ZmRAD51C Is Essential for Double-Strand Break Repair and Homologous Recombination in Maize Meiosis


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
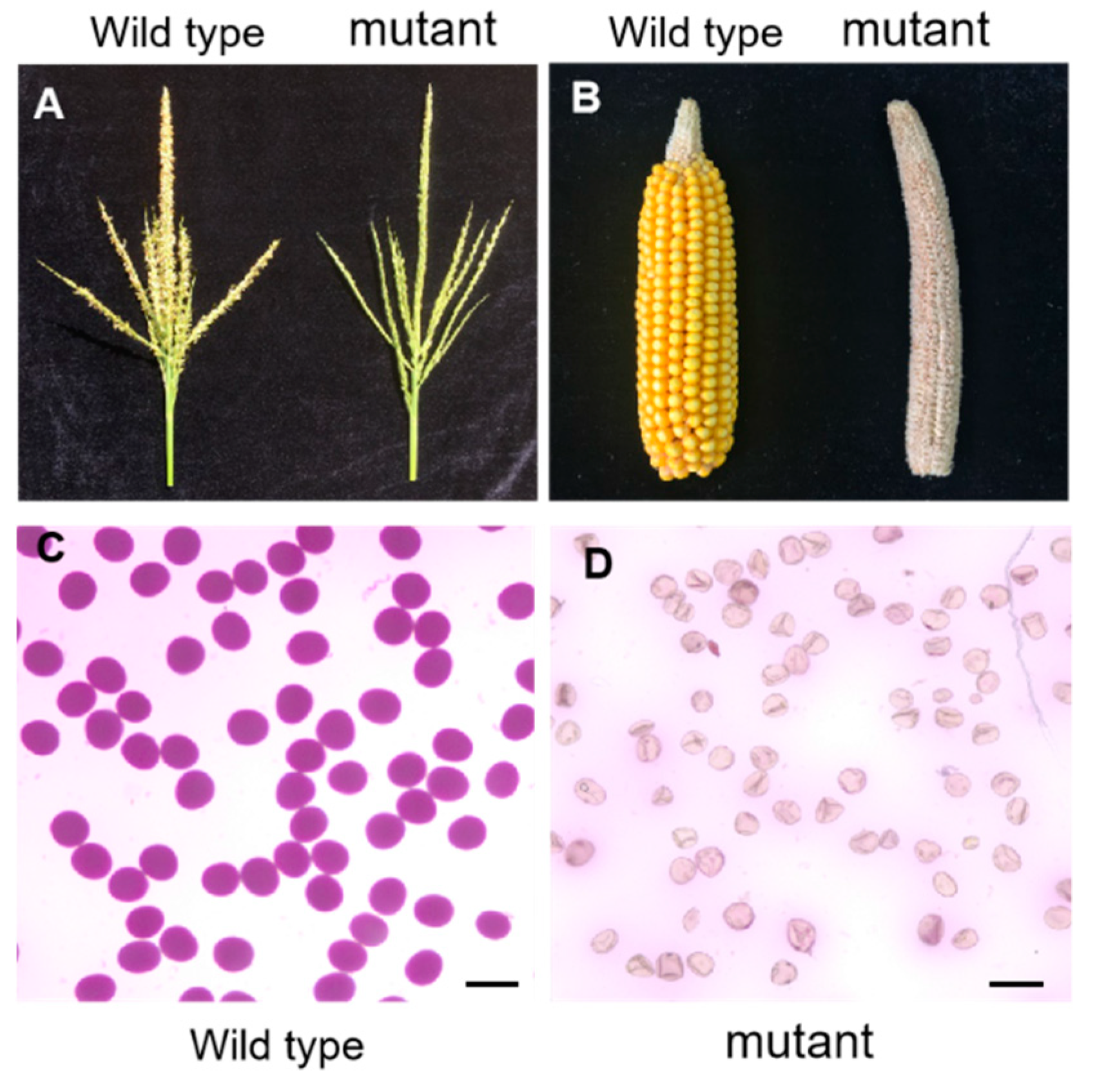
2.1. Characterization of a Sterile Mutant
2.2. Map-Based Cloning and Characterization of ZmRAD51C
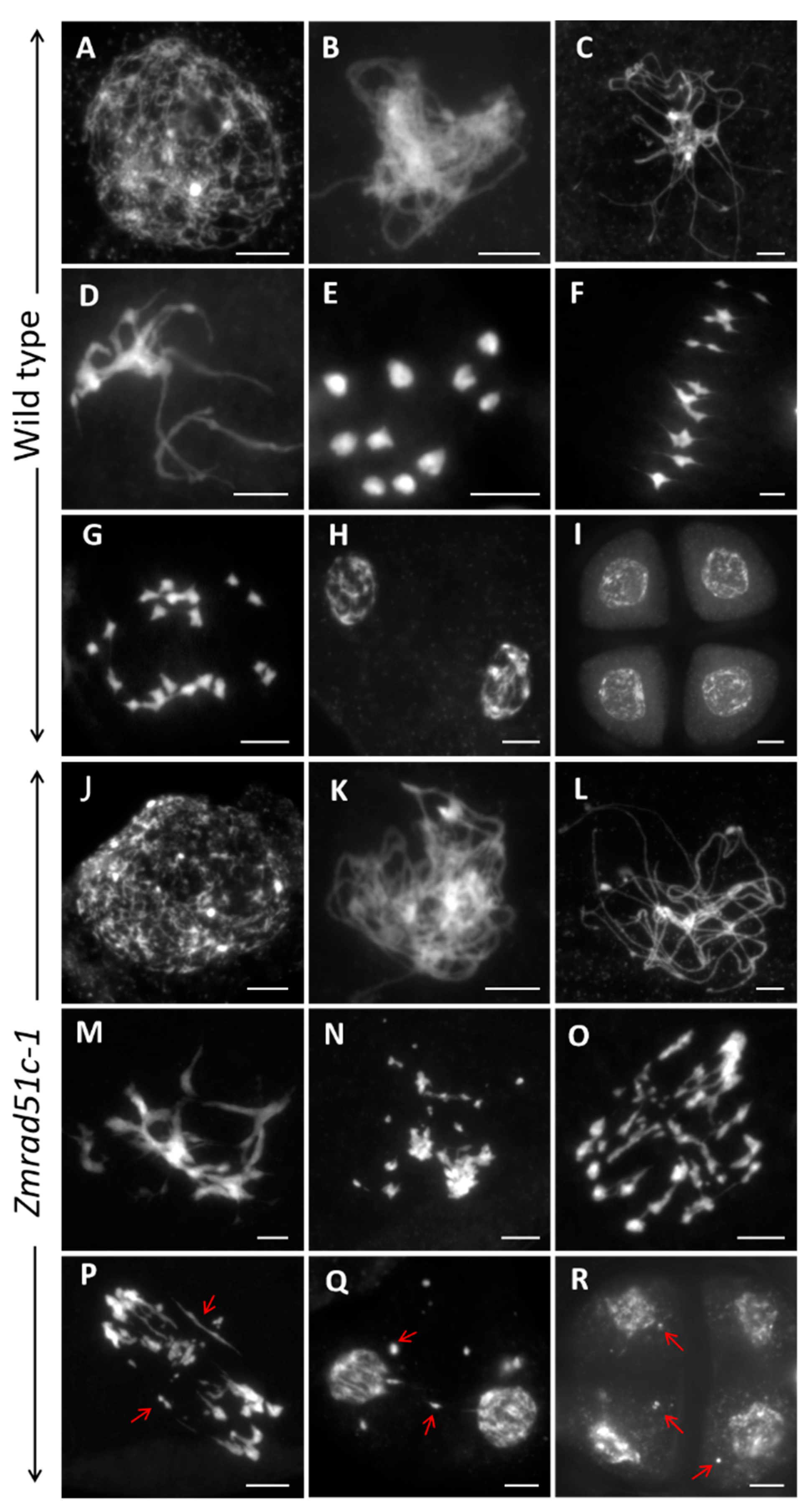
2.3. Abnormal Meiotic Chromosome Behaviors in Zmrad51c
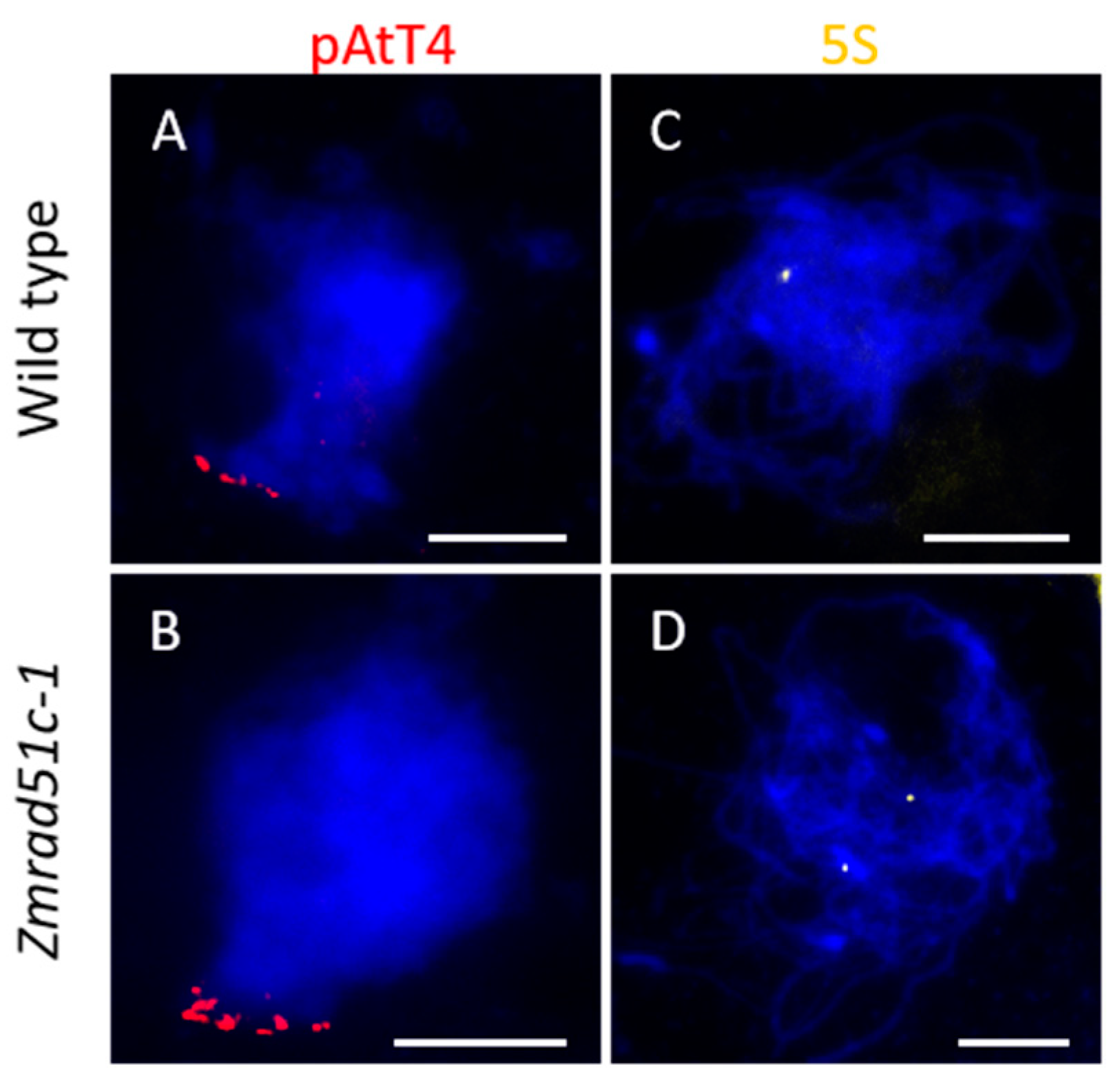
2.4. Normal Telomere Bouquet Clustering but Deficient Homologous Chromosome Pairing in Zmrad51c
2.5. Normal Axial Element Installation but Defective Central Element Installation in Zmrad51c
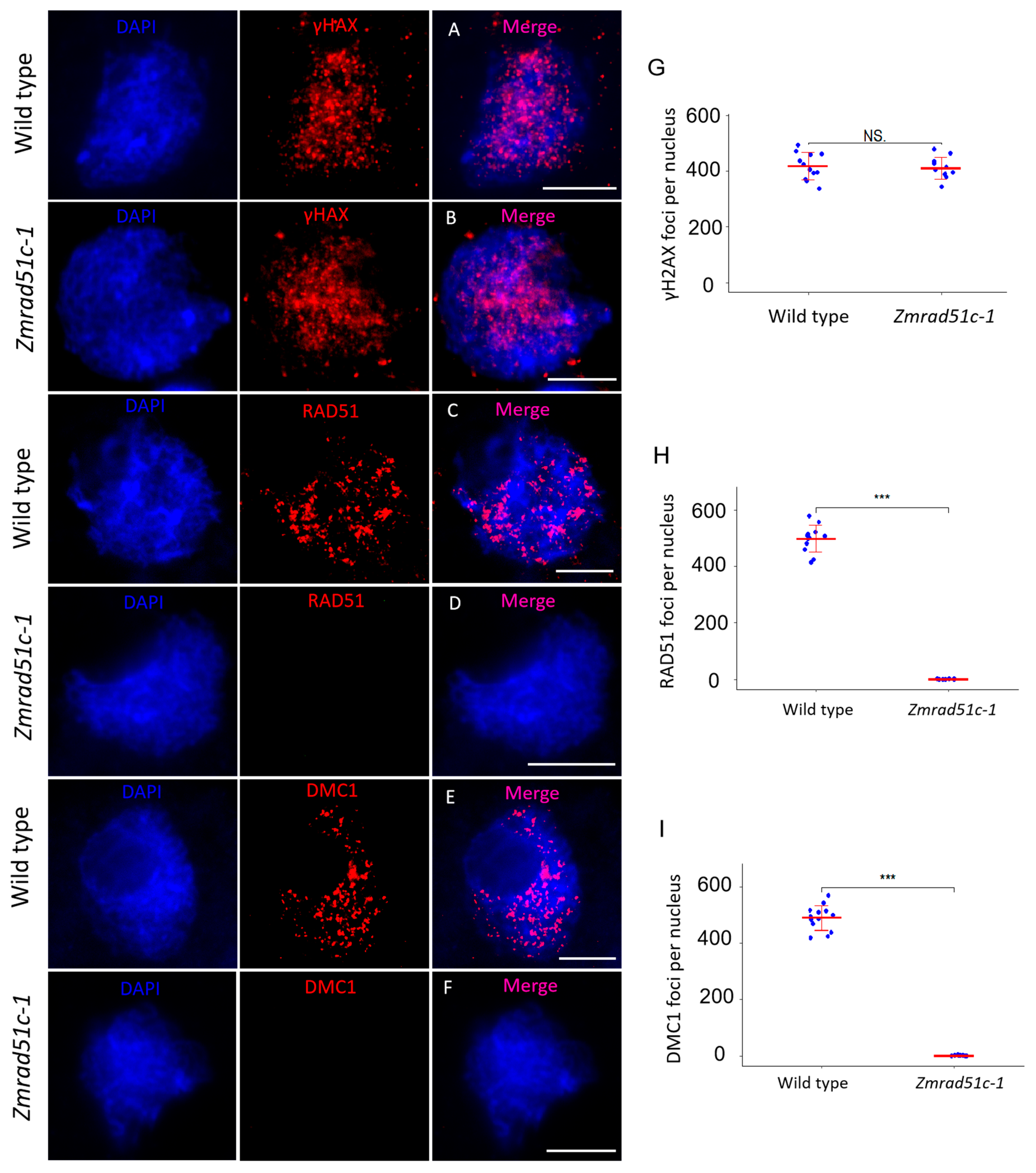
2.6. Normal DSB Formation but Defective Loading of RAD51 and DMC1 in Zmrad51c
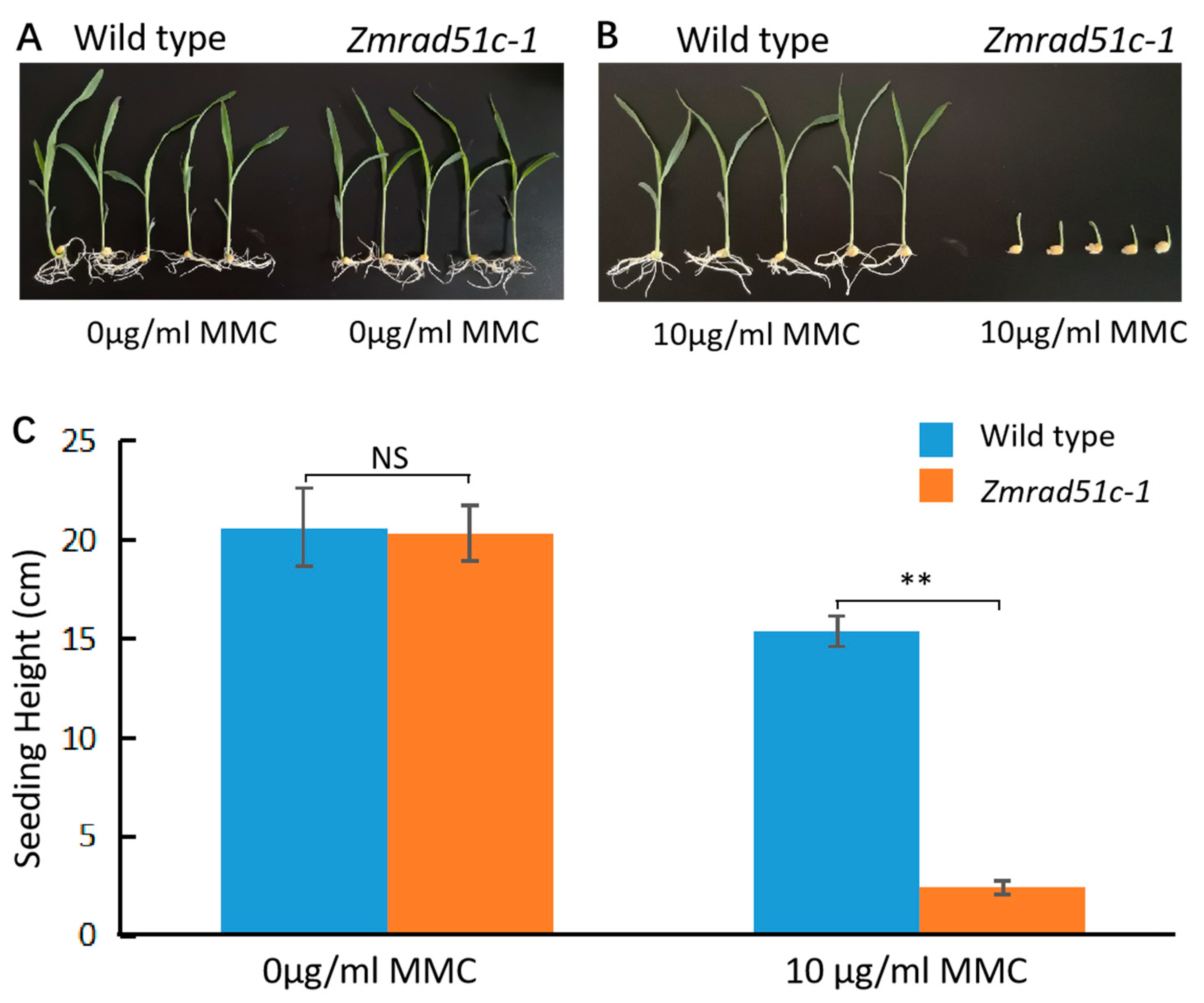
2.7. Zmrad51c Mutants Are Hypersensitive to DNA Damage Agent
3. Discussions
3.1. Functional Conservation of RAD51C in Different Organisms
3.2. Abnormal Chromosome Fragmentation in Zmrad51c
4. Materials and Methods
4.1. Plant Materials
4.2. Map-Based Cloning of Zmrad51c-1
4.3. cDNA Cloning and Real-Time qPCR for Transcript Expression Assay
4.4. Pollen Viability Observation
4.5. Meiotic Chromosome Preparation
4.6. FISH Analysis
4.7. Fluorescence Immunolocalization Assays
4.8. DNA Damage Agent Mitomycin C (MMC) Treatment
4.9. Image Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| DSB | Double strand break |
| HR | Homologous recombination |
| SC | Synaptonemal complex |
| AE | Axial elements |
| CE | Central elements |
| TF | Transverse filament |
References
- De Massy, B. Initiation of meiotic recombination: How and where? Conservation and specificities among eukaryotes. Annu. Rev. Genet. 2013, 47, 563–599. [Google Scholar] [CrossRef] [PubMed]
- Edlinger, B.; Schlogelhofer, P. Have a break: Determinants of meiotic DNA double strand break (DSB) formation and processing in plants. J. Exp. Bot. 2011, 62, 1545–1563. [Google Scholar] [CrossRef] [PubMed]
- Keeney, S. Spo11 and the Formation of DNA Double-Strand Breaks in Meiosis. Genome Dyn. Stab. 2008, 2, 81–123. [Google Scholar] [CrossRef] [PubMed]
- Keeney, S.; Neale, M.J. Initiation of meiotic recombination by formation of DNA double-strand breaks: Mechanism and regulation. Biochem Soc. Trans. 2006, 34, 523–525. [Google Scholar] [CrossRef]
- Cole, F.; Keeney, S.; Jasin, M. Evolutionary conservation of meiotic DSB proteins: More than just Spo11. Genes Dev. 2010, 24, 1201–1207. [Google Scholar] [CrossRef]
- Alani, E.; Padmore, R.; Kleckner, N. Analysis of wild-type and rad50 mutants of yeast suggests an intimate relationship between meiotic chromosome synapsis and recombination. Cell 1990, 61, 419–436. [Google Scholar] [CrossRef]
- Cao, L.; Alani, E.; Kleckner, N. A pathway for generation and processing of double-strand breaks during meiotic recombination in S. cerevisiae. Cell 1990, 61, 1089–1101. [Google Scholar] [CrossRef]
