Low Molecular Seleno-Aminopolysaccharides Protect the Intestinal Mucosal Barrier of Rats under Weaning Stress

Abstract
:1. Introduction
2. Results
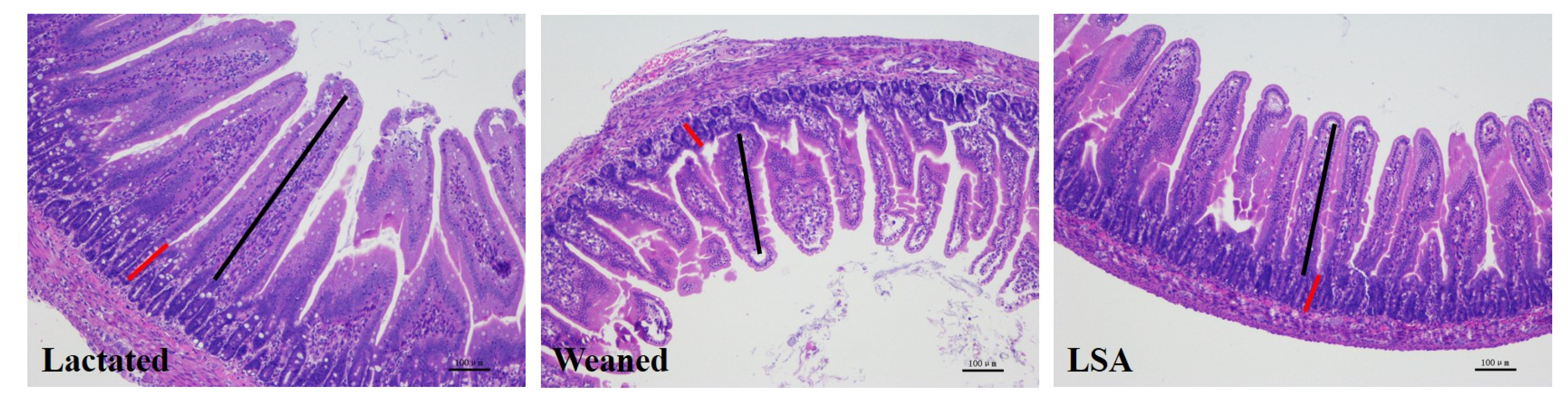
2.1. Effects of LSA on Intestinal Tissue Morphology of Weaning Rats
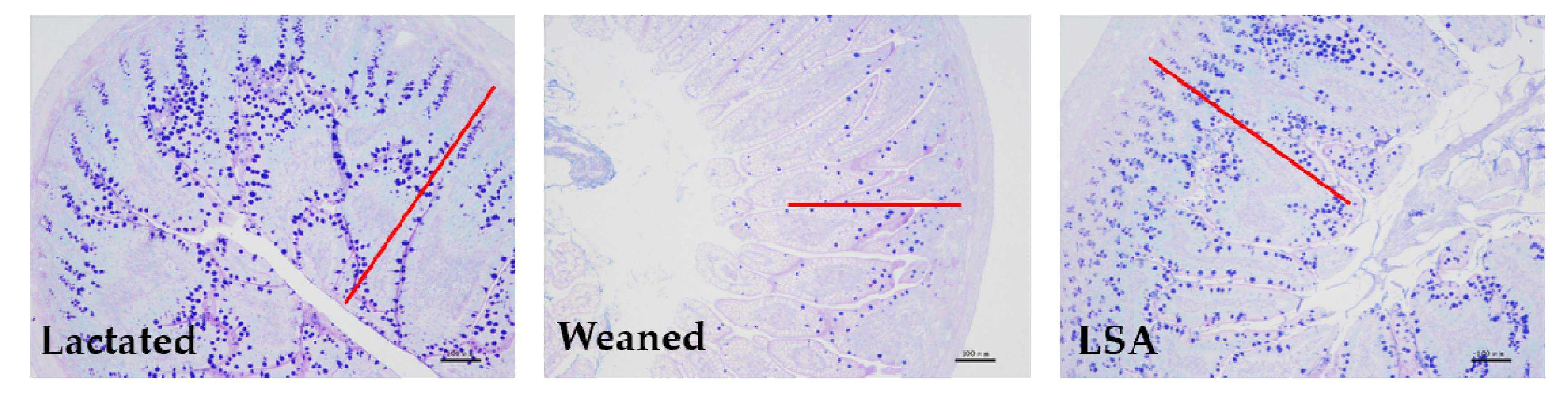
2.2. Effects of LSA on Number of Goblet Cells and Intestinal Mucosal Thickness in Weaning Rats
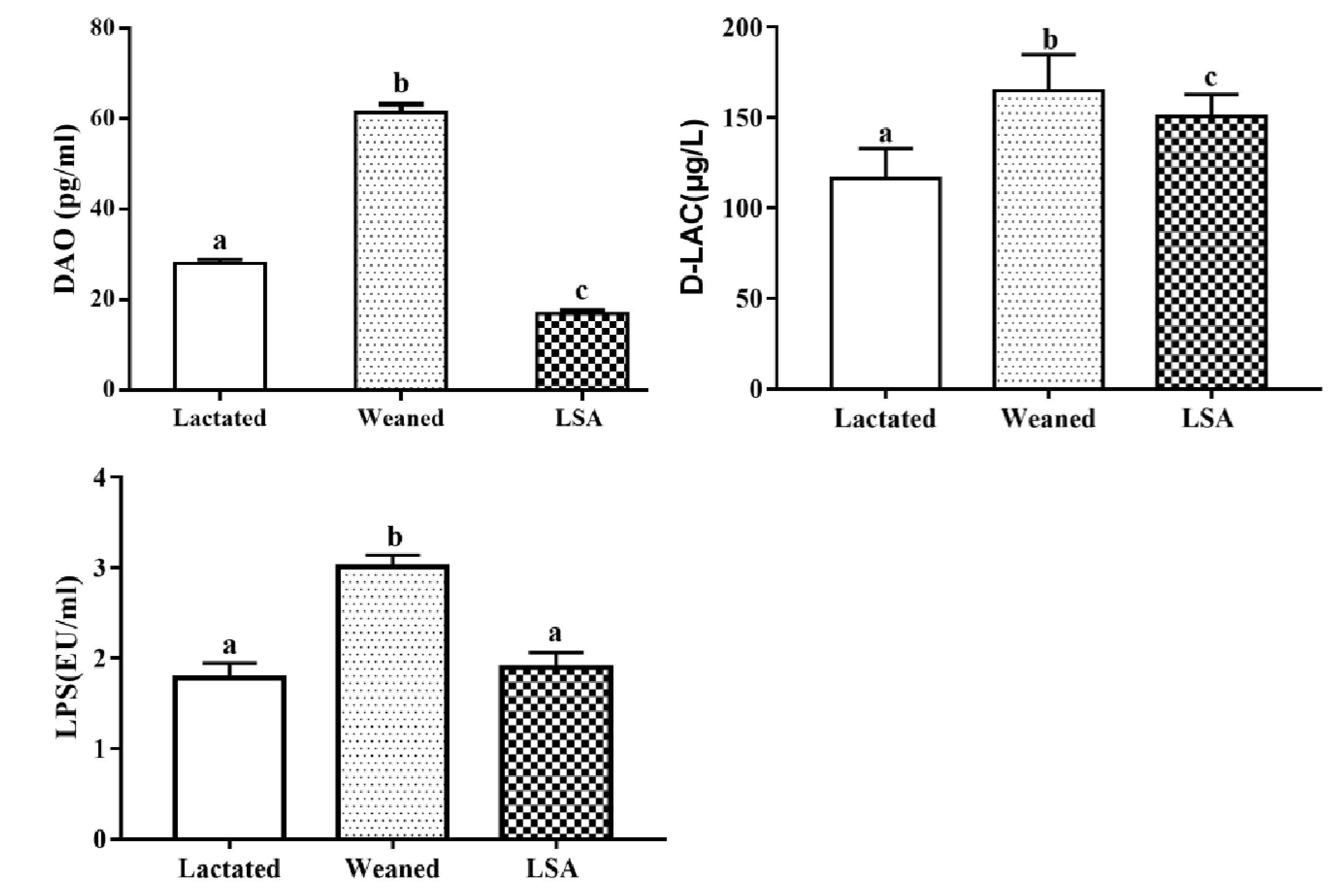
2.3. Effects of LSA on the Level of DAO, D-LA and LPS in Serum
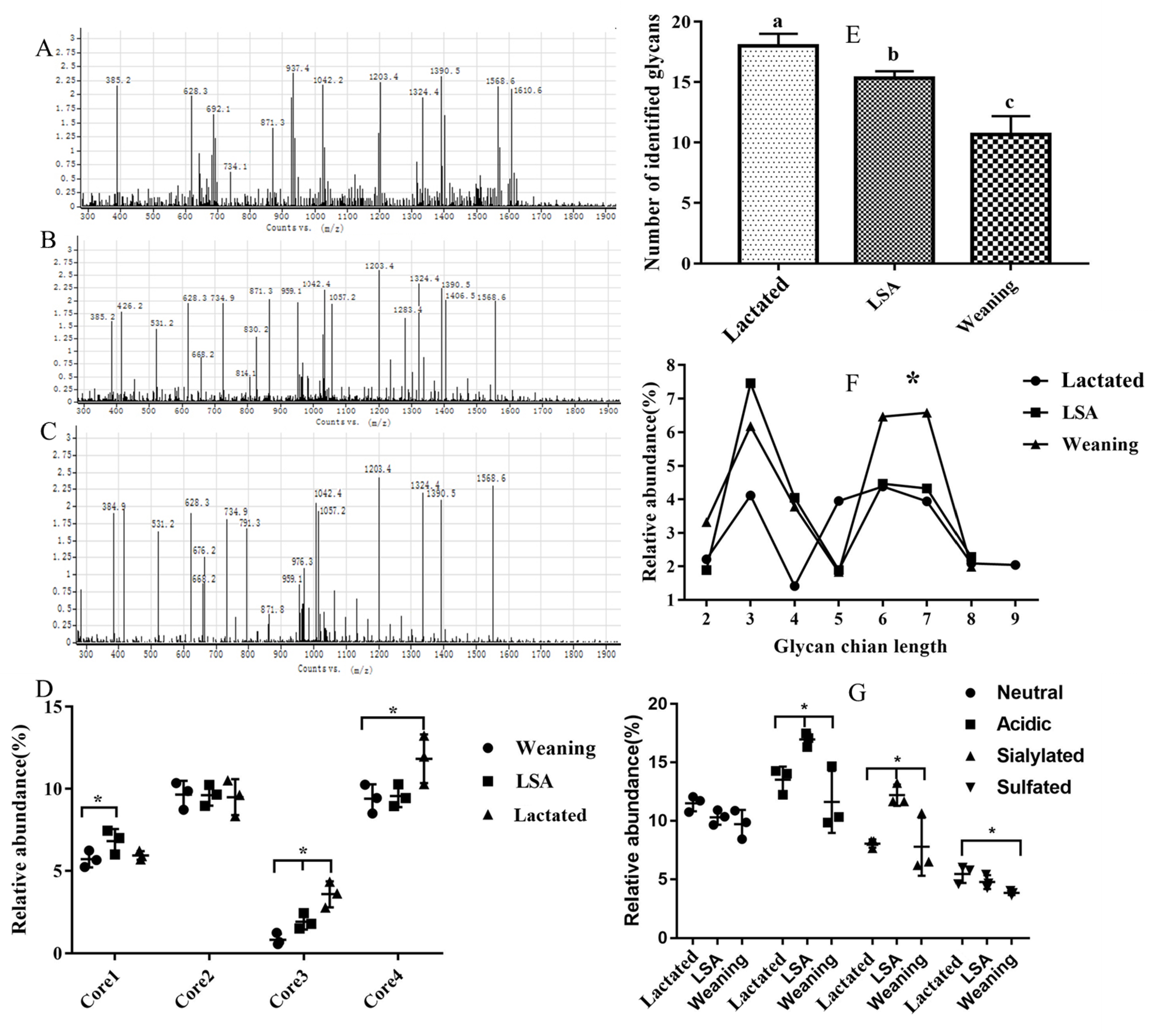
2.4. Effects of LSA on the Structure of Intestinal Mucin MUC2
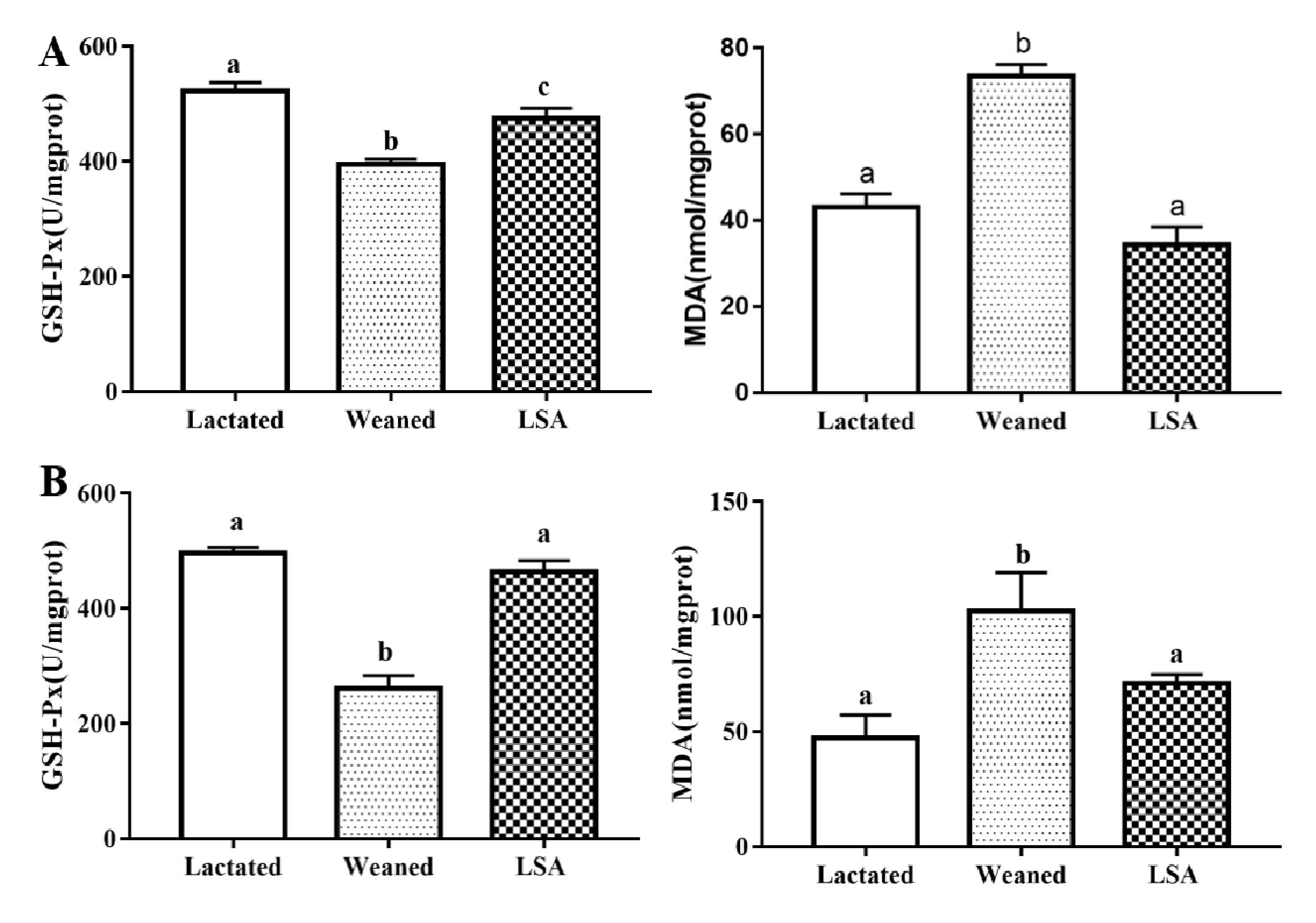
2.5. Effects of LSA on the Antioxidant Indexes in Serum
2.6. Effects of LSA on the Antioxidant Indexes in Jejunum and Ileum Homogenate
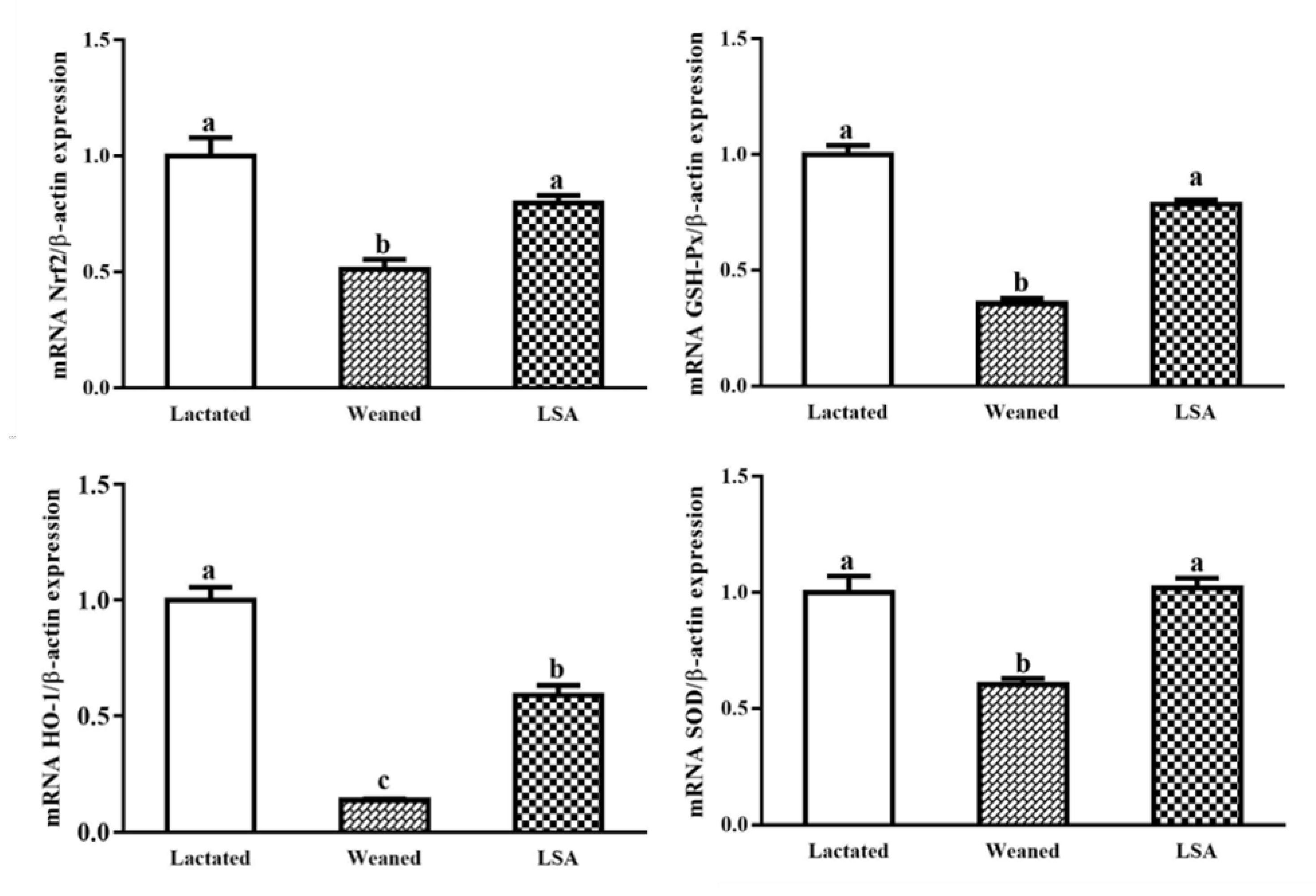
2.7. Effects of LSA on the Expression of Antioxidant Genes in Ileum
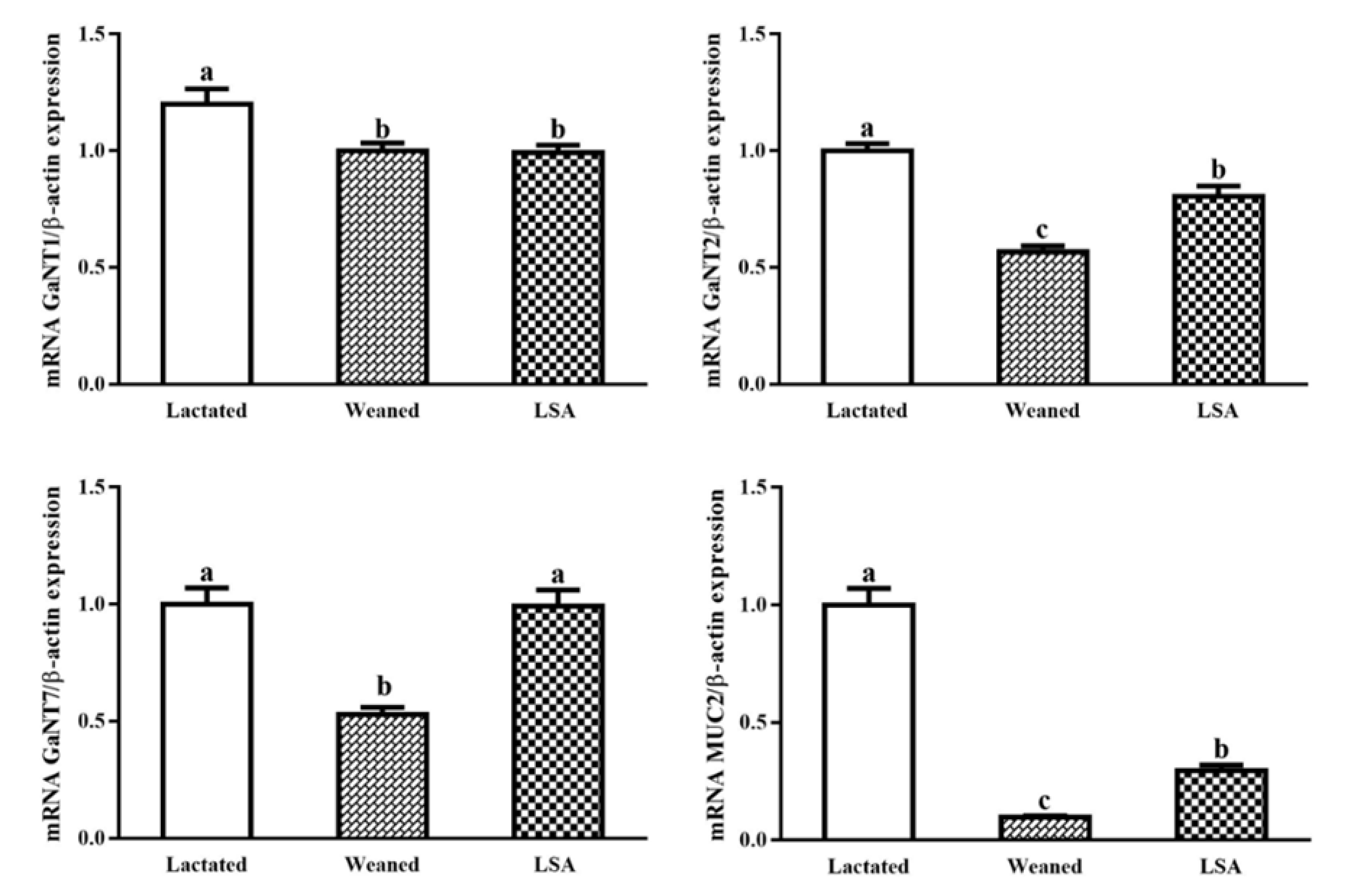
2.8. Effect of LSA on mRNA Expression of Glycosyltransferase and Mucin Genes in Intestinal Mucosa of Weaning Rats
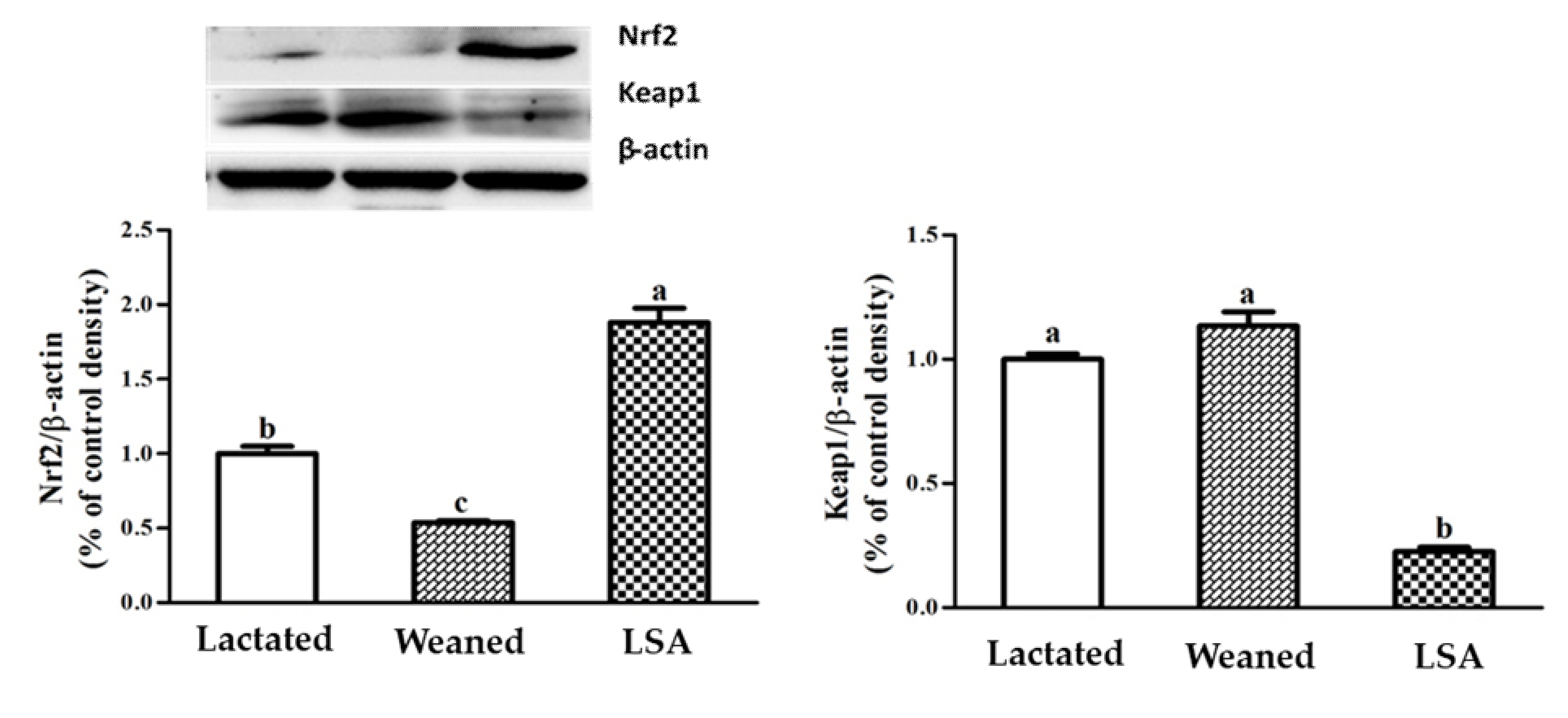
2.9. Effects of LSA Expression Levels of Transcription Factors in Intestinal Mucosa of Weaning Rats
3. Discussion
4. Materials and Methods
4.1. Materials and Reagents
4.2. Experimental Animals
4.3. Sample Collection
4.4. Histopathological Observation
4.5. Thickness of Intestinal Mucosa Layer and the Distribution and Number of Goblet Cells Were Observed by AB-PAS Staining
4.6. Determination of Intestinal Permeability by Detecting DAO, D-LA, LPS Levels in Serum
4.7. Determination of the Structure of Intestinal Mucin MUC2
4.8. Determination of Antioxidant indexes in Serum
4.9. Quantitative Real-Time Polymerase Chain Reaction (RT-PCR)
