Proteomic Analysis of Urinary Extracellular Vesicles Reveals a Role for the Complement System in Medullary Sponge Kidney Disease

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Characterization of Exosomes and Microvesicles
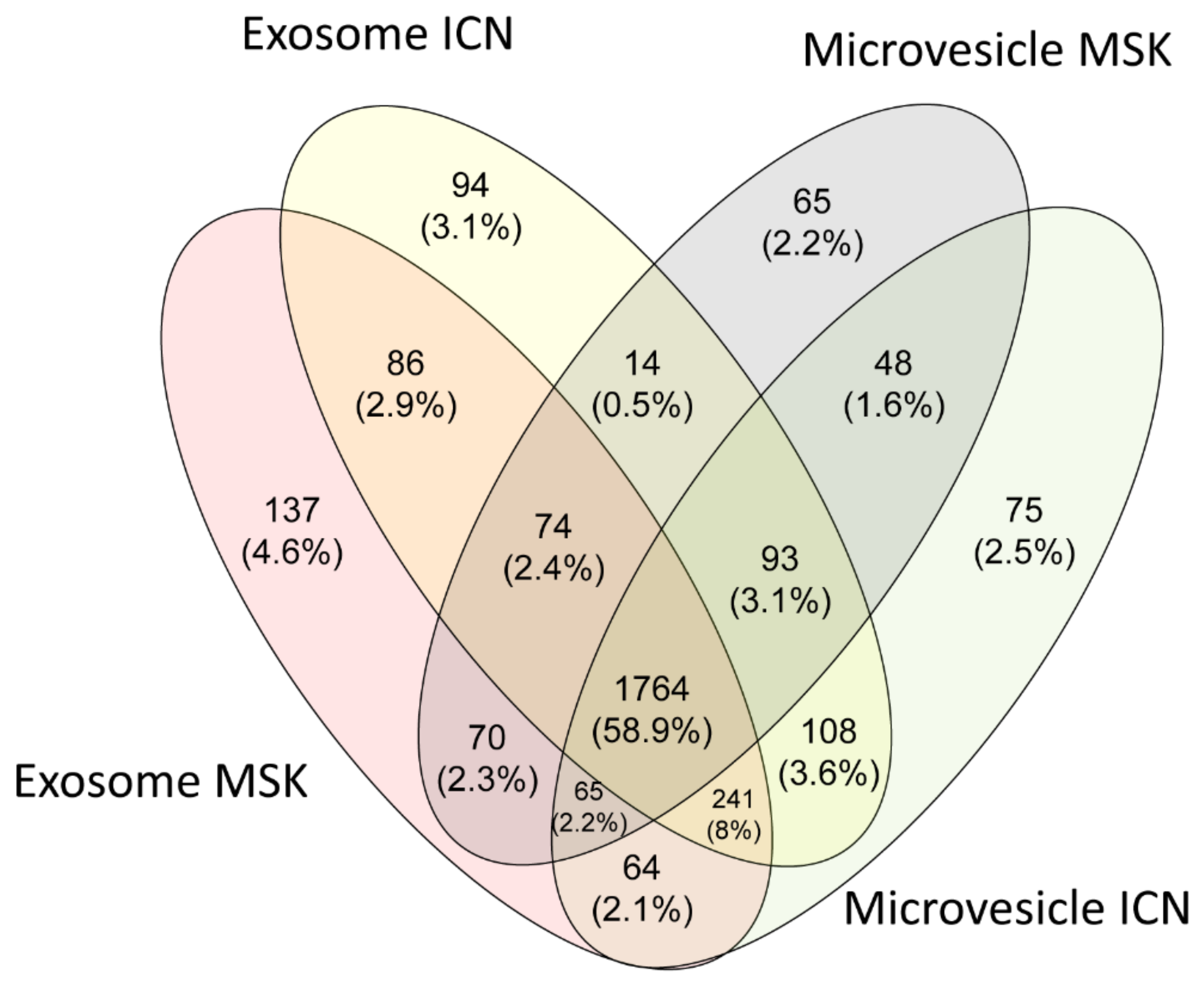
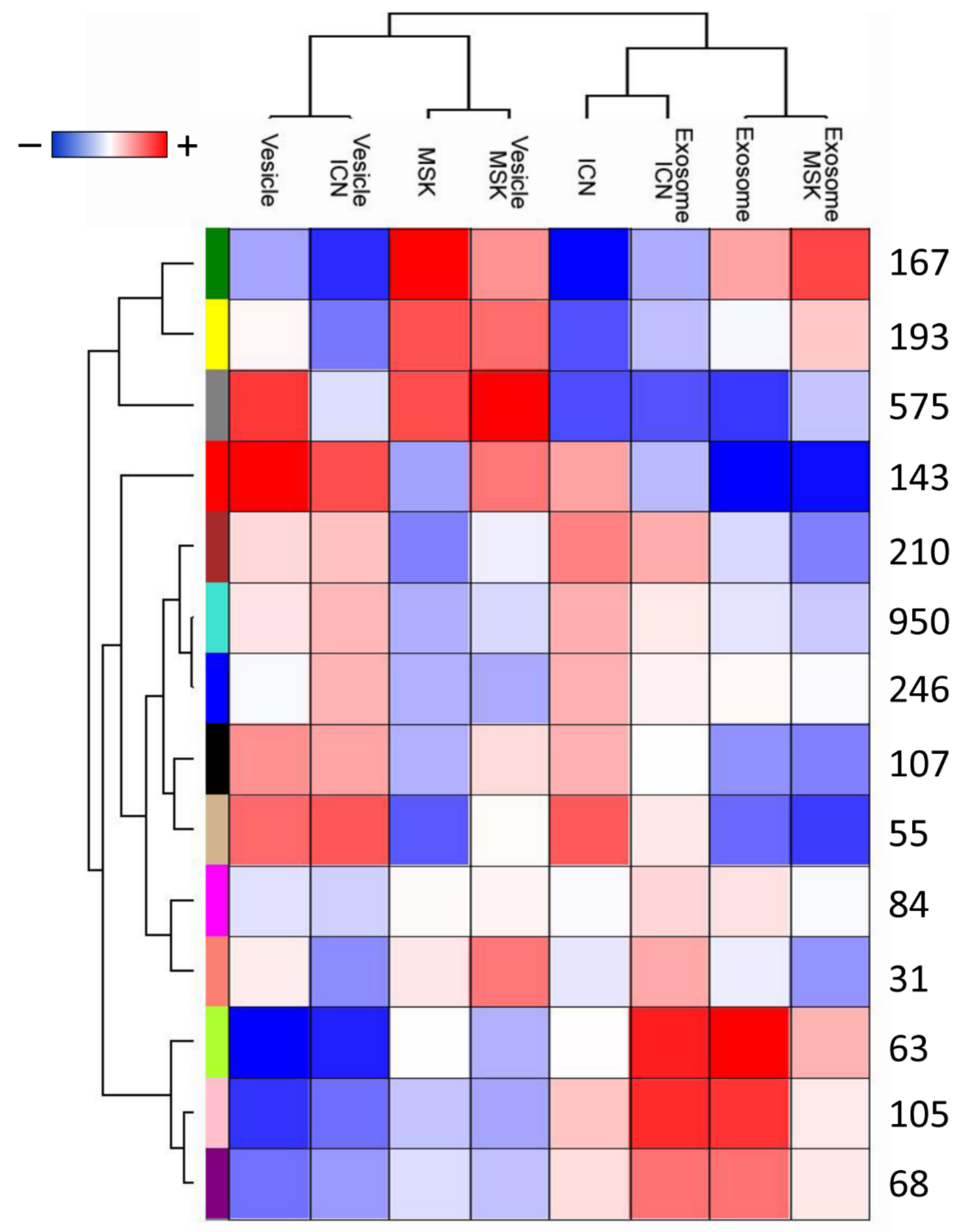
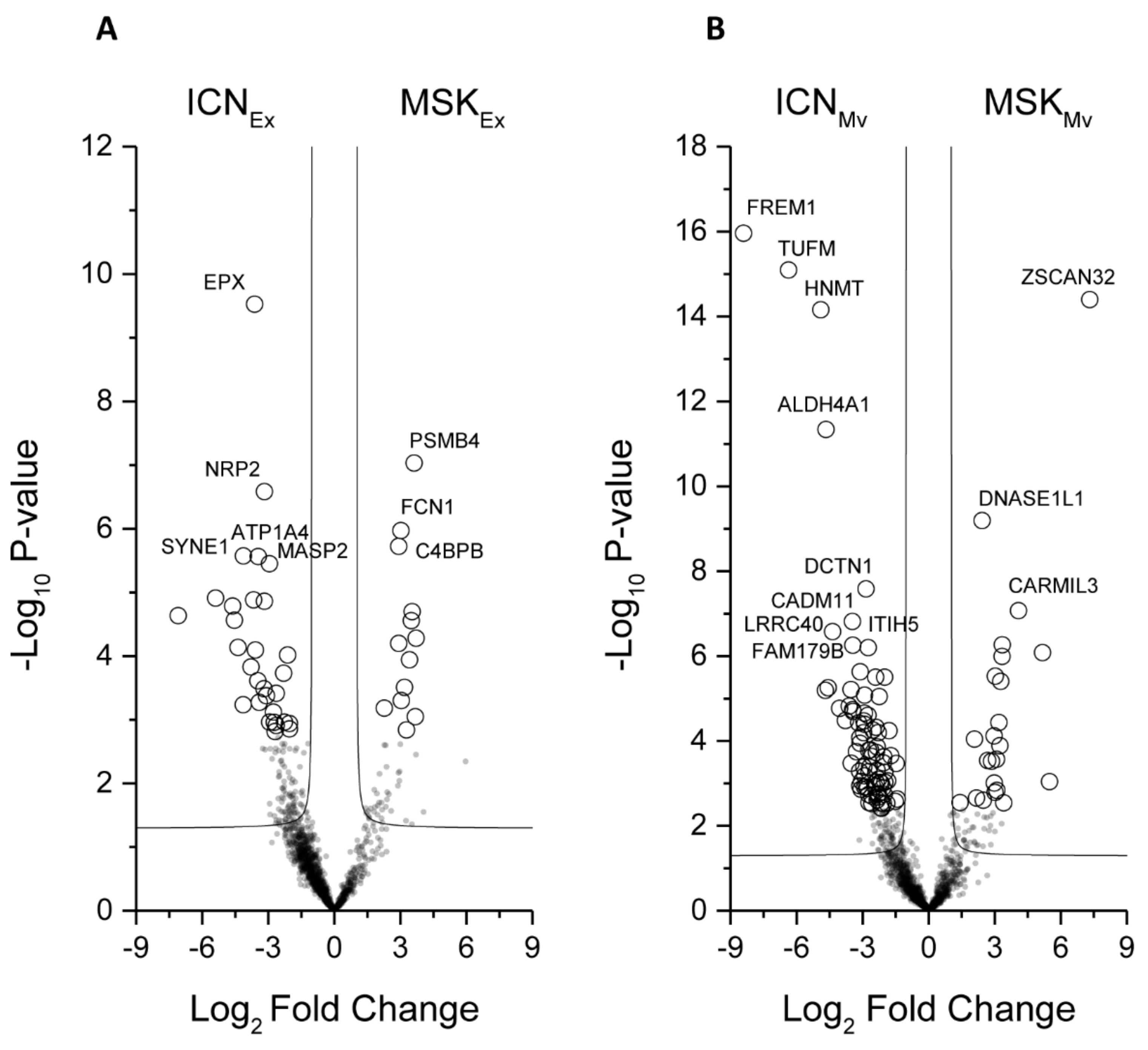
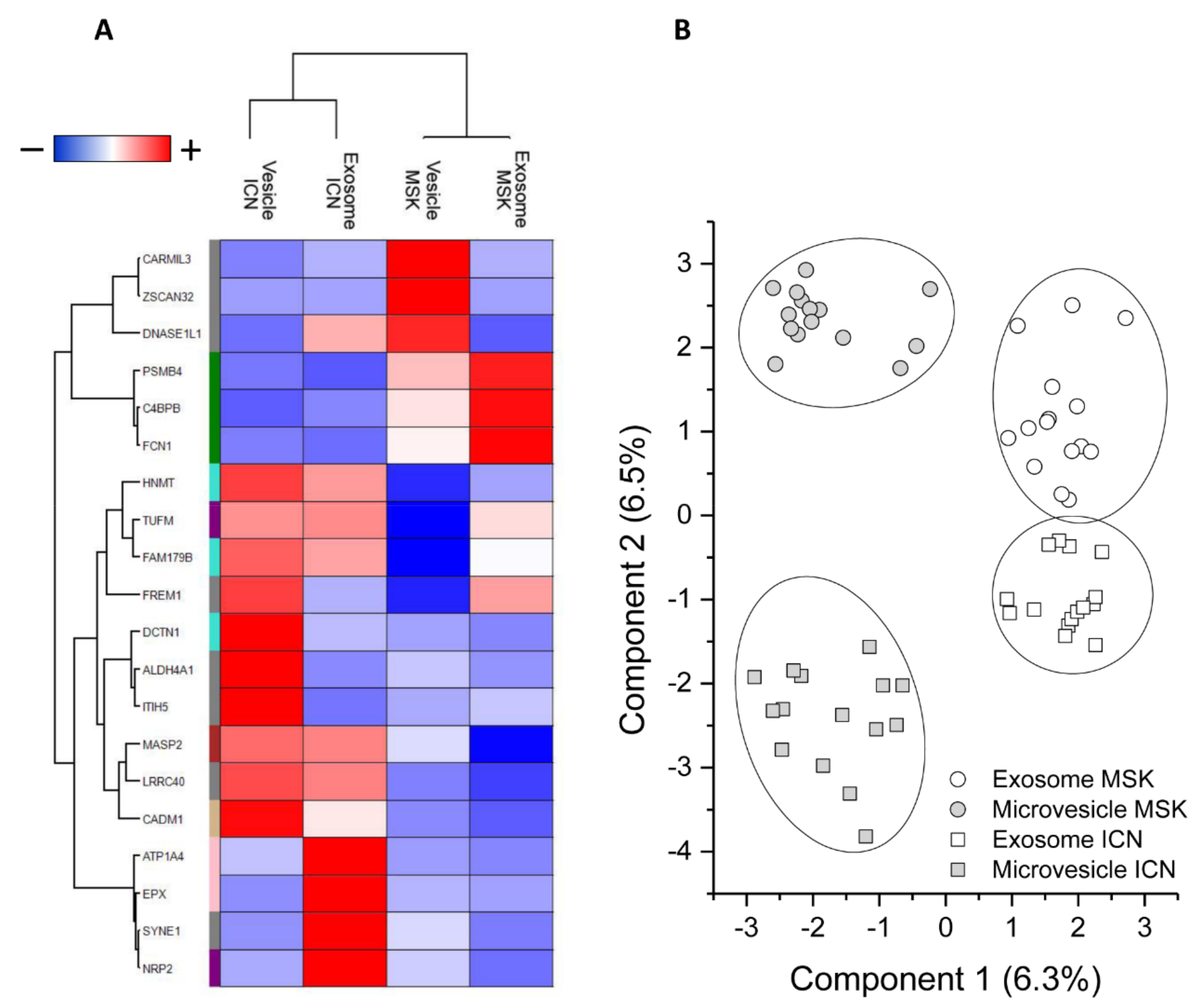
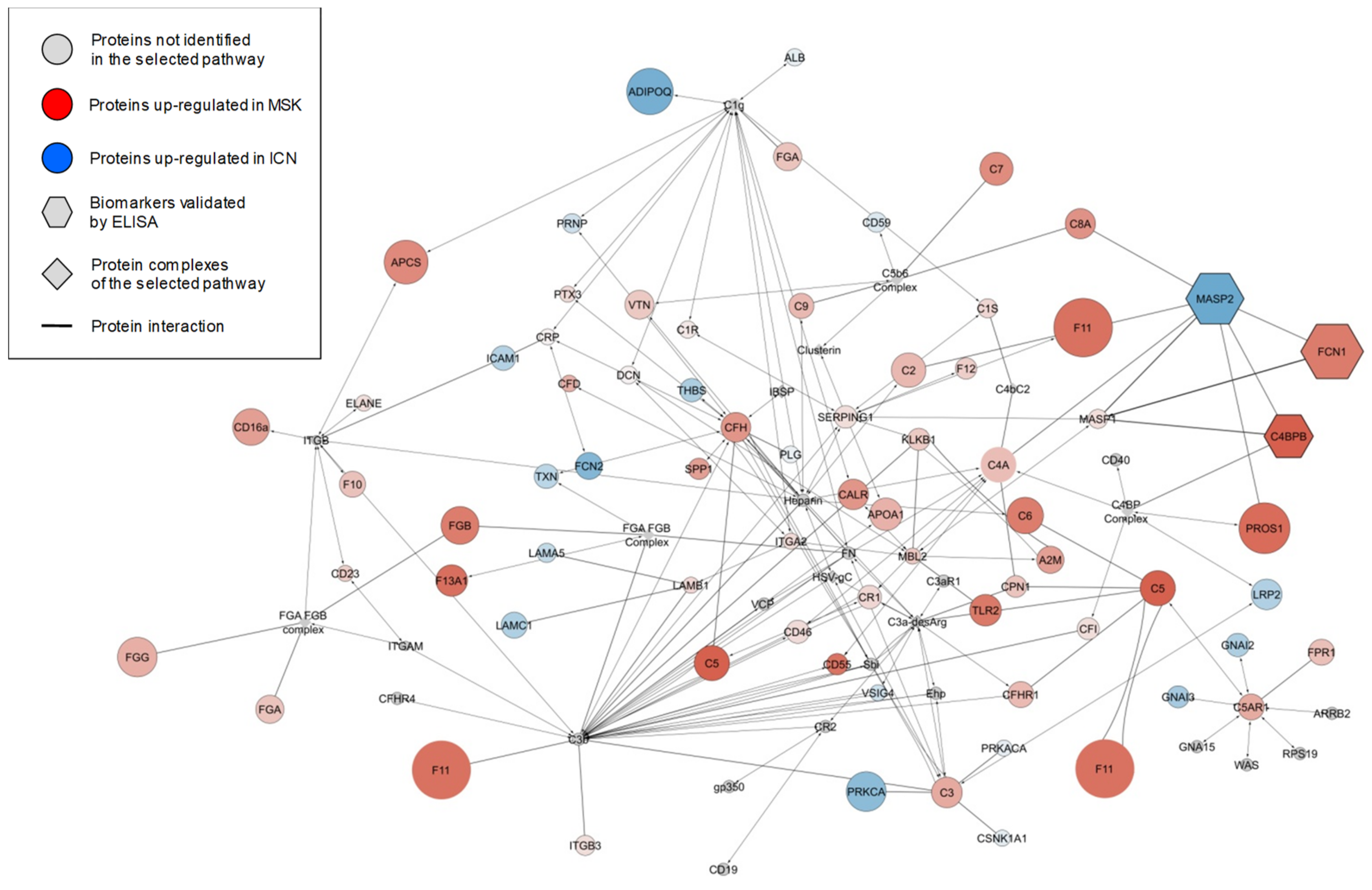
2.2. Protein Composition of Exosomes and Microvesicles
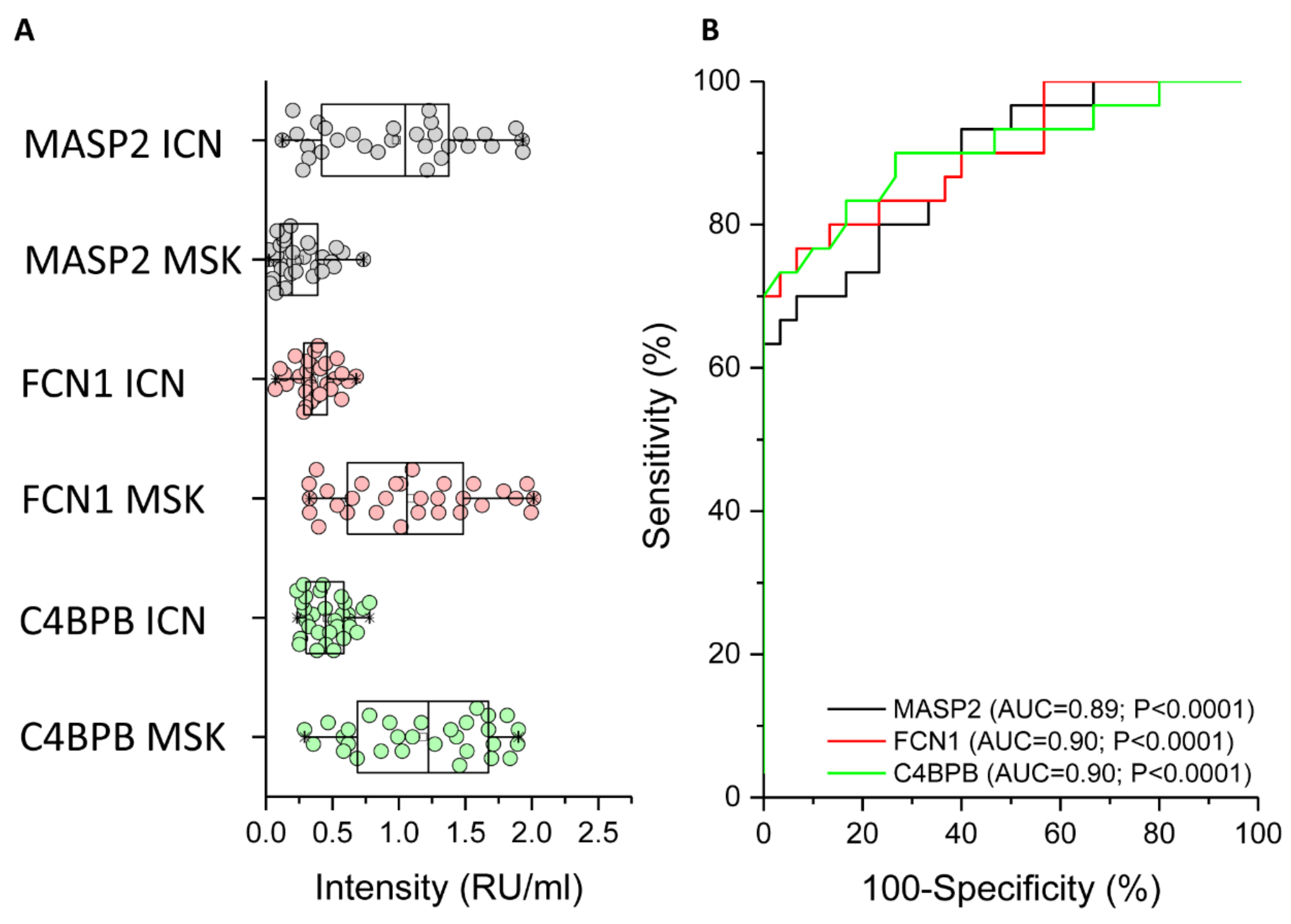
2.3. ELISA Analysis of MASP2, FCN1, and C4BPB to Confirm the MS Data
3. Discussion
4. Materials and Methods
4.1. Patients
4.2. Isolation of Microvesicles and Exosomes
4.3. Dynamic Light Scattering
4.4. Western Blot
4.5. Mass Spectrometry
4.6. MS Data Analysis
4.7. ELISA
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
Abbreviations
| MSK | medullary sponge kidney |
| ICN | idiopathic calcium nephrolithiasis |
| LAMA2 | laminin subunit α2 |
| SVM | support vector machine |
| PLS-DA | partial least squares discriminant analysis |
| ADPKD | autosomal dominant polycystic kidney disease |
| MS | mass spectrometry |
| SPP1 | osteopontin |
| PBS | phosphate-buffered saline |
| DLS | dynamic light scattering |
| iST | in-StageTip |
| SDB-RPS | poly (styrene divinylbenzene) reversed-phase sulfonate |
| TFA | trifluoroacetic acid |
| BSA | bovine serum albumin |
| PBST | PBS plus 0.1% Tween-20 |
| HRP | horseradish peroxidase |
| ROC | Received operating characteristic |
| AUC | area under the curve |
| CI | confidence interval |
| FCN1 | Ficolin 1 |