- Ivanov, E.L.; Korolev, V.G.; Fabre, F. XRS2, a DNA repair gene of Saccharomyces cerevisiae, is needed for meiotic recombination. Genetics 1992, 132, 651–664. [Google Scholar]
- McKee, A.H.; Kleckner, N. A general method for identifying recessive diploid-specific mutations in Saccharomyces cerevisiae, its application to the isolation of mutants blocked at intermediate stages of meiotic prophase and characterization of a new gene SAE2. Genetics 1997, 146, 797–816. [Google Scholar]
- Nairz, K.; Klein, F. mre11S—A yeast mutation that blocks double-strand-break processing and permits nonhomologous synapsis in meiosis. Genes Dev. 1997, 11, 2272–2290. [Google Scholar] [CrossRef]
- Prinz, S.; Amon, A.; Klein, F. Isolation of COM1, a new gene required to complete meiotic double-strand break-induced recombination in Saccharomyces cerevisiae. Genetics 1997, 146, 781–795. [Google Scholar] [PubMed]
- Longhese, M.P.; Bonetti, D.; Guerini, I.; Manfrini, N.; Clerici, M. DNA double-strand breaks in meiosis: Checking their formation, processing and repair. DNA Repair 2009, 8, 1127–1138. [Google Scholar] [CrossRef] [PubMed]
- Mimitou, E.P.; Symington, L.S. DNA end resection: Many nucleases make light work. DNA Repair 2009, 8, 983–995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bishop, D.K.; Park, D.; Xu, L.; Kleckner, N. DMC1: A meiosis-specific yeast homolog of E. coli recA required for recombination, synaptonemal complex formation, and cell cycle progression. Cell 1992, 69, 439–456. [Google Scholar] [CrossRef]
- Shinohara, A.; Ogawa, H.; Ogawa, T. Rad51 protein involved in repair and recombination in S. cerevisiae is a RecA-like protein. Cell 1992, 69, 457–470. [Google Scholar] [CrossRef]
- Cloud, V.; Chan, Y.L.; Grubb, J.; Budke, B.; Bishop, D.K. Rad51 is an accessory factor for Dmc1-mediated joint molecule formation during meiosis. Science 2012, 337, 1222–1225. [Google Scholar] [CrossRef]
- Allers, T.; Lichten, M. Differential timing and control of noncrossover and crossover recombination during meiosis. Cell 2001, 106, 47–57. [Google Scholar] [CrossRef]
- Borner, G.V.; Kleckner, N.; Hunter, N. Crossover/noncrossover differentiation, synaptonemal complex formation, and regulatory surveillance at the leptotene/zygotene transition of meiosis. Cell 2004, 117, 29–45. [Google Scholar] [CrossRef]
- Seitz, E.M.; Brockman, J.P.; Sandler, S.J.; Clark, A.J.; Kowalczykowski, S.C. RadA protein is an archaeal RecA protein homolog that catalyzes DNA strand exchange. Genes Dev. 1998, 12, 1248–1253. [Google Scholar] [CrossRef]
- Baumann, P.; West, S.C. Role of the human RAD51 protein in homologous recombination and double-stranded-break repair. Trends Biochem. Sci. 1998, 23, 247–251. [Google Scholar] [CrossRef]
