Gene Losses and Variations in Chloroplast Genome of Parasitic Plant Macrosolen and Phylogenetic Relationships within Santalales

 , ,
, ,
Abstract
:1. Introduction
2. Results
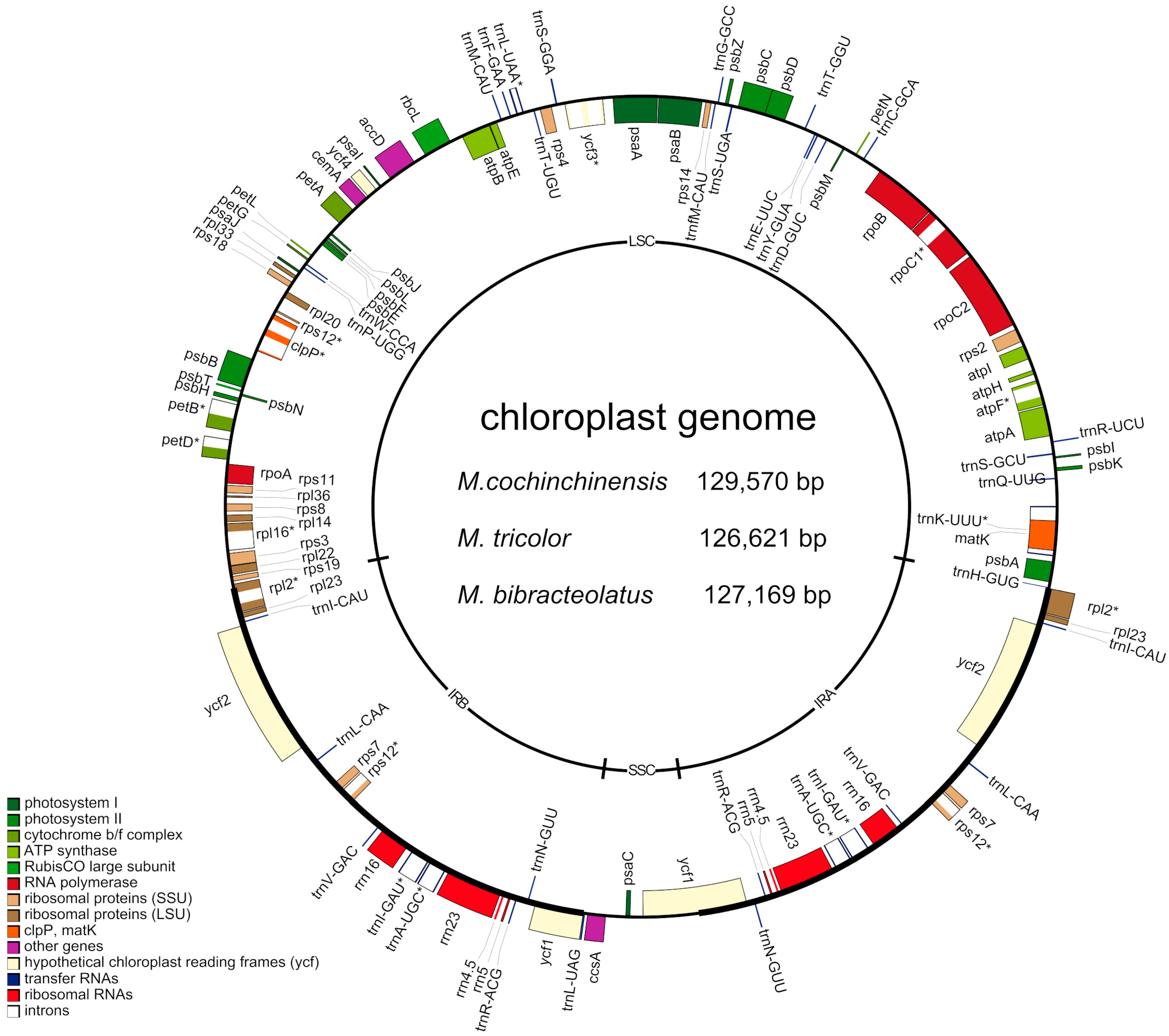
2.1. Complete Chloroplast Genomes of Three Macrosolen Species
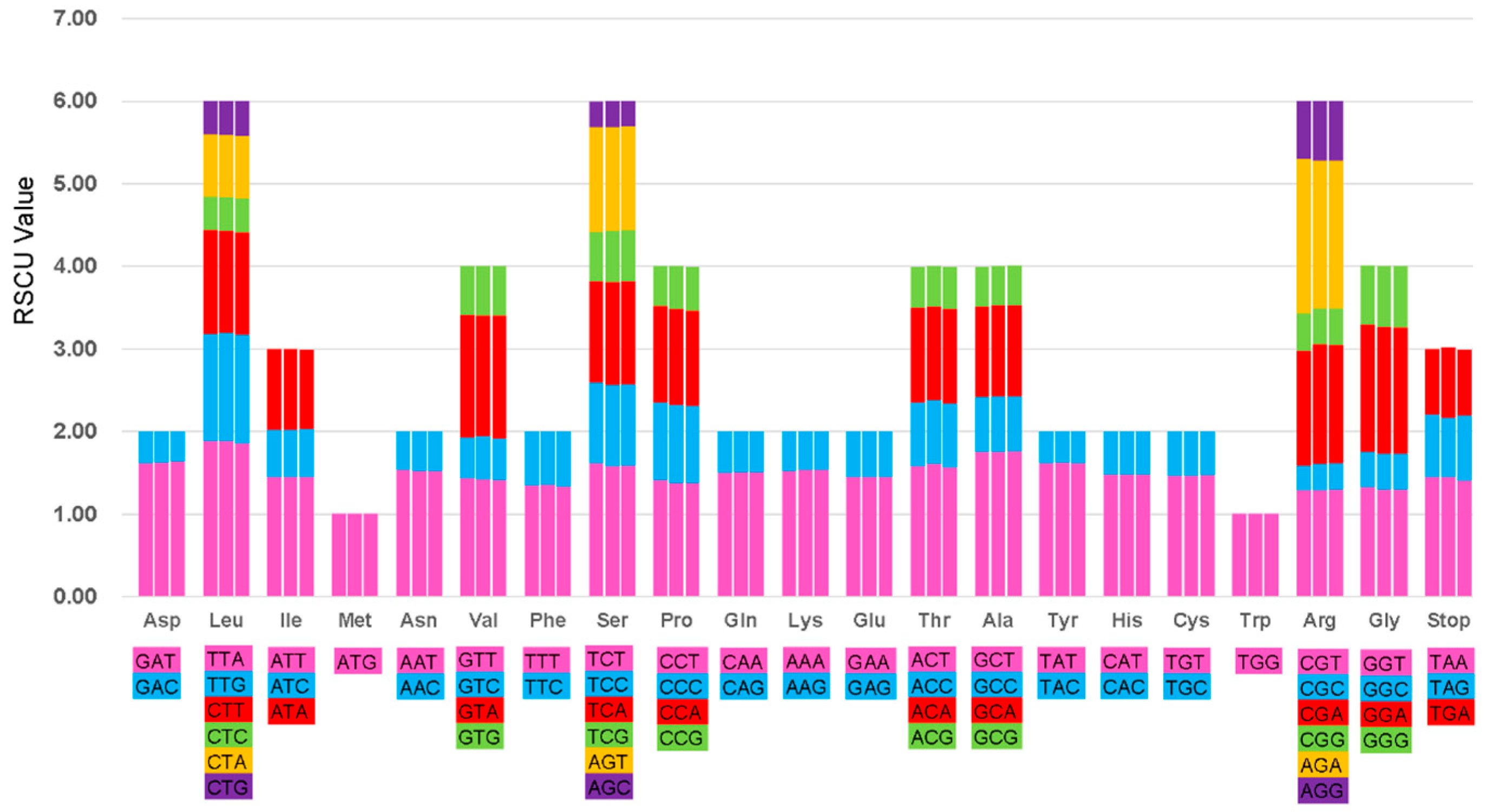
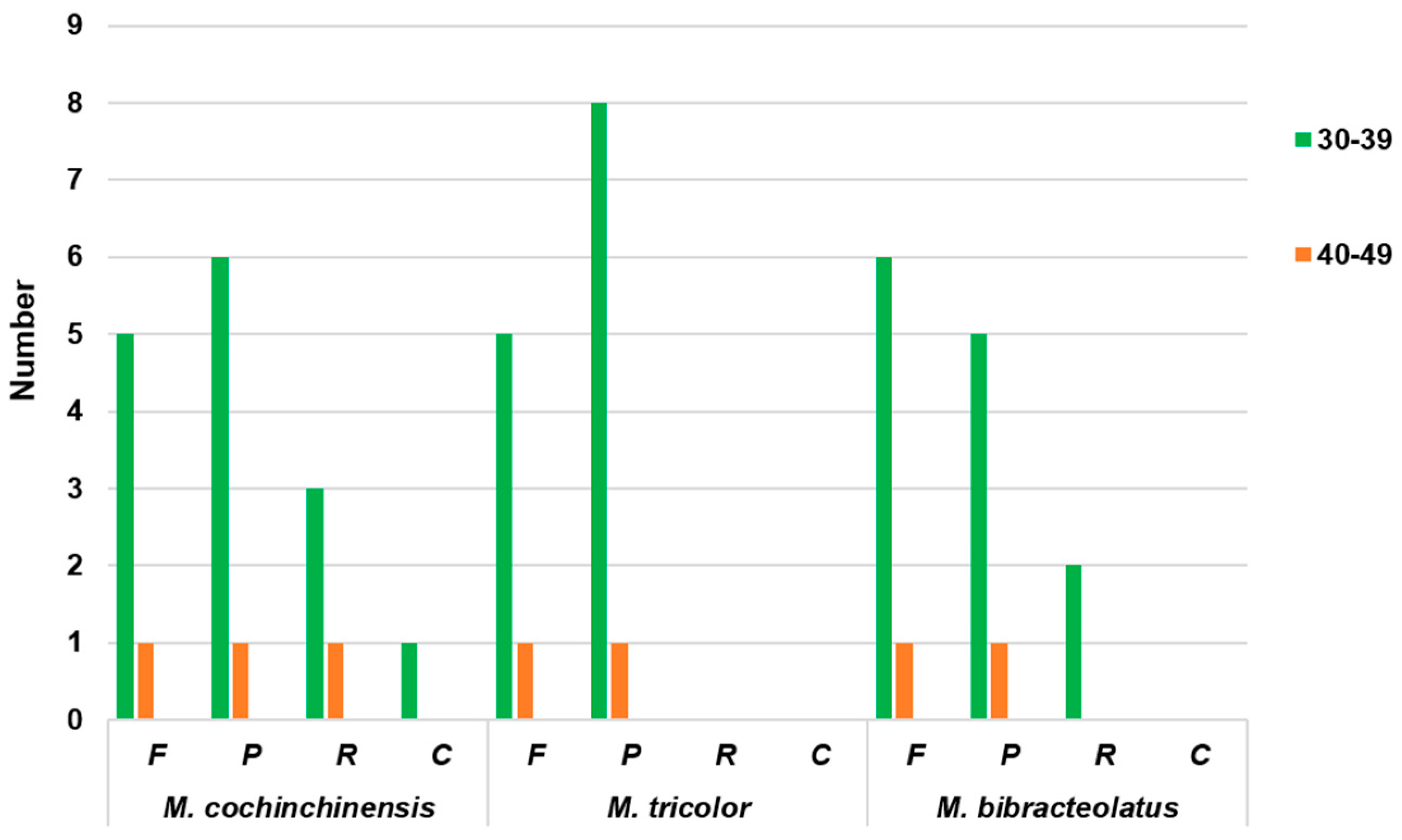
2.2. Codon Usage Analyses and RNA Editing Sites
2.3. IR Constriction and Expansion
2.4. Simple Sequence Repeats (SSRs) and Repeat Structure Analyses
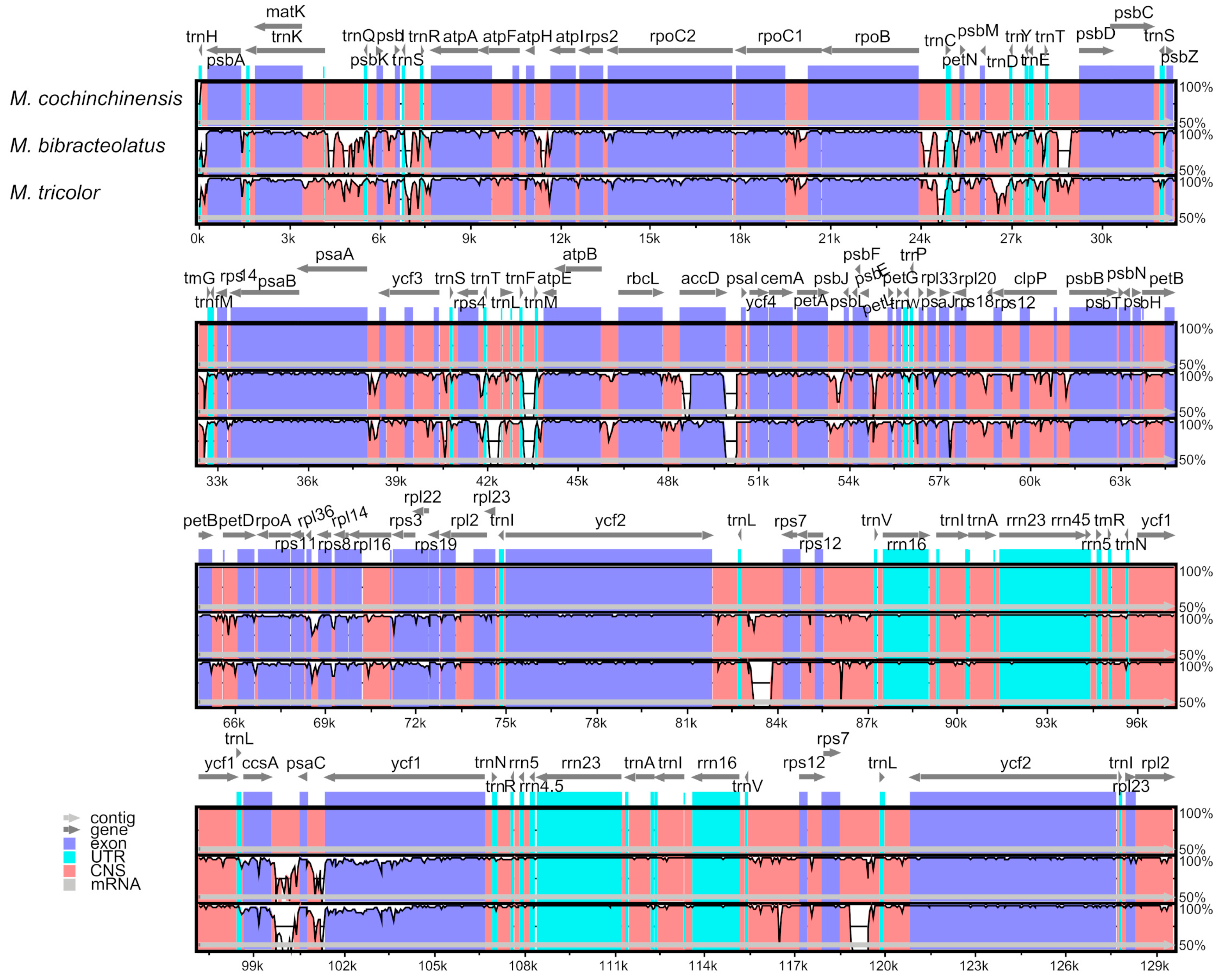
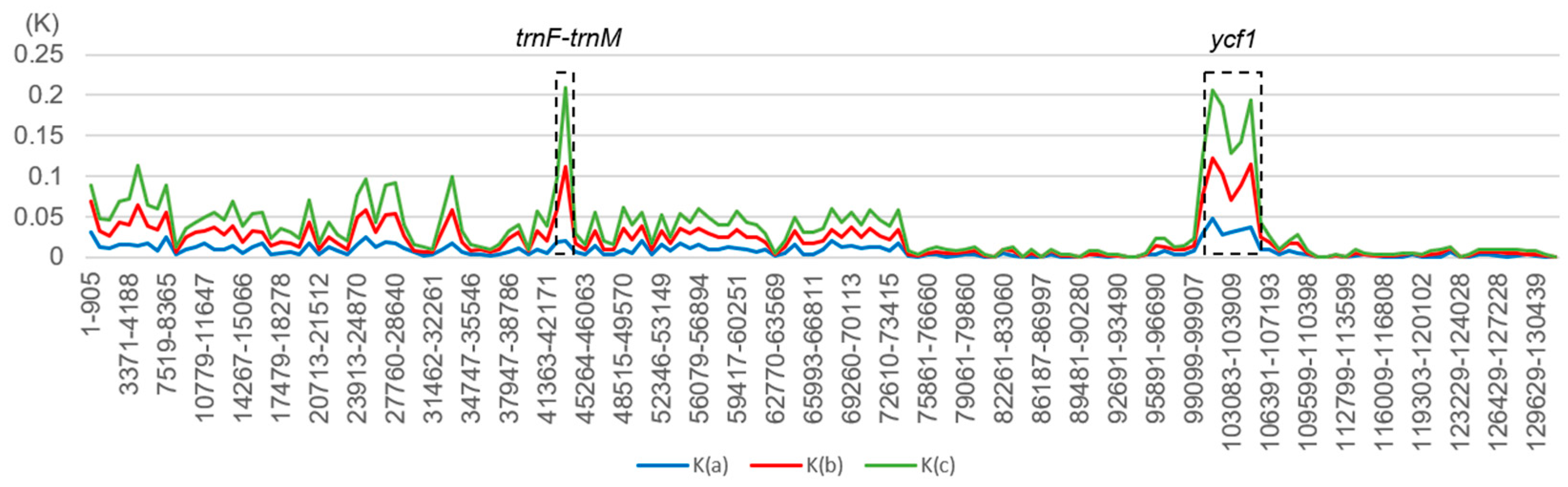
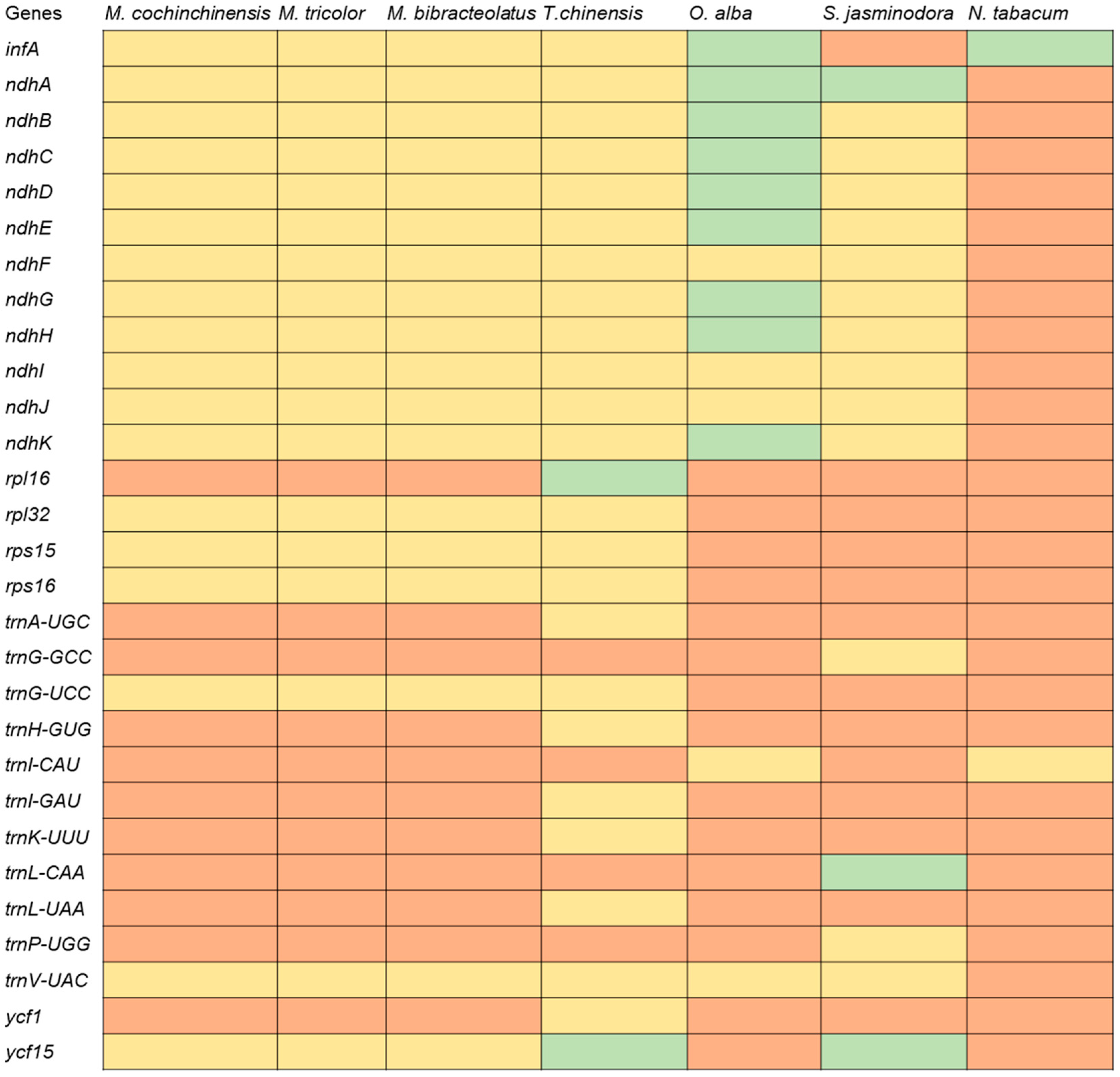
2.5. Comparative Genomic Analyses
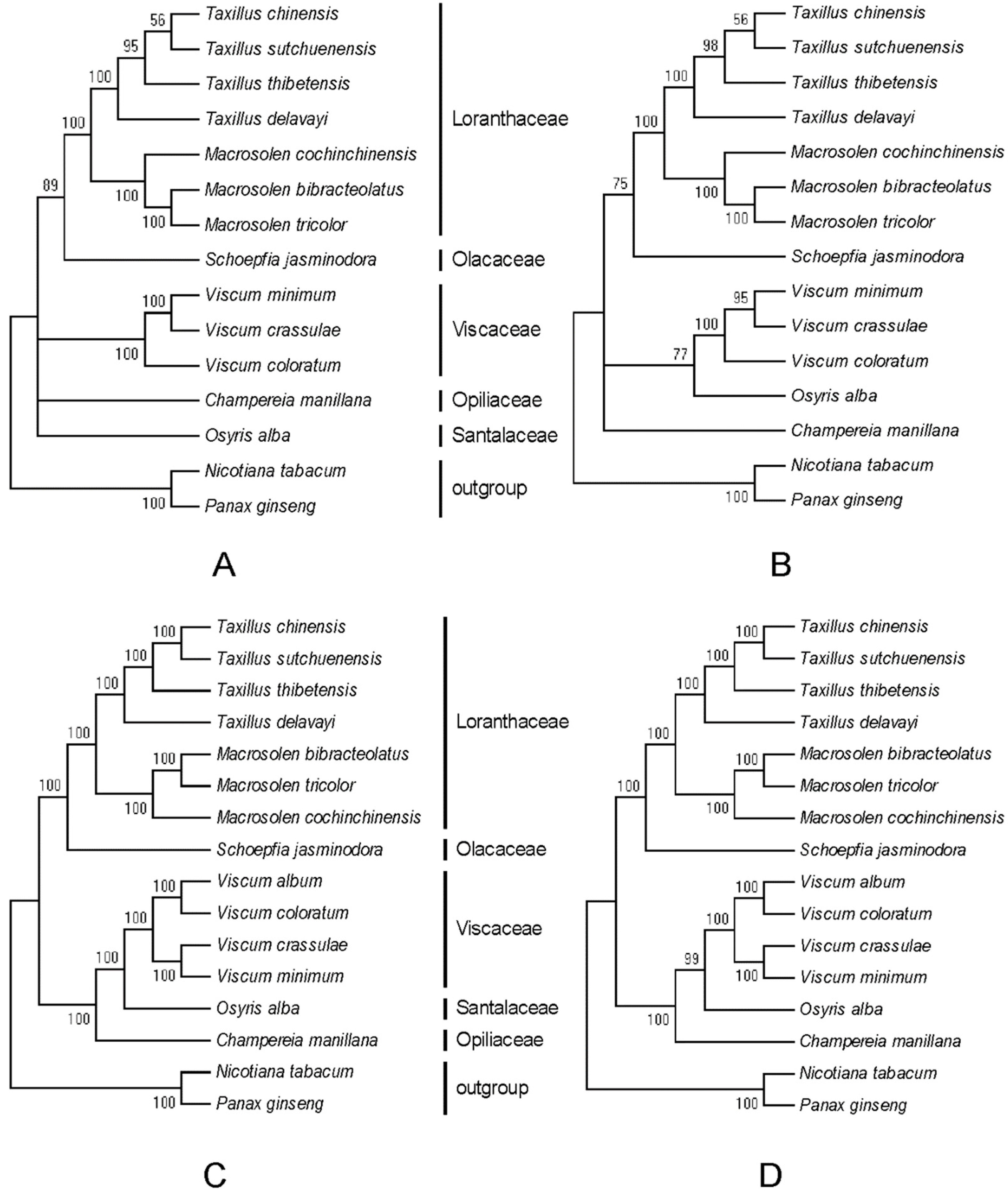
2.6. Phylogenetic Analyses
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. DNA Extraction, Sequencing and Assembly
4.3. Genome Comparison and Phylogenetic Analyses
4.4. Other Analyses
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| LSC | Large single copy |
| SSC | Small single copy |
| IR | Inverted repeat |
| MP | Maximum parsimony |
| ML | Maximum likelihood |
| RSCU | Relative synonymous codon usage |
| SSR | Simple Sequence Repeats |
| NCBI | National Center for Biotechnology Information |
References