| MASP2 | Mannan-binding lectin serine protease 2 |
| C4BPB | Complement component 4-binding protein β |
| PTX3 | pentraxin 3 |
| CRP | C-reactive protein |
References
- Fabris, A.; Anglani, F.; Lupo, A.; Gambaro, G. Medullary sponge kidney: State of the art. Nephrol. Dial. Transplant. 2013, 28, 1111–1119. [Google Scholar] [CrossRef] [PubMed]
- Indridason, O.S.; Thomas, L.; Berkoben, M. Medullary sponge kidney associated with congenital hemihypertrophy. J. Am. Soc. Nephrol. 1996, 7, 1123–1130. [Google Scholar] [PubMed]
- El-Sawi, M.; Shahein, A.R. Medullary sponge kidney presenting in a neonate with distal renal tubular acidosis and failure to thrive: A case report. J. Med. Case Rep. 2009, 3, 6656. [Google Scholar] [CrossRef] [PubMed]
- Patriquin, H.B.; O’Regan, S. Medullary sponge kidney in childhood. AJR Am. J. Roentgenol. 1985, 145, 315–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lambrianides, A.L.; John, D.R. Medullary sponge disease in horseshoe kidney. Urology 1987, 29, 426–427. [Google Scholar] [CrossRef]
- Gambaro, G.; Fabris, A.; Citron, L.; Tosetto, E.; Anglani, F.; Bellan, F.; Conte, M.; Bonfante, L.; Lupo, A.; D’Angelo, A. An unusual association of contralateral congenital small kidney reduced renal function and hyperparathyroidism in sponge kidney patients: On the track of the molecular basis. Nephrol. Dial. Transplant. 2005, 20, 1042–1047. [Google Scholar] [CrossRef] [PubMed]
- Janjua, M.U.; Long, X.D.; Mo, Z.H.; Dong, C.S.; Jin, P. Association of medullary sponge kidney and hyperparathyroidism with RET G691S/S904S polymorphism: A case report. J. Med. Case Rep. 2018, 12, 197. [Google Scholar] [CrossRef]
