Fruit Industry Technology Innovation Team Project of Taizhou.
Acknowledgments
We wish to thank Zhongyan Wei for her helpful suggestions to the manuscript. We also would like to thank the two anonymous reviewers for critical review of our manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Wu, G.A.; Terol, J.; Ibanez, V.; Lopez-Garcia, A.; Perez-Roman, E.; Borreda, C.; Domingo, C.; Tadeo, F.R.; Carbonell-Caballero, J.; Alonso, R.; et al. Genomics of the origin and evolution of Citrus. Nature 2018, 554, 311–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Febres, V.; Fisher, L.; Khalaf, A.; Moore, G.A. Citrus Transformation: Challenges and Prospects. In Genetic transformation; Alvarez, M., Ed.; IntechOpen: London, UK, 2011; pp. 101–122. [Google Scholar]
- Upadhyay, A. Transgenic Research in Fruit Crops. In Genetic Engineering of Horticultural Crops; Rout, G.R., Peter, K.V., Eds.; Academic Press: Cambridge, MA, USA, 2018; pp. 63–87. [Google Scholar]
- Prabha, R.; Singh, D.P.; Verma, M.K. Microbial Interactions and Perspectives for Bioremediation of Pesticides in the Soils. In Plant-Microbe Interactions in Agro-Ecological Perspectives; Singh, D.P., Singh, H.B., Prabha, R., Eds.; Springer: Singapore, 2017; Volume 2, pp. 649–671. [Google Scholar]
- Gimenes-Fernandes, N.; Barbosa, J.C.; Ayres, A.; Massari, C. Plantas doentes não detectadas nas inspeções dificultam a erradicação do cancro cítrico. Summa Phytopathol. 2000, 26, 320–325. [Google Scholar] [CrossRef]
- Pan, X.L.; Dong, F.S.; Wu, X.H.; Xu, J.; Liu, X.G.; Zheng, Y.Q. Progress of the discovery, application, and control technologies of chemical pesticides in China. J. Integr. Agric. 2018, 18, 840–853. [Google Scholar] [CrossRef]
- Grosser, J.; Gmitter, F.; Tusa, N.; Chandler, J. Somatic hybrid plants from sexually incompatible woody species: Citrus reticulata and Citropsis gilletiana. Plant Cell Rep. 1990, 8, 656–659. [Google Scholar] [CrossRef] [PubMed]
- Vanloqueren, G.; Baret, P.V. How agricultural research systems shape a technological regime that develops genetic engineering but locks out agroecological innovations. Res. Policy 2009, 38, 971–983. [Google Scholar] [CrossRef]
- Gong, X.Q.; Liu, J.H. Genetic transformation and genes for resistance to abiotic and biotic stresses in Citrus and its related genera. Plant Cell Tiss. Org. 2012, 113, 137–147. [Google Scholar] [CrossRef]
- Orsenigo, L. The Emergence of Biotechnology: Institutions and Markets in Industrial Innovation; Pinter Publishers Ltd.: London, UK, 1989. [Google Scholar]
- Cohen, S.N.; Chang, A.C.; Boyer, H.W.; Helling, R.B. Construction of biologically functional bacterial plasmids in vitro. Proc. Natl. Acad. Sci. USA 1973, 70, 3240–3244. [Google Scholar] [CrossRef]
- Melchers, L.S.; Stuiver, M.H. Novel genes for disease-resistance breeding. Curr. Opin. Plant. Biol. 2000, 3, 147–152. [Google Scholar] [CrossRef]
- Horsch, R.B.; Fraley, R.T.; Rogers, S.G.; Sanders, P.R.; Lloyd, A.; Hoffmann, N. Inheritance of functional foreign genes in plants. Science 1984, 223, 496–498. [Google Scholar] [CrossRef]
- Miedaner, T. Breeding strategies for improving plant resistance to diseases. In Advances in Plant Breeding Strategies: Agronomic, Abiotic and Biotic Stress Traits; Al-Khayri, J.M., Jain, S.M., Johnson, D.V., Eds.; Springer: Cham, Switzerland, 2016; pp. 561–599. [Google Scholar]
- Toriyama, K.; Arimoto, Y.; Uchimiya, H.; Hinata, K. Transgenic rice plants after direct gene transfer into protoplasts. Nat. Biotechnol. 1988, 6, 1072. [Google Scholar] [CrossRef]
- Gordon-Kamm, W.J.; Spencer, T.M.; Mangano, M.L.; Adams, T.R.; Daines, R.J.; Start, W.G.; O’Brien, J.V.; Chambers, S.A.; Adams, W.R.; Willetts, N.G. Transformation of maize cells and regeneration of fertile transgenic plants. Plant Cell 1990, 2, 603–618. [Google Scholar] [CrossRef] [PubMed]
- Vasilets, L.A.; Schwarz, W. Regulation of endogenous and expressed Na+/K+ pumps in Xenopus oocytes by membrane potential and stimulation of protein kinases. J. Membrane Biol. 1992, 125, 119–132. [Google Scholar] [CrossRef] [PubMed]
- Rai, M.K.; Shekhawat, N. Recent advances in genetic engineering for improvement of fruit crops. Plant Cell Tiss. Org. 2014, 116, 1–15. [Google Scholar] [CrossRef]
- Peña, L.; Martín-Trillo, M.; Juárez, J.; Pina, J.A.; Navarro, L.; Martínez-Zapater, J.M. Constitutive expression of Arabidopsis LEAFY or APETALA1 genes in citrus reduces their generation time. Nat. Biotechnol. 2001, 19, 263. [Google Scholar] [CrossRef] [PubMed]
- Alquezar, B.; Rodrigo, M.J.; Zacarías, L. Regulation of carotenoid biosynthesis during fruit maturation in the red-fleshed orange mutant Cara Cara. Phytochemistry 2008, 69, 1997–2007. [Google Scholar] [CrossRef] [PubMed]
- Pons, E.; Alquézar, B.; Rodríguez, A.; Martorell, P.; Genovés, S.; Ramón, D.; Rodrigo, M.J.; Zacarías, L.; Pena, L. Metabolic engineering of β-carotene in orange fruit increases its in vivo antioxidant properties. Plant Biotechnol. J. 2014, 12, 17–27. [Google Scholar] [CrossRef]
- Dutt, M.; Barthe, G.; Irey, M.; Grosser, J. Transgenic Citrus Expressing an ArabidopsisNPR1 Gene Exhibit Enhanced Resistance against Huanglongbing (HLB; Citrus Greening). PLoS ONE 2015, 10, e0137134. [Google Scholar] [CrossRef]
- Alquézar, B.; Rodríguez, A.; de la Peña, M.; Peña, L. Genomic Analysis of Terpene Synthase Family and Functional Characterization of Seven Sesquiterpene Synthases from Citrus sinensis. Front. Plant Sci. 2017, 8, 1481. [Google Scholar] [CrossRef]
- Abel, P.P.; Nelson, R.S.; De, B.; Hoffmann, N.; Rogers, S.G.; Fraley, R.T.; Beachy, R.N. Delay of disease development in transgenic plants that express the tobacco mosaic virus coat protein gene. Science 1986, 232, 738–743. [Google Scholar] [CrossRef]
- Kobayashi, S.; Uchimiya, H. Expression and integration of a foreign gene in orange (Citrus sinensis Osb.) protoplasts by direct DNA transfer. Jpn. J. Genet. 1989, 64, 91–97. [Google Scholar] [CrossRef]
- Vardi, A.; Bleichman, S.; Aviv, D. Genetic transformation of Citrus protoplasts and regeneration of transgenic plants. Plant Sci. 1990, 69, 199–206. [Google Scholar] [CrossRef]