4.10. Western Blot Analysis
4.11. Statistical Analysis of Data
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- McLamb, B.L.; Gibson, A.J.; Overman, E.L.; Stahl, C.; Moeser, A.J. Early weaning stress in pigs impairs innate mucosal immune responses to enterotoxigenic E. coli challenge and exacerbates intestinal injury and clinical disease. PLoS ONE 2013, 8, e59838. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, A.L.; Blikslager, A.T. Impaired intestinal barrier function and relapsing digestive disease: Lessons from a porcine model of early life stress. Neurogastroenterol. Motil. 2017, 29, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Demaude, J.; Salvador-Cartier, C.; Fioramonti, J.; Ferrier, L.; Bueno, L. Phenotypic changes in colonocytes following acute stress or activation of mast cells in mice: Implications for delayed epithelial barrier dysfunction. Gut 2006, 55, 655–661. [Google Scholar] [CrossRef]
- Moeser, A.J.; Klok, C.V.; Ryan, K.A.; Wooten, J.G.; Little, D.; Cook, V.L.; Blikslager, A.T. Stress signaling pathways activated by weaning mediate intestinal dysfunction in the pig. Am. J. Physiol. Liver Physiol. 2007, 292, G173–G181. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.H.; Xiao, K.; Luan, Z.S.; Song, J. Early weaning increases intestinal permeability, alters expression of cytokine and tight junction proteins, and activates mitogen-activated protein kinases in pigs1. J. Anim. Sci. 2013, 91, 1094–1101. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Jia, Z.; Misra, H.; Li, Y.R. Oxidative stress and redox signaling mechanisms of alcoholic liver disease: Updated experimental and clinical evidence. J. Dig. Dis. 2012, 13, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Grisham, M. Oxidants and free radicals in inflammatory bowel disease. Lancet 1994, 344, 859–861. [Google Scholar] [CrossRef]
