Trypanosoma brucei J-Protein 2 Functionally Co-Operates with the Cytosolic Hsp70 and Hsp70.4 Proteins

Biotechnology Innovation Centre, Rhodes University, Grahamstown 6140, South Africa
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2019, 20(23), 5843; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20235843
Submission received: 8 October 2019
/
Revised: 5 November 2019
/
Accepted: 7 November 2019
/
Published: 21 November 2019
(This article belongs to the Special Issue The Role of Heat Shock Proteins: From Molecular Mechanisms to Therapeutics)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:The etiological agent of African trypanosomiasis, Trypanosoma brucei (Tb), has been identified to possess an expanded and diverse group of heat shock proteins, which have been implicated in cytoprotection, differentiation, and subsequently progression and transmission of the disease. Heat shock protein 70 (Hsp70) is a highly conserved and ubiquitous molecular chaperone that is important in maintaining protein homeostasis in the cell. Its function is regulated by a wide range of co-chaperones, and inhibition of these functions and interactions with co-chaperones are emerging as potential therapeutic targets for numerous diseases. This study sought to biochemically characterize the cytosolic TbHsp70 and TbHsp70.4 proteins and to investigate if they functionally co-operate with the Type I J-protein, Tbj2. Expression of TbHsp70 was shown to be heat inducible, while TbHsp70.4 was constitutively expressed. The basal ATPase activities of TbHsp70.4 and TbHsp70 were stimulated by Tbj2. It was further determined that Tbj2 functionally co-operated with TbHsp70 and TbHsp70.4 as the J-protein was shown to stimulate the ability of both proteins to mediate the refolding of chemically denatured β-galactosidase. This study provides further insight into this important class of proteins, which may contribute to the development of new therapeutic strategies to combat African Trypanosomiasis.
1. Introduction
African trypanosomiasis, a neglected tropical disease, afflicts humans as well as domestic and wild animals, and has a detrimental impact on the socioeconomic development in sub-Saharan Africa [1]. There is an urgent need for the development of more effective and safer chemotherapies to treat the disease, due to existing drug toxicity, growing parasite resistance, and the lack of a vaccine [2]. The Hsp70/J-protein chaperone machinery has been found to play an integral role in the development, differentiation, and survival of protozoan parasites as they transition through the various stages of their life cycle [3]. In Trypanosoma cruzi and several Leishmania spp., heat shock proteins have been shown to play an essential role in stress-induced stage differentiation and are important for disease progression and transmission [4,5], making this protein family an attractive chemotherapeutic target. The completion of the Trypanosoma brucei (T. brucei) genome has expedited transcriptome and proteome analyses and revealed that the extracellular parasite has an expanded and diverse Hsp70 and J-protein complement, with the parasite possessing cytosolic Hsp70 members that display atypical Hsp70 features [6].
The Hsp70 protein family is ubiquitous and plays an integral role in protein quality control and maintaining protein homeostasis under normal and stressful conditions [7,8]. Evolution has given rise to multiple homologous Hsp70 genes, with Hsp70 members found in all the major subcellular compartments within the cell [9,10]. The cytosol of eukaryotic cells has been shown to possess two major Hsp70 isoforms: stress-inducible Hsp70 and constitutively expressed Hsc70 [9]. The structure of eukaryotic cytosolic Hsp70s are highly conserved and are typically comprised of an N-terminal nucleotide binding domain and a C-terminal domain with a substrate binding domain (SBD) and a 10 kDa α-helical domain with a conserved EEVD motif [11,12].
Hsp70 functions as either an ATP-dependent refoldase, which entails folding nascent and unfolded polypeptides into their respective native states, or displays ATP-independent holdase activity, which involves binding to unfolded polypeptide aggregates and retrieving them back into solution [13,14]. Although Hsp70 chaperones can interact with a vast array of client proteins, their substrate specificity and activity are often driven by a cohort of proteins known as co-chaperones [7,14], which include nucleotide exchange factors (NEFs), tetratricopeptide repeat (TPR) domain containing proteins, and the Hsp40 family, otherwise known as the J proteins [14,15]. The J-protein family is one of the most important classes of Hsp70 co-chaperones, and all contain a highly conserved J-domain [16]. Members of this family are further classified into four types, I–IV [17,18]. Type I J-proteins are comprised of an N-terminal J-domain with a conserved HPD motif, a glycine-phenylalanine rich region, four zinc finger motifs (zinc finger domain), and a C-terminal peptide binding domain [17]. Type II J-proteins have a glycine-methionine rich region instead of the zinc finger domain [16]. Both type I and type II J-proteins serve to bind substrate polypeptides and target them to Hsp70 for refolding [16,17]. Type III J-proteins only possess the J-domain, however, it is not necessarily located at the N-terminal [17]. Type IV J-proteins also possess the J-domain, but the HPD motif is non-conserved or absent [18].
A recent in silico investigation revealed that the T. brucei genome was found to encode 12 members of the Hsp70 superfamily, with 8 members from the Hsp70/HSPA family and 4 Hsp110/HSPH family members [6]. The same study identified 67 putative J-proteins, with 5 type I J-proteins [6]. Phenotypic knockdown of T. brucei genes using RNAi, conducted by Alsford and colleagues [19], demonstrated that the Hsp70/J-protein machinery plays a prominent role in trypanosome biology, as the loss of certain members of these protein families impacted the survival and fitness of the parasite at various stages of its life cycle. Of significance to this study are the cytosolic Hsp70 proteins, TbHsp70 and TbHsp70.4, and their potential interactions with the cytosolic Type I J-protein, Tbj2.
It has been proposed that TbHsp70 is cytosolic and is an integral component of the heat shock response as it may play a key role in providing cytoprotection against cellular stress [6]. The Hsp70 orthologue in Leishmania chagasi has been linked to the parasite’s resistance to macrophage-induced oxidative stress [20], and has also been shown in several Leishmania spp. to be linked to parasite resistance to pentavalent antimonial treatment, as it induces Hsp70 expression, which in turn provides stress tolerance against the drug [21,22]. Based on phylogeny, TbHsp70.4 was found to form a distinct Hsp70/HSPA group unique to kinetoplastid parasites, with no obvious mammalian orthologues and a divergent C-terminal EEVD motif [6]. The Hsp70.4 orthologue in Leishmania major was shown to be cytoplasmic and constitutively expressed [23]. Tbj2 was identified to be an essential stress inducible Type I J-protein, as knockdown via RNAi revealed a severe growth defect and it was shown to reside in the parasite cytosol [24]. Tbj2 was determined to possess holdase activity when chemically denatured rhodanese and thermally aggregated malate dehydrogenase (MDH) were used as substrates [25].
Few TbHsp70/J-protein interactions have been biochemically characterized. This study aimed to investigate a potential functional co-operation between TbHsp70 and TbHsp70.4 with the co-chaperone Tbj2. The expression of TbHsp70 was determined to be heat inducible, whereas TbHsp70.4 was constitutive. Tbj2 stimulated the ATPase activities of both TbHsp70 and TbHsp70.4. TbHsp70 and TbHsp70.4 were both demonstrated to suppress the aggregation of thermally induced MDH in a dose dependent manner, and this was further enhanced by Tbj2. Furthermore, Tbj2 stimulated the refolding abilities of the Hsp70s of chemically denatured β-galactosidase. Tbj2 does functionally co-operate with both TbHsp70 and TbHsp70.4 and is part of the functional chaperone network that has been implicated in the proliferation and growth of parasitic cells. An increase in knowledge will enhance our comprehension of this important class of proteins and improve our understanding of the biology of the parasite, which may contribute to the development of new therapeutic strategies to combat African Trypanosomiasis.