- Wicke, S.; Naumann, J. Molecular evolution of plastid genomes in parasitic flowering plants. Adv. Bot. Res. 2018, 85, 315–347. [Google Scholar]
- Wang, L.; Dong, W.; Zhou, S. Structural mutations and reorganizations in chloroplast genomes of flowering plants. Acta Bot. Boreali Occident. Sin. 2012, 32, 1282–1288. [Google Scholar]
- Huang, X.; Guan, K.; Li, A. Biological traits and their ecological significances of parasitic plants: A review. Chin. J. Ecol. 2011, 30, 1838–1844. [Google Scholar]
- Wolfe, K.H.; Morden, C.W.; Palmer, J.D. Function and evolution of a minimal plastid genome from a nonphotosynthetic parasitic plant. Proc. Natl. Acad. Sci. USA 1992, 89, 10648–10652. [Google Scholar] [CrossRef]
- Petersen, G.; Cuenca, A.; Seberg, O. Plastome evolution in hemiparasitic mistletoes. Genome Biol. Evol. 2015, 7, 2520–2532. [Google Scholar] [CrossRef]
- Su, H.; Hu, J. The complete chloroplast genome of hemiparasitic flowering plant Schoepfia jasminodora. Mitochondrial DNA Part B 2016, 1, 767–769. [Google Scholar] [CrossRef]
- Li, Y.; Zhou, J.G.; Chen, X.L.; Cui, Y.X.; Xu, Z.C.; Li, Y.H.; Song, J.Y.; Duan, B.Z.; Yao, H. Gene losses and partial deletion of small single-copy regions of the chloroplast genomes of two hemiparasitic Taxillus species. Sci. Rep. 2017, 7, 12834. [Google Scholar] [CrossRef]