- Ziegler, A.; Gonzalez, L.; Blikslager, A. Large Animal Models: The Key to Translational Discovery in Digestive Disease Research. Cell. Mol. Gastroenterol. Hepatol. 2016, 2, 716–724. [Google Scholar] [CrossRef]
- Papp, L.V.; Lu, J.; Holmgren, A.; Khanna, K.K. From Selenium to Selenoproteins: Synthesis, Identity, and Their Role in Human Health. Antioxidants Redox Signal. 2007, 9, 775–806. [Google Scholar] [CrossRef]
- Andoh, A.; Hirashima, M.; Maeda, H.; Hata, K.; Inatomi, O.; Tsujikawa, T.; Sasaki, M.; Takahashi, K.; Fujiyama, Y. Serum selenoprotein-P levels in patients with inflammatory bowel disease. Nutr. 2005, 21, 574–579. [Google Scholar] [CrossRef]
- Gao, Y.; Yuan, Y.J.; Zhou, X.J.; Wang, F.X.; Zhang, J. Preparation and characterization of polysaccharide selenate of tamarind seeds. Chin. J. Phar. Ana. 2012. [Google Scholar] [CrossRef]
- Wang, J.; Zhao, B.; Wang, X.; Yao, J.; Zhang, J. Synthesis of selenium-containing polysaccharides and evaluation of antioxidant activity in vitro. Int. J. Boil. Macromol. 2012, 51, 987–991. [Google Scholar] [CrossRef] [PubMed]
- Wen, Z.-S.; Ma, L.; Xiang, X.-W.; Tang, Z.; Guan, R.-F.; Qu, Y.-L. Protective effect of low molecular-weight seleno-aminopolysaccharides against H2O2-induecd oxidative stress in intestinal epithelial cells. Int. J. Boil. Macromol. 2018, 112, 745–753. [Google Scholar] [CrossRef] [PubMed]