- Shinohara, A.; Ogawa, H.; Matsuda, Y.; Ushio, N.; Ikeo, K.; Ogawa, T. Cloning of human, mouse and fission yeast recombination genes homologous to RAD51 and recA. Nat. Genet. 1993, 4, 239–243. [Google Scholar] [CrossRef] [PubMed]
- Tebbs, R.S.; Zhao, Y.; Tucker, J.D.; Scheerer, J.B.; Siciliano, M.J.; Hwang, M.; Liu, N.; Legerski, R.J.; Thompson, L.H. Correction of chromosomal instability and sensitivity to diverse mutagens by a cloned cDNA of the XRCC3 DNA repair gene. Proc. Natl. Acad. Sci. USA 1995, 92, 6354–6358. [Google Scholar] [CrossRef] [PubMed]
- Albala, J.S.; Thelen, M.P.; Prange, C.; Fan, W.; Christensen, M.; Thompson, L.H.; Lennon, G.G. Identification of a novel human RAD51 homolog, RAD51B. Genomics 1997, 46, 476–479. [Google Scholar] [CrossRef] [PubMed]
- Dosanjh, M.K.; Collins, D.W.; Fan, W.; Lennon, G.G.; Albala, J.S.; Shen, Z.; Schild, D. Isolation and characterization of RAD51C, a new human member of the RAD51 family of related genes. Nucl. Acids Res. 1998, 26, 1179–1184. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Lamerdin, J.E.; Tebbs, R.S.; Schild, D.; Tucker, J.D.; Shen, M.R.; Brookman, K.W.; Siciliano, M.J.; Walter, C.A.; Fan, W.; et al. XRCC2 and XRCC3, new human Rad51-family members, promote chromosome stability and protect against DNA cross-links and other damages. Mol. Cell 1998, 1, 783–793. [Google Scholar] [CrossRef]
- Pittman, D.L.; Weinberg, L.R.; Schimenti, J.C. Identification, characterization, and genetic mapping of Rad51d, a new mouse and human RAD51/RecA-related gene. Genomics 1998, 49, 103–111. [Google Scholar] [CrossRef]
- Schild, D.; Lio, Y.C.; Collins, D.W.; Tsomondo, T.; Chen, D.J. Evidence for simultaneous protein interactions between human Rad51 paralogs. J. Biol. Chem. 2000, 275, 16443–16449. [Google Scholar] [CrossRef]
- Masson, J.Y.; Tarsounas, M.C.; Stasiak, A.Z.; Stasiak, A.; Shah, R.; McIlwraith, M.J.; Benson, F.E.; West, S.C. Identification and purification of two distinct complexes containing the five RAD51 paralogs. Genes Dev. 2001, 15, 3296–3307. [Google Scholar] [CrossRef] [Green Version]
- Liu, N.; Schild, D.; Thelen, M.P.; Thompson, L.H. Involvement of Rad51C in two distinct protein complexes of Rad51 paralogs in human cells. Nucl. Acids Res. 2002, 30, 1009–1015. [Google Scholar] [CrossRef] [Green Version]
- Miller, K.A.; Sawicka, D.; Barsky, D.; Albala, J.S. Domain mapping of the Rad51 paralog protein complexes. Nucl. Acids Res. 2004, 32, 169–178. [Google Scholar] [CrossRef]
- Li, W.; Yang, X.; Lin, Z.; Timofejeva, L.; Xiao, R.; Makaroff, C.A.; Ma, H. The AtRAD51C gene is required for normal meiotic chromosome synapsis and double-stranded break repair in Arabidopsis. Plant Physiol. 2005, 138, 965–976. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Masson, J.Y.; Shah, R.; O’Regan, P.; West, S.C. RAD51C is required for Holliday junction processing in mammalian cells. Science 2004, 303, 243–246. [Google Scholar] [CrossRef] [PubMed]