- Molina, J.; Hazzouri, K.M.; Nickrent, D.; Geisler, M.; Meyer, R.S.; Pentony, M.M.; Flowers, J.M.; Pelser, P.; Barcelona, J.; Inovejas, S.A.; et al. Possible loss of the chloroplast genome in the parasitic flowering plant Rafflesia lagascae (Rafflesiaceae). Mol. Biol. Evol. 2014, 31, 793–803. [Google Scholar] [CrossRef]
- Li, X.; Zhang, T.C.; Qiao, Q.; Ren, Z.; Zhao, J.; Yonezawa, T.; Hasegawa, M.; Crabbe, M.J.; Li, J.; Zhong, Y. Complete chloroplast genome sequence of holoparasite Cistanche deserticola (Orobanchaceae) reveals gene loss and horizontal gene transfer from its host Haloxylon ammodendron (Chenopodiaceae). PLoS ONE 2013, 8, e58747. [Google Scholar] [CrossRef]
- Frailey, D.C.; Chaluvadi, S.R.; Vaughn, J.N.; Coatney, C.G.; Bennetzen, J.L. Gene loss and genome rearrangement in the plastids of five Hemiparasites in the family Orobanchaceae. BMC Plant Biol. 2018, 18, 30. [Google Scholar] [CrossRef]
- Samigullin, T.H.; Logacheva, M.D.; Penin, A.A.; Vallejo-Roman, C.M. Complete plastid genome of the recent holoparasite Lathraea squamaria reveals earliest stages of plastome reduction in orobanchaceae. PLoS ONE 2016, 11, e0150718. [Google Scholar] [CrossRef] [PubMed]