- Gu, L.-X.; Wen, Z.-S.; Xiang, X.-W.; Ma, L.; Wang, X.-B.; Ma, J.-Y.; Qu, Y.-L. Immunomodulatory effect of low molecular-weight seleno-aminopolysaccharides in intestinal epithelial cells. Int. J. Boil. Macromol. 2017, 99, 570–577. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.B.; Wen, Z.S.; Zhang, P.; Qu, Y.L.; Xu, Z.R. Acute toxicity of low molecular seleno-aminopolysaccharide to mice. Chin. J. Vet. Sci. 2017, 249, 1790–1796. [Google Scholar]
- Pié, S.; Lallès, J.P.; Blazy, F.; Laffitte, J.; Sève, B.; Oswald, I.P. Weaning is associated with an upregulation of expression of inflamatory cytokines in the intestine of piglets. J. Nutr. 2004, 134, 641–647. [Google Scholar] [CrossRef]
- France, M.M.; Turner, J.R. The mucosal barrier at a glance. J. Cell Sci. 2017, 130, 307–314. [Google Scholar] [CrossRef]
- Johansson, M.E.; Larsson, J.M.; Hansson, G.C. The two mucus layers of colon are organized by the MUC2 mucin, whereas the outer layer is a legislator of host-microbial interactions. Proc. Natl. Acad. Sci. USA 2011, 108, 4659–4665. [Google Scholar] [CrossRef]
- Jakobsson, H.E.; Rodríguez-Piñeiro, A.M.; Schütte, A.; Ermund, A.; Boysen, P.; Bemark, M.; Sommer, F.; Bäckhed, F.; Hansson, G.C.; Johansson, M.E. The composition of the gut microbiota shapes the colon mucus barrier. EMBO Rep. 2015, 16, 164–177. [Google Scholar] [CrossRef]
- Lindén, S.K.; Sheng, Y.H.; Every, A.L.; Miles, K.M.; Skoog, E.C.; Florin, T.H.J.; Sutton, P.; McGuckin, M.A. MUC1 Limits Helicobacter pylori Infection both by Steric Hindrance and by Acting as a Releasable Decoy. PLoS Pathog. 2009, 5, e1000617. [Google Scholar] [CrossRef]
- Karlsson, N.G.; Herrmann, A.; Karlsson, H.E.; Johansson, M.; Carlstedt, I.; Hansson, G.C. The glycosylation of rat intestinal Muc2 mucin varies between rat strains and the small and large intestine. A study of O-linked oligosaccharides by a mass spectrometric approach. J. Boil. Chem. 1997, 272, 27025–27034. [Google Scholar] [CrossRef] [PubMed]