- Hidaka, T.; Omura, M.; Ugaki, M.; Tomiyama, M.; Kato, A.; Ohsima, M.; Motoyoshi, F. Agrobacterium-mediated transformation and regeneration of Citrus spp. from suspension cells. Jpn. J. Breed. 1990, 40, 199–207. [Google Scholar] [CrossRef]
- Hidaka, T.; Omura, M. Transformation of Citrus protoplasts by electroporation. J. Jpn. Soc. Hortic. Sci. 1993, 62, 371–376. [Google Scholar] [CrossRef]
- Yao, J.L.; Wu, J.H.; Gleave, A.P.; Morris, B.A. Transformation of citrus embryogenic cells using particle bombardment and production of transgenic embryos. Plant Sci. 1996, 113, 175–183. [Google Scholar] [CrossRef]
- Almeida, W.A.B.D.; Mourão Filho, F.D.A.A.; Mendes, B.M.J.; Pavan, A.; Rodriguez, A.P.M. Agrobacterium-mediated transformation of Citrus sinensis and Citrus limonia epicotyl segments. Sci. Agr. 2003, 60, 23–29. [Google Scholar] [CrossRef]
- Molinari, H.; Bespalhok, J.; Kobayashi, A.; Pereira, L.; Vieira, L. Agrobacteriumtumefaciens-mediated transformation of Swingle citrumelo (Citrus paradisi Macf. × Poncirustrifoliata L. Raf.) using thin epicotyl sections. Sci. Hortic. Amst. 2004, 99, 379–385. [Google Scholar] [CrossRef]
- Ahmad, M.; Mirza, B. An efficient protocol for transient transformation of intact fruit and transgene expression in Citrus. Plant Mol. Biol. Rep. 2005, 23, 419–420. [Google Scholar] [CrossRef]
- Dutt, M.; Grosser, J. An embryogenic suspension cell culture system for Agrobacterium-mediated transformation of citrus. Plant Cell Rep. 2010, 29, 1251–1260. [Google Scholar] [CrossRef]
- Al Bachchu, M.A.; Jin, S.B.; Park, J.W.; Sun, H.J.; Yun, S.H.; Lee, H.Y.; Lee, D.S.; Hong, Q.C.; Kim, Y.W.; Riu, K.Z. Agrobacterium-mediated transformation using embryogenic calli in satsuma mandarin (Citrus unshiu Marc.) cv. Miyagawa Wase. Hortic. Environ. Biotechnol. 2011, 52, 170–175. [Google Scholar] [CrossRef]
- He, Y.; Chen, S.; Peng, A.; Zou, X.; Xu, L.; Lei, T.; Liu, X.; Yao, L. Production and evaluation of transgenic sweet orange (Citrus sinensis Osbeck) containing bivalent antibacterial peptide genes (Shiva A and Cecropin B) via a novel Agrobacterium-mediated transformation of mature axillary buds. Sci. Hortic. Amst. 2011, 128, 99–107. [Google Scholar] [CrossRef]
- Fávero, P.; Mourão Filho, F.D.A.A.; Stipp, L.C.L.; Mendes, B.M.J. Genetic transformation of three sweet orange cultivars from explants of adult plants. Acta. Physiol. Plant. 2012, 34, 471–477. [Google Scholar] [CrossRef]
- Khan, E.U.; Fu, X.Z.; Liu, J.H. Agrobacterium-mediated genetic transformation and regeneration of transgenic plants using leaf segments as explants in Valencia sweet orange. Plant Cell Tiss. Org. 2012, 109, 383–390. [Google Scholar] [CrossRef]
- Lopez, C.; Cervera, M.; Fagoaga, C.; Moreno, P.; Navarro, L.; Flores, R.; Pena, L. Accumulation of transgene-derived siRNAs is not sufficient for RNAi-mediated protection against Citrus tristeza virus in transgenic Mexican lime. Mol. Plant Pathol. 2010, 11, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Soler, N.; Plomer, M.; Fagoaga, C.; Moreno, P.; Navarro, L.; Flores, R.; Peña, L. Transformation of Mexican lime with an intron-hairpin construct expressing untranslatable versions of the genes coding for the three silencing suppressors of Citrus tristeza virus confers complete resistance to the virus. Plant Biotechnol. J. 2012, 10, 597–608. [Google Scholar] [CrossRef] [PubMed]
- Donmez, D.; Simsek, O.; Izgu, T.; Aka Kacar, Y.; Yalcin Mendi, Y. Genetic transformation in citrus. Sci. World J. 2013, 2013, 491207. [Google Scholar] [CrossRef]
- Arora, L.; Narula, A. Gene editing and crop improvement using CRISPR-Cas9 system. Front. Plant Sci. 2017, 8, 1932. [Google Scholar] [CrossRef]
- Limera, C.; Sabbadini, S.; Sweet, J.B.; Mezzetti, B. New biotechnological tools for the genetic improvement of major woody fruit species. Front. Plant Sci. 2017, 8, 1418. [Google Scholar] [CrossRef]
- Yin, K.; Gao, C.; Qiu, J.L. Progress and prospects in plant genome editing. Nat. Plants 2017, 3, 17107. [Google Scholar] [CrossRef]
- Jia, H.; Wang, N. Targeted genome editing of sweet orange using Cas9/sgRNA. PLoS ONE 2014, 9, e93806. [Google Scholar] [CrossRef]
- Jia, H.; Orbovic, V.; Jones, J.B.; Wang, N. Modification of the PthA4 effector binding elements in Type I CsLOB1 promoter using Cas9/sgRNA to produce transgenic Duncan grapefruit alleviating XccΔpthA4:dCs LOB1.3 infection. Plant Biotechnol. J. 2016, 14, 1291–1301. [Google Scholar] [CrossRef]
- Jia, H.; Zhang, Y.; Orbovic, V.; Xu, J.; White, F.F.; Jones, J.B.; Wang, N. Genome editing of the disease susceptibility gene CsLOB1 in citrus confers resistance to citrus canker. Plant Biotechnol. J. 2017, 15, 817–823. [Google Scholar] [CrossRef] [PubMed]
- Peng, A.; Chen, S.; Lei, T.; Xu, L.; He, Y.; Wu, L.; Yao, L.; Zou, X. Engineering canker-resistant plants through CRISPR/Cas9-targeted editing of the susceptibility gene CsLOB1 promoter in citrus. Plant Biotechnol. J. 2017, 15, 1509–1519. [Google Scholar] [CrossRef] [PubMed]
- Jia, H.; Orbović, V.; Wang, N. CRISPR-LbCas12a-mediated modification of citrus. Plant Biotechnol. J. 2019. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Chen, S.; Peng, A.; Xie, Z.; He, Y.; Zou, X. CRISPR/Cas9-mediated editing of CsWRKY22 reduces susceptibility to Xanthomonas citri subsp. citri in Wanjincheng orange (Citrus sinensis (L.) Osbeck). Plant Biotechnol. Rep. 2019, 1–10. [Google Scholar] [CrossRef]
- Gottwald, T.R.; Graham, J.H.; Schubert, T.S. Citrus canker: The pathogen and its impact. Plant Health Progress 2002, 10, 32. [Google Scholar] [CrossRef]
- Das, A.K. Citrus canker-A review. J. Appl. Hort. 2003, 5, 52–60. [Google Scholar]
- Dutta, S.; Bendre, R.; Padhye, S.; Ahmed, F.; Sarkar, F. Synthesis, antioxidant properties and antiproliferative activities of tetrameric copper and copper-zinc metal complexes of catecholamine Schiff base ligand. Synth. React. Inorg. M. 2005, 35, 3–10. [Google Scholar] [CrossRef]