2. Results and Discussion
2.1. Recombinant Protein Production and Purification
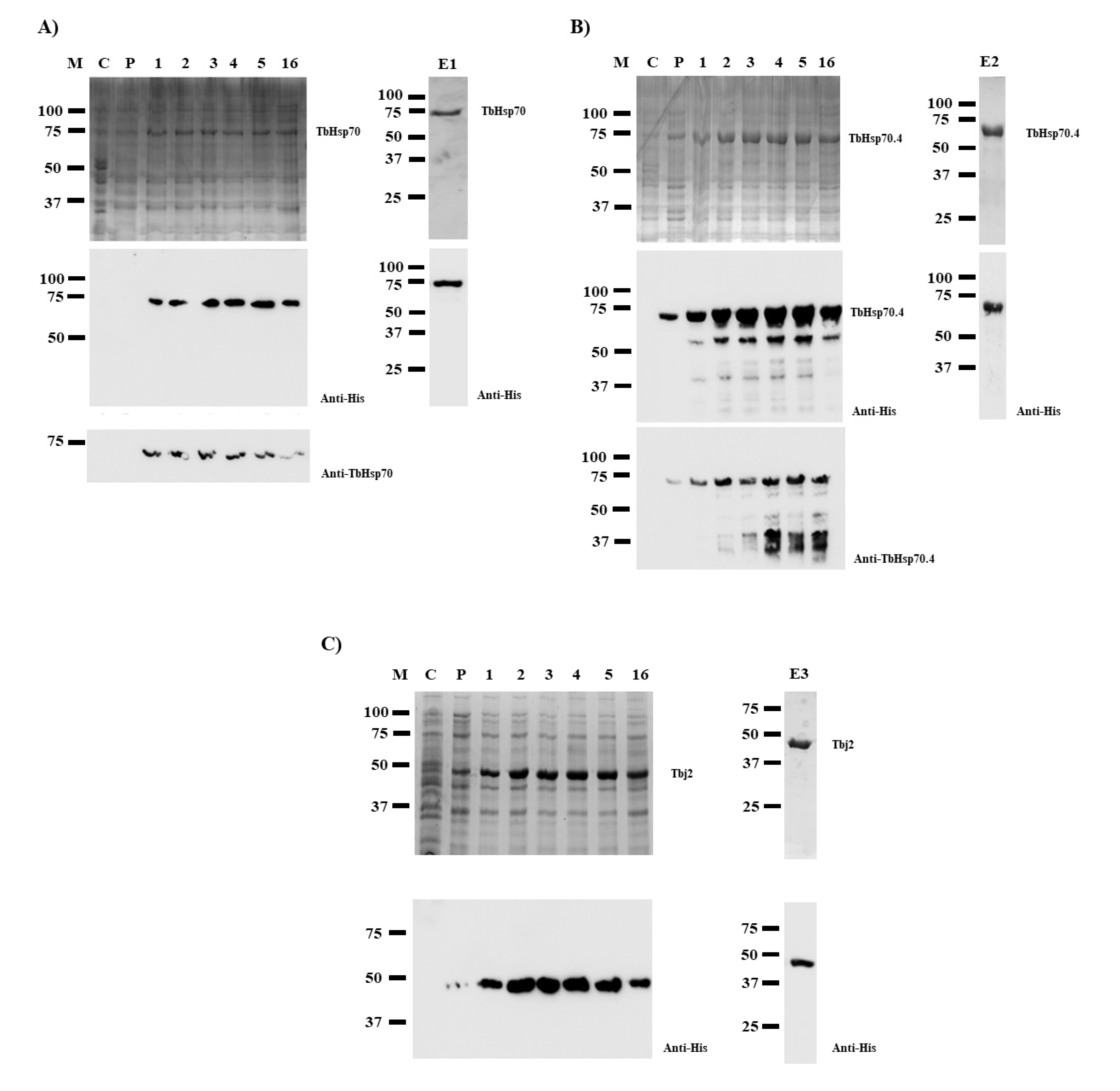
SDS-PAGE analyses revealed that TbHsp70 and TbHsp70.4 proteins were successfully expressed in E. coli XL1 Blue cells as species of approximately 71 kDa (Figure 1A,B). However, Western blot analyses revealed protein species of lower molecular weights when TbHsp70.4 were produced in E. coli, along with the full-length protein using both anti-TbHsp70.4 and anti-His antibodies (Figure 1B). These protein species were either N-terminally His-tagged truncated versions of the full-length TbHsp70.4 protein or products of incomplete synthesis. Despite this, the full-length TbHsp70.4 recombinant protein was purified by nickel affinity chromatography. Western blot analyses of the eluted proteins produced after purification by nickel affinity chromatography revealed that both His-tagged TbHsp70 and TbHsp70.4 proteins were successfully purified (Figure 1, lanes E1 and E2). Tbj2 was expressed in E. coli BL21 (DE3) cells (Figure 1C), and subsequently purified using nickel affinity chromatography as previously described [25]. Western blot analysis of the eluted protein is shown in Figure 1, lane E3.
2.2. Protein Expression of TbHsp70 and TbHsp70.4 Is Modulated in Response to Heat Stress
The complex life cycle of T. brucei displays cell forms with different morphology and functional characteristics that enable the parasites to interact with both its insect vector and a mammalian host, undergoing several environmental variations in the process [26]. Hsp70 proteins have been shown in eukaryotic and prokaryotic cells to be self-protective proteins that maintain cell homeostasis against a wide variety of stressors as an adaptive response [27]. SDS-PAGE and Western blot analyses were used to assess the protein expression of TbHsp70 and TbHsp70.4 in T. brucei parasites at the bloodstream stage under normal conditions (37 °C) and heat shock conditions (42 °C), respectively, for 1 h (Figure 2A). Both Hsp70 proteins were shown to be expressed under normal culture conditions of 37 °C (Figure 2A). Even though there was no notable difference in the protein levels of TbHsp70 and TbHsp70.4 on the SDS-PAGE gel, TbHsp70 protein expression was shown to be up-regulated in response to heat shock, whereas TbHsp70.4 was slightly down-regulated as shown in the Western blot analyses (Figure 2A,B). The differences in the protein levels observed for TbHsp70 and TbHsp70.4 for the various treatments were not due to differences in loading, as the levels of actin, the loading control, remained unchanged (Figure 2A, lower panel).
The mRNA levels of TbHsp70 have been shown previously to be up-regulated in both transgenic bloodstream and procyclic form parasites in response to heat shock [28], as well as conditions that mimic mammalian infection [29]. Expression of TcHsp70, an orthologue of TbHsp70 in Trypanosoma cruzi, has also been investigated, and it has been shown that TcHsp70 synthesis increases 4- to 5-fold after heat shock [30]. Thus, the up-regulation in the expression of TbHsp70 is indicative that the Hsp70 protein may play an integral role in cytoprotection of T. brucei parasites in response to various environmental stresses. The protein expression of TbHsp70.4 in response to heat stress is consistent with the findings observed for its orthologue in Leishmania major (LmjHsp70.4) [23]. The protein expression of LmHsp70.4 in promastigote parasites was shown to have no difference in expression after a period of heat stress [23]. In the cytoplasm of mammalian cells, the housekeeping functions are exerted by the constitutively expressed, cognate Hsp70 isoform, HSPA8 [31,32]. These functions include protein biogenesis, degradation, and protein translocation [8], which could infer that TbHsp70.4 may play a similar role in T. brucei. The modulation in protein expression of these Hsp70 proteins could indicate that Hsp70.4 and Hsp70 proteins in T. brucei and other kinetoplastid parasites may represent the cognate and inducible Hsp70 isoforms, respectively. As mentioned previously, Tbj2 has been shown to be heat stress inducible and resides within the cytosol [24]. Both TbHsp70 and TbHsp70.4 have been shown to localize in the cytosol of the parasite [33]. Thus, the localization for both T. brucei Hsp70s and Tbj2 may suggest possible colocalization, and that these proteins could functionally co-operate within the cytosol of the parasite.
2.3. TbHsp70 and TbHsp70.4 Both Possess Intrinsic ATPase Activity, Which Is Stimulated by Tbj2
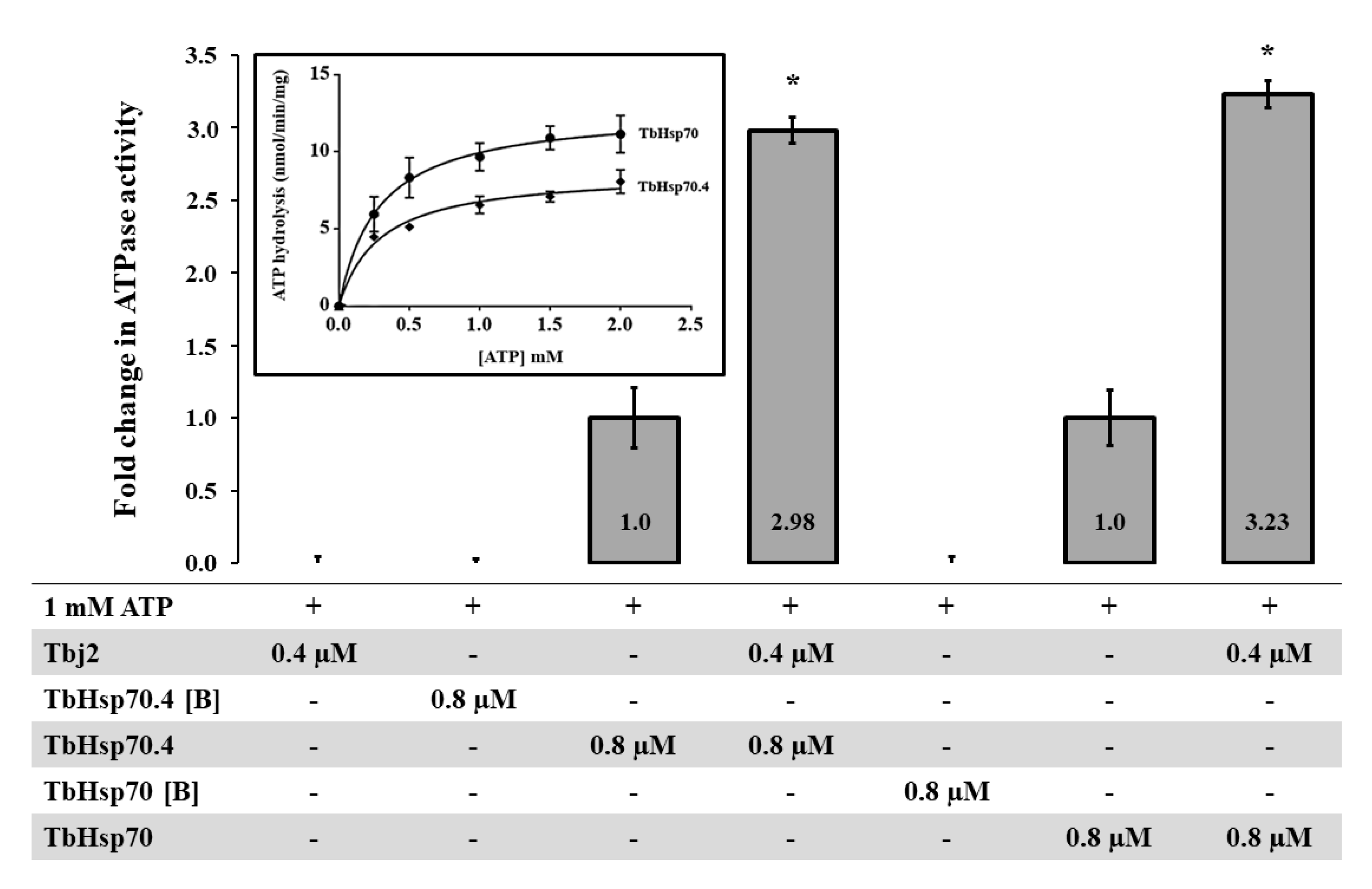
The basal ATPase activities for both TbHsp70 and TbHsp70.4 were determined using a colorimetric assay and were conducted under variable concentrations (0–2000 µM) of ATP. The Michaelis–Menten plots were generated from three independent batches of TbHsp70 and TbHsp70.4 (Figure 3). Both TbHsp70 and TbHsp70.4 exhibited intrinsic ATPase activity (Figure 3), though TbHsp70 was found to have a higher basal ATPase activity than that of TbHsp70.4.