- Kuznetsov, S.; Pellegrini, M.; Shuda, K.; Fernandez-Capetillo, O.; Liu, Y.; Martin, B.K.; Burkett, S.; Southon, E.; Pati, D.; Tessarollo, L.; et al. RAD51C deficiency in mice results in early prophase I arrest in males and sister chromatid separation at metaphase II in females. J. Cell Biol. 2007, 176, 581–592. [Google Scholar] [CrossRef] [PubMed]
- Suwaki, N.; Klare, K.; Tarsounas, M. RAD51 paralogs: Roles in DNA damage signalling, recombinational repair and tumorigenesis. Semin. Cell Dev. Biol. 2011, 22, 898–905. [Google Scholar] [CrossRef]
- Lio, Y.C.; Mazin, A.V.; Kowalczykowski, S.C.; Chen, D.J. Complex formation by the human Rad51B and Rad51C DNA repair proteins and their activities in vitro. J. Biol. Chem. 2003, 278, 2469–2478. [Google Scholar] [CrossRef] [PubMed]
- Wiese, C.; Collins, D.W.; Albala, J.S.; Thompson, L.H.; Kronenberg, A.; Schild, D. Interactions involving the Rad51 paralogs Rad51C and XRCC3 in human cells. Nucl. Acids Res. 2002, 30, 1001–1008. [Google Scholar] [CrossRef] [Green Version]
- Masson, J.Y.; Stasiak, A.Z.; Stasiak, A.; Benson, F.E.; West, S.C. Complex formation by the human RAD51C and XRCC3 recombination repair proteins. Proc. Natl. Acad. Sci. USA 2001, 98, 8440–8446. [Google Scholar] [CrossRef] [Green Version]
- Bishop, D.K.; Ear, U.; Bhattacharyya, A.; Calderone, C.; Beckett, M.; Weichselbaum, R.R.; Shinohara, A. Xrcc3 is required for assembly of Rad51 complexes in vivo. J. Biol. Chem. 1998, 273, 21482–21488. [Google Scholar] [CrossRef]
- Lin, Z.; Kong, H.; Nei, M.; Ma, H. Origins and evolution of the recA/RAD51 gene family: Evidence for ancient gene duplication and endosymbiotic gene transfer. Proc. Natl. Acad. Sci. USA 2006, 103, 10328–10333. [Google Scholar] [CrossRef]
- Abe, K.; Osakabe, K.; Nakayama, S.; Endo, M.; Tagiri, A.; Todoriki, S.; Ichikawa, H.; Toki, S. Arabidopsis RAD51C gene is important for homologous recombination in meiosis and mitosis. Plant Physiol. 2005, 139, 896–908. [Google Scholar] [CrossRef]
- Osakabe, K.; Yoshioka, T.; Ichikawa, H.; Toki, S. Molecular cloning and characterization of RAD51-like genes from Arabidopsis thaliana. Plant Mol. Biol. 2002, 50, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Bleuyard, J.Y.; Gallego, M.E.; Savigny, F.; White, C.I. Differing requirements for the Arabidopsis Rad51 paralogs in meiosis and DNA repair. Plant J. 2005, 41, 533–545. [Google Scholar] [CrossRef] [PubMed]
- Bleuyard, J.Y.; White, C.I. The Arabidopsis homologue of Xrcc3 plays an essential role in meiosis. EMBO J. 2004, 23, 439–449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, B.; Wang, M.; Tang, D.; Li, Y.; Xu, M.; Gu, M.; Cheng, Z.; Yu, H. XRCC3 is essential for proper double-strand break repair and homologous recombination in rice meiosis. J. Exp. Bot. 2015, 66, 5713–5725. [Google Scholar] [CrossRef] [Green Version]