- Rinaldi, D.; Leite, R., Jr. Adaptation of Xanthomonas axonopodis pv. citri population to the presence of copper compounds in nature. Proc. Int. Soc. Citric 2000, 2, 1064. [Google Scholar]
- Behlau, F.; Belasque, J., Jr.; Bergamin Filho, A.; Graham, J.; Leite, R., Jr.; Gottwald, T. Copper sprays and windbreaks for control of citrus canker on young orange trees in southern Brazil. Crop Prot. 2008, 27, 807–813. [Google Scholar] [CrossRef]
- Zhang, X.; Francis, M.I.; Dawson, W.O.; Graham, J.H.; Orbović, V.; Triplett, E.W.; Mou, Z. Over-expression of the Arabidopsis NPR1 gene in citrus increases resistance to citrus canker. Eur. J. Plant Pathol. 2010, 128, 91–100. [Google Scholar] [CrossRef]
- Zasloff, M. Antibiotic peptides as mediators of innate immunity. Curr. Opin. Immunol. 1992, 4, 3–7. [Google Scholar] [CrossRef]
- Boman, H. Antibacterial peptides: Basic facts and emerging concepts. J. Intern. Med. 2003, 254, 197–215. [Google Scholar] [CrossRef] [PubMed]
- Boscariol, R.L.; Monteiro, M.; Takahashi, E.K.; Chabregas, S.M.; Vieira, M.L.C.; Vieira, L.G.E.; Pereira, L.F.P.; Mourao Filho, F.D.A.A.; Cardoso, S.C.; Christiano, R.S.C. Attacin A gene from Tricloplusia ni reduces susceptibility to Xanthomonas axonopodis pv. citri in transgenic Citrus sinensis ‘Hamlin’. J. Am. Soc. Hortic. Sci. 2006, 131, 530–536. [Google Scholar] [CrossRef]
- Cardoso, S.C.; Barbosa-Mendes, J.M.; Boscariol-Camargo, R.L.; Christiano, R.S.C.; Filho, A.B.; Vieira, M.L.C.; Mendes, B.M.J.; Mourão Filho, F.D.A.A. Transgenic Sweet Orange (Citrus sinensis L. Osbeck) Expressing the attacin A Gene for Resistance to Xanthomonas citri subsp. citri. Plant Mol. Biol. Rep. 2009, 28, 185–192. [Google Scholar] [CrossRef]
- Sala Junior, V.; Celloto, V.R.; Vieira, L.G.E.; Gonçalves, J.E.; Gonçalves, R.A.C.; de Oliveira, A.J.B. Floral nectar chemical composition of floral nectar in conventional and transgenic sweet orange, Citrus sinensis (L.) Osbeck, expressing an antibacterial peptide. Plant Syst. Evol. 2008, 275, 1–7. [Google Scholar] [CrossRef]
- Kobayashi, A.K.; Vieira, L.G.E.; Bespalhok Filho, J.C.; Leite, R.P.; Pereira, L.F.P.; Molinari, H.B.C.; Marques, V.V. Enhanced resistance to citrus canker in transgenic sweet orange expressing the sarcotoxin IA gene. Eur. J. Plant Pathol. 2017, 149, 865–873. [Google Scholar] [CrossRef] [Green Version]
- Hao, G.; Zhang, S.; Stover, E. Transgenic expression of antimicrobial peptide D2A21 confers resistance to diseases incited by Pseudomonas syringae pv. tabaci and Xanthomonas citri, but not Candidatus Liberibacter asiaticus. PLoS ONE 2017, 12, e0186810. [Google Scholar]
- Furman, N.; Kobayashi, K.; Zanek, M.C.; Calcagno, J.; Garcia, M.L.; Mentaberry, A. Transgenic sweet orange plants expressing a dermaseptin coding sequence show reduced symptoms of citrus canker disease. J. Biotechnol. 2013, 167, 412–419. [Google Scholar] [CrossRef]
- Chern, M.; Fitzgerald, H.A.; Canlas, P.E.; Navarre, D.A.; Ronald, P.C. Over expression of a rice NPR1 homolog leads to constitutive activation of defense response and hypersensitivity to light. Mol. Plant Microbe Interact. 2005, 18, 511–520. [Google Scholar] [CrossRef]
- Lin, W.C.; Lu, C.F.; Wu, J.W.; Cheng, M.L.; Lin, Y.M.; Yang, N.S.; Black, L.; Green, S.K.; Wang, J.F.; Cheng, C.P. Transgenic tomato plants expressing the Arabidopsis NPR1 gene display enhanced resistance to a spectrum of fungal and bacterial diseases. Transgenic Res. 2004, 13, 567–581. [Google Scholar] [CrossRef]
- Makandar, R.; Essig, J.S.; Schapaugh, M.A.; Trick, H.N.; Shah, J. Genetically engineered resistance to Fusarium head blight in wheat by expression of Arabidopsis NPR1. Mol. Plant Microbe Interact. 2006, 19, 123–129. [Google Scholar] [CrossRef] [PubMed]
- Malnoy, M.; Jin, Q.; Borejsza-Wysocka, E.; He, S.; Aldwinckle, H. Over expression of the apple MpNPR1 gene confers increased disease resistance in Malus × domestica. Mol. Plant Microbe Interact. 2007, 20, 1568–1580. [Google Scholar] [CrossRef] [PubMed]
- Potlakayala, S.D.; DeLong, C.; Sharpe, A.; Fobert, P.R. Conservation of NON-EXPRESSOR OF PATHOGENESIS-RELATED GENES1 function between Arabidopsis thaliana and Brassica napus. Physiol. Mol. Plant Pathol. 2007, 71, 174–183. [Google Scholar] [CrossRef]
- Chen, X.; Barnaby, J.Y.; Sreedharan, A.; Huang, X.; Orbović, V.; Grosser, J.W.; Wang, N.; Dong, X.; Song, W.Y. Over-expression of the citrus gene CtNH1 confers resistance to bacterial canker disease. Physiol. Mol. Plant Pathol. 2013, 84, 115–122. [Google Scholar] [CrossRef]
- Flor, H.H. Current status of the gene-for-gene concept. Annu. Rev. Phytopathol. 1971, 9, 275–296. [Google Scholar] [CrossRef]
- Tai, T.H.; Dahlbeck, D.; Clark, E.T.; Gajiwala, P.; Pasion, R.; Whalen, M.C.; Stall, R.E.; Staskawicz, B.J. Expression of the Bs2 pepper gene confers resistance to bacterial spot disease in tomato. Proc. Natl. Acad. Sci. USA 1999, 96, 14153–14158. [Google Scholar] [CrossRef]
- Sendín, L.; Filippone, M.; Orce, I.; Rigano, L.; Enrique, R.; Peña, L.; Vojnov, A.; Marano, M.; Castagnaro, A. Transient expression of pepper Bs2 gene in Citrus limon as an approach to evaluate its utility for management of citrus canker disease. Plant Pathol. 2012, 61, 648–657. [Google Scholar] [CrossRef]
- Sendín, L.N.; Orce, I.G.; Gómez, R.L.; Enrique, R.; Bournonville, C.F.G.; Noguera, A.S.; Vojnov, A.A.; Marano, M.R.; Castagnaro, A.P.; Filippone, M.P. Inducible expression of Bs2 R gene from Capsicum chacoense in sweet orange (Citrus sinensis L. Osbeck) confers enhanced resistance to citrus canker disease. Plant Mol. Biol. 2017, 93, 607–621. [Google Scholar] [CrossRef]
- Song, W.Y.; Wang, G.L.; Chen, L.L.; Kim, H.S.; Pi, L.Y.; Holsten, T.; Gardner, J.; Wang, B.; Zhai, W.X.; Zhu, L.H. A receptor kinase-like protein encoded by the rice disease resistance gene, Xa21. Science 1995, 270, 1804–1806. [Google Scholar] [CrossRef]
- Omar, A.; Song, W.Y.; Grosser, J. Introduction of Xa21, a Xanthomonas-resistance gene from rice, into ‘Hamlin’sweet orange [Citrus sinensis (L.) Osbeck] using protoplast-GFP co-transformation or single plasmid transformation. J. Hortic. Sci. Biotech. 2007, 82, 914–923. [Google Scholar] [CrossRef]