- Ria, P.; Fabris, A.; Dalla Gassa, A.; Zaza, G.; Lupo, A.; Gambaro, G. New non-renal congenital disorders associated with medullary sponge kidney (MSK) support the pathogenic role of GDNF and point to the diagnosis of MSK in recurrent stone formers. Urolithiasis 2017, 45, 359–362. [Google Scholar] [CrossRef]
- Osther, P.J.; Hansen, A.B.; Røhl, H.F. Renal acidification defects in medullary sponge kidney. Br. J. Urol. 1988, 61, 392–394. [Google Scholar] [CrossRef]
- Fabris, A.; Bernich, P.; Abaterusso, C.; Marchionna, N.; Canciani, C.; Nouvenne, A.; Zamboni, M.; Lupo, A.; Gambaro, G. Bone disease in medullary sponge kidney and effect of potassium citrate treatment. Clin. J. Am. Soc. Nephrol. 2009, 4, 1974–1979. [Google Scholar] [CrossRef]
- McPhail, E.F.; Gettman, M.T.; Patterson, D.E.; Rangel, L.J.; Krambeck, A.E. Nephrolithiasis in medullary sponge kidney: Evaluation of clinical and metabolic features. Urology 2012, 79, 277–281. [Google Scholar] [CrossRef] [PubMed]
- Fabris, A.; Bruschi, M.; Santucci, L.; Candiano, G.; Granata, S.; Dalla Gassa, A.; Antonucci, N.; Petretto, A.; Ghiggeri, G.M.; Gambaro, G.; et al. Proteomic-based research strategy identified laminin subunit alpha 2 as a potential urinary-specific biomarker for the medullary sponge kidney disease. Kidney Int. 2017, 91, 459–468. [Google Scholar] [CrossRef] [PubMed]
- Colognato, H.; Yurchenco, P.D. Form and function: The laminin family of heterotrimers. Dev. Dyn. 2000, 218, 213–234. [Google Scholar] [CrossRef]