- Larsson, J.M.H.; Crespo, J.G.; Johansson, M.E.V.; Eklund, L.; Karlsson, H.; Sjövall, H.; Hansson, G.C. Altered O-glycosylation profile of MUC2 mucin occurs in active ulcerative colitis and is associated with increased inflammation. Inflamm. Bowel Dis. 2011, 17, 2299–2307. [Google Scholar] [CrossRef] [PubMed]
- Larsson, J.M.; Karlsson, H.; Sjövall, H.; Hansson, G.C. A complex, but uniform O-glycosylation of the human MUC2 mucin from colonic biopsies analyzed by nanoLC/MS. Glycobiology 2009, 19, 756–766. [Google Scholar] [CrossRef] [PubMed]
- Thomsson, K.; Holmén-Larsson, J.M.; Ångström, J.; Johansson, M.E.V.; Xia, L.; Hansson, G.C. Detailed O-glycomics of the Muc2 mucin from colon of wild-type, core 1- and core 3-transferase-deficient mice highlights differences compared with human MUC2. Glycobiology 2012, 22, 1128–1139. [Google Scholar] [CrossRef]
- Brockhausen, I. Sulphotransferases acting on mucin-type oligosaccharides. Biochem. Soc. Trans. 2003, 31, 318–325. [Google Scholar] [CrossRef]
- An, G.; Wei, B.; Xia, B.; McDaniel, J.M.; Ju, T.; Cummings, R.D.; Braun, J.; Xia, L. Increased susceptibility to colitis and colorectal tumors in mice lacking core 3–derived O-glycans. J. Exp. Med. 2007, 204, 1417–1429. [Google Scholar] [CrossRef]
- Sommer, F.; Adam, N.; Johansson, M.E.V.; Xia, L.; Hansson, G.C.; Bäckhed, F. Altered Mucus Glycosylation in Core 1 O-Glycan-Deficient Mice Affects Microbiota Composition and Intestinal Architecture. PLoS ONE 2014, 9, e85254. [Google Scholar] [CrossRef]