As mentioned, the basal ATPase activity of Hsp70 is modulated by a cohort of co-chaperones, with the J-protein family being the most prominent. J-proteins stimulate the rate-limiting ATP hydrolysis step in the Hsp70 catalytic cycle, which facilitates substrate capture. Using a colorimetric assay, the effect of Tbj2 on the basal ATPase activity of TbHsp70 and TbHsp70.4 was explored. The basal ATPase activity of TbHsp70 and TbHsp70.4 were represented as 1, with modulation represented as a fold change in basal ATPase activity (Figure 3). Tbj2 was shown to moderately stimulate both the basal ATPase activities of TbHsp70 and TbHsp704 by approximately 3-fold (Figure 3). None of Tbj2, TbHsp70, or TbHsp70.4 denatured by boiling displayed any ATPase activity (Figure 3), indicating that the increased phosphate release was due to stimulation of the Hsp70 ATPase activity. The magnitude of Hsp70 ATPase activity stimulation by Tbj2 is comparable to the study by Burger et al. [25] describing stimulation of TbHsp70.c and TcHsp70B by Tbj2. Tcj2, Trypanosoma cruzi orthologue to Tbj2, was shown to stimulate the basal ATPase activity of TcHsp70B by 1.5-fold [34]. Thus, the results of these findings indicate that Tbj2 can functionally co-operate with either of the cytosolic T. brucei Hsp70s.
2.4. TbHsp70 and TbHsp70.4 Both Suppress the Thermal Aggregation of Malate Dehydrogenase (MDH)
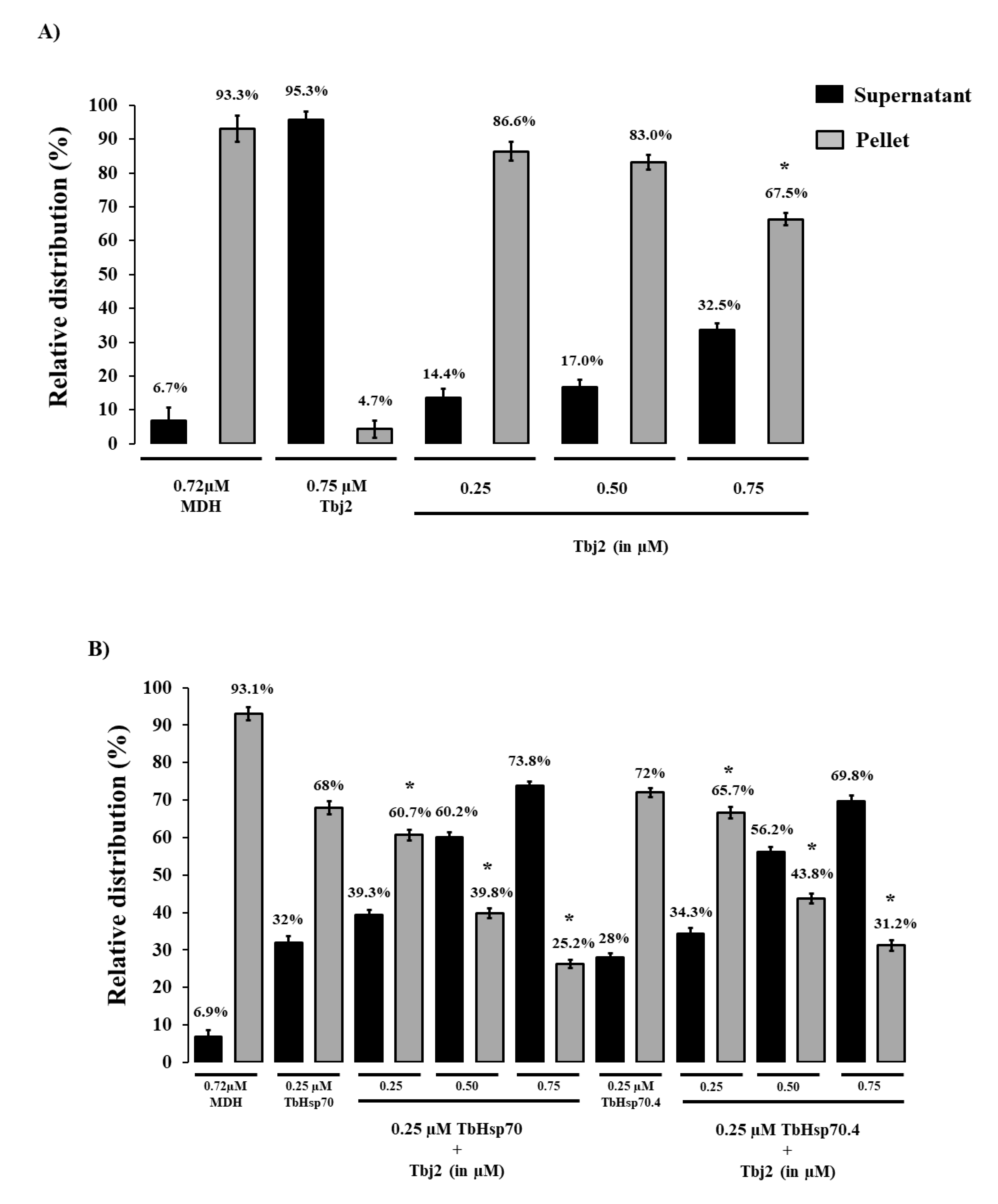
The holdase function of TbHsp70 and TbHsp70.4 in the presence and absence of Tbj2 was determined by assessing its ability to prevent the thermally-induced aggregation of the model substrate, malate dehydrogenase (MDH). The MDH aggregation suppression assay was adapted from [35]. MDH was subjected to heat stress at 48 °C and, as expected, the protein aggregated in the absence of chaperones (Figure 4). In the presence of BSA, a non-chaperone protein, MDH also aggregated in response to heat stress (Figure A1). In the absence of MDH, both T. brucei Hsp70s and Tbj2 were found to be stable under the assay conditions as no significant aggregation was observed (Figure 4A and Figure A1). The addition of either TbHsp70 or TbHsp70.4 at varying concentrations (0.25–0.75 µM) were both shown to suppress the thermally induced aggregation of MDH in a dose dependent manner (Figure A1). Though, TbHsp70 was shown to be more capable at suppressing the aggregation of MDH than was TbHsp70.4 (Figure A1). TbHsp70 was shown to be heat inducible, and thus a more proficient holdase function may be necessary for effective maintenance of the proteostasis under stressful conditions.