- Mendes, B.M.J.; Cardoso, S.; Boscariol-Camargo, R.; Cruz, R.; Mourão Filho, F.; Bergamin Filho, A. Reduction in susceptibility to Xanthomonas axonopodis pv. citri in transgenic Citrus sinensis expressing the rice Xa21 gene. Plant Pathol. 2010, 59, 68–75. [Google Scholar]
- Li, D.L.; Xiao, X.; Guo, W.W. Production of Transgenic Anliucheng Sweet Orange (Citrus sinensis Osbeck) with Xa21 Gene for Potential Canker Resistance. J. Integr. Agr. 2014, 13, 2370–2377. [Google Scholar] [CrossRef]
- Omar, A.A.; Murata, M.M.; El-Shamy, H.A.; Graham, J.H.; Grosser, J.W. Enhanced resistance to citrus canker in transgenic mandarin expressing Xa21 from rice. Transgenic Res. 2018, 27, 179–191. [Google Scholar] [CrossRef] [PubMed]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef]
- Gechev, T.S.; Van Breusegem, F.; Stone, J.M.; Denev, I.; Laloi, C. Reactive oxygen species as signals that modulate plant stress responses and programmed cell death. Bioessays 2006, 28, 1091–1101. [Google Scholar] [CrossRef]
- Hao, G.; Pitino, M.; Duan, Y.; Stover, E. Reduced susceptibility to Xanthomonas citri in transgenic citrus expressing the FLS2 receptor from Nicotiana benthamiana. Mol. Plant Microbe Interact. 2016, 29, 132–142. [Google Scholar] [CrossRef]
- De Oliveira, M.L.; de Lima Silva, C.C.; Abe, V.Y.; Costa, M.G.; Cernadas, R.A.; Benedetti, C.E. Increased resistance against citrus canker mediated by a citrus mitogen-activated protein kinase. Mol. Plant Microbe Interact. 2013, 26, 1190–1199. [Google Scholar] [CrossRef]
- Barbosa-Mendes, J.M.; Filho, F.D.A.A.M.; Filho, A.B.; Harakava, R.; Beer, S.V.; Mendes, B.M.J. Genetic transformation of Citrus sinensis cv. Hamlin with hrpN gene from Erwinia amylovora and evaluation of the transgenic lines for resistance to citrus canker. Sci. Hortic. Amst. 2009, 122, 109–115. [Google Scholar] [CrossRef]
- Fu, X.Z.; Chen, C.W.; Wang, Y.; Liu, J.H.; Moriguchi, T. Ectopic expression of MdSPDS1 in sweet orange (Citrus sinensis Osbeck) reduces canker susceptibility: Involvement of H2O2 production and transcriptional alteration. BMC Plant Biol. 2011, 11, 55. [Google Scholar] [CrossRef]
- Qin, L.; Luo, S.; Deng, Z.; Yan, J.; Li, N.; Yuan, F. Enhanced resistance to canker disease and tolerances to biotic and abiotic stresses in terf1 transgenic sweet orange. In II International Symposium on Citrus Biotechnology; Gentile, A., La Malfa, S., Eds.; International Society for Horticultural Science: Catania, Italy, 2009; pp. 165–172. [Google Scholar]
- Yang, L.; Hu, C.; Li, N.; Zhang, J.; Yan, J.; Deng, Z. Transformation of sweet orange [Citrus sinensis (L.) Osbeck] with pthA-nls for acquiring resistance to citrus canker disease. Plant Mol. Biol. 2011, 75, 11–23. [Google Scholar] [CrossRef]
- Hao, G.; Stover, E.; Gupta, G. Overexpression of a modified plant thionin enhances disease Resistance to citrus canker and huanglongbing (HLB). Front. Plant Sci. 2016, 7, 1078. [Google Scholar] [CrossRef] [PubMed]
- Bové, J.M. Huanglongbing: A destructive, newly-emerging, century-old disease of citrus. J. Plant Pathol. 2006, 88, 7–37. [Google Scholar]
- Robertson, C.J.; Zhang, X.; Gowda, S.; Orbović, V.; Dawson, W.O.; Mou, Z. Overexpression of the Arabidopsis NPR1 protein in citrus confers tolerance to Huanglongbing. J. Citrus Pathol. 2018, 5, 1–8. [Google Scholar]
- Munir, S.; He, P.; Wu, Y.; He, P.; Khan, S.; Huang, M.; Cui, W.; He, P.; He, Y. Huanglongbing control: Perhaps the end of the beginning. Microb. Ecol. 2018, 76, 192–204. [Google Scholar] [CrossRef]
- Gottwald, T.R. Current epidemiological understanding of citrus huanglongbing. Annu. Rev. Phytopathol. 2010, 48, 119–139. [Google Scholar] [CrossRef]
- Wang, N.; Trivedi, P. Citrus huanglongbing: A newly relevant disease presents unprecedented challenges. Phytopathology 2013, 103, 652–665. [Google Scholar] [CrossRef]
- Da Graça, J.; Korsten, L. Citrus huanglongbing: Review, present status and future strategies. In Diseases of Fruits and Vegetables Volume I; Naqvi, S.A.M.H., Ed.; Springer: Dordrecht, The Netherlands, 2004; pp. 229–245. [Google Scholar]
- Zhang, M.; Guo, Y.; Powell, C.A.; Doud, M.S.; Yang, C.; Duan, Y. Effective antibiotics against ‘Candidatus Liberibacter asiaticus’ in HLB-affected citrus plants identified via the graft-based evaluation. PLoS ONE 2014, 9, e111032. [Google Scholar] [CrossRef]
- Miles, G.P.; Stover, E.; Ramadugu, C.; Keremane, M.L.; Lee, R.F. Apparent tolerance to Huanglongbing in Citrus and Citrus-related germplasm. HortScience 2017, 52, 31–39. [Google Scholar] [CrossRef]
- Wang, Y.; Zhou, L.; Yu, X.; Stover, E.; Luo, F.; Duan, Y. Transcriptome profiling of Huanglongbing (HLB) tolerant and susceptible citrus plants reveals the role of basal resistance in HLB tolerance. Front. Plant Sci. 2016, 7, 933. [Google Scholar] [CrossRef]
- Felipe, R.T.A.; Mourão Filho, F.D.A.A.; Lopes, S.A.; Mendes, B.M.J.; Behling, M.; Pereira Junior, E.V. Reaction of sweet orange cultivars expressing the attacin A gene to ‘Candidatus Liberibacter asiaticus’ infection. Pesqui. Agropecu. Bras. 2013, 48, 1440–1448. [Google Scholar] [CrossRef]
- Rocha Tavano, E.C.D.; Erpen, L.; Aluisi, B.; Harakava, R.; Lopes, J.R.S.; Vieira, M.L.C.; Piedade, S.M.D.S.; Mendes, B.M.J.; Mourao Filho, A.A. Sweet orange genetic transformation with the attacin A gene under the control of phloem-specific promoters and inoculation with Candidatus Liberibacter asiaticus. J. Hortic. Sci. Biotech. 2019, 94, 210–219. [Google Scholar] [CrossRef]
- Zou, X.; Jiang, X.; Xu, L.; Lei, T.; Peng, A.; He, Y.; Yao, L.; Chen, S. Transgenic citrus expressing synthesized cecropin B genes in the phloem exhibits decreased susceptibility to Huanglongbing. Plant Mol. Biol. 2017, 93, 341–353. [Google Scholar] [CrossRef] [PubMed]
- Bar-Joseph, M.; Marcus, R.; Lee, R.F. The continuous challenge of citrus tristeza virus control. Annu. Rev. Phytopathol. 1989, 27, 291–316. [Google Scholar] [CrossRef]
- Dominguez, A.; Guerri, J.; Cambra, M.; Navarro, L.; Moreno, P.; Pena, L. Efficient production of transgenic citrus plants expressing the coat protein gene of citrus tristeza virus. Plant Cell Rep. 2000, 19, 427–433. [Google Scholar] [CrossRef]
