Let-7f: A New Potential Circulating Biomarker Identified by miRNA Profiling of Cells Isolated from Human Abdominal Aortic Aneurysm

 and
and
Abstract
:1. Introduction
2. Results
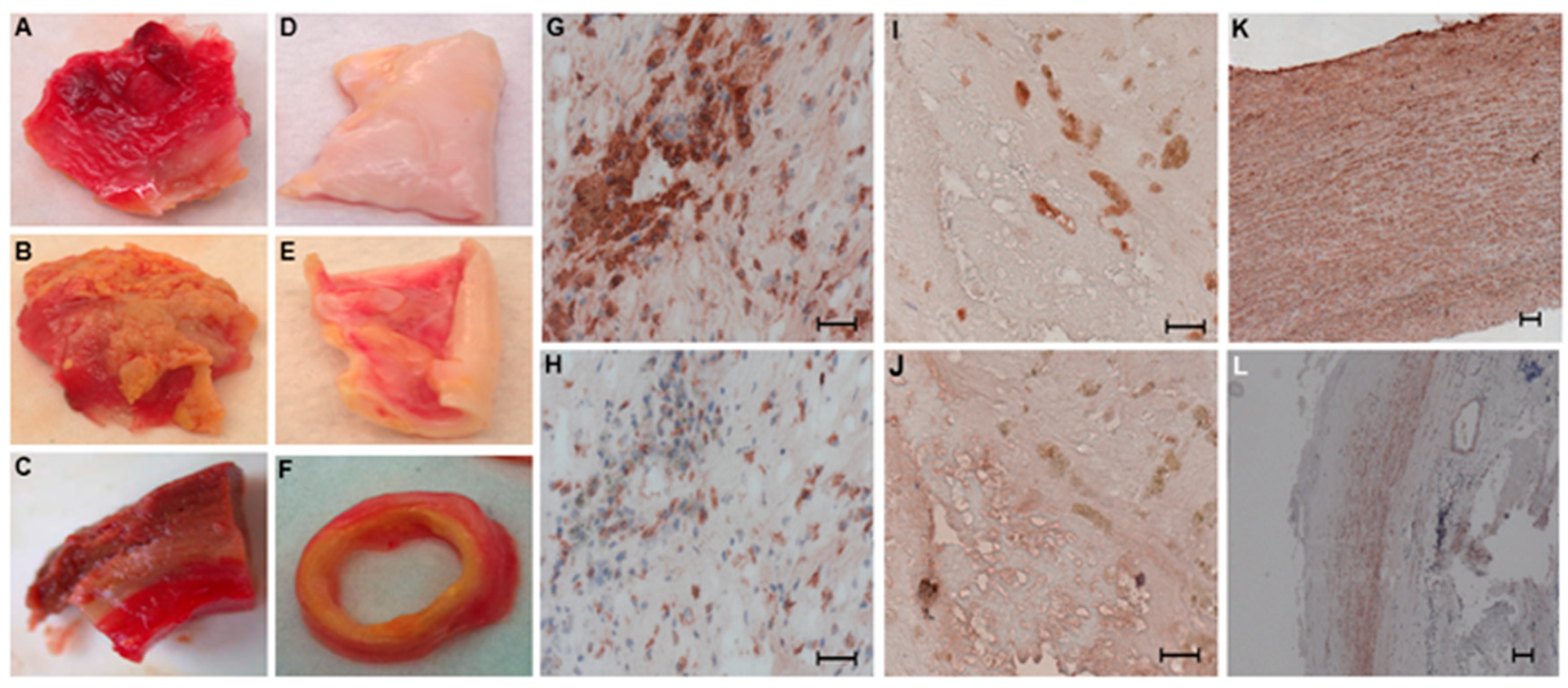
2.1. Distribution of SMCs and Macrophages in Human Aneurysmal and Control Aortic Wall Samples
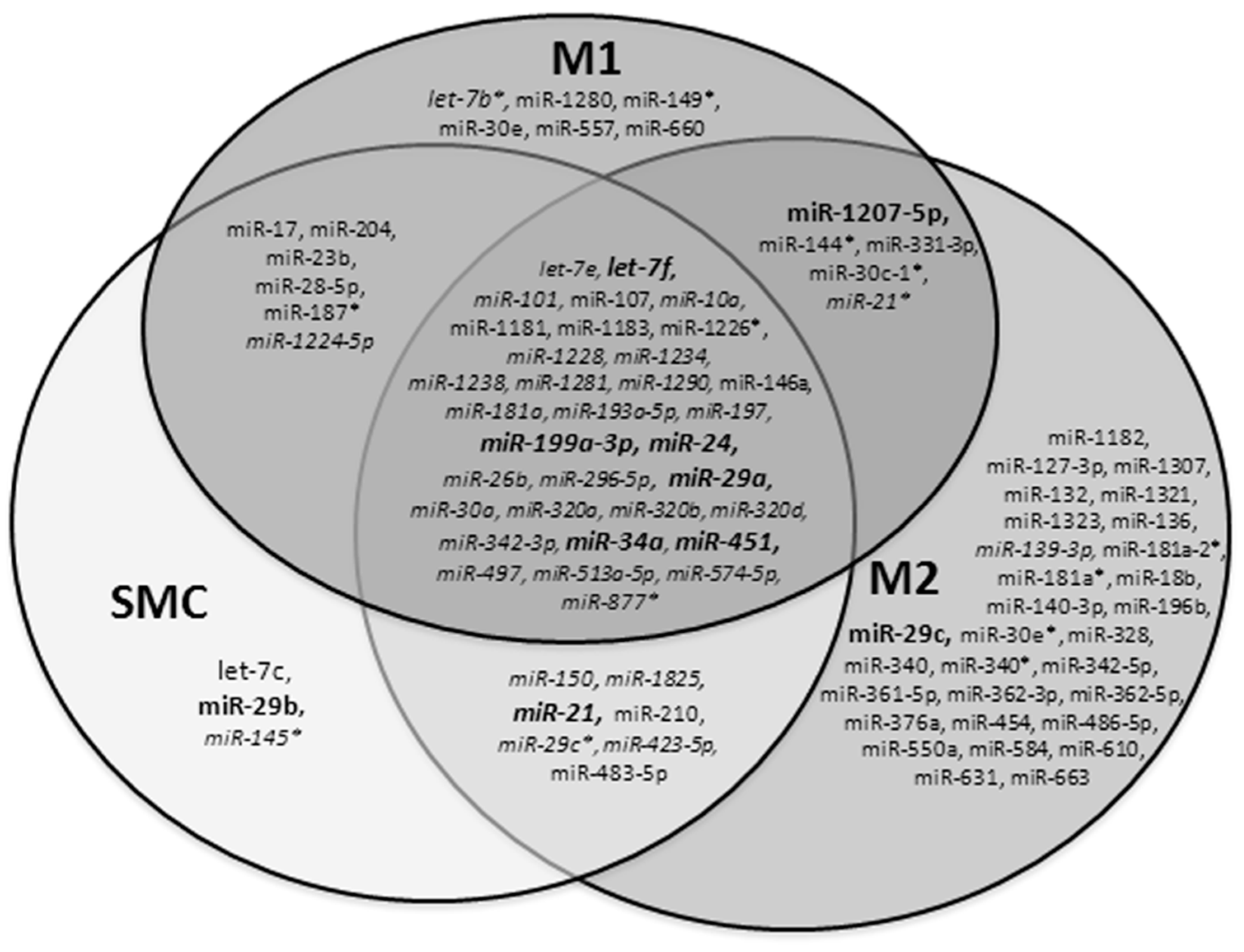
2.2. Profile of miRNAs in SMCs and Macrophages Isolated by LCM in Human Aneurysmal and Control Aortas
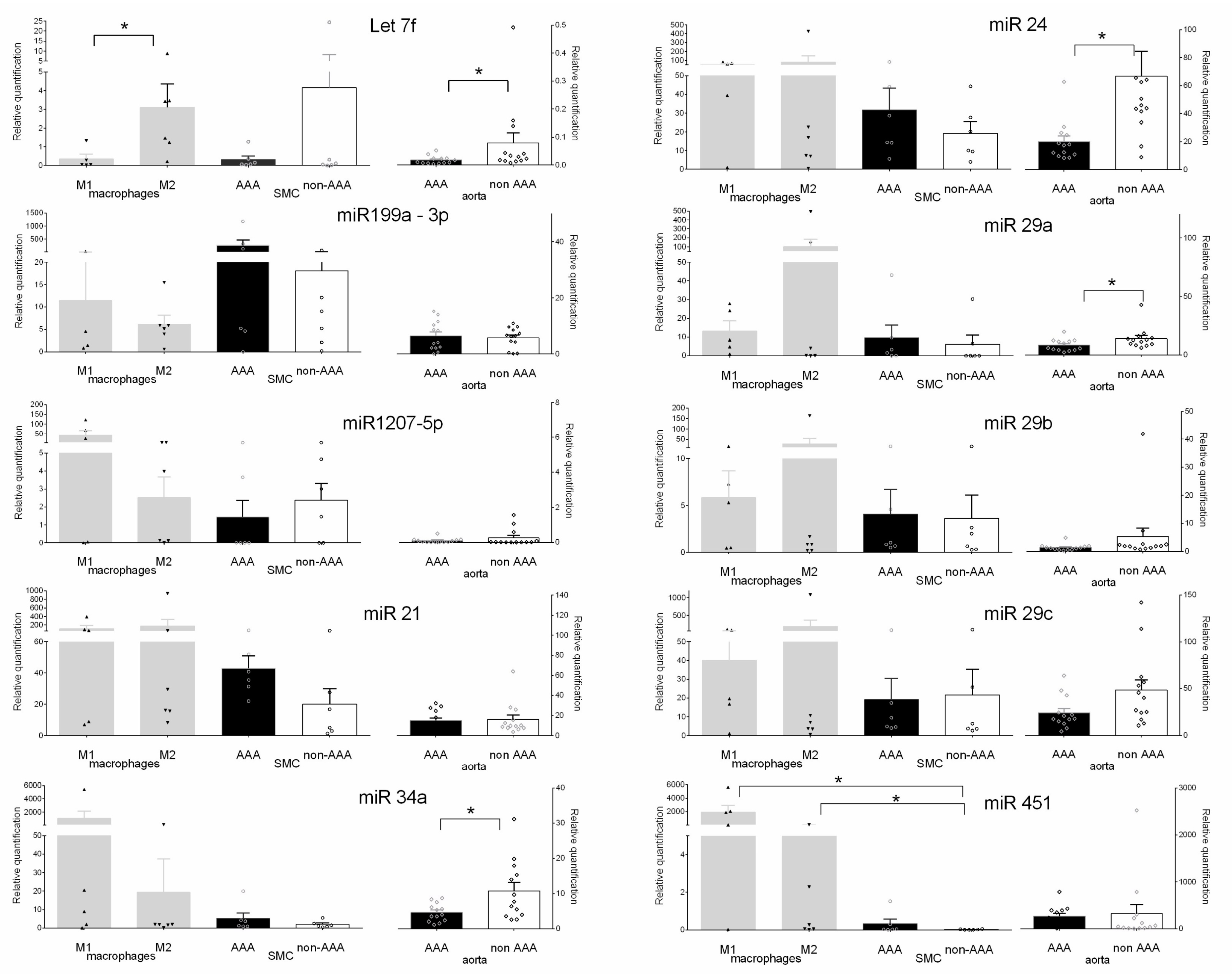
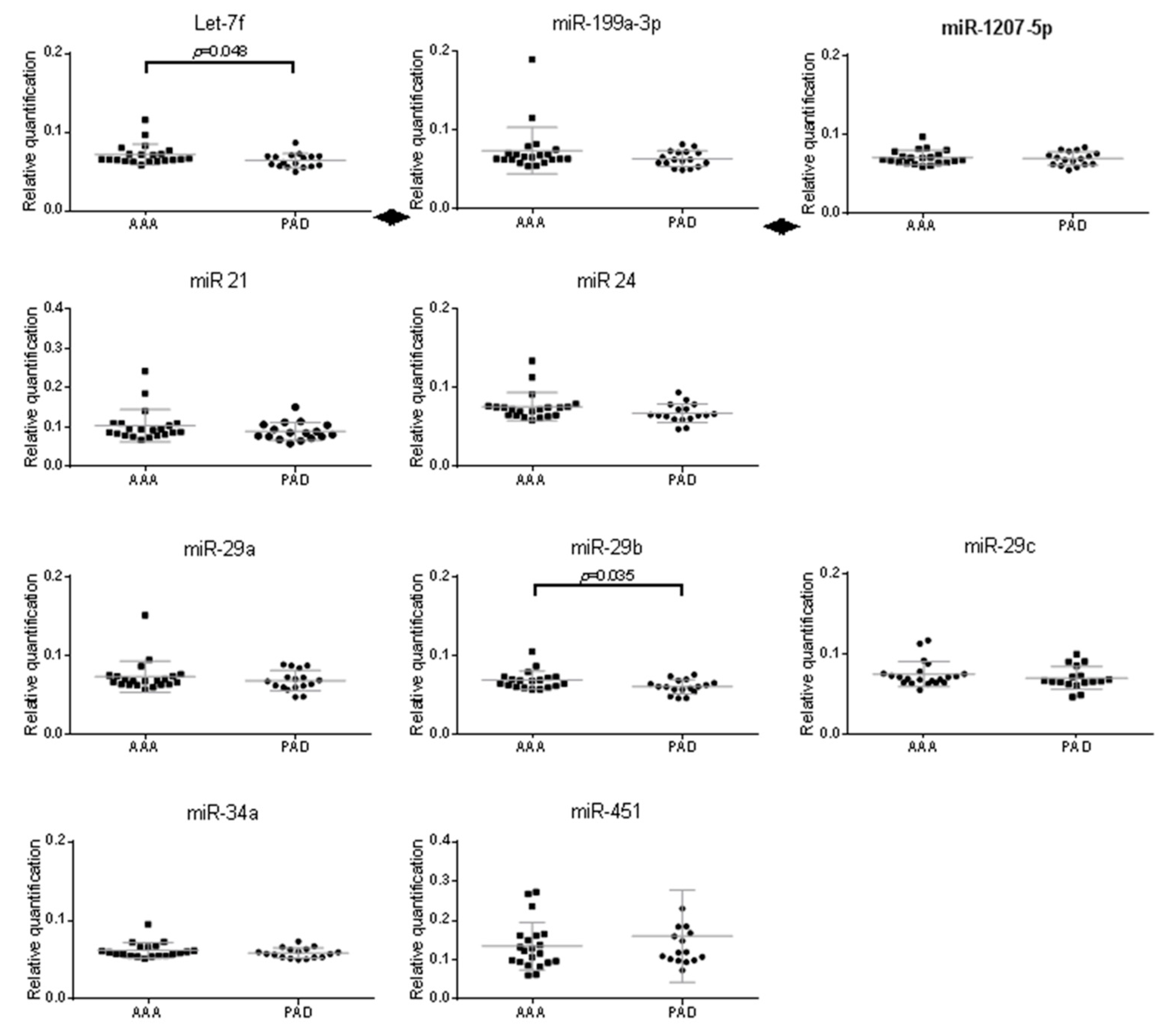
2.3. Validation of miRNAs Expression in Aneurysmal Cells
3. Discussion
4. Materials and Methods
4.1. Human Abdominal Aorta Samples
4.2. Human Plasma Samples
4.3. Histological Analysis and Immunohistochemistry
4.4. Laser Capture Microdissection (LCM)
4.5. Microarray Analysis
4.6. miRNA Quantification by q-RT-PCR
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AAA | Abdominal aortic aneurysm |
| LCM | Laser capture microdissection |
| miRNA | microRNA |
| MMP | Metalloprotease |
| MR | Mannose receptor |
| SMC | Smooth muscle cell |
| PAD | Peripheral arterial disease |
References