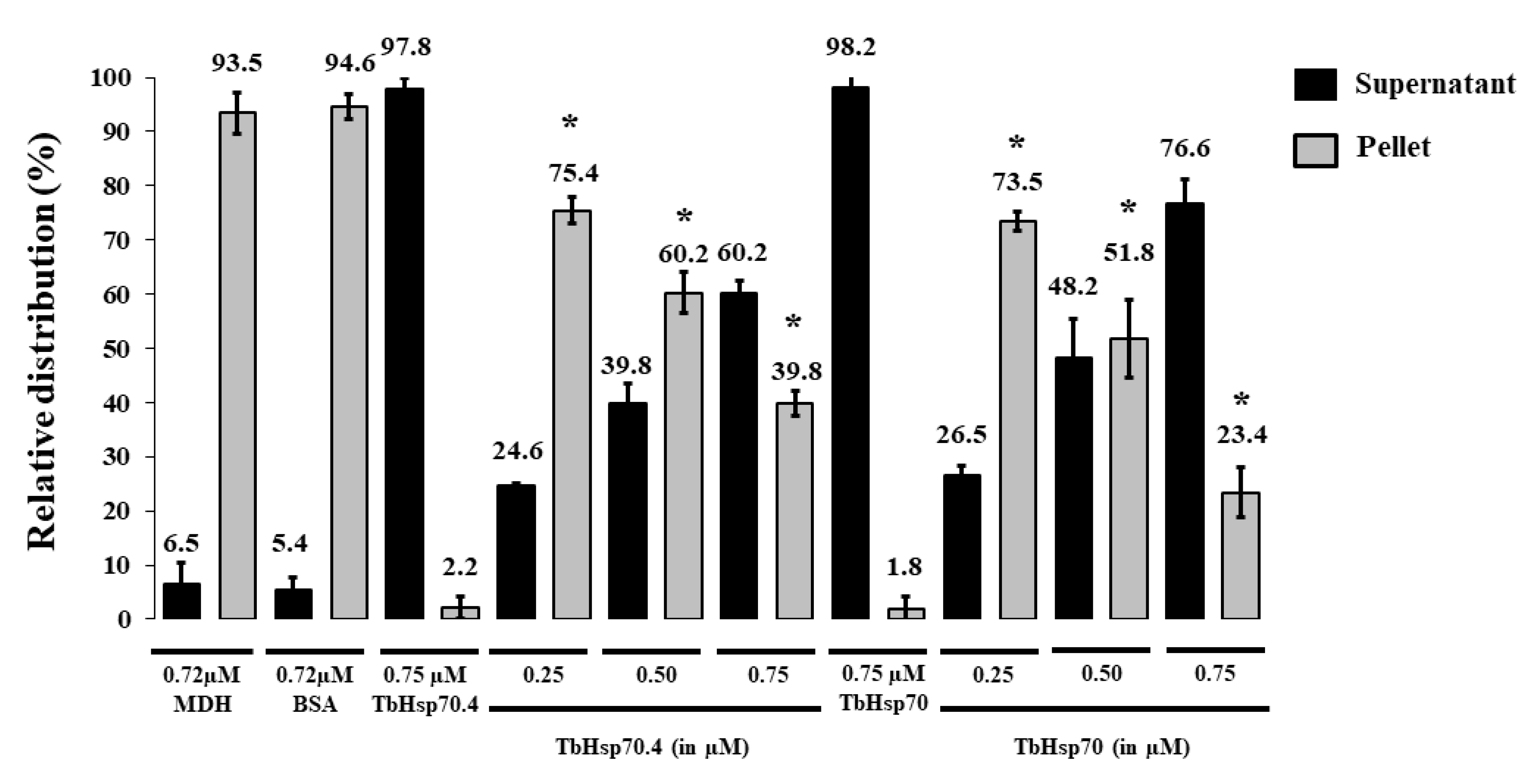
The addition of Tbj2 at varying concentrations was also found to suppress the thermal aggregation of MDH (Figure 4A), which is consistent with previous findings [25]. The Type I J-protein subfamily has been shown to possess intrinsic chaperone activity to select and deliver nascent polypeptides to their Hsp70 chaperone partner [36]. The demonstrated ability of Tbj2 to bind and suppress protein aggregates indicates that this J-protein may also play a prominent role in parasite cytoprotection. The ability of Tbj2 to enhance the holdase function of the T. brucei Hsp70s was assessed by maintaining a constant Hsp70 concentration and varying that of the J-protein (Figure 4B). TbHsp70 and TbHsp70.4 (0.25 µM) suppressed MDH aggregation suppression by 32% and 28%, respectively (Figure 4B). The addition of Tbj2 at equimolar concentration (0.25 µM) to TbHsp70 and TbHsp70.4 resulted in 39.3% and 34.3% MDH aggregation suppression, respectively (Figure 4B). Increasing concentrations of Tbj2 resulted in 60.2% (0.5 µM) and 73.8% (0.75 µM) MDH aggregation suppression for TbHsp70, and 56.2% (0.5 µM) and 69.8% (0.75 µM) MDH aggregation suppression for TbHsp70.4 (Figure 4B). Despite Tbj2 being shown to possess independent chaperone activity (Figure 4A), the modulatory effect of Tbj2 on the holdase activity of the T. brucei Hsp70s was not found to be additive, indicating a demonstrated co-operative effect and thus prompting Tbj2 as a potential co-chaperone to both TbHsp70 and TbHsp70.4.
2.5. Tbj2 Stimulates the Refoldase Activities of TbHsp70 and TbHsp70.4
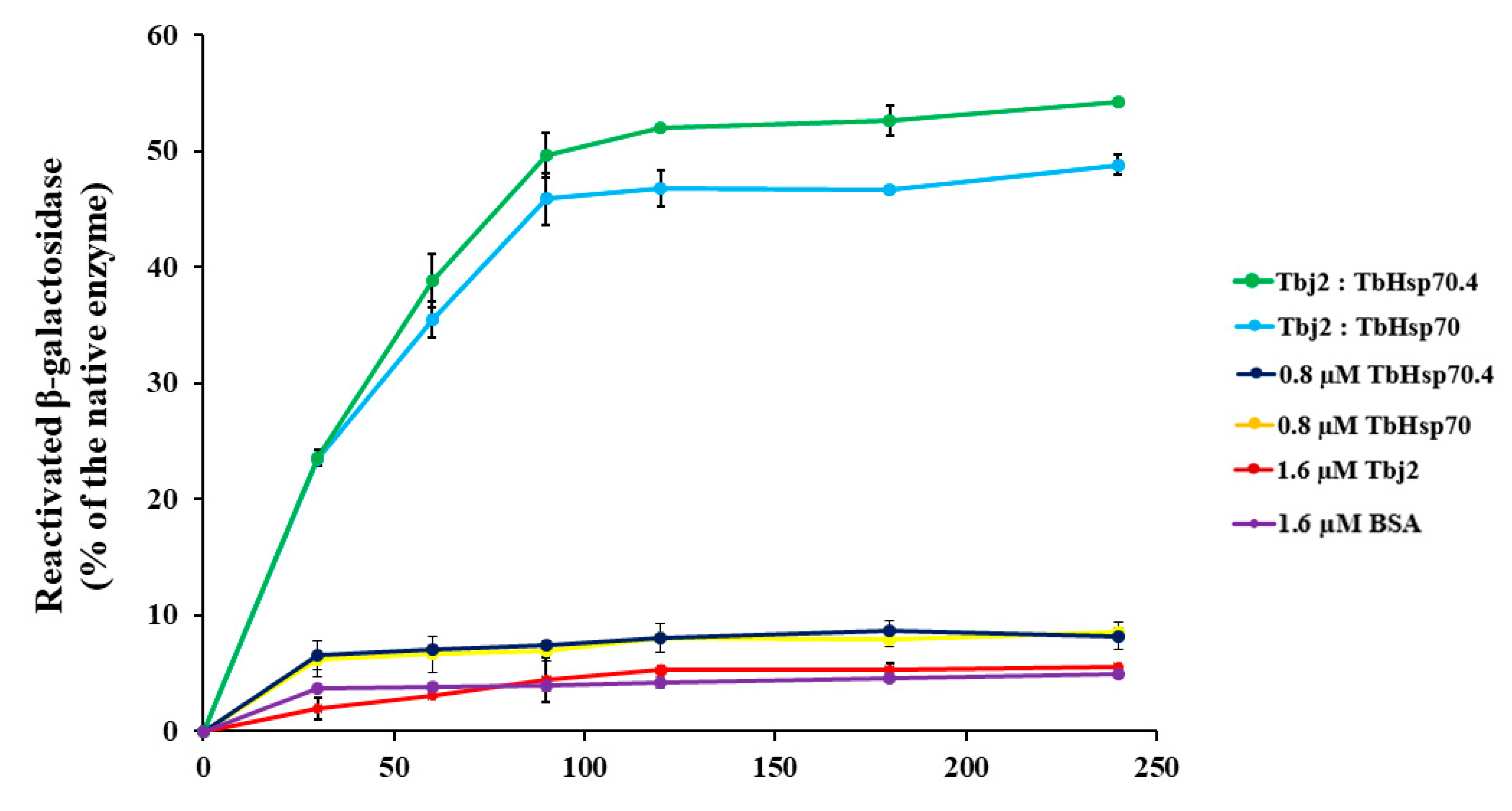
A β-galactosidase refolding assay, adapted from [37], was used to investigate the refoldase activity of TbHsp70 and TbHsp70.4 in the presence and absence of Tbj2. β-galactosidase recognizes and cleaves the chromogenic substrate, o-nitrophenyl-β-D-galactoside (ONPG), into ortho-nitrophenol (ONP) and galactose. Absorbance at 420 nm measures the amount of ONP produced in the reaction, which reflects the ability of the molecular chaperones to refold chemically denatured β-galactosidase. In these experiments, the activity of β-galactosidase without denaturation was considered as 100%. It was shown that the T. brucei Hsp70s were not able to proficiently mediate β-galactosidase refolding (Figure 5). Human Hsp70 has also been demonstrated to be unable to independently mediate the refolding of chemically denatured β-galactosidase [37] or thermally denatured luciferase [38]. It is suggested that the Hsp70 preferentially maintains the non-native substrate in a ‘folding-competent’ state that, upon addition of a J-protein co-chaperone, leads to refolding of the client protein [37]. The addition of Tbj2 to TbHsp70 or TbHsp70.4 resulted in the stimulation of the refoldase activity of the Hsp70s, and approximately 50% of the non-native β-galactosidase being refolded (Figure 5). These findings, however, are comparable with those reported for the refolding of non-native β-galactosidase mediated by human Hsp70 and the Type I J-protein, Hdj1, which was shown to reactivate approximately 40% of the denatured enzyme [37]. However, the stimulation of the refoldase activity of both TbHsp70 and TbHsp70.4 by Tbj2 indicates a functional Hsp70/J-protein association.
3. Materials and Methods
3.1. The pQE2/TbHsp70 Expression Vector
The codon optimized coding sequence for expression of TbHsp70 (TriTrypDB accession number: Tb927.11.11330) in Escherichia coli was synthesized and supplied by the GenScript Corporation (Piscataway, NJ, USA). The TbHsp70 coding region was inserted into the pQE2 expression vector (Qiagen, Valencia, CA, USA) using NdeI and HindIII restriction sites. The integrity of the pQE2/TbHsp70 vector which was used to express the N-terminal His-tagged TbHsp70 was verified by restriction analysis and DNA sequencing.