- Domínguez, A.; de Mendoza, A.H.; Guerri, J.; Cambra, M.; Navarro, L.; Moreno, P.; Peña, L. Pathogen-derived resistance to Citrus tristeza virus (CTV) in transgenic Mexican lime (Citrus aurantifolia (Christ.) Swing.) plants expressing its p25 coat protein gene. Mol. Breed. 2002, 10, 1–10. [Google Scholar]
- Fagoaga, C.; López, C.; de Mendoza, A.H.; Moreno, P.; Navarro, L.; Flores, R.; Peña, L. Post-transcriptional gene silencing of the p23 silencing suppressor of Citrus tristeza virus confers resistance to the virus in transgenic Mexican lime. Plant Mol. Biol. 2006, 60, 153–165. [Google Scholar] [CrossRef]
- Ananthakrishnan, G.; Orbović, V.; Pasquali, G.; Ćalović, M.; Grosser, J. Transfer of citrus tristeza virus (CTV)-derived resistance candidate sequences to four grapefruit cultivars through Agrobacterium-mediated genetic transformation. In Vitro Cell. Dev. Plant 2007, 43, 593–601. [Google Scholar] [CrossRef]
- Muniz, F.; De Souza, A.; Stipp, L.C.L.; Schinor, E.; Freitas, W.; Harakava, R.; Stach-Machado, D.; Rezende, J.A.M.; Mourão Filho, F.; Mendes, B. Genetic transformation of Citrus sinensis with Citrus tristeza virus (CTV) derived sequences and reaction of transgenic lines to CTV infection. Biol. Plant. 2012, 56, 162–166. [Google Scholar] [CrossRef]
- Olivares-Fuster, O.; Fleming, G.; Albiach-Marti, M.; Gowda, S.; Dawson, W.; Crosser, J. Citrus tristeza virus (CTV) resistance in transgenic citrus based on virus challenge of protoplasts. In Vitro Cell. Dev. Plant 2003, 39, 567–572. [Google Scholar] [CrossRef]
- Cervera, M.; Esteban, O.; Gil, M.; Gorris, M.T.; Martínez, M.C.; Peña, L.; Cambra, M. Transgenic expression in citrus of single-chain antibody fragments specific to Citrus tristeza virus confers virus resistance. Transgenic Res. 2010, 19, 1001–1015. [Google Scholar] [CrossRef]
- Waterhouse, P.M.; Wang, M.B.; Lough, T. Gene silencing as an adaptive defence against viruses. Nature 2001, 411, 834. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.; Zhang, Y.; Yang, J.; Zhong, Y. Expression of hairpin RNA (hpRNA) targeting the three CTV-silencing suppressor genes confers sweet orange with stem-pitting CTV tolerance. J. Hortic. Sci. Biotechnol. 2017, 92, 465–474. [Google Scholar] [CrossRef]
- Cheng, C.Z.; Yang, J.W.; Yan, H.B.; Bei, X.J.; Zhang, Y.Y.; Lu, Z.M.; Zhong, G.Y. Expressing p20 hairpin RNA of Citrus tristeza virus confers Citrus aurantium with tolerance/resistance against stem pitting and seedling yellow CTV strains. J. Integr. Agric. 2015, 14, 1767–1777. [Google Scholar] [CrossRef]
- Kayim, M.; Barthe, G.; Beretta, J.; Derrick, K. Transgenic citrus plants expressing the coat protein gene of Citrus psorosis virus. Phytopathology 2005, 95, S52. [Google Scholar]
- Reyes, C.A.; Zanek, M.C.; Velázquez, K.; Costa, N.; Plata, M.I.; Garcia, M.L. Generation of sweet orange transgenic lines and evaluation of citrus psorosis virus-derived resistance against psorosis A and psorosis B. J. Phytopathol. 2011, 159, 531–537. [Google Scholar] [CrossRef]
- Reyes, C.A.; De Francesco, A.; Peña, E.J.; Costa, N.; Plata, M.I.; Sendin, L.; Castagnaro, A.P.; García, M.L. Resistance to Citrus psorosis virus in transgenic sweet orange plants is triggered by coat protein–RNA silencing. J. Biotechnol. 2011, 151, 151–158. [Google Scholar] [CrossRef]
- Gentile, A.; Deng, Z.; La Malfa, S.; Distefano, G.; Domina, F.; Vitale, A.; Polizzi, G.; Lorito, M.; Tribulato, E. Enhanced resistance to Phoma tracheiphila and Botrytis cinerea in transgenic lemon plants expressing a Trichoderma harzianum chitinase gene. Plant Breed. 2007, 126, 146–151. [Google Scholar] [CrossRef]
- Distefano, G.; La Malfa, S.; Vitale, A.; Lorito, M.; Deng, Z.; Gentile, A. Defence-related gene expression in transgenic lemon plants producing an antimicrobial Trichoderma harzianum endochitinase during fungal infection. Transgenic Res. 2008, 17, 873–879. [Google Scholar] [CrossRef]
- Fagoaga, C.; Rodrigo, I.; Conejero, V.; Hinarejos, C.; Tuset, J.J.; Arnau, J.; Pina, J.A.; Navarro, L.; Peña, L. Increased tolerance to Phytophthora citrophthora in transgenic orange plants constitutively expressing a tomato pathogenesis related protein PR-5. Mol. Breed. 2001, 7, 175–185. [Google Scholar] [CrossRef]
- Mondal, S.; Dutt, M.; Grosser, J.; Dewdney, M. Transgenic citrus expressing the antimicrobial gene AttacinE (attE) reduces the susceptibility of ‘Duncan’ grapefruit to the citrus scab caused by Elsinoë fawcettii. Eur. J. Plant Pathol. 2012, 133, 391–404. [Google Scholar] [CrossRef]
- Rodríguez, A.; Andrés, V.S.; Cervera, M.; Redondo, A.; Alquézar, B.; Shimada, T.; Gadea, J.; Rodrigo, M.; Zacarías, L.; Palou, L. The monoterpene limonene in orange peels attracts pests and microorganisms. Plant Signal. Behav. 2011, 6, 1820–1823. [Google Scholar] [CrossRef] [Green Version]
- Parmar, N.; Singh, K.H.; Sharma, D.; Singh, L.; Kumar, P.; Nanjundan, J.; Khan, Y.J.; Chauhan, D.K.; Thakur, A.K. Genetic engineering strategies for biotic and abiotic stress tolerance and quality enhancement in horticultural crops: A comprehensive review. 3 Biotech 2017, 7, 239. [Google Scholar] [CrossRef]
- Jansing, J.; Schiermeyer, A.; Schillberg, S.; Fischer, R.; Bortesi, L. Genome Editing in Agriculture: Technical and Practical Considerations. Int. J. Mol. Sci. 2019, 20, 2888. [Google Scholar] [CrossRef]
- Pennisi, E. The CRISPR Craze. Science 2013, 341, 833–836. [Google Scholar] [CrossRef]
- Jiang, W.; Zhou, H.; Bi, H.; Fromm, M.; Yang, B.; Weeks, D.P. Demonstration of CRISPR/Cas9/sgRNA-mediated targeted gene modification in Arabidopsis, tobacco, sorghum and rice. Nucleic Acids Res. 2013, 41, e188. [Google Scholar] [CrossRef]
- Wang, Y.; Cheng, X.; Shan, Q.; Zhang, Y.; Liu, J.; Gao, C.; Qiu, J.L. Simultaneous editing of three homoeoalleles in hexaploid bread wheat confers heritable resistance to powdery mildew. Nat. Biotechnol. 2014, 32, 947. [Google Scholar] [CrossRef]