- Brown, L.; Powell, J. Risk factors for aneurysm rupture in patients kept under ultrasound surveillance. UK Small Aneurysm Trial Participants. Ann. Surg. 1999, 230, 289–296. [Google Scholar] [CrossRef] [PubMed]
- Sakalihasan, N.; Limet, R.D. Abdominal aortic aneurysm. Lancet 2005, 365, 1577–1589. [Google Scholar] [CrossRef]
- Chaikof, E.; Dalman, R.; Eskandari, M.; Jackson, B.; Lee, W.; Mansour, M.; Mastracci, T.; Mell, M.; Murad, M.; Nguyen, L.; et al. The Society for Vascular Surgery practice guidelines on the care of patients with an abdominal aortic aneurysm. J. Vasc. Surg. 2018, 67, 2–77. [Google Scholar] [CrossRef] [PubMed]
- Ascoli Marchetti, A.; Pratesi, G.; Di Giulio, L.; Battistini, M.; Massoud, R.; Ippoliti, A. EVAR and OPEN treatment of abdominal aortic aneurysm: What is the role of MMP-9 in the follow-up? J. Med. Vasc. 2017, 42, 21–28. [Google Scholar] [CrossRef] [Green Version]
- Sangiorgi, G.; Biondi-Zoccai, G.; Pizzuto, A.; Martelli, E. Biochemical Markers for Diagnosis and Follow-up of Aortic Diseases: An Endless Search for the Holy Grail. J. Endovasc. Ther. 2019. [Google Scholar] [CrossRef]
- Quintana, R.; Taylor, W. Cellular Mechanisms of Aortic Aneurysm Formation. Circ. Res. 2019, 124, 607–618. [Google Scholar] [CrossRef]