- Su, H.J.; Barkman, T.J.; Hao, W.; Jones, S.S.; Naumann, J.; Skippington, E.; Wafula, E.K.; Hu, J.M.; Palmer, J.D.; de Pamphilis, C.W. Novel genetic code and record-setting AT-richness in the highly reduced plastid genome of the holoparasitic plant Balanophora. Proc. Natl. Acad. Sci. USA 2019, 116, 934–943. [Google Scholar] [CrossRef] [PubMed]
- Bellot, S.; Renner, S.S. The plastomes of two species in the endoparasite genus pilostyles (Apodanthaceae) each retain just five or six possibly functional genes. Genome Biol. Evol. 2016, 8, 189–201. [Google Scholar] [CrossRef] [PubMed]
- Petersen, G.; Zervas, A.; Pedersen, H.; Seberg, O. Contracted genes and dwarfed plastome in mycoheterotrophic Sciaphila thaidanica (Triuridaceae, Pandanales). Genome Biol. Evol. 2018, 10, 976–981. [Google Scholar] [CrossRef] [PubMed]
- Schelkunov, M.I.; Nuraliev, M.S.; Logacheva, M.D. Rhopalocnemis phalloides has one of the most reduced and mutated plastid genomes known. Peer J. 2019, 7, e7500. [Google Scholar] [CrossRef]
- Graham, S.W.; Lam, V.K.; Merckx, V.S. Plastomes on the edge: The evolutionary breakdown of mycoheterotroph plastid genomes. New Phytol. 2017, 214, 48. [Google Scholar] [CrossRef]