- Tang, D.; Miao, C.; Li, Y.; Wang, H.; Liu, X.; Yu, H.; Cheng, Z. OsRAD51C is essential for double-strand break repair in rice meiosis. Front. Plant. Sci. 2014, 5, 167. [Google Scholar] [CrossRef] [Green Version]
- Kou, Y.; Chang, Y.; Li, X.; Xiao, J.; Wang, S. The rice RAD51C gene is required for the meiosis of both female and male gametocytes and the DNA repair of somatic cells. J. Exp. Bot. 2012, 63, 5323–5335. [Google Scholar] [CrossRef] [Green Version]
- Strable, J.; Scanlon, M.J. Maize (Zea mays): A model organism for basic and applied research in plant biology. Cold Spring Harb. Protoc. 2009, 2009, pdb.emo132. [Google Scholar] [CrossRef]
- Wang, C.J.; Harper, L.; Cande, W.Z. High-resolution single-copy gene fluorescence in situ hybridization and its use in the construction of a cytogenetic map of maize chromosome 9. Plant Cell 2006, 18, 529–544. [Google Scholar] [CrossRef]
- Pawlowski, W.P. Chromosome organization and dynamics in plants. Curr. Opin. Plant Biol. 2010, 13, 640–645. [Google Scholar] [CrossRef]
- Wang, C.J. Analyzing maize meiotic chromosomes with super-resolution structured illumination microscopy. Methods Mol. Biol. 2013, 990, 67–78. [Google Scholar] [CrossRef]
- Chang, P.; Tseng, Y.F.; Chen, P.Y.; Wang, C.R. Using Flow Cytometry to Isolate Maize Meiocytes for Next Generation Sequencing: A Time and Labor Efficient Method. Curr. Protoc. Plant. Biol. 2018, 3, e20068. [Google Scholar] [CrossRef] [PubMed]
- Chang, M.T.; Neuffer, M.G. Chromosomal Behavior During Microsporogenesis. In The Maize Handbook; Freeling, M., Walbot, V., Eds.; Springer: New York, NY, USA, 1994; pp. 460–475. [Google Scholar] [CrossRef]
- Timofejeva, L.; Skibbe, D.S.; Lee, S.; Golubovskaya, I.; Wang, R.; Harper, L.; Walbot, V.; Cande, W.Z. Cytological characterization and allelism testing of anther developmental mutants identified in a screen of maize male sterile lines. G3 (BethesdaMd.) 2013, 3, 231–249. [Google Scholar] [CrossRef] [PubMed]
- Alexander, M.P. Differential staining of aborted and nonaborted pollen. Stain Technol. 1969, 44, 117–122. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Liu, J.; Ren, W.; Yang, Q.; Chai, Z.; Chen, R.; Wang, L.; Zhao, J.; Lang, Z.; Wang, H.; et al. Gene-Indexed Mutations in Maize. Mol. Plant 2018, 11, 496–504. [Google Scholar] [CrossRef] [PubMed]
- Blokhina, Y.P.; Nguyen, A.D. The telomere bouquet is a hub where meiotic double-strand breaks, synapsis, and stable homolog juxtaposition are coordinated in the zebrafish, Danio rerio. PLoS Genet. 2019, 15, e1007730. [Google Scholar] [CrossRef]