- Michel, J. Anoikis in the cardiovascular system: Known and unknown extracellular mediators. Arter. Thromb. Vasc. Biol. 2003, 23, 2146–2154. [Google Scholar] [CrossRef]
- Thompson, M. Controlling the expansion of abdominal aortic aneurysms. Br. J. Surg. 2003, 90, 897–898. [Google Scholar] [CrossRef]
- Reeps, C.; Pelisek, J.; Seidl, S.; Schuster, T.; Zimmermann, A.; Kuehnl, A.; Eckstein, H. Inflammatory Infiltrates and Neovessels Are Relevant Sources of MMPs in Abdominal Aortic Aneurysm Wall. Pathobiology 2009, 76, 243–252. [Google Scholar] [CrossRef]
- Lohoefer, F.; Reeps, C.; Lipp, C.; Rudelius, M.; Zimmermann, A.; Ockert, S.; Eckstein, H.; Pelisek, J. Histopathological analysis of cellular localization of cathepsins in abdominal aortic aneurysm wall. Int. J. Exp. Pathol. 2012, 93, 252–258. [Google Scholar] [CrossRef]
- Koch, A.; Haines, G.; Rizzo, R.; Radosevich, J.; Pope, R.; Robinson, P.; Pearce, W. Human abdominal aortic aneurysms. Immunophenotypic analysis suggesting an immune-mediated response. Am. J. Pathol. 1990, 37, 1199–1213. [Google Scholar]
- Rizzo, R.J.; McCarthy, W.J.; Dixit, S.N.; Lilly, M.P.; Shively, V.P.; Flinn, W.R.; Yao, J.S. Collagen types and matrix protein content in human abdominal aortic aneurysms. J. Vasc. Surg. 1989, 10, 365–373. [Google Scholar] [CrossRef] [Green Version]