- Lallemand, F.; Logacheva, M.; Le Clainche, I.; Berard, A.; Zheleznaia, E.; May, M.; Jakalski, M.; Delannoy, E.; Le Paslier, M.C.; Selosse, M.A. Thirteen new plastid genomes from mixotrophic and autotrophic species provide insights into heterotrophy evolution in Neottieae orchids. Genome Biol. Evol. 2019, 11, 2457–2467. [Google Scholar] [CrossRef]
- Clegg, M.T.; Gaut, B.S.; Learn, G.H., Jr.; Morton, B.R. Rates and patterns of chloroplast DNA evolution. Proc. Natl. Acad. Sci. USA 1994, 91, 6795–6801. [Google Scholar] [CrossRef]
- Dyall, S.D.; Brown, M.T.; Johnson, P.J. Ancient invasions: From endosymbionts to organelles. Science 2004, 304, 253–257. [Google Scholar] [CrossRef]
- Palmer, J.D. Comparative organization of chloroplast genomes. Annu. Rev. Genet. 1985, 19, 325–354. [Google Scholar] [CrossRef]
- Wicke, S.; Schneeweiss, G.M.; dePamphilis, C.W.; Müller, K.F.; Quandt, D. The evolution of the plastid chromosome in land plants: Gene content, gene order, gene function. Plant Mol. Biol. 2011, 76, 273–297. [Google Scholar] [CrossRef] [PubMed]
- Powell, W.; Morgante, M.; Andre, C.; Hanafey, M.; Vogel, J.; Tingey, S.; Rafalski, A. The comparison of RFLP, RAPD, AFLP and SSR (microsatellite) markers for germplasm analysis. Mol. Breed. 1996, 2, 225–238. [Google Scholar] [CrossRef]