- Golubovskaya, I.N.; Harper, L.C.; Pawlowski, W.P.; Schichnes, D.; Cande, W.Z. The pam1 gene is required for meiotic bouquet formation and efficient homologous synapsis in maize (Zea mays L.). Genetics 2002, 162, 1979–1993. [Google Scholar]
- Harper, L.; Golubovskaya, I.; Cande, W.Z. A bouquet of chromosomes. J. Cell Sci. 2004, 117, 4025–4032. [Google Scholar] [CrossRef] [Green Version]
- Klutstein, M.; Fennell, A.; Fernandez-Alvarez, A.; Cooper, J.P. The telomere bouquet regulates meiotic centromere assembly. Nat. Cell Biol. 2015, 17, 458–469. [Google Scholar] [CrossRef]
- Richards, E.J.; Ausubel, F.M. Isolation of a higher eukaryotic telomere from Arabidopsis thaliana. Cell 1988, 53, 127–136. [Google Scholar] [CrossRef]
- Prieto, P.; Martin, A.; Cabrera, A. Chromosomal distribution of telomeric and telomeric-associated sequences in Hordeum chilense by in situ hybridization. Hereditas 2004, 141, 122–127. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Arumuganathan, K. Physical mapping of 45S and 5S rDNA on maize metaphase and sorted chromosomes by FISH. Hereditas 2001, 134, 141–145. [Google Scholar] [CrossRef] [PubMed]
- Kato, A.; Lamb, J.C.; Birchler, J.A. Chromosome painting using repetitive DNA sequences as probes for somatic chromosome identification in maize. Proc. Natl. Acad. Sci. USA 2004, 101, 13554–13559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cahoon, C.K.; Hawley, R.S. Regulating the construction and demolition of the synaptonemal complex. Nat. Struct. Mol. Biol. 2016, 23, 369–377. [Google Scholar] [CrossRef] [PubMed]
- Golubovskaya, I.N.; Hamant, O.; Timofejeva, L.; Wang, C.J.; Braun, D.; Meeley, R.; Cande, W.Z. Alleles of afd1 dissect REC8 functions during meiotic prophase I. J. Cell Sci. 2006, 119, 3306–3315. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, S.J.; Caryl, A.P.; Jones, G.H.; Franklin, F.C. Asy1, a protein required for meiotic chromosome synapsis, localizes to axis-associated chromatin in Arabidopsis and Brassica. J. Cell Sci. 2002, 115, 3645–3655. [Google Scholar] [CrossRef] [PubMed]
- Golubovskaya, I.N.; Wang, C.J.; Timofejeva, L.; Cande, W.Z. Maize meiotic mutants with improper or non-homologous synapsis due to problems in pairing or synaptonemal complex formation. J. Exp. Bot. 2011, 62, 1533–1544. [Google Scholar] [CrossRef] [PubMed]
- Geric, M.; Gajski, G.; Garaj-Vrhovac, V. gamma-H2AX as a biomarker for DNA double-strand breaks in ecotoxicology. Ecotoxicol. Environ. Saf. 2014, 105, 13–21. [Google Scholar] [CrossRef]
- Turinetto, V.; Giachino, C. Multiple facets of histone variant H2AX: A DNA double-strand-break marker with several biological functions. Nucl. Acids Res. 2015, 43, 2489–2498. [Google Scholar] [CrossRef]
- Valdiglesias, V.; Giunta, S.; Fenech, M.; Neri, M.; Bonassi, S. gammaH2AX as a marker of DNA double strand breaks and genomic instability in human population studies. Mutat. Res. 2013, 753, 24–40. [Google Scholar] [CrossRef]