- Soong, G.; Muir, A.; Gomez, M.I.; Waks, J.; Reddy, B.; Planet, P.; Singh, P.K.; Kanetko, Y.; Wolfgang, M.C.; Hsiao, Y.-S.; et al. Bacterial neuraminidase facilitates mucosal infection by participating in biofilm production. J. Clin. Investig. 2006, 116, 2297–2305. [Google Scholar] [CrossRef]
- Venkatakrishnan, V.; Quintana-Hayashi, M.P.; Mahu, M.; Haesebrouck, F.; Pasmans, F.; Lindén, S.K. Brachyspira hyodysenteriae Infection Regulates Mucin Glycosylation Synthesis Inducing an Increased Expression of Core-2 O-Glycans in Porcine Colon. J. Proteome Res. 2017, 16, 1728–1742. [Google Scholar] [CrossRef]
- Tran, D.T.; Hagen, K.G.T. Mucin-type O-Glycosylation during Development*. J. Boil. Chem. 2013, 288, 6921–6929. [Google Scholar] [CrossRef]
- Kong, Y.; Joshi, H.J.; Schjoldager, K.T.; Madsen, T.D.; Gerken, T.A.; Vester-Christensen, M.B.; Wandall, H.H.; Bennett, E.P.; Levery, S.B.; Vakhrushev, S.Y.; et al. Probing polypeptide GalNAc-transferase isoform substrate specificities by in vitro analysis. Glycobiology 2015, 25, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Bennett, E.P.; Hassan, H.S.; Hollingsworth, M.A.; Clausen, H. A novel human UDP-N-acetyl-D-Galactosamine; poylypeptide N-acetylgalactosaminyltransferase, Ga1NAc-T7, with specificity for partial Ga1NAc-glycosylated acceptor substrates. FEBS Lett. 1999, 460, 226–230. [Google Scholar] [CrossRef] [Green Version]
- Di Meo, S.; Reed, T.T.; Venditti, P.; Victor, V.M. Role of ROS and RNS Sources in Physiological and Pathological Conditions. Oxidative Med. Cell. Longev. 2016, 2016, 1–44. [Google Scholar] [CrossRef] [PubMed]
- Cai, B.; Wan, P.; Chen, H.; Chen, D.; Chen, X.; Sun, H.; Pan, J. Composition characterization of oyster polysaccharides from Crassostrea hongkongensis and their protective effect against H2O2-induced oxidative damage in IEC-6 cells. Int. J. Biol. Macromol. 2019, 124, 246–254. [Google Scholar] [CrossRef]