- Vijayakumar, S.; Dang, S.; Marinkovich, M.P.; Lazarova, Z.; Yoder, B.; Torres, V.E.; Wallace, D.P. Aberrant expression of laminin-332 promotes cell proliferation and cyst growth in ARPKD. Am. J. Physiol. Renal. Physiol. 2014, 306, F640–F654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joly, D.; Berissi, S.; Bertrand, A.; Strehl, L.; Patey, N.; Knebelmann, B. Laminin 5 regulates polycystic kidney cell proliferation and cyst formation. J. Biol. Chem. 2006, 281, 29181–29189. [Google Scholar] [CrossRef]
- Shannon, M.B.; Patton, B.L.; Harvey, S.J.; Miner, J.H. A hypomorphic mutation in the mouse laminin alpha5 gene causes polycystic kidney disease. J. Am. Soc. Nephrol. 2006, 17, 1913–1922. [Google Scholar] [CrossRef]
- Weir, M.L.; Oppizzi, M.L.; Henry, M.D.; Onishi, A.; Campbell, K.P.; Bissell, M.J.; Muschler, J.L. Dystroglycan loss disrupts polarity and beta-casein induction in mammary epithelial cells by perturbing laminin anchoring. J. Cell Sci. 2006, 119 (Pt 19), 4047–4058. [Google Scholar] [CrossRef]
- Bruschi, M.; Granata, S.; Santucci, L.; Candiano, G.; Fabris, A.; Antonucci, N.; Petretto, A.; Bartolucci, M.; Del Zotto, G.; Antonini, F.; et al. Proteomic Analysis of Urinary Microvesicles and Exosomes in Medullary Sponge Kidney Disease and Autosomal Dominant Polycystic Kidney Disease. Clin. J. Am. Soc. Nephrol. 2019, 14, 834–843. [Google Scholar] [CrossRef]
- Kleinman, J.G.; Wesson, J.A.; Hughes, J. Osteopontin and calcium stone formation. Nephron Physiol. 2004, 98, 43–47. [Google Scholar] [CrossRef]
- Verhulst, A.; Persy, V.P.; Van Rompay, A.R.; Verstrepen, W.A.; Helbert, M.F.; De Broe, M.E. Osteopontin synthesis and localization along the human Nephron. J. Am. Soc. Nephrol. 2002, 13, 1210–1218. [Google Scholar]