3.2. Heterologous Expression and Purification of TbHsp70 and TbHsp70.4
E. coli XL1 Blue cells transformed with either pQE2/TbHsp70 or pQE2/TbHsp70.4 were grown at 37 °C in 2× YT medium supplemented with 100 μg/mL ampicillin and grown to mid-logarithmic phase (A600 0.4–0.6). Protein production was induced by the addition of 1 mM IPTG (isopropyl-β-D-thiogalactopyranoside), and the bacterial cultures were incubated at 37 °C for 3 h for TbHsp70 and 1 h for TbHsp70.4. Bacterial cells expressing TbHsp70 or TbHsp70.4 were harvested by centrifugation (10,000× g; 15 min; 4 °C) and the cell pellet was resuspended in lysis buffer (100 mM Tris-HCl, pH 7.5, 300 mM NaCl, 20 mM imidazole, 1 mM PMSF, 1 mg/mL lysozyme), allowed to stand for 30 min at room temperature, and then frozen at −80 °C overnight. The cells were then thawed on ice and sonicated at 4 °C. The resulting lysate was cleared by centrifugation (13,000× g, 40 min, 4 °C) and the supernatant was incubated with cOmplete His-tag purification resin (Roche, Mannheim, Germany) and allowed to bind overnight at 4 °C with gentle agitation. The resin was then pelleted by centrifugation (4500× g; 4 min) to remove unbound proteins and washed three times using native wash buffer (100 mM Tris-HCl, pH 7.5, 300 mM NaCl, 50 mM imidazole, 1 mM PMSF) to remove non-specific contaminants. The bound protein was eluted three times by re-suspending the resin in elution buffer (10 mM Tris-HCl, pH 7.5, 300 mM NaCl, 750 mM imidazole). The eluted proteins were extensively dialyzed using SnakeSkin dialysis tubing (Pierce-MWCO 10,000; Thermo Scientific, Waltham, MA, USA) into either dialysis buffer (DB; 10 mM Tris, pH 7.5, 100 mM NaCl, 0.5 mM DTT, 10% (v/v) glycerol, 50 mM KCl, 2 mM MgCl2) or into the appropriate assay buffer for functional studies and then subsequently concentrated against PEG 20,000 (Merck, Darmstadt, Germany). The protein yield was estimated using the Bradford assay (Sigma-Aldrich, St. Louis, MO, USA) with BSA as the standard. SDS-PAGE (10%) and Western blot analysis using mouse monoclonal anti-His primary antibody and HRP-conjugated goat anti-mouse IgG secondary antibody (Santa Cruz Biotechnology, Santa Cruz, CA, USA) were conducted to assess the expression and purification of the recombinant proteins. TbHsp70 and TbHsp70.4 protein expression in E. coli was also confirmed by Western blot using rabbit-polyclonal anti-TbHsp70 and rabbit-polyclonal anti-TbHsp70.4 [39,40], respectively. HRP-conjugated goat anti-rabbit (Santa Cruz Biotechnology Inc., USA) was used as the secondary antibody. Imaging of the protein bands on the blot was conducted using the ECL kit (Thermo Scientific, USA) as per manufacturer’s instructions. Images we captured using the ChemiDoc Imaging system (Bio-Rad, Hercules, CA, USA).
3.3. Purification of Tbj2
Recombinant N-terminal His-tagged Tbj2 was purified under native conditions from E. coli BL21 (DE3) cells as previously described [25]. Samples were dialyzed in DB or into the appropriate assay buffer for functional studies.
3.4. Investigation of Heat-Induced Expression of TbHsp70 and TbHsp70.4 in T. b. brucei Lister 927 V221 Parasites
Wild type T. b. brucei Lister 927 v221 bloodstream form lysates (106 cells/mL) were used for the heat stress inducibility experiment. The bloodstream form of the T. b. brucei Lister 927 v221 strain trypanosome parasites were cultured in filter sterilized complete Iscoves Modified Dulbeccos Media (IMDM) based HM1-9 medium (IMDM base powder, 3.6 mM sodium bicarbonate, 1 mM hypoxanthine, 1 mM sodium pyruvate, 0.16 mM thymidine, 0.05 mM bathocuprone sulphate acid, 10% (v/v) heat inactivated Foetal Bovine Serum, 1.5 mM L-cysteine, 0.2 mM β-mercaptoethanol, pH 7.5) in a humidified chamber at 37 °C with an atmosphere of 5% CO2. Separate 25 mL cultures of cells were exposed to heat shock at 42 °C for a period of 120 min in plugged flasks, allowing no entry of CO2. A control experiment was performed under the same conditions maintaining the temperature at 37 °C. Cell lysates were harvested by centrifugation at 800× g for 10 min, washed twice in 1× PBS buffer, and repelleted prior to resuspension in SDS-PAGE loading buffer to a final cell count of 5 × 105 cells/µL. The lysates (5 × 106 cells per lane) were resolved on a 10% SDS-PAGE gel. Differences in TbHsp70 and TbHsp70.4 protein expression were detected using rabbit-polyclonal anti-TbHsp70 and rabbit-polyclonal anti-TbHsp70.4, respectively, and goat anti-rabbit IgG HRP-conjugated secondary antibody (Santa Cruz Biotechnology Inc., Santa Cruz, CA, USA) in subsequent Western blot analysis. Actin was also probed as a loading control using mouse monoclonal anti-actin antibody and HRP-conjugated goat anti-mouse IgG secondary antibody (Santa Cruz Biotechnology Inc., Santa Cruz, CA, USA). Images were acquired using the ChemiDoc Imaging system (Bio-Rad, Hercules, CA, USA), and densitometric analyses of bands were conducted using the Image Lab v5.1 built 8 (Bio-Rad, Hercules, CA, USA).
3.5. MDH Aggregation Suppression Assay
The capacity of TbHsp70 and TbHsp70.4 to suppress the thermally-induced aggregation of the model substrate, malate dehydrogenase (MDH) from porcine heart (Sigma-Aldrich, St. Louis, MO, USA.), alone or in the presence of Tbj2 was adapted from [35]. Briefly, the reaction was initiated by adding 0.72 µM MDH to varying concentrations of the T. brucei Hsp70s in assay buffer (50 mM Tris-HCl pH 7.4, 100 mM NaCl) either alone or in combination with Tbj2. The reaction was then heated at 48 °C for an hour. After incubation, the samples were centrifuged at 13,000× g for 10 min to separate the soluble and insoluble fractions. These fractions were analyzed using 10% SDS-PAGE, and subsequently quantified using densitometric analysis using the Image Lab v5.1 built 8 (Bio-Rad, Hercules, CA, USA). A negative control of BSA (0.75 µM) was added to illustrate that MDH-aggregation suppression was due to the holdase function of the T. brucei molecular chaperones and not simply due to the presence of a second protein in the assay system. As a control, the aggregation of the chaperones was monitored in the assay buffer without MDH. Each assay was conducted in triplicate and three independently purified batches of proteins were used.
3.6. ATPase Activity Assay
The determination of the basal ATPase activity of TbHsp70 and TbHsp70.4 were performed using a high throughput colorimetric ATPase assay kit (Innova Biosciences, Cambridge, UK). Summarily, the method allows for the quantification of the inorganic phosphate (Pi) released from ATP hydrolysis by an enzyme. Briefly, the Hsp70s were prepared in ATPase assay buffer (100 mM Tris-HCl, 7.5, 2 mM MgCl2, 50 mM KCl, 0.5 mM DTT) and incubated with varying ATP concentrations (0–2 mM) for 1 h at 37 °C. The samples containing Pi hydrolyzed from ATP were incubated with the PiColorLock™ solution, which is a malachite green dye solution that in the presence of Pi changes absorbance due to the generation of molybdate-phosphate complexes. The absorbance was measured at 595 nm using a Powerwave 96-well plate reader (BioTek Instruments Inc., Winooski, VT, USA), and absorbance values were converted to phosphate concentrations using a standard curve of absorbance vs. phosphate concentration based on a set of Pi standards provided by the supplier and assayed along with the samples. All samples were corrected for spontaneous breakdown of ATP observed in a control experiment in the absence of protein. A Michaelis–Menten kinetic plot was constructed and used to determine the kinetic specific activity (Vmax) and Km for the basal ATPase activity of TbHsp70 and TbHsp70.4. A non-linear regression curve was fitted to the Michaelis–Menten plot using GraphPad Prism® (v. 7.0; San Diego, CA, USA.) software. The assay was conducted in triplicate on three independently purified batches of proteins.
To investigate the modulatory effect of Tbj2 on the basal ATPase activity of the T. brucei Hsp70s, the molecular chaperones at indicated concentrations were prepared in ATPase assay buffer (100 mM Tris-HCl, 7.5, 2 mM MgCl2, 50 mM KCl, 0.5 mM DTT) and incubated with 1 mM ATP for 1 h at 37 °C. The color development and absorbance measurement procedures were conducted as previously described. All samples were corrected for spontaneous breakdown of ATP observed in a control experiment in the absence of protein. Additional control of the respective boiled Hsp70 protein was used to cater for the spontaneous hydrolysis of ATP. Any background ATP hydrolysis observed for Tbj2 was corrected for by subtracting this activity from the reactions containing these proteins. The modulatory effect on the ATPase activity of the T. brucei Hsp70s was represented as fold change with the basal ATPase activity of the Hsp70s taken as 1. The assay was conducted in triplicate on three independently purified batches of protein.