- Aviello, G.; Knaus, U.G. ROS in gastrointestinal inflammation: Rescue or Sabotage. Br. J. Pharmacol. 2017, 174, 1704–1718. [Google Scholar] [CrossRef] [Green Version]
- Birben, E.; Sahiner, U.M.; Sackesen, C.; Erzurum, S.; Kalayci, O. Oxidative Stress and Antioxidant Defense. World Allergy Organ. J. 2012, 5, 9–19. [Google Scholar] [CrossRef] [Green Version]
- Hayes, J.D.; McMahon, M. NRF2 and KEAP1 mutations: Permanent activation of an adaptive response in cancer. Trends Biochem. Sci. 2009, 34, 176–188. [Google Scholar] [CrossRef]
- Vachharajani, T.J.; Work, J.; Issekutz, A.C.; Granger, D.N. Heme oxygenase modulates selectin expression in different regional vascular beds. Am. J. Physiol. Circ. Physiol. 2000, 278, H1613–H1617. [Google Scholar] [CrossRef]
- Reszka, E.; Wieczorek, E.; Jablonska, E.; Janasik, B.; Fendler, W.; Wasowicz, W. Association between plasma selenium level and Nrf2 target genes expression in humans. J. Trace Elem. Med. Biol. 2014, 30, 102–106. [Google Scholar] [CrossRef]
- Schulz, B.L.; Packer, N.; Karlsson, N.G. Small-scale analysis of O-linked oligosaccharides from glycoproteins and mucins separated by gel electrophoresis. Anal. Chem. 2002, 174, 6088–6097. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | VH (μm) | CD (μm) | VH/CD |
|---|---|---|---|
| Lactated | 464.29 ± 14.20 a | 124.56 ± 12.30 a | 3.72 ± 1.24 a |
| Weaned | 358.25 ± 24.10 b | 109.64 ± 8.46 a | 3.26 ± 0.89 b |
| LSA | 421.54 ± 18.70 a | 113.57 ± 5.68 a | 3.76 ± 0.95 a |
| Gene | Gene Accession Number | Primer Sequence 5′-3′ | PCR Product Size (bp) | Tm |
|---|---|---|---|---|