- Worcester, E.M.; Beshensky, A.M. Osteopontin inhibits nucleation of calcium oxalate crystals. Ann. N. Y. Acad. Sci. 1995, 760, 375–387. [Google Scholar] [CrossRef] [PubMed]
- Wesson, J.A.; Worcester, E. Formation of hydrated calcium oxalates in the presence of poly-l-aspartic acid. Scan. Microsc. 1996, 10, 415–424. [Google Scholar]
- Cowley, B.D., Jr.; Ricardo, S.D.; Nagao, S.; Diamond, J.R. Increased renal expression of monocyte chemoattractant protein-1 and osteopontin in ADPKD in rats. Kidney Int. 2001, 60, 2087–2096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gauer, S.; Urbschat, A.; Gretz, N.; Hoffmann, S.C.; Kränzlin, B.; Geiger, H.; Obermüller, N. Kidney Injury Molecule-1 Is Specifically Expressed in Cystically-Transformed Proximal Tubules of the PKD/Mhm (cy/+) Rat Model of Polycystic Kidney Disease. Int. J. Mol. Sci. 2016, 17, 802. [Google Scholar] [CrossRef] [PubMed]
- Pisitkun, T.; Shen, R.F.; Knepper, M.A. Identification and proteomic profiling of exosomes in human urine. Proc. Natl. Acad. Sci. USA 2004, 101, 13368–13373. [Google Scholar] [CrossRef] [Green Version]
- Moon, P.G.; You, S.; Lee, J.E.; Hwang, D.; Baek, M.C. Urinary exosomes and proteomics. Mass Spectrom. Rev. 2011, 30, 1185–1202. [Google Scholar] [CrossRef]
- Piñero, J.; Bravo, À.; Queralt-Rosinach, N.; Gutiérrez-Sacristán, A.; Deu-Pons, J.; Centeno, E.; García-García, J.; Sanz, F.; Furlong, L.I. DisGeNET: A comprehensive platform integrating information on human disease-associated genes and variants. Nucleic Acids Res. 2017, 45, D833–D839. [Google Scholar] [CrossRef]