3.7. β-galactosidase Refolding Assay
Investigation of the ability of the T. brucei Hsp70s to refold chemically denatured β-galactosidase alone or in combination with Tbj2 was carried out as previously described [37]. The catalytic hydrolysis activity of the refolded β-galactosidase using 2-Nitrophenyl β-D-galactopyranoside (ONPG) as a chromogenic substrate [41] was measured in relation to native β-galactosidase and used as a means to determine the refoldase activity of Hsp70. After denaturation of β-galactosidase (Sigma-Aldrich, St. Louis, MO, USA) for 30 min at 30 °C in denaturation buffer (25 mM HEPES, pH 7.5, 5 mM MgCl2, 50 mM KCl, 5 mM β-mercaptoethanol, and 6 M guanidine-HCl), denatured β-galactosidase to a final concentration of 3.4 nM was diluted 1:125-fold into refolding buffer (25 mM HEPES pH 7.5, 5 mM MgCl2, 50 mM KCl, 2 mM ATP, and 10 mM DTT) supplemented with the T. brucei Hsp70s alone or in the presence of Tbj2 at indicated concentrations, and incubated for 2 h at 37 °C. Reactions containing 1.6 µM BSA (no molecular chaperones) with native and denatured β-galactosidase were conducted to serve as positive and negative controls, respectively. The negative control consisted of BSA (1.6 µM) in the presence of denatured β-galactosidase enzyme to determine the kinetics of spontaneous refolding of the denatured β -galactosidase enzyme. The activity of β-galactosidase was measured at various time points by mixing 10 µL of each refolding reaction with 10 µL of ONPG, followed by incubation at 37 °C for 15 min. Assays were terminated by the addition of 0.5 M sodium carbonate. Absorbance was recorded at 420 nm for each reaction. The percentage refolding activity was calculated relative to the activity of native β-galactosidase. The assay was conducted in triplicate on three independently purified batches of protein.
4. Conclusions
It has become increasingly evident that the Hsp70/J-protein machinery is essential to the survival, pathogenicity, and differentiation of kinetoplastid parasites. However, further elucidation of the molecular details of the Hsp70/J-protein chaperone interactions and pathways are required, as some of these pathways may represent a novel means of chemotherapeutic intervention for African trypanosomiasis. This study aimed to investigate the cytosolic Hsp70 system through in vitro biochemical characterization of the chaperone properties of the predicted cytosolic T. brucei Hsp70s, TbHsp70, and TbHsp70.4, and their potential partnership with the cytosolic Type I J-protein, Tbj2. We have provided the first evidence that TbHsp70 and TbHsp70.4 represent the stress inducible and cognate Hsp70 isoforms, respectively. Thus, TbHsp70 may represent an essential component in cytoprotection of parasites, whereas TbHsp70.4 fulfils crucial housekeeping roles. Tbj2 was shown to stimulate the basal ATPase activity of both Hsp70s and functionally co-operate in mediating the reactivation of client proteins. Overall, this study provides a greater understanding of the T. brucei Hsp70 chaperone system. Further studies, however, are required to explore the T. brucei Hsp70/J-protein machinery and to identify small-molecule inhibitors capable of disrupting these systems.
Author Contributions
S.J.B. and A.B. designed the research; S.J.B. performed all experiments. S.J.B. analyzed the data; S.J.B. and A.B. wrote the manuscript.
Funding
This work was funded by a grant from the National Research Foundation (NRF), grant number 87663. S.J.B. was a recipient of an NRF Doctoral Innovation Scholarship.
Acknowledgments
The pET28a/Tbj2 and pQE2/TbHsp70.4 expression vectors were kindly provided by Michael Ludewig (Rhodes University, South Africa). The T. b. brucei Lister 927 variant 221 strain was a kind donation from George Cross (Rockefeller University, New York, U.S.A.). Thanks to Michelle Isaacs (Rhodes University) for T. b. brucei culturing at the parasite facility, which is supported by the South African Medical Research Council (MRC). The rabbit polyclonal anti-TbHsp70 and rabbit polyclonal anti-TbHsp70.4 antibodies were generous gifts from Ariel Louwrier (StressMarq Biosciences Inc., Canada) and Jay Bangs (University of Buffalo, New York, U.S.A.), respectively.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
Appendix A
Figure A1.
TbHsp70 and TbHsp70.4 suppress the thermal aggregation of MDH. The holdase function of recombinant TbHsp70 and TbHsp70.4 was conducted by monitoring the heat-induced aggregation of MDH (0.72 µM) in vitro at 48 °C and by quantifying the pellet (insoluble; grey bars) and supernatant (soluble; black bars) fractions after heat exposure. The thermal aggregation of MDH in the presence of a non-chaperone, BSA, is shown. TbHsp70.4 and TbHsp70 in the absence of MDH were not prone to aggregation under the assay conditions. Standard deviations were obtained from three replicate assays on three independent batches of purified protein. A statistically significant difference between a reaction and MDH alone are indicated by * (p < 0.05) above the reaction using a Student’s t-test.
Figure A1.
TbHsp70 and TbHsp70.4 suppress the thermal aggregation of MDH. The holdase function of recombinant TbHsp70 and TbHsp70.4 was conducted by monitoring the heat-induced aggregation of MDH (0.72 µM) in vitro at 48 °C and by quantifying the pellet (insoluble; grey bars) and supernatant (soluble; black bars) fractions after heat exposure. The thermal aggregation of MDH in the presence of a non-chaperone, BSA, is shown. TbHsp70.4 and TbHsp70 in the absence of MDH were not prone to aggregation under the assay conditions. Standard deviations were obtained from three replicate assays on three independent batches of purified protein. A statistically significant difference between a reaction and MDH alone are indicated by * (p < 0.05) above the reaction using a Student’s t-test.

References
- Alsan, M. The effect of the tsetse fly on African development. Am. Econ. Rev. 2015, 105, 382–410. [Google Scholar] [CrossRef]
- Barrett, M.P.; Croft, S.L. Management of trypanosomiasis and leishmaniasis. Br. Med. Bull. 2012, 104, 175–196. [Google Scholar] [CrossRef] [PubMed]
- Shonhai, A.; Maier, A.G.; Przyborski, J.M.; Blatch, G.L. Intracellular protozoan parasites of humans: The role of molecular chaperones in development and pathogenesis. Protein Pept. Lett. 2011, 18, 143–157. [Google Scholar] [CrossRef] [PubMed]
- Requena, J.M.; Montalvo, A.M.; Fraga, J. Molecular chaperones of Leishmania: Central players in many stress-related and-unrelated physiological processes. BioMed Res. Int. 2015, 2015, 301326. [Google Scholar] [CrossRef] [PubMed]
- Urmenyi, T.P.; Silva, R.; Rondinelli, E. The heat shock proteins of Trypanosoma cruzi. In Proteins and Proteomics of Leishmania and Trypanosoma; Santos, A., Branquinha, M., d’Avila-Levy, C., Kneipp, L., Sodré, C., Eds.; Springer: Dordrecht, The Netherlands, 2014; Volume 74, pp. 119–135. [Google Scholar]
- Bentley, S.J.; Jamabo, M.; Boshoff, A. The Hsp70/J-protein machinery of the African trypanosome, Trypanosoma brucei. Cell Stress Chaperones 2019, 24, 125–148. [Google Scholar] [CrossRef] [PubMed]
- Mayer, M.P.; Bukau, B. Hsp70 chaperones: Cellular functions and molecular mechanism. Cell Mol. Life Sci. 2005, 62, 670–684. [Google Scholar] [CrossRef] [PubMed]
- Bukau, B.; Weissman, J.; Horwich, A. Molecular chaperones and protein quality control. Cell 2006, 125, 443–451. [Google Scholar] [CrossRef]
- Daugaard, M.; Rohde, M.; Jäättelä, M. The heat shock protein 70 family: Highly homologous proteins with overlapping and distinct functions. FEBS Lett. 2007, 581, 3702–3710. [Google Scholar] [CrossRef]
- Brocchieri, L.; de Macario, E.C.; Macario, A.J.L. Hsp70 genes in the human genome: Conservation and differentiation patterns predict a wide array of overlapping and specialized functions. BMC Evol. Biol. 2008, 8, 19. [Google Scholar] [CrossRef]
- Flaherty, K.M.; Deluca-Flaherty, C.; McKay, D.B. Three-dimensional structure of the ATPase fragment of a 70K heat shock cognate protein. Nature 1990, 346, 623–628. [Google Scholar] [CrossRef]
- Wang, T.F.; Chang, J.H.; Wang, C. Identification of the peptide binding domain of hsc70. J. Biol. Chem. 1993, 268, 26049–26051. [Google Scholar] [PubMed]
- Slepenkov, S.V.; Witt, S.N. The unfolding story of the Escherichia coli Hsp70 DnaK: Is DnaK a holdase or an unfoldase? Mol. Microbiol. 2002, 45, 1197–1206. [Google Scholar] [CrossRef] [PubMed]
- Clerico, E.M.; Tilitsky, J.M.; Meng, W.; Gierasch, L.M. How hsp70 molecular machines interact with their substrates to mediate diverse physiological functions. J. Mol. Biol. 2015, 427, 1575–1588. [Google Scholar] [CrossRef] [PubMed]