| SOD | NM_017050.1 | F: TCTAAGAAACATGGCGGTCC R: CAGTTAGCAGGCCAGCAGAT | 312 | 60 |
| GSH-Px | nm_030826.4 | F: CTCTCCGCGGTGGCACAGT R: CCACCACCGGGTCGGACATAC | 290 | 64 |
| Nrf2 | XM_006234398.3 | F: GCTGCCATTAGTCAGTCGCTCTC R: ACCGTGCCTTCAGTGTGCTTC | 104 | 63 |
| HO-1 | NM_012580.2 | F: CGTGCTCGCATGAACACTCT R: GGCGGTCTTAGCCTCTTCTGT | 72 | 61 |
| GalNT1 | XM_006254471.3 | F: ACGACAAGCGTGGTGATT R: CTCGCAGTGAGCATCTAA | 291 | 60 |
| GalNT2 | NM_001106196.1 | F: TGCCAAGCAACAACAAGA R: TTAGCAGCGGACATCGTG | 183 | 60 |
| GalNT7 | XM_017600074.1 | F: CAGAGCCCACTGAGCAGGAT R: TGAATCTGTCATCTCCAGGAGCTA | 97 | 63 |
| β-actin | NM_031144.3 | F: AGTGTGACGTTGACATCCGTA R: GCCAGAGCAGTAATCTCCTTCT | 112 | 59 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wen, Z.-S.; Du, M.; Tang, Z.; Zhou, T.-Y.; Zhang, Z.-S.; Song, H.-H.; Xiang, X.-W.; Han, X.-Y. Low Molecular Seleno-Aminopolysaccharides Protect the Intestinal Mucosal Barrier of Rats under Weaning Stress. Int. J. Mol. Sci. 2019, 20, 5727. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20225727
Wen Z-S, Du M, Tang Z, Zhou T-Y, Zhang Z-S, Song H-H, Xiang X-W, Han X-Y. Low Molecular Seleno-Aminopolysaccharides Protect the Intestinal Mucosal Barrier of Rats under Weaning Stress. International Journal of Molecular Sciences. 2019; 20(22):5727. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20225727
Chicago/Turabian StyleWen, Zheng-Shun, Ming Du, Zhen Tang, Tian-Yi Zhou, Zhong-Shan Zhang, Hou-Hui Song, Xing-Wei Xiang, and Xin-Yan Han. 2019. "Low Molecular Seleno-Aminopolysaccharides Protect the Intestinal Mucosal Barrier of Rats under Weaning Stress" International Journal of Molecular Sciences 20, no. 22: 5727. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20225727