- Gordon, S. Alternative activation of macrophages. Nat. Rev. Immunol. 2003, 3, 23–35. [Google Scholar] [CrossRef] [PubMed]
- Stein, M.; Keshav, S.; Harris, N.; Gordon, S. Interleukin 4 Potently Enhances Murine Macrophage Mannose Receptor Activity: A Marker of Alternative Immunologic Macrophage Activation. J. Exp. Med. 1992, 176, 287–292. [Google Scholar] [CrossRef]
- Boytard, L.; Spear, R.; Chinetti-Gbaguidi, G.; Acosta-Martin, A.E.; Vanhoutte, J.; Lamblin, N.; Staels, B.; Amouyel, P.; Haulon, S.; Pinet, F. Role of proinflammatory CD68+ mannose receptor-macrophages in peroxiredoxin-1 expression and in abdominal aortic aneurysms in humans. Arter. Thromb. Vasc. Biol. 2013, 33, 431–438. [Google Scholar] [CrossRef]
- Sho, E.; Sho, M.; Nanjo, H.; Kawamura, K.; Masuda, H.; Dalman, R. Comparison of cell-type-specific vs. transmural aortic gene expression in experimental aneurysms. J. Vasc. Surg. 2005, 41, 844–852. [Google Scholar] [CrossRef]
- Spear, R.; Boytard, L.; Blervaque, R.; Chwastyniak, M.; Hot, D.; Vanhoutte, J.; Staels, B.; Lemoine, Y.; Lamblin, N.; Pruvot, F.-R.; et al. Adventitial Tertiary Lymphoid Organs as Potential Source of MicroRNA Biomarkers for Abdominal Aortic Aneurysm. Int. J. Mol. Sci. 2015, 16, 11276–11293. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, P.S.; Parkin, R.K.; Kroh, E.M.; Fritz, B.R.; Wyman, S.K.; Pogosova-Agadjanyan, E.L.; Peterson, A.; Noteboom, J.; O’Briant, K.C.; Allen, A.; et al. Circulating microRNAs as stable blood-based markers for cancer detection. Proc. Natl. Acad. Sci. USA 2008, 105, 10513–10518. [Google Scholar] [CrossRef] [Green Version]
- Maegdefessel, L.; Azuma, J.; Toh, R.; Merk, D.R.; Deng, A.; Chin, J.T.; Raaz, U.; Schoelmerich, A.M.; Raiesdana, A.; Leeper, N.J.; et al. Inhibition of microRNA-29b reduces murine abdominal aortic aneurysm development. J. Clin. Invest. 2012, 122, 497–506. [Google Scholar] [CrossRef] [Green Version]
- Maegdefessel, L.; Maegdefessel, L.; Azuma, J.; Toh, R.; Deng, A.; Merk, D.R.; Raiesdana, A.; Leeper, N.J.; Raaz, U.; Schoelmerich, A.M.; et al. MicroRNA-21 Blocks Abdominal Aortic Aneurysm Development and Nicotine-Augmented Expansion. Sci. Transl. Med. 2012, 22, 122ra22. [Google Scholar] [CrossRef]
- Pahl, M.C.; Derr, K.; Gäbel, G.; Hinterseher, I.; Elmore, J.R.; Schworer, C.M.; Peeler, T.C.; Franklin, D.P.; Gray, J.L.; Carey, D.J.; et al. MicroRNA expression signature in human abdominal aortic aneurysms. BMC Med. Genom. 2012, 5, 25. [Google Scholar] [CrossRef] [PubMed]