- Char, S.N.; Neelakandan, A.K.; Nahampun, H.; Frame, B.; Main, M.; Spalding, M.H.; Becraft, P.W.; Meyers, B.C.; Walbot, V.; Wang, K. An Agrobacterium-delivered CRISPR/Cas9 system for high-frequency targeted mutagenesis in maize. Plant Biotechnol. J. 2017, 15, 257–268. [Google Scholar] [CrossRef]
- Nishitani, C.; Hirai, N.; Komori, S.; Wada, M.; Okada, K.; Osakabe, K.; Yamamoto, T.; Osakabe, Y. Efficient genome editing in apple using a CRISPR/Cas9 system. Sci. Rep. 2016, 6, 31481. [Google Scholar] [CrossRef]
- Ren, C.; Liu, X.; Zhang, Z.; Wang, Y.; Duan, W.; Li, S.; Liang, Z. CRISPR/Cas9-mediated efficient targeted mutagenesis in Chardonnay (Vitis vinifera L.). Sci. Rep. 2016, 6, 32289. [Google Scholar] [CrossRef]
- Song, G.; Jia, M.; Chen, K.; Kong, X.; Khattak, B.; Xie, C.; Li, A.; Mao, L. CRISPR/Cas9: A powerful tool for crop genome editing. Crop J. 2016, 4, 75–82. [Google Scholar] [CrossRef] [Green Version]
- Jia, H.; Zou, X.; Orbovic, V.; Wang, N. Genome Editing in Citrus Tree with CRISPR/Cas9. In Plant Genome Editing with CRISPR Systems; Qi, Y., Ed.; Methods in Molecular Biology; Humana Press: New York, NY, USA, 2019; Volume 1917, pp. 235–241. [Google Scholar]
- Xu, Z.S.; Feng, K.; Xiong, A.S. CRISPR/Cas9-mediated multiply targeted mutagenesis in orange and purple carrot plants. Mol. Biotechnol. 2019, 61, 191–199. [Google Scholar] [CrossRef]
- Knott, G.J.; Doudna, J.A.J.S. CRISPR-Cas guides the future of genetic engineering. Science 2018, 361, 866–869. [Google Scholar] [CrossRef] [Green Version]
- Ali, Z.; Abulfaraj, A.; Idris, A.; Ali, S.; Tashkandi, M.; Mahfouz, M.M. CRISPR/Cas9-mediated viral interference in plants. Genome Biol. 2015, 16, 238. [Google Scholar] [CrossRef] [Green Version]
- Baltes, N.J.; Hummel, A.W.; Konecna, E.; Cegan, R.; Bruns, A.N.; Bisaro, D.M.; Voytas, D.F. Conferring resistance to geminiviruses with the CRISPR–Cas prokaryotic immune system. Nat. Plants 2015, 1, 15145. [Google Scholar] [CrossRef]
- Malnoy, M.; Viola, R.; Jung, M.H.; Koo, O.J.; Kim, S.; Kim, J.S.; Velasco, R.; Nagamangala Kanchiswamy, C. DNA-free genetically edited grapevine and apple protoplast using CRISPR/Cas9 ribonucleoproteins. Front. Plant Sci. 2016, 7, 1904. [Google Scholar] [CrossRef]
- Jia, H.; Wang, N. Xcc-facilitated agroinfiltration of citrus leaves: A tool for rapid functional analysis of transgenes in citrus leaves. Plant Cell Rep. 2014, 33, 1993–2001. [Google Scholar] [CrossRef]
- Hu, Y.; Zhang, J.; Jia, H.; Sosso, D.; Li, T.; Frommer, W.B.; Yang, B.; White, F.F.; Wang, N.; Jones, J.B. Lateral organ boundaries 1 is a disease susceptibility gene for citrus bacterial canker disease. Proc. Natl. Acad. Sci. USA 2014, 111, E521–E529. [Google Scholar] [CrossRef]
- Ledford, H. Geneticists enlist engineered virus and CRISPR to battle citrus disease. Nat. News 2017, 545, 277. [Google Scholar] [CrossRef]
- Wang, N.; Pierson, E.A.; Setubal, J.C.; Xu, J.; Levy, J.G.; Zhang, Y.; Li, J.; Rangel, L.T.; Martins, J., Jr. The Candidatus Liberibacter–host interface: Insights into pathogenesis mechanisms and disease control. Annu. Rev. Phytopathol. 2017, 55, 451–482. [Google Scholar] [CrossRef]
- Zhang, Y.; Malzahn, A.A.; Sretenovic, S.; Qi, Y. The emerging and uncultivated potential of CRISPR technology in plant science. Nat. Plants 2019, 5, 778–794. [Google Scholar] [CrossRef]
- Zhu, C.; Zheng, X.; Huang, Y.; Ye, J.; Chen, P.; Zhang, C.; Zhao, F.; Xie, Z.; Zhang, S.; Wang, N. Genome sequencing and CRISPR/Cas9 gene editing of an early flowering Mini-Citrus (Fortunella hindsii). Plant Biotechnol. J. 2019, 17, 2199–2210. [Google Scholar] [CrossRef]
- Hunter, W.B.; Sinisterra-Hunter, X.H. Emerging RNA Suppression Technologies to Protect Citrus Trees from Citrus Greening Disease Bacteria. Adv. Insect Physiol. 2018, 55, 163–197. [Google Scholar]
- Goulin, E.H.; Galdeano, D.M.; Granato, L.M.; Matsumura, E.E.; Dalio, R.J.D.; Machado, M.A. RNA interference and CRISPR: Promising approaches to better understand and control citrus pathogens. Microbiol. Res. 2019, 226, 1–9. [Google Scholar] [CrossRef]
- Andrade, C.; Hunter, W.B. RNA interference-natural gene-based technology for highly specific pest control (HiSPeC). RNA Interf. 2016, 391–409. [Google Scholar] [CrossRef]
- Andrade, E.C.; Hunter, W.B. RNAi feeding bioassay: Development of a non-transgenic approach to control Asian citrus psyllid and other hemipterans. Entomol. Exp. Appl. 2017, 162, 389–396. [Google Scholar] [CrossRef]
- Galdeano, D.M.; Breton, M.C.; Lopes, J.R.S.; Falk, B.W.; Machado, M.A. Oral delivery of double-stranded RNAs induces mortality in nymphs and adults of the Asian citrus psyllid, Diaphorina citri. PLoS ONE 2017, 12, e0171847. [Google Scholar] [CrossRef]
- Ghosh, S.K.B.; Hunter, W.B.; Park, A.L.; Gundersen-Rindal, D.E. Double strand RNA delivery system for plant-sap-feeding insects. PLoS ONE 2017, 12, e0171861. [Google Scholar] [CrossRef]
- Ghosh, S.K.B.; Hunter, W.B.; Park, A.L.; Gundersen-Rindal, D.E. Double-stranded RNA oral delivery methods to induce RNA interference in phloem and plant-sap-feeding hemipteran insects. JoVE J. Vis. Exp. 2018, 135, e57390. [Google Scholar] [CrossRef]
- Kruse, A.; Fattah-Hosseini, S.; Saha, S.; Johnson, R.; Warwick, E.; Sturgeon, K.; Mueller, L.; MacCoss, M.J.; Shatters Jr, R.G.; Heck, M.C. Combining’omics and microscopy to visualize interactions between the Asian citrus psyllid vector and the Huanglongbing pathogen Candidatus Liberibacter asiaticus in the insect gut. PLoS ONE 2017, 12, e0179531. [Google Scholar] [CrossRef]
- Taning, C.N.; Andrade, E.C.; Hunter, W.B.; Christiaens, O.; Smagghe, G. Asian Citrus Psyllid RNAi Pathway-RNAi evidence. Sci. Rep. 2016, 6, 38082. [Google Scholar] [CrossRef]
- Ammar, E.D.; Walter, A.J.; Hall, D.G. New excised-leaf assay method to test inoculativity of Asian citrus psyllid (Hemiptera: Psyllidae) with Candidatus Liberibacter asiaticus associated with citrus huanglongbing disease. J. Econ. Entomol. 2013, 106, 25–35. [Google Scholar] [CrossRef]