- Niu, Z.; Zhu, S.; Pan, J.; Li, L.; Sun, J.; Ding, X. Comparative analysis of Dendrobium plastomes and utility of plastomic mutational hotspots. Sci. Rep. 2017, 7, 2073. [Google Scholar] [CrossRef]
- Zhou, Y.; Nie, J.; Xiao, L.; Hu, Z.; Wang, B. Comparative chloroplast genome analysis of rhubarb botanical origins and the development of specific identification markers. Molecules 2018, 23, 2811. [Google Scholar] [CrossRef]
- Chen, X.; Zhou, J.; Cui, Y.; Wang, Y.; Duan, B.; Yao, H. Identification of Ligularia herbs using the complete chloroplast genome as a super-barcode. Front. Pharmacol. 2018, 9, 695. [Google Scholar] [CrossRef]
- Niu, Z.; Pan, J.; Xue, Q.; Zhu, S.; Liu, W.; Ding, X. Plastome-wide comparison reveals new SNV resources for the authentication of Dendrobium huoshanense and its corresponding medicinal slice (Huoshan Fengdou). Acta Pharm. Sin. B 2018, 8, 466–477. [Google Scholar] [CrossRef]
- Wu, M.; Li, Q.; Hu, Z.; Li, X.; Chen, S. The complete Amomum kravanh chloroplast genome sequence and phylogenetic analysis of the commelinids. Molecules 2017, 22, 1875. [Google Scholar] [CrossRef]
- Flora Reipublicae Popularis Sinicae (FRPS). Available online: http://www.iplant.cn/ (accessed on 28 October 2019).