- Kin, K.; Miyagawa, S.; Fukushima, S.; Shirakawa, Y.; Torikai, K.; Shimamura, K.; Daimon, T.; Kawahara, Y.; Kuratani, T.; Sawa, Y. Tissue- and Plasma-Specifi MicroRNA Signatures for Atherosclerotic Abdominal Aortic Aneurysm. J. Am. Hear. Assoc. 2012, 1, e000745. [Google Scholar] [CrossRef]
- Iyer, V.; Rowbotham, S.; Biros, E.; Bingley, J.; Golledge, J. A systematic review investigating the association of microRNAs with human abdominal aortic aneurysms. Atherosclerosis 2017, 261, 78–89. [Google Scholar] [CrossRef] [PubMed]
- Lamblin, N.; Ratajczak, P.; Hot, D.; Dubois, E.; Chwastyniak, M.; Beseme, O.; Drobecq, H.; Lemoine, Y.; Koussa, M.; Amouyel, P.; et al. Profile of macrophages in human abdominal aortic aneurysms: A transcriptomic, proteomic, and antibody protein array study. J. Proteome Res. 2010, 9, 3720–3729. [Google Scholar] [CrossRef] [PubMed]
- Busch, A.; Eken, S.; Maegdefessel, L. Prospective and therapeutic screening value of non-coding RNA as biomarkers in cardiovascular disease. Ann. Transl. Med. 2016, 4, 236. [Google Scholar] [CrossRef]
- Bartel DP MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297.