- Salih, M.; Demmers, J.A.; Bezstarosti, K.; Leonhard, W.N.; Losekoot, M.; van Kooten, C.; Gansevoort, R.T.; Peters, D.J.; Zietse, R.; Hoorn, E.J.; et al. Proteomics of Urinary Vesicles Links Plakins and Complement to Polycystic Kidney Disease. J. Am. Soc. Nephrol. 2016, 27, 3079–3092. [Google Scholar] [CrossRef] [Green Version]
- Runza, V.L.; Schwaeble, W.; Männel, D.N. Ficolins: Novel pattern recognition molecules of the innate immune response. Immunobiology 2008, 213, 297–306. [Google Scholar] [CrossRef]
- Kjaer, T.R.; Hansen, A.G.; Sørensen, U.B.; Nielsen, O.; Thiel, S.; Jensenius, J.C. Investigations on the pattern recognition molecule M-ficolin: Quantitative aspects of bacterial binding and leukocyte association. J. Leukoc. Biol. 2011, 90, 425–437. [Google Scholar] [CrossRef]
- Thiel, S. Complement activating soluble pattern recognition molecules with collagen-like regions, mannan-binding lectin, ficolins and associated proteins. Mol. Immunol. 2007, 44, 3875–3888. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Yang, L.; Ang, Z.; Yoong, S.L.; Tran, T.T.; Anand, G.S.; Tan, N.S.; Ho, B.; Ding, J.L. Secreted M-ficolin anchors onto monocyte transmembrane G protein-coupled receptor 43 and cross talks with plasma C-reactive protein to mediate immune signaling and regulate host defense. J. Immunol. 2010, 185, 6899–6910. [Google Scholar] [CrossRef] [PubMed]
