A Paradigm in Immunochemistry, Revealed by Monoclonal Antibodies to Spatially Distinct Epitopes on Syntenin-1

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