- Meimaridou, E.; Gooljar, S.B.; Chapple, J.P. From hatching to dispatching: The multiple cellular roles of the Hsp70 molecular chaperone machinery. J. Mol. Endocrinol. 2009, 42, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Hennessy, F.; Nicoll, W.S.; Zimmermann, R.; Cheetham, M.E.; Blatch, G.L. Not all J domains are created equal: Implications for the specificity of Hsp40–Hsp70 interactions. Protein Sci. 2005, 14, 1697–1709. [Google Scholar] [CrossRef]
- Cheetham, M.E.; Caplan, A.J. Structure, function and evolution of DnaJ: Conservation and adaptation of chaperone function. Cell Stress Chaperones 1998, 3, 28–36. [Google Scholar] [CrossRef]
- Botha, M.; Pesce, E.R.; Blatch, G.L. The Hsp40 proteins of Plasmodium falciparum and other apicomplexa: Regulating chaperone power in the parasite and the host. Int. J. Biochem. Cell Biol. 2007, 39, 1781–1803. [Google Scholar] [CrossRef]
- Alsford, S.; Turner, D.J.; Obado, S.O.; Sanchez-Flores, A.; Glover, L.; Berriman, M.; Hertz-Fowler, C.; Horn, D. High-throughput phenotyping using parallel sequencing of RNA interference targets in the African trypanosome. Genome Res. 2011, 21, 915–924. [Google Scholar] [CrossRef]
- Miller, M.A.; McGowan, S.E.; Gantt, K.R.; Champion, M.; Novick, S.L.; Andersen, K.A.; Bacchi, C.J.; Yarlett, N.; Britigan, B.E.; Wilson, M.E. Inducible resistance to oxidant stress in the protozoan Leishmania chagasi. J. Biol. Chem. 2000, 275, 33883–33889. [Google Scholar] [CrossRef]
- Maharjan, M.; Madhubala, R. Heat shock protein 70 (HSP70) expression in antimony susceptible/resistant clinical isolates of Leishmania donovani. Nepal J. Biotechnol. 2015, 3, 22–28. [Google Scholar] [CrossRef]
- Codonho, B.S.; Costa, S.; Peloso Ede, F.; Joazeiro, P.P.; Gadelha, F.R.; Giorgio, S. HSP70 of Leishmania amazonensis alters resistance to different stresses and mitochondrial bioenergetics. Mem. Inst. Oswaldo Cruz. 2016, 111, 460–468. [Google Scholar] [CrossRef] [PubMed]
- Searle, S.; Smith, D.F. Leishmania major: Characterisation and expression of a cytoplasmic stress-related protein. Exp. Parasitol. 1993, 77, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Ludewig, M.H.; Boshoff, A.; Horn, D.; Blatch, G.L. Trypanosoma brucei J protein 2 is a stress inducible and essential Hsp40. Int. J. Biochem. Cell Biol. 2015, 60, 93–98. [Google Scholar] [CrossRef]
- Burger, A.; Ludewig, M.L.; Boshoff, A. Investigating the chaperone properties of a novel heat shock protein, Hsp70.c, from Trypanosoma brucei. J. Parasitol. Res. 2014, 2014, 172582. [Google Scholar] [CrossRef] [PubMed]
- Fenn, K.; Matthews, K.R. The cell biology of Trypanosoma brucei differentiation. Curr. Opin. Microbiol. 2007, 10, 539–546. [Google Scholar] [CrossRef] [PubMed]
- Snoeckx, L.H.; Cornelussen, R.N.; Van Nieuwenhoven, F.A.; Reneman, R.S.; Van Der Vusse, G.J. Heat shock proteins and cardiovascular pathophysiology. Physiol. Rev. 2001, 81, 1461–1497. [Google Scholar] [CrossRef]
- Häusler, T.; Clayton, C. Post-transcriptional control of hsp70 mRNA in Trypanosoma brucei. Mol. Biochem. Parasitol. 1996, 76, 57–71. [Google Scholar] [CrossRef]
- Van der Ploeg, L.H.; Giannini, S.H.; Cantor, C.R. Heat shock genes: Regulatory role for differentiation in parasitic protozoa. Science 1985, 228, 1443–1446. [Google Scholar] [CrossRef]
- de Carvalho, E.F.; de Castro, F.T.; Rondinelli, E.; Soares, C.M.; Carvalho, J.F. HSP70 gene expression in Trypanosoma cruzi is regulated at different levels. J. Cell Physiol. 1990, 143, 439–444. [Google Scholar] [CrossRef]
- Vos, M.J.; Hageman, J.; Carra, S.; Kampinga, H.H. Structural and functional diversities between members of the human HSPB, HSPH, HSPA, and DNAJ chaperone families. Biochemistry 2008, 47, 7001–7011. [Google Scholar] [CrossRef]
- Kampinga, H.H.; Craig, E.A. The HSP70 chaperone machinery: J proteins as drivers of functional specificity. Nat. Rev. Mol. Cell Biol. 2010, 11, 579–592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dean, S.; Sunter, J.D.; Wheeler, R.J. TrypTag.org: A trypanosome genome-wide protein localisation resource. Trends Parasitol. 2017, 33, 80–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edkins, A.L.; Ludewig, M.H.; Blatch, G.L. A Trypanosoma cruzi heat shock protein 40 is able to stimulate the adenosine triphosphate hydrolysis activity of heat shock protein 70 and can substitute for a yeast heat shock protein 40. Int. J. Biochem. Cell Biol. 2004, 36, 1585–1598. [Google Scholar] [CrossRef] [PubMed]
- Zininga, T.; Pooe, O.J.; Makhado, P.B.; Ramatsui, L.; Prinsloo, E.; Achilonu, I.; Dirr, H.; Shonhai, A. Polymyxin B inhibits the chaperone activity of Plasmodium falciparum Hsp70. Cell Stress Chaperones 2017, 22, 707–715. [Google Scholar] [CrossRef] [Green Version]
- Cyr, D.M.; Ramos, C.H. Specification of Hsp70 function by Type I and Type II Hsp40. Subcell. Biochem. 2015, 78, 91–102. [Google Scholar]
- Freeman, B.C.; Morimoto, R.I. The human cytosolic molecular chaperones hsp90, hsp70 (hsc70) and hdj-1 have distinct roles in recognition of a non-native protein and protein refolding. EMBO J. 1996, 15, 2969–2979. [Google Scholar] [CrossRef]
- Johnson, B.D.; Schumacher, R.J.; Ross, E.D. Hop modulates Hsp70/Hsp90 interactions in protein folding. J. Biol. Chem. 1998, 273, 3679–3686. [Google Scholar] [CrossRef] [Green Version]
- Bangs, J.D.; Uyetake, L.; Brickman, M.; Balber, A.E.; Boothroyd, J.C. Molecular cloning and cellular localization of a BiP homologue in Trypanosoma brucei: Divergent ER retention signals in a lower eukaryote. J. Cell Sci. 1993, 105, 1101–1113. [Google Scholar]
- Bangs, J.D.; Brouch, E.M.; Ransom, D.M.; Roggy, J.L. A soluble secretory reporter system in Trypanosoma brucei: Studies on endoplasmic reticulum targeting. J. Biol. Chem. 1996, 271, 18387–18393. [Google Scholar] [CrossRef] [Green Version]
- Rotman, B. Regulation of enzymatic activity in the intact cell: The beta-D-galactosidase of Escherichia coli. J. Bacteriol. 1958, 76, 1–14. [Google Scholar]
Figure 1.
Expression and purification of recombinant TbHsp70, TbHsp70.4, and Tbj2. TbHsp70 and TbHsp70.4 were both expressed in E. coli XL1 Blue cells, and Tbj2 was expressed in E. coli BL21 (DE3) cells. SDS-PAGE (10%) and Western blot images representing the expression and purification of the recombinant forms of TbHsp70 (A), TbHsp70.4 (B), and Tbj2 (C). Lane M: Marker in kilodalton (kDa) (Precision Plus ProteinTM All Blue Prestained Protein Standard) is shown on the left-hand side; Lane C: The total extract for cells transformed with a neat pQE2 plasmid (TbHsp70 and TbHsp70.4) or transformed with a neat pET28a plasmid (Tbj2). Lane P: The total cell extract of E. coli XL1 Blue cells transformed with either pQE2/TbHsp70 or pQE2/TbHsp70.4, and E. coli BL21 (DE3) cells transformed with pET28a/Tbj2 prior to 1 mM IPTG induction; Lanes 1–16: Total cell lysate obtained 1–16 h post IPTG induction, respectively. Lanes E1–3: TbHsp70, TbHsp70.4, and Tbj2 proteins eluted from the affinity matrix using 500 mM imidazole. Lower panels: Western blot analysis using anti-His antibody to confirm expression and purification of recombinant TbHsp70, TbHsp70.4, and Tbj2. Western blot analysis using anti-TbHsp70 and anti-TbHsp70.4 to confirm expression of recombinant TbHsp70 and TbHsp70.4 were also used, respectively.
Figure 1.
Expression and purification of recombinant TbHsp70, TbHsp70.4, and Tbj2. TbHsp70 and TbHsp70.4 were both expressed in E. coli XL1 Blue cells, and Tbj2 was expressed in E. coli BL21 (DE3) cells. SDS-PAGE (10%) and Western blot images representing the expression and purification of the recombinant forms of TbHsp70 (A), TbHsp70.4 (B), and Tbj2 (C). Lane M: Marker in kilodalton (kDa) (Precision Plus ProteinTM All Blue Prestained Protein Standard) is shown on the left-hand side; Lane C: The total extract for cells transformed with a neat pQE2 plasmid (TbHsp70 and TbHsp70.4) or transformed with a neat pET28a plasmid (Tbj2). Lane P: The total cell extract of E. coli XL1 Blue cells transformed with either pQE2/TbHsp70 or pQE2/TbHsp70.4, and E. coli BL21 (DE3) cells transformed with pET28a/Tbj2 prior to 1 mM IPTG induction; Lanes 1–16: Total cell lysate obtained 1–16 h post IPTG induction, respectively. Lanes E1–3: TbHsp70, TbHsp70.4, and Tbj2 proteins eluted from the affinity matrix using 500 mM imidazole. Lower panels: Western blot analysis using anti-His antibody to confirm expression and purification of recombinant TbHsp70, TbHsp70.4, and Tbj2. Western blot analysis using anti-TbHsp70 and anti-TbHsp70.4 to confirm expression of recombinant TbHsp70 and TbHsp70.4 were also used, respectively.