- Tanio, M.; Wakamatsu, K.; Kohno, T. Binding site of C-reactive protein on M-ficolin. Mol. Immunol. 2009, 47, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Gout, E.; Moriscot, C.; Doni, A.; Dumestre-Pérard, C.; Lacroix, M.; Pérard, J.; Schoehn, G.; Mantovani, A.; Arlaud, G.J.; Thielens, N.M. M-ficolin interacts with the long pentraxin PTX3: A novel case of cross-talk between soluble pattern-recognition molecules. J. Immunol. 2011, 186, 5815–5822. [Google Scholar] [CrossRef]
- Zhou, J.; Ouyang, X.; Schoeb, T.R.; Bolisetty, S.; Cui, X.; Mrug, S.; Yoder, B.K.; Johnson, M.R.; Szalai, A.J.; Mrug, M. Kidney injury accelerates cystogenesis via pathways modulated by heme oxygenase and complement. J. Am. Soc. Nephrol. 2012, 23, 1161–1171. [Google Scholar] [CrossRef] [PubMed]
- Su, Z.; Wang, X.; Gao, X.; Liu, Y.; Pan, C.; Hu, H.; Beyer, R.P.; Shi, M.; Zhou, J.; Zhang, J.; et al. Excessive activation of the alternative complement pathway in autosomal dominant polycystic kidney disease. J. Intern. Med. 2014, 276, 470–485. [Google Scholar] [CrossRef] [Green Version]
- Lai, X.; Bacallao, R.L.; Blazer-Yost, B.L.; Hong, D.; Mason, S.B.; Witzmann, F.A. Characterization of the renal cyst fluid proteome in autosomal dominant polycystic kidney disease (ADPKD) patients. Proteom. Clin. Appl. 2008, 2, 1140–1152. [Google Scholar] [CrossRef] [Green Version]
- Bakun, M.; Niemczyk, M.; Domanski, D.; Jazwiec, R.; Perzanowska, A.; Niemczyk, S.; Kistowski, M.; Fabijanska, A.; Borowiec, A.; Paczek, L.; et al. Urine proteome of autosomal dominant polycystic kidney disease patients. Clin. Proteom. 2012, 9, 13. [Google Scholar] [CrossRef]
- Rawal, N.; Rajagopalan, R.; Salvi, V.P. Stringent regulation of complement lectin pathway C3/C5 convertase by C4b-binding protein (C4BP). Mol. Immunol. 2009, 46, 2902–2910. [Google Scholar] [CrossRef] [Green Version]