- Raiol-Junior, L.L.; Baia, A.D.; Luiz, F.Q.; Fassini, C.G.; Marques, V.V.; Lopes, S.A. Improvement in the Excised Citrus Leaf Assay to Investigate Inoculation of ‘Candidatus Liberibacter asiaticus’ by the Asian Citrus Psyllid Diaphorina citri. Plant Dis. 2017, 101, 409–413. [Google Scholar] [CrossRef]
- Russell, C.W.; Pelz-Stelinski, K.S. Development of an artificial diet and feeding system for juvenile stages of the A sian citrus psyllid, Diaphorina citri. Entomol. Exp. Appl. 2015, 154, 171–176. [Google Scholar] [CrossRef]
- Killiny, N.; Hajeri, S.; Tiwari, S.; Gowda, S.; Stelinski, L.L. Double-stranded RNA uptake through topical application, mediates silencing of five CYP4 genes and suppresses insecticide resistance in Diaphorina citri. PLoS ONE 2014, 9, e110536. [Google Scholar] [CrossRef]
- Killiny, N.; Kishk, A. Delivery of dsRNA through topical feeding for RNA interference in the citrus sap piercing-sucking hemipteran, Diaphorina citri. Arch. Insect Biochem. 2017, 95, e21394. [Google Scholar] [CrossRef]
- Yu, X.; Gowda, S.; Killiny, N. Double-stranded RNA delivery through soaking mediates silencing of the muscle protein 20 and increases mortality to the Asian citrus psyllid, Diaphorina citri. Pest Manag. Sci. 2017, 73, 1846–1853. [Google Scholar] [CrossRef]
- Zhang, L.; Reed, R.D. A Practical Guide to CRISPR/Cas9 Genome Editing in Lepidoptera. In Diversity and Evolution of Butterfly Wing Patterns; Sekimura, T., Nijhout, H., Eds.; Springer: Singapore, 2017; pp. 155–172. [Google Scholar]
- Zheng, Z.; Bao, M.; Wu, F.; Chen, J.; Deng, X. Predominance of single prophage carrying a CRISPR/cas system in “Candidatus Liberibacter asiaticus” strains in southern China. PLoS ONE 2016, 11, e0146422. [Google Scholar] [CrossRef]
- Shang, F.; Xiong, Y.; Xia, W.K.; Wei, D.D.; Wei, D.; Wang, J.J. Identification, characterization and functional analysis of a chitin synthase gene in the brown citrus aphid, Toxoptera citricida (Hemiptera, Aphididae). Insect Mol. Biol. 2016, 25, 422–430. [Google Scholar] [CrossRef]
- Jing, T.X.; Tan, Y.; Ding, B.Y.; Dou, W.; Wei, D.; Wang, J.J. NADPH–cytochrome P450 reductase mediates the resistance of Aphis (Toxoptera) citricidus (Kirkaldy) to abamectin. Front. Plant Sci. 2018, 9, 986. [Google Scholar] [CrossRef]
- Rosa, C.; Kamita, S.G.; Dequine, H.; Wuriyanghan, H.; Lindbo, J.A.; Falk, B.W. RNAi effects on actin mRNAs in Homalodisca vitripennis cells. J. RNAi Gene Silenc. 2010, 6, 361. [Google Scholar]
- Rosa, C.; Kamita, S.G.; Falk, B.W. RNA interference is induced in the glassy winged sharpshooter Homalodisca vitripennis by actin dsRNA. Pest Manag. Sci. 2012, 68, 995–1002. [Google Scholar] [CrossRef]
- Roeschlin, R.A.; Uviedo, F.; García, L.; Molina, M.C.; Favaro, M.A.; Chiesa, M.A.; Tasselli, S.; Franco-Zorrilla, J.M.; Forment, J.; Gadea, J.; et al. PthA4AT, a 7.5-repeats transcription activator-like (TAL) effector from Xanthomonas citri ssp. citri, triggers citrus canker resistance. Mol. Plant Pathol. 2019, 20, 1394–1407. [Google Scholar] [CrossRef]
- Shimo, H.M.; Terassi, C.; Lima Silva, C.C.; de Lima Zanella, J.; Mercaldi, G.F.; Rocco, S.A.; Benedetti, C.E. Role of the Citrus sinensis RNA deadenylase CsCAF1 in citrus canker resistance. Mol. Plant Pathol. 2019, 20, 1105–1118. [Google Scholar] [CrossRef]
- Glandorf, D.; Bakker, P.; Van Loon, L.J. Influence of the production of antibacterial and antifungal proteins by transgenic plants on the saprophytic soil microflora. Acta Bot. Neerl. 1997, 46, 85–104. [Google Scholar] [CrossRef]
- Bulgarelli, D.J.S. How manipulating the plant microbiome could improve agriculture. Scientist 2018, 32, 51422. [Google Scholar]
- Ageitos, J.M.; Sanchez-Perez, A.; Calo-Mata, P.; Villa, T.G. Antimicrobial peptides (AMPs): Ancient compounds that represent novel weapons in the fight against bacteria. Biochem. Pharmacol. 2017, 133, 117–138. [Google Scholar] [CrossRef]
- Weinhold, A.; Dorcheh, E.K.; Li, R.; Rameshkumar, N.; Baldwin, I.T. Antimicrobial peptide expression in a wild tobacco plant reveals the limits of host-microbe-manipulations in the field. eLife 2018, 7, e28715. [Google Scholar] [CrossRef]
- Cunningham, F.J.; Goh, N.S.; Demirer, G.S.; Matos, J.L.; Landry, M.P. Nanoparticle-mediated delivery towards advancing plant genetic engineering. Trends Biotechnol. 2018, 36, 882–897. [Google Scholar] [CrossRef]
- Wang, J.W.; Grandio, E.G.; Newkirk, G.M.; Demirer, G.S.; Butrus, S.; Giraldo, J.P.; Landry, M.P. Nanoparticle-Mediated Genetic Engineering of Plants. Mol. Plant 2019, 12, 1037–1040. [Google Scholar] [CrossRef]
- Dyab, A.K.; Mohamed, L.A.; Taha, F. Non-aqueous olive oil-in-glycerin (o/o) Pickering emulsions: Preparation, characterization and in vitro aspirin release. J. Disper. Sci. Technol. 2018, 39, 890–900. [Google Scholar] [CrossRef]
- Naqvi, S.; Maitra, A.; Abdin, M.; Akmal, M.; Arora, I.; Samim, M. Calcium phosphate nanoparticle mediated genetic transformation in plants. J. Mater. Chem. 2012, 22, 3500–3507. [Google Scholar] [CrossRef]
- Pasupathy, K.; Lin, S.; Hu, Q.; Luo, H.; Ke, P.C. Direct plant gene delivery with a poly (amidoamine) dendrimer. Biotechnol. J. Healthc. Nutr. Technol. 2008, 3, 1078–1082. [Google Scholar] [CrossRef]
- Burlaka, O.; Pirko, Y.V.; Yemets, A.; Blume, Y.B. Plant genetic transformation using carbon nanotubes for DNA delivery. Cytol. Genet. 2015, 49, 349–357. [Google Scholar] [CrossRef]
- Chang, F.P.; Kuang, L.Y.; Huang, C.A.; Jane, W.N.; Hung, Y.; Yue-ie, C.H.; Mou, C.Y. A simple plant gene delivery system using mesoporous silica nanoparticles as carriers. J. Mater. Chem. B 2013, 1, 5279–5287. [Google Scholar] [CrossRef]
- Jiang, L.; Ding, L.; He, B.; Shen, J.; Xu, Z.; Yin, M.; Zhang, X. Systemic gene silencing in plants triggered by fluorescent nanoparticle-delivered double-stranded RNA. Nanoscale 2014, 6, 9965–9969. [Google Scholar] [CrossRef]
- Gao, Y.; Zhao, Y. Specific and heritable gene editing in Arabidopsis. Proc. Natl. Acad. Sci. USA 2014, 111, 4357–4358. [Google Scholar] [CrossRef]
- Kanchiswamy, C.N.; Sargent, D.J.; Velasco, R.; Maffei, M.E.; Malnoy, M. Looking forward to genetically edited fruit crops. Trends Biotechnol. 2015, 33, 62–64. [Google Scholar] [CrossRef]