Figure 2.
Expression of TbHsp70 and TbHsp70.4 in T. b. brucei parasites is modulated in response to heat stress at the blood stage. (A) (i) SDS-PAGE (10%) and (ii) Western blot analyses of the expression of TbHsp70 and TbHsp70.4 by T. b. brucei parasites cultured under normal (37 °C) and heat shock (42 °C) conditions, respectively; Lane M (molecular weight markers in kDa); parasite lysate harvested from bloodstream T. b. brucei 927 V221 cells in vitro at 37 °C and 42 °C (5 × 106 cells/lane), respectively. Lower panels: Detection of the protein expression levels of TbHsp70 and TbHsp70.4 by Western blot analysis using anti-TbHsp70 and anti-TbHsp70.4, respectively. Actin was used as a loading control to confirm that loading was equivalent in each lane. (B) Densitometric analysis illustrating the fold change in protein expression levels of the T. brucei Hsp70s in response to heat shock. A student’s t-test was used to validate the expression of the protein at 42 °C compared to 37 °C (p < 0.005). The experiment was performed in triplicate using three different whole cell lysates and the figure represents the findings of a typical experiment.
Figure 2.
Expression of TbHsp70 and TbHsp70.4 in T. b. brucei parasites is modulated in response to heat stress at the blood stage. (A) (i) SDS-PAGE (10%) and (ii) Western blot analyses of the expression of TbHsp70 and TbHsp70.4 by T. b. brucei parasites cultured under normal (37 °C) and heat shock (42 °C) conditions, respectively; Lane M (molecular weight markers in kDa); parasite lysate harvested from bloodstream T. b. brucei 927 V221 cells in vitro at 37 °C and 42 °C (5 × 106 cells/lane), respectively. Lower panels: Detection of the protein expression levels of TbHsp70 and TbHsp70.4 by Western blot analysis using anti-TbHsp70 and anti-TbHsp70.4, respectively. Actin was used as a loading control to confirm that loading was equivalent in each lane. (B) Densitometric analysis illustrating the fold change in protein expression levels of the T. brucei Hsp70s in response to heat shock. A student’s t-test was used to validate the expression of the protein at 42 °C compared to 37 °C (p < 0.005). The experiment was performed in triplicate using three different whole cell lysates and the figure represents the findings of a typical experiment.

Figure 3.
Stimulation of TbHsp70 and TbHsp70.4 ATPase activity by co-chaperone Tbj2. Submolar concentrations of recombinant Tbj2 were used to assess the ability of the co-chaperone to stimulate the basal ATPase activities of TbHsp70 and TbHsp70.4. The inorganic phosphate release was monitored by direct colorimetry at 595 nm wavelength. The curves represent the basal ATPase activities of recombinant TbHsp70 and TbHsp70.4, respectively, expressed as mean (±) SD. The assay was carried out in triplicate from three independent batches of TbHsp70 and TbHsp70.4. ATP hydrolysis by the Tbj2 was also analyzed separately and boiled samples of TbHsp70 and TbHsp70.4, indicated as [B], were included as negative controls. The ‘+’symbols represent components present in a reaction while ‘- ‘represents those that were absent. The averaged data from three independent experiments done in triplicate using three independent batches of purified proteins for each experiment are shown with error bars indicated on each bar to represent standard deviation. Statistically significant difference of the ATPase activity of the TbHsp70s alone relative to the ATPase activity of the TbHsp70s in the presence of Tbj2 are indicated by * (p < 0.05) above the reaction.
Figure 3.
Stimulation of TbHsp70 and TbHsp70.4 ATPase activity by co-chaperone Tbj2. Submolar concentrations of recombinant Tbj2 were used to assess the ability of the co-chaperone to stimulate the basal ATPase activities of TbHsp70 and TbHsp70.4. The inorganic phosphate release was monitored by direct colorimetry at 595 nm wavelength. The curves represent the basal ATPase activities of recombinant TbHsp70 and TbHsp70.4, respectively, expressed as mean (±) SD. The assay was carried out in triplicate from three independent batches of TbHsp70 and TbHsp70.4. ATP hydrolysis by the Tbj2 was also analyzed separately and boiled samples of TbHsp70 and TbHsp70.4, indicated as [B], were included as negative controls. The ‘+’symbols represent components present in a reaction while ‘- ‘represents those that were absent. The averaged data from three independent experiments done in triplicate using three independent batches of purified proteins for each experiment are shown with error bars indicated on each bar to represent standard deviation. Statistically significant difference of the ATPase activity of the TbHsp70s alone relative to the ATPase activity of the TbHsp70s in the presence of Tbj2 are indicated by * (p < 0.05) above the reaction.

Figure 4.
Tbj2 enhances the aggregation suppression activity of TbHsp70 and TbHsp70.4. (A) The ability of Tbj2, at varying concentrations, to bind malate dehydrogenase (MDH) was monitored during the heat-induced aggregation of MDH (0.72 µM) in vitro at 48 °C and by quantifying the pellet (insoluble; grey bars) and supernatant (soluble; black bars) fractions after heat exposure. (B) The ability of TbHsp70 and TbHsp70.4 in the presence and absence of Tbj2 to suppress the heat-induced aggregation of MDH (0.72 µM) in vitro at 48 °C was assessed by quantifying the pellet (insoluble; grey bars) and supernatant (soluble; black bars) fractions after heat exposure. Suppression of the thermal aggregation of MDH by 0.25 µM TbHsp70 and TbHsp70.4 is enhanced by the addition of varying concentrations of Tbj2. Tbj2, TbHsp70.4, and TbHsp70 in the absence of MDH were not prone to aggregation under the assay conditions. Standard deviations were obtained from three replicate assays on three independent batches of purified protein. A statistically significant difference between a reaction and MDH alone are indicated by * (p < 0.05) above the bar using a Student’s t-test.
Figure 4.
Tbj2 enhances the aggregation suppression activity of TbHsp70 and TbHsp70.4. (A) The ability of Tbj2, at varying concentrations, to bind malate dehydrogenase (MDH) was monitored during the heat-induced aggregation of MDH (0.72 µM) in vitro at 48 °C and by quantifying the pellet (insoluble; grey bars) and supernatant (soluble; black bars) fractions after heat exposure. (B) The ability of TbHsp70 and TbHsp70.4 in the presence and absence of Tbj2 to suppress the heat-induced aggregation of MDH (0.72 µM) in vitro at 48 °C was assessed by quantifying the pellet (insoluble; grey bars) and supernatant (soluble; black bars) fractions after heat exposure. Suppression of the thermal aggregation of MDH by 0.25 µM TbHsp70 and TbHsp70.4 is enhanced by the addition of varying concentrations of Tbj2. Tbj2, TbHsp70.4, and TbHsp70 in the absence of MDH were not prone to aggregation under the assay conditions. Standard deviations were obtained from three replicate assays on three independent batches of purified protein. A statistically significant difference between a reaction and MDH alone are indicated by * (p < 0.05) above the bar using a Student’s t-test.

Figure 5.
Tbj2 stimulates the refoldase activity of TbHsp70 and TbHsp70.4. Chemically-denatured β-galactosidase (3.4 nM) was diluted into refolding buffer containing the indicated Hsp70 and/or Tbj2 mixtures. β-galactosidase assays were performed as previously described [37]. Recovery of activity of denatured β-galactosidase by the various molecular chaperone mixtures was measured by its ability to convert substrate o-nitrophenyl-β-D-galactoside (ONPG) to yellow ortho-nitrophenol (ONP), and this absorbance was recorded at 420 nm. The negative control consisted of BSA (1.6 µM) added to denatured β-galactosidase enzyme to determine the kinetics of spontaneous refolding of the denatured β-galactosidase enzyme. Results are expressed as % β-galactosidase activity of the refolded enzyme in relation to native enzyme. Standard deviations were obtained from three replicate assays on three independent batches of purified protein.
Figure 5.
Tbj2 stimulates the refoldase activity of TbHsp70 and TbHsp70.4. Chemically-denatured β-galactosidase (3.4 nM) was diluted into refolding buffer containing the indicated Hsp70 and/or Tbj2 mixtures. β-galactosidase assays were performed as previously described [37]. Recovery of activity of denatured β-galactosidase by the various molecular chaperone mixtures was measured by its ability to convert substrate o-nitrophenyl-β-D-galactoside (ONPG) to yellow ortho-nitrophenol (ONP), and this absorbance was recorded at 420 nm. The negative control consisted of BSA (1.6 µM) added to denatured β-galactosidase enzyme to determine the kinetics of spontaneous refolding of the denatured β-galactosidase enzyme. Results are expressed as % β-galactosidase activity of the refolded enzyme in relation to native enzyme. Standard deviations were obtained from three replicate assays on three independent batches of purified protein.

© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Bentley, S.J.; Boshoff, A. Trypanosoma brucei J-Protein 2 Functionally Co-Operates with the Cytosolic Hsp70 and Hsp70.4 Proteins. Int. J. Mol. Sci. 2019, 20, 5843. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20235843
AMA Style
Bentley SJ, Boshoff A. Trypanosoma brucei J-Protein 2 Functionally Co-Operates with the Cytosolic Hsp70 and Hsp70.4 Proteins. International Journal of Molecular Sciences. 2019; 20(23):5843. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20235843
Chicago/Turabian StyleBentley, Stephen John, and Aileen Boshoff. 2019. "Trypanosoma brucei J-Protein 2 Functionally Co-Operates with the Cytosolic Hsp70 and Hsp70.4 Proteins" International Journal of Molecular Sciences 20, no. 23: 5843. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20235843
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.