- Salvadori, M.; Rosso, G.; Bertoni, E. Complement involvement in kidney diseases: From physiopathology to therapeutical targeting. World J. Nephrol. 2015, 4, 169–184. [Google Scholar] [CrossRef]
- Nolan, J.P.; Duggan, E. Analysis of Individual Extracellular Vesicles by Flow Cytometry. Methods Mol. Biol. 2018, 1678, 79–92. [Google Scholar] [PubMed]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bruschi, M.; Granata, S.; Candiano, G.; Fabris, A.; Petretto, A.; Ghiggeri, G.M.; Gambaro, G.; Zaza, G. Proteomic Analysis of Urinary Extracellular Vesicles Reveals a Role for the Complement System in Medullary Sponge Kidney Disease. Int. J. Mol. Sci. 2019, 20, 5517. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20215517
Bruschi M, Granata S, Candiano G, Fabris A, Petretto A, Ghiggeri GM, Gambaro G, Zaza G. Proteomic Analysis of Urinary Extracellular Vesicles Reveals a Role for the Complement System in Medullary Sponge Kidney Disease. International Journal of Molecular Sciences. 2019; 20(21):5517. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20215517
Chicago/Turabian StyleBruschi, Maurizio, Simona Granata, Giovanni Candiano, Antonia Fabris, Andrea Petretto, Gian Marco Ghiggeri, Giovanni Gambaro, and Gianluigi Zaza. 2019. "Proteomic Analysis of Urinary Extracellular Vesicles Reveals a Role for the Complement System in Medullary Sponge Kidney Disease" International Journal of Molecular Sciences 20, no. 21: 5517. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20215517