- Friesner, J.D.; Liu, B.; Culligan, K.; Britt, A.B. Ionizing radiation-dependent gamma-H2AX focus formation requires ataxia telangiectasia mutated and ataxia telangiectasia mutated and Rad3-related. Mol. Biol. Cell 2005, 16, 2566–2576. [Google Scholar] [CrossRef]
- Pawlowski, W.P.; Golubovskaya, I.N.; Cande, W.Z. Altered nuclear distribution of recombination protein RAD51 in maize mutants suggests the involvement of RAD51 in meiotic homology recognition. Plant. Cell 2003, 15, 1807–1816. [Google Scholar] [CrossRef] [PubMed]
- Etedali, F.; Baghban Kohnehrouz, B.; Valizadeh, M.; Gholizadeh, A.; Malboobi, M.A. Genome wide cloning of maize meiotic recombinase Dmc1 and its functional structure through molecular phylogeny. Genet. Mol. Res. GMR 2011, 10, 1636–1649. [Google Scholar] [CrossRef] [PubMed]
- De Silva, I.U.; McHugh, P.J.; Clingen, P.H.; Hartley, J.A. Defects in interstrand cross-link uncoupling do not account for the extreme sensitivity of ERCC1 and XPF cells to cisplatin. Nucl. Acids Res. 2002, 30, 3848–3856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasaki, M.S.; Takata, M.; Sonoda, E.; Tachibana, A.; Takeda, S. Recombination repair pathway in the maintenance of chromosomal integrity against DNA interstrand crosslinks. Cytogenet. Genome Res. 2004, 104, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Drexler, G.A.; Rogge, S.; Beisker, W.; Eckardt-Schupp, F.; Zdzienicka, M.Z.; Fritz, E. Spontaneous homologous recombination is decreased in Rad51C-deficient hamster cells. DNA Repair 2004, 3, 1335–1343. [Google Scholar] [CrossRef]
- Kuznetsov, S.G.; Haines, D.C.; Martin, B.K.; Sharan, S.K. Loss of Rad51c leads to embryonic lethality and modulation of Trp53-dependent tumorigenesis in mice. Cancer Res. 2009, 69, 863–872. [Google Scholar] [CrossRef]
- Smeenk, G.; de Groot, A.J.; Romeijn, R.J.; van Buul, P.P.; Zdzienicka, M.Z.; Mullenders, L.H.; Pastink, A.; Godthelp, B.C. Rad51C is essential for embryonic development and haploinsufficiency causes increased DNA damage sensitivity and genomic instability. Mutat. Res. 2010, 689, 50–58. [Google Scholar] [CrossRef]
- Min, A.; Im, S.A.; Yoon, Y.K.; Song, S.H.; Nam, H.J.; Hur, H.S.; Kim, H.P.; Lee, K.H.; Han, S.W.; Oh, D.Y.; et al. RAD51C-deficient cancer cells are highly sensitive to the PARP inhibitor olaparib. Mol. Cancer Ther. 2013, 12, 865–877. [Google Scholar] [CrossRef]
- Yonetani, Y.; Hochegger, H.; Sonoda, E.; Shinya, S.; Yoshikawa, H.; Takeda, S.; Yamazoe, M. Differential and collaborative actions of Rad51 paralog proteins in cellular response to DNA damage. Nucl. Acids Res. 2005, 33, 4544–4552. [Google Scholar] [CrossRef]
- Hu, Q.; Tang, D.; Wang, H. The Exonuclease Homolog OsRAD1 Promotes Accurate Meiotic Double-Strand Break Repair by Suppressing Nonhomologous End Joining. Plant Physiol. 2016, 172, 1105–1116. [Google Scholar]