- Kent, O.; McCall, M.; Cornish, T.; Halushka, M. Lessons from miR-143/145: The importance of cell-type localization of miRNAs. Nucleic Acids Res. 2004, 42, 7528–7538. [Google Scholar] [CrossRef]
- Henderson, E.; Geng, Y.; Sukhova, G.; Whittemore, A.; Knox, J.; Libby, P. Death of smooth muscle cells and expression of mediators of apoptosis by T lymphocytes in human abdominal aortic aneurysms. Circulation 1999, 99, 96–104. [Google Scholar] [CrossRef]
- Chinetti-Gbaguidi, G.; Baron, M.; Bouhlel, M.A.; Vanhoutte, J.; Copin, C.; Sebti, Y.; Derudas, B.; Bories, G.; Tailleux, A.; Haulon, S.; et al. Human Atherosclerotic Plaque Alternative Macrophages Display Low Cholesterol Handling but High Phagocytosis Because of Distinct Activities of the PPARg and LXRa Pathways LOX. Circ. Res. 2011, 108, 985–995. [Google Scholar] [CrossRef]
- Cheuk, B.L.Y.; Cheng, S.W.K. Identification and characterization of microRNAs in vascular smooth muscle cells from patients with abdominal aortic aneurysms. J. Vasc. Surg. 2014, 59, 202–209. [Google Scholar] [CrossRef] [Green Version]
- Biros, E.; Moran, C.S.; Wang, Y.; Walker, P.J.; Cardinal, J.; Golledge, J. microRNA profiling in patients with abdominal aortic aneurysms: The significance of miR-155. Clin. Sci. 2014, 803, 795–803. [Google Scholar] [CrossRef] [PubMed]
- Pritchard, C.; Kroh, E.; Wood, B.; Arroyo, J.; Dougherty, K.; Miyaji, M.; Tait, J.; Tewari, M. Blood cell origin of circulating microRNAs: A cautionary note for cancer biomarker studies. Nat. Rev. Genet. 2012, 13, 358–369. [Google Scholar] [CrossRef] [PubMed]
- Stather, P.; Sylvius, N.; Sidloff, D.A.; Dattani, N.; Verissimo, A.; Wild, J.B.; Butt, H.Z. Identification of microRNAs associated with abdominal aortic. Br. J. Surg. 2015, 101, 755–766. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Cao, H.; Zhuang, J.; Wan, J.; Guan, M.; Yu, B.; Li, X.; Zhang, W. Identification of miR-130a, miR-27b and miR-210 as serum biomarkers for atherosclerosis obliterans. Clin. Chim. Acta 2011, 412, 66–70. [Google Scholar] [CrossRef]
- Jiang, S.; Yan, W.; Wang, S.; Baltimore, D. Dual mechanisms of posttranscriptional regulation of Tet2 by Let-7 microRNA in macrophages. Proc. Natl. Acad. Sci. USA 2019, 116, 12416–12421. [Google Scholar] [CrossRef] [Green Version]
- Matthews, E.; Rowbotham, S.; Moxon, J.; Jones, R.; Vega de Ceniga, M.; Golledge, J. Meta-analysis of the association between peripheral artery disease and growth of abdominal aortic aneurysms. Br. J. Surg. 2017, 104, 1765–1774. [Google Scholar] [CrossRef]
- Tenorio, E.; Braga, A.; Tirapelli, D.; Ribeiro, M.; Piccinato, C.; Joviliano, E. Expression in Whole Blood Samples of miRNA-191 and miRNA-455-3p in Patients with AAA and Their Relationship to Clinical Outcomes after Endovascular Repair. Ann. Vasc. Surg. 2018, 50, 209–217. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| AAA (n = 24) | PAD (n = 18) | |
|---|---|---|
| Age, years | 68.0 ± 6.1 | 62.3 ± 6.6 |
| Body mass index, kg/m2 | 27.4 ± 3.7 | 26.3 ± 4.1 |
| Aortic diameter | 56.1 ± 2.3 | |
| Cardiovascular risk factors, n (%) | ||
| Current smoking | 4 (17) | 4 (22) |
| Past smoking | 17 (71) | 13 (72) |
| Hypercholesterolemia | 14 (58) | 12 (67) |
| Hypertension | 15 (63) | 10 (56) |
| Diabetes mellitus | 4 (17) | 5 (28) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Spear, R.; Boytard, L.; Blervaque, R.; Chwastyniak, M.; Hot, D.; Vanhoutte, J.; Lamblin, N.; Amouyel, P.; Pinet, F. Let-7f: A New Potential Circulating Biomarker Identified by miRNA Profiling of Cells Isolated from Human Abdominal Aortic Aneurysm. Int. J. Mol. Sci. 2019, 20, 5499. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20215499
Spear R, Boytard L, Blervaque R, Chwastyniak M, Hot D, Vanhoutte J, Lamblin N, Amouyel P, Pinet F. Let-7f: A New Potential Circulating Biomarker Identified by miRNA Profiling of Cells Isolated from Human Abdominal Aortic Aneurysm. International Journal of Molecular Sciences. 2019; 20(21):5499. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20215499
Chicago/Turabian StyleSpear, Rafaelle, Ludovic Boytard, Renaud Blervaque, Maggy Chwastyniak, David Hot, Jonathan Vanhoutte, Nicolas Lamblin, Philippe Amouyel, and Florence Pinet. 2019. "Let-7f: A New Potential Circulating Biomarker Identified by miRNA Profiling of Cells Isolated from Human Abdominal Aortic Aneurysm" International Journal of Molecular Sciences 20, no. 21: 5499. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20215499