- Gong, Z.; Wang, Z.; Xu, L.; Xu, G.; Wu, J. Studies on medicinal plants of Loranthaceae in China. Chin. Wild Plant Resour. 1996, 1, 11–15. [Google Scholar]
- Zhao, Q.; Xu, Q.; Zhang, H. Pharmacognostic identification on crude drug of Macrosolen cochinchinensis. Chin. J. Ethnomed. Ethnopharm. 1998, 34, 1–3. [Google Scholar]
- Li, Y.; Lu, D.; Zhao, M.; Zhu, K. Research on the developments and applications for medicinal plants of Loranthaceae in Guangxi. Guangxi Med. J. 2006, 28, 1695–1698. [Google Scholar]
- Sharp, P.M.; Li, W.H. The codon Adaptation Index--a measure of directional synonymous codon usage bias, and its potential applications. Nucleic Acids Res. 1987, 15, 1281–1295. [Google Scholar] [CrossRef] [PubMed]
- Powell, W.; Morgante, M.; Mcdevitt, R.; Vendramin, G.G.; Rafalski, J.A. Polymorphic simple sequence repeat regions in chloroplast genomes: Applications to the population genetics of pines. Proc. Natl. Acad. Sci. USA 1995, 92, 7759–7763. [Google Scholar] [CrossRef] [PubMed]
- Akkaya, S.M. Length polymorphism of simple sequence repeat DNA in soybean. Genetics 1992, 132, 1131–1139. [Google Scholar] [PubMed]
- Rozas, J.; Ferrer-Mata, A.; Sanchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sanchez-Gracia, A. DnaSP 6: DNA Sequence Polymorphism Analysis of Large Data Sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef] [PubMed]
- Shin, H.W.; Lee, N.S. Understanding plastome evolution in Hemiparasitic Santalales: Complete chloroplast genomes of three species, Dendrotrophe varians, Helixanthera parasitica, and Macrosolen cochinchinensis. PLoS ONE 2018, 13, e0200293. [Google Scholar] [CrossRef]
- Kim, H.T.; Kim, J.S.; Moore, M.J.; Neubig, K.M.; Williams, N.H.; Whitten, W.M.; Kim, J.H. Seven new complete plastome sequences reveal rampant independent loss of the ndh gene family across orchids and associated instability of the inverted repeat/small single-copy region boundaries. PLoS ONE 2015, 10, e0142215. [Google Scholar] [CrossRef]
- Mcneal, J.R.; Kuehl, J.V.; Boore, J.L.; Pamphilis, C.W.D. Complete plastid genome sequences suggest strong selection for retention of photosynthetic genes in the parasitic plant genus Cuscuta. BMC Plant Biol. 2007, 7, 57. [Google Scholar] [CrossRef] [Green Version]
- Der, J.P.; Nickrent, D.L. A molecular phylogeny of Santalaceae (Santalales). Syst. Bot. 2008, 33, 107–116. [Google Scholar] [CrossRef]
- Zhou, J.; Chen, X.; Cui, Y.; Sun, W.; Li, Y.; Wang, Y.; Song, J.; Yao, H. Molecular structure and phylogenetic analyses of complete chloroplast genomes of two Aristolochia medicinal species. Int. J. Mol. Sci. 2017, 18, 1839. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Frazer, K.A.; Lior, P.; Alexander, P.; Rubin, E.M.; Inna, D. VISTA: Computational tools for comparative genomics. Nucleic Acids Res. 2004, 32, W273. [Google Scholar] [CrossRef] [PubMed]
- CPGAVAS2. Available online: http://47.96.249.172:16018/analyzer/extractSeq (accessed on 30 July 2019).
- Mower, J.P. The PREP suite: Predictive RNA editors for plant mitochondrial genes, chloroplast genes and user-defined alignments. Nucleic Acids Res. 2009, 37, 253–259. [Google Scholar] [CrossRef] [PubMed]
- Misa-Microsatellite Identification Tool. Available online: Pgrc.ipk-gatersleben.de/misa/ (accessed on 2 June 2017).