- Hu, Q.; Zhang, C.; Xue, Z.; Ma, L.; Liu, W.; Shen, Y.; Ma, B.; Cheng, Z. OsRAD17 Is Required for Meiotic Double-Strand Break Repair and Plays a Redundant Role With OsZIP4 in Synaptonemal Complex Assembly. Front. Plant. Sci. 2018, 9, 1236. [Google Scholar] [CrossRef] [Green Version]
- Vannier, J.B.; Depeiges, A.; White, C.; Gallego, M.E. Two roles for Rad50 in telomere maintenance. EMBO J. 2006, 25, 4577–4585. [Google Scholar] [CrossRef] [Green Version]
- Samanic, I.; Simunic, J.; Riha, K.; Puizina, J. Evidence for distinct functions of MRE11 in Arabidopsis meiosis. PLoS ONE 2013, 8, e78760. [Google Scholar] [CrossRef]
- McVey, M.; Lee, S.E. MMEJ repair of double-strand breaks (director’s cut): Deleted sequences and alternative endings. Trends Genet. 2008, 24, 529–538. [Google Scholar] [CrossRef]
- Shrivastav, M.; De Haro, L.P.; Nickoloff, J.A. Regulation of DNA double-strand break repair pathway choice. Cell Res. 2008, 18, 134–147. [Google Scholar] [CrossRef]
- Yang, N.; Xu, X.W.; Wang, R.R.; Peng, W.L.; Cai, L.; Song, J.M.; Li, W.; Luo, X.; Niu, L.; Wang, Y.; et al. Contributions of Zea mays subspecies mexicana haplotypes to modern maize. Nat. Commun. 2017, 8, 1874. [Google Scholar] [CrossRef]
- Sun, S.; Zhou, Y.; Chen, J.; Shi, J. Extensive intraspecific gene order and gene structural variations between Mo17 and other maize genomes. Nat. Genet. 2018, 50, 1289–1295. [Google Scholar] [CrossRef] [Green Version]
- Johnson-Brousseau, S.A.; McCormick, S. A compendium of methods useful for characterizing Arabidopsis pollen mutants and gametophytically-expressed genes. Plant. J. 2004, 39, 761–775. [Google Scholar] [CrossRef]
- Ji, J.; Tang, D.; Wang, K.; Wang, M.; Che, L.; Li, M.; Cheng, Z. The role of OsCOM1 in homologous chromosome synapsis and recombination in rice meiosis. Plant. J. 2012, 72, 18–30. [Google Scholar] [CrossRef]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jing, J.; Zhang, T.; Wang, Y.; Cui, Z.; He, Y. ZmRAD51C Is Essential for Double-Strand Break Repair and Homologous Recombination in Maize Meiosis. Int. J. Mol. Sci. 2019, 20, 5513. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20215513
Jing J, Zhang T, Wang Y, Cui Z, He Y. ZmRAD51C Is Essential for Double-Strand Break Repair and Homologous Recombination in Maize Meiosis. International Journal of Molecular Sciences. 2019; 20(21):5513. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20215513
Chicago/Turabian StyleJing, Juli, Ting Zhang, Yazhong Wang, Zhenhai Cui, and Yan He. 2019. "ZmRAD51C Is Essential for Double-Strand Break Repair and Homologous Recombination in Maize Meiosis" International Journal of Molecular Sciences 20, no. 21: 5513. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20215513