- Gilbert, N.; Gewin, V.; Tollefson, J.; Sachs, J.; Potrykus, I.J.N. How to feed a hungry world. Nature 2010, 466, 531–532. [Google Scholar]
- Coleman-Derr, D.; Tringe, S.G. Building the crops of tomorrow: Advantages of symbiont-based approaches to improving abiotic stress tolerance. Front. Microbiol. 2014, 5, 283. [Google Scholar] [CrossRef]
- Christou, P. Plant genetic engineering and agricultural biotechnology 1983–2013. Trends Biotechnol. 2013, 31, 125–127. [Google Scholar] [CrossRef]
- Dong, O.X.; Ronald, P.C. Genetic Engineering for Disease Resistance in Plants: Recent Progress and Future Perspectives. Plant Physiol. 2019, 180, 26–38. [Google Scholar] [CrossRef] [Green Version]
- Giller, K.E. Genetically Engineered Crops: Experiences and Prospects; The National Academies Press: Washington, DC, USA, 2016. [Google Scholar]
- Ishii, T.; Araki, M. A future scenario of the global regulatory landscape regarding genome-edited crops. GM Crops Food 2017, 8, 44–56. [Google Scholar] [CrossRef]
- Waltz, E. With a free pass, CRISPR-edited plants reach market in record time. Nat. Biotechnol. 2018, 36, 6–7. [Google Scholar] [CrossRef]
- Wolt, J.D.; Wang, K.; Yang, B. The regulatory status of genome-edited crops. Plant Biotechnol. J. 2016, 14, 510–518. [Google Scholar] [CrossRef]
- Fritsche, S.; Poovaiah, C.; MacRae, E.; Thorlby, G. A New Zealand perspective on the application and regulation of gene editing. Front. Plant Sci. 2018, 9. [Google Scholar] [CrossRef]
- Whelan, A.I.; Lema, M.A. Regulatory framework for gene editing and other new breeding techniques (NBTs) in Argentina. GM Crops Food 2015, 6, 253–265. [Google Scholar] [CrossRef] [Green Version]
- Davison, J.; Ammann, K. New GMO regulations for old: Determining a new future for EU crop biotechnology. GM Crops Food 2017, 8, 13–34. [Google Scholar] [CrossRef] [Green Version]
- Callaway, E. CRISPR plants now subject to tough GM laws in European Union. Nature 2018, 560, 16–17. [Google Scholar] [CrossRef]
Figure 1.
History of the genetic transformation of citrus.
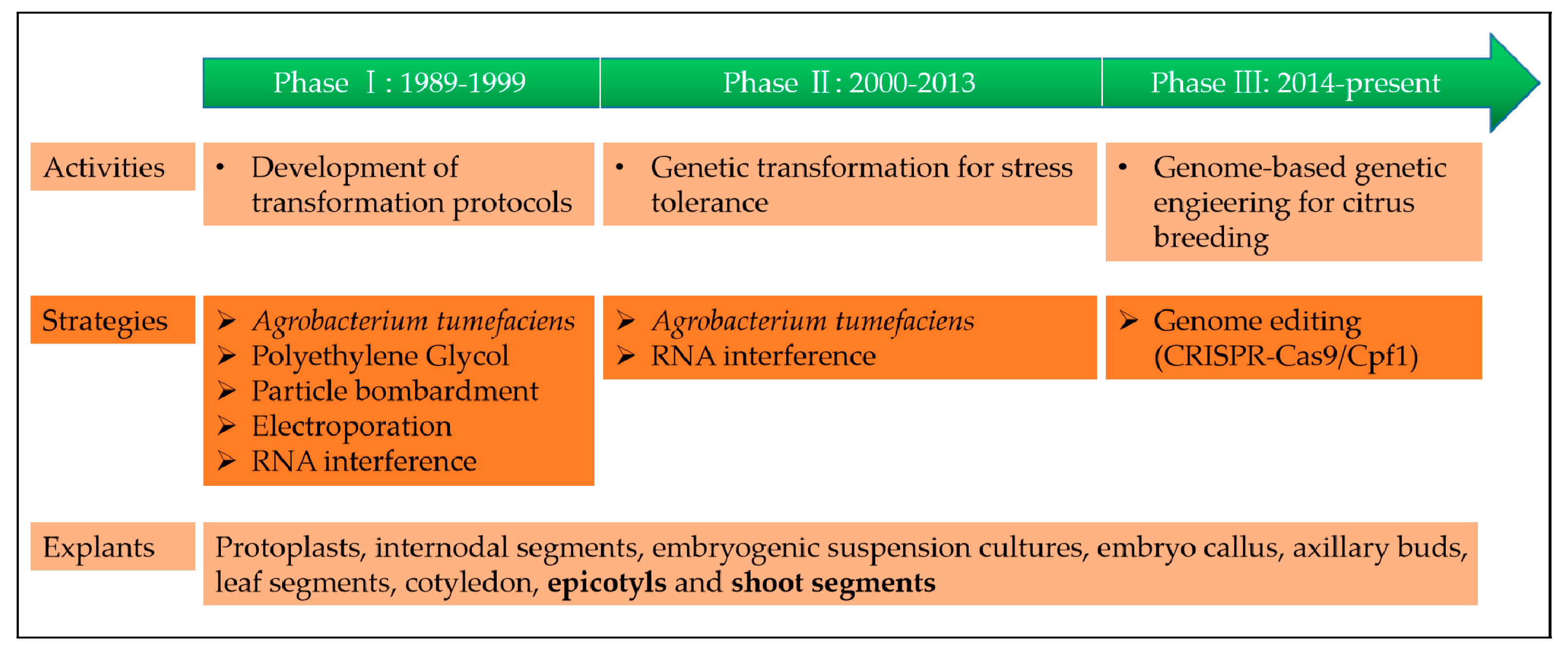

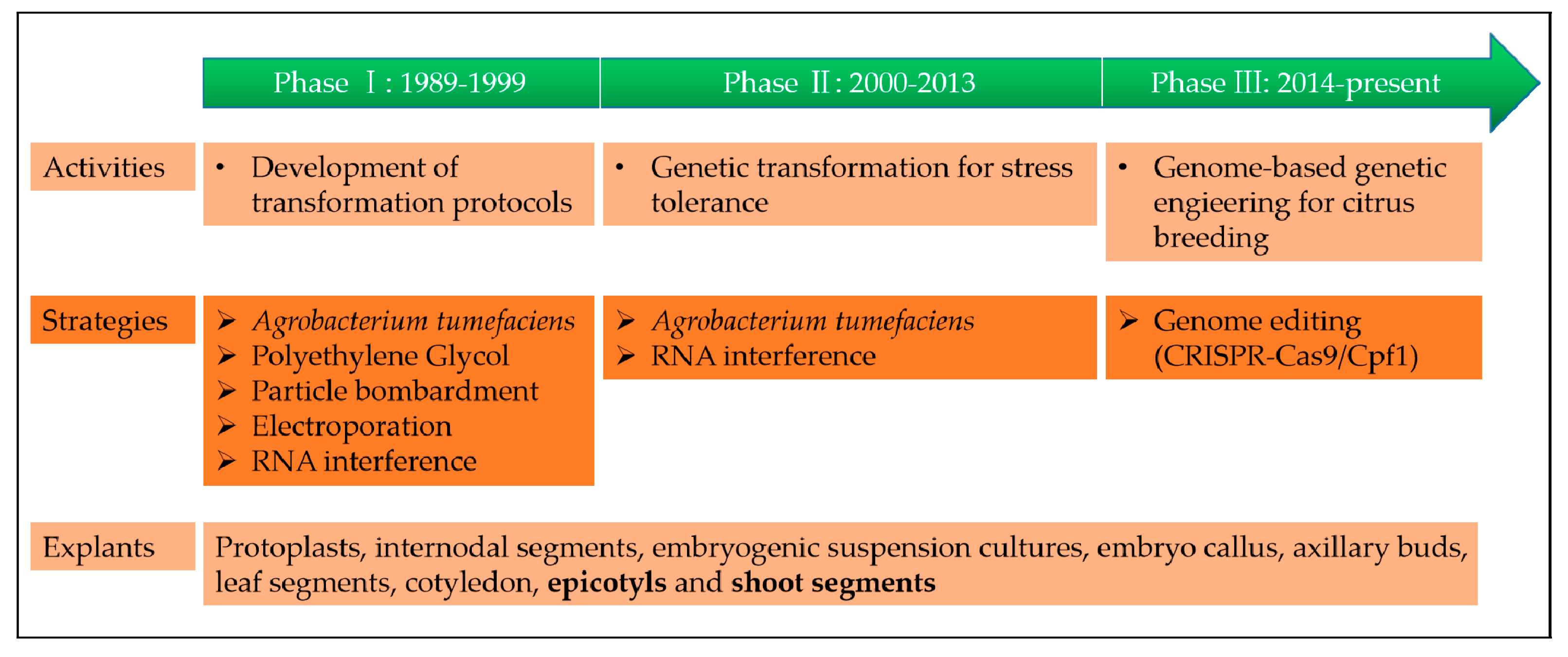{kind=link}
Table 1.
The genes used in the genetic transformation of citrus to impart resistance to canker.
| Genes | Sources | Type | Species | References |
|---|---|---|---|---|
| Attacin A (attA) | Trichoplusia ni | Antimicrobial peptide | C. sinensis cv. Hamlin/Natal/Pera/Valencia | [58,59] |
| Cecropin B and Shiva A | Synthetic | Antimicrobial peptide | C. sinensis (L.) Osbeck | [35] |
| Stx IA | Sarcophaga peregrina | Antimicrobial peptide | C. sinensis (L.) Osbeck | [60,61] |
| D2A21 | Synthetic | Antimicrobial peptide | Carrizo citrange | [62] |
| Dermaseptin gene | Synthetic | Antimicrobial peptide | C. sinensis cv. Pineapple | [63] |
| AtNPR1 | Arabidopsis thaliana | Key positive regulator of systemic acquired resistance (SAR) | C. paradisi Macf.; C. sinensis cv. Hamlin | [55] |
| CtNH1 | Citrus maxima | Key positive regulator of SAR | C. paradisi Macf. | [69] |
| Bs2 | Capsicum annuum | Resistance gene | C. sinensis cv. Hamlin/Natal/Pera/Valencia/Anliucheng; W. Murcott tangor | [72,73] |
| Xa21 | Oryza longistaminata | Resistance gene | C. limon cv. Eureka Frost Nuclear; C. sinensis cv. Pineapple | [75,76,77,78] |
| NbFLS2 | Nicotiana benthamiana | Leucine-rich repeat (LRR) receptor–like kinase gene | Carrizo citrange | [81] |
| CsMAPK1 | Citrus sinensis | Mitogen-activated protein kinase gene | Troyer citrange | [82] |
| hrpN | Erwinia amylovora | Hairpin gene | C. sinensis cv. Hamlin/Valencia | [83] |
| MdSPDS1 | Malus domestica | Spermidine synthase gene | C. sinensis cv. Anliucheng | [84] |
| terf1 | Solanum lycopersicum | Transcription factor | C. sinensis (L.) Osbeck | [85] |
| pthA-nls | Xanthomonas axonopdis pv.citri | Pathogenesis gene | C. sinensis (L.) Osbeck | [86] |
| Modified theonin | Synthetic | Cysteine-rich peptide | Carrizo citrange | [87] |
Table 2.
The genes used for imparting resistance to fungal and viral diseases in citrus.
| Genes | Sources | Type | Fungi and Virus Diseases | Species | References |
|---|---|---|---|---|---|
| CTV-CP | Citrus Tristeza virus | CTV coat protein gene | Tristeza virus | C. sinensis cv. Valencia/Hamlin | [105] |
| CTV-392/393 | Citrus Tristeza virus | CTV-derived gene | C. sinensis cv. Itaborai | [106] | |
| 3DF1scFv | Hybridoma 3DF1 cell | Monoclonal antibody | C. aurantifolia (Christm.) Swing | [107] | |
| p20/23/25 | Citrus Tristeza virus | Silencing suppressor protein | C. aurantifolia (Christm.) Swing.; C. paradisi Macf. C. sinensis (L.) Osbeck | [38,39,102,103,104,109,110,116] | |
| CPsV-CP ihpCP | Citrus Psorosis virus | Coat protein gene/siRNA | Psorosis virus | C. sinensis (L.) Osbeck; | [111,112,113] |
| chit42 | Trichoderma harzianum | Endochitinase | Mal secco and gray mold | C. limon (L.) Burm. f. | [114,115] |
| PR-5 | Solanum lycopersicum | Pathogenesis-related protein | Root rot and gummosis | C. sinensis cv. Pineapple | [116] |
| Attacin E(attE) | Antimicrobial peptide | Hyalophoracecropia | Citrus scab | C. paradisi Macf. | [117] |
| CitMTSE1 | Citrus sinensis | Limonene synthase gene | Black spot | C. sinensis (L.) Osbeck | [118] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Sun, L.; Nasrullah; Ke, F.; Nie, Z.; Wang, P.; Xu, J. Citrus Genetic Engineering for Disease Resistance: Past, Present and Future. Int. J. Mol. Sci. 2019, 20, 5256. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20215256
AMA Style
Sun L, Nasrullah, Ke F, Nie Z, Wang P, Xu J. Citrus Genetic Engineering for Disease Resistance: Past, Present and Future. International Journal of Molecular Sciences. 2019; 20(21):5256. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20215256
Chicago/Turabian StyleSun, Lifang, Nasrullah, Fuzhi Ke, Zhenpeng Nie, Ping Wang, and Jianguo Xu. 2019. "Citrus Genetic Engineering for Disease Resistance: Past, Present and Future" International Journal of Molecular Sciences 20, no. 21: 5256. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20215256
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.