- Kurtz, S.; Choudhuri, J.V.; Ohlebusch, E.; Schleiermacher, C.; Stoye, J.; Giegerich, R. REPuter: The manifold applications of repeat analysis on a genomic scale. Nucleic Acids Res. 2001, 29, 4633–4642. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | M. cochinchinensis | M. tricolor | M. bibracteolatus |
|---|---|---|---|
| Accession No. | MH161424 | MH161425 | MH161423 |
| Genome size (bp) | 129,570 | 126,621 | 127,169 |
| LSC length (bp) | 73,052 | 71,895 | 70,692 |
| SSC length (bp) | 5724 | 5320 | 5587 |
| IRs length (bp) | 25,397 | 24,703 | 25,445 |
| GC content (%) | 37.3 | 37.7 | 37.9 |
| Number of genes | 113 | 113 | 113 |
| Number of protein-coding genes | 68 | 68 | 68 |
| Number of tRNAs | 35 | 35 | 35 |
| Number of rRNAs | 8 | 8 | 8 |
| Number of pseudogenes | 2 | 2 | 2 |
| No. | Group of Genes | Gene Names | Number |
|---|---|---|---|
| 1 | Photosystem I | psaA, psaB, psaC, psaI, psaJ | 5 |
| 2 | Photosystem II | psbA, psbB, psbC, psbD, psbE, psbF, psbH, psbI, psbJ, psbK, psbL, psbM, psbN, psbT, psbZ | 15 |
| 3 | Cytochrome b/f complex | petA, petB *, petD *, petG, petL, petN | 6 |
| 4 | ATP synthase | atpA, atpB, atpE, atpF *, atpH, atpI | 6 |
| 5 | NADH dehydrogenase | - | 0 |
| 6 | RubisCO large subuni | rbcL | 1 |
| 7 | RNA polymerase | rpoA, rpoB, rpoC1 *, rpoC2 | 4 |
| 8 | Ribosomal proteins (SSU) | rps2, rps3, rps4, rps7 (×2), rps8, rps11, rps12 ** (×2), rps14, rps18, rps19 | 12(2) |
| 9 | Ribosomal proteins (LSU) | rpl2 * (×2), rpl14, rpl16 *, rpl20, rpl22, rpl23 (×2), rpl33, rpl36 | 10(2) |
| 10 | Proteins of unknown function | ycf1(×2), ycf2(×2), ycf3 **, ycf4 | 6(2) |
| 11 | Transfer RNAs | 35 tRNAs (4 contain an intron, 7 in the IRs) | 35(7) |
| 12 | Ribosomal RNAs | rrn4.5 (×2), rrn5(×2), rrn16 (×2), rrn23 (×2) | 8(4) |
| 13 | Other genes | accD, clpP **, matK, ccsA, cemA | 5 |
| SSR Type | Repeat Unit | Amount | Ratio (%) | ||||
|---|---|---|---|---|---|---|---|
| ① | ② | ③ | ① | ② | ③ | ||
| mono | A/T | 161 | 159 | 153 | 95.3 | 95.8 | 94.4 |
| C/G | 8 | 7 | 9 | 4.7 | 4.2 | 5.6 | |
| di | AC/GT | 5 | 4 | 4 | 9.6 | 8.5 | 9.3 |
| AG/CT | 13 | 14 | 13 | 25 | 29.8 | 30.2 | |
| AT/TA | 34 | 29 | 26 | 64.4 | 61.7 | 60.5 | |
| tri | AAT/ATT | 4 | 4 | 0 | 66.7 | 66.7 | 0 |
| ATC/ATG | 2 | 2 | 2 | 33.3 | 33.3 | 100 | |
| tetra | AAAG/CTTT | 3 | 3 | 3 | 33.3 | 42.9 | 30 |
| AATC/ATTG | 1 | 1 | 0 | 11.1 | 14.3 | 0 | |
| ACAG/CTGT | 1 | 1 | 1 | 11.1 | 14.3 | 10 | |
| AAAT/ATTT | 3 | 1 | 3 | 33.3 | 14.3 | 30 | |
| AATG/ATTC | 1 | 0 | 1 | 11.1 | 0 | 10 | |
| AGAT/ATCT | 0 | 1 | 1 | 0 | 14.3 | 10 | |
| ACAT/ATGT | 0 | 0 | 1 | 0 | 0 | 10 | |
| penta | AATAT/ATATT | 1 | 0 | 0 | 100 | 0 | 0 |
| hexa | ATATCC/ATATGG | 1 | 0 | 0 | 100 | 0 | 0 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nie, L.; Cui, Y.; Wu, L.; Zhou, J.; Xu, Z.; Li, Y.; Li, X.; Wang, Y.; Yao, H. Gene Losses and Variations in Chloroplast Genome of Parasitic Plant Macrosolen and Phylogenetic Relationships within Santalales. Int. J. Mol. Sci. 2019, 20, 5812. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20225812
Nie L, Cui Y, Wu L, Zhou J, Xu Z, Li Y, Li X, Wang Y, Yao H. Gene Losses and Variations in Chloroplast Genome of Parasitic Plant Macrosolen and Phylogenetic Relationships within Santalales. International Journal of Molecular Sciences. 2019; 20(22):5812. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20225812
Chicago/Turabian StyleNie, Liping, Yingxian Cui, Liwei Wu, Jianguo Zhou, Zhichao Xu, Yonghua Li, Xiwen Li, Yu Wang, and Hui Yao. 2019. "Gene Losses and Variations in Chloroplast Genome of Parasitic Plant Macrosolen and Phylogenetic Relationships within Santalales" International Journal of Molecular Sciences 20, no. 22: 5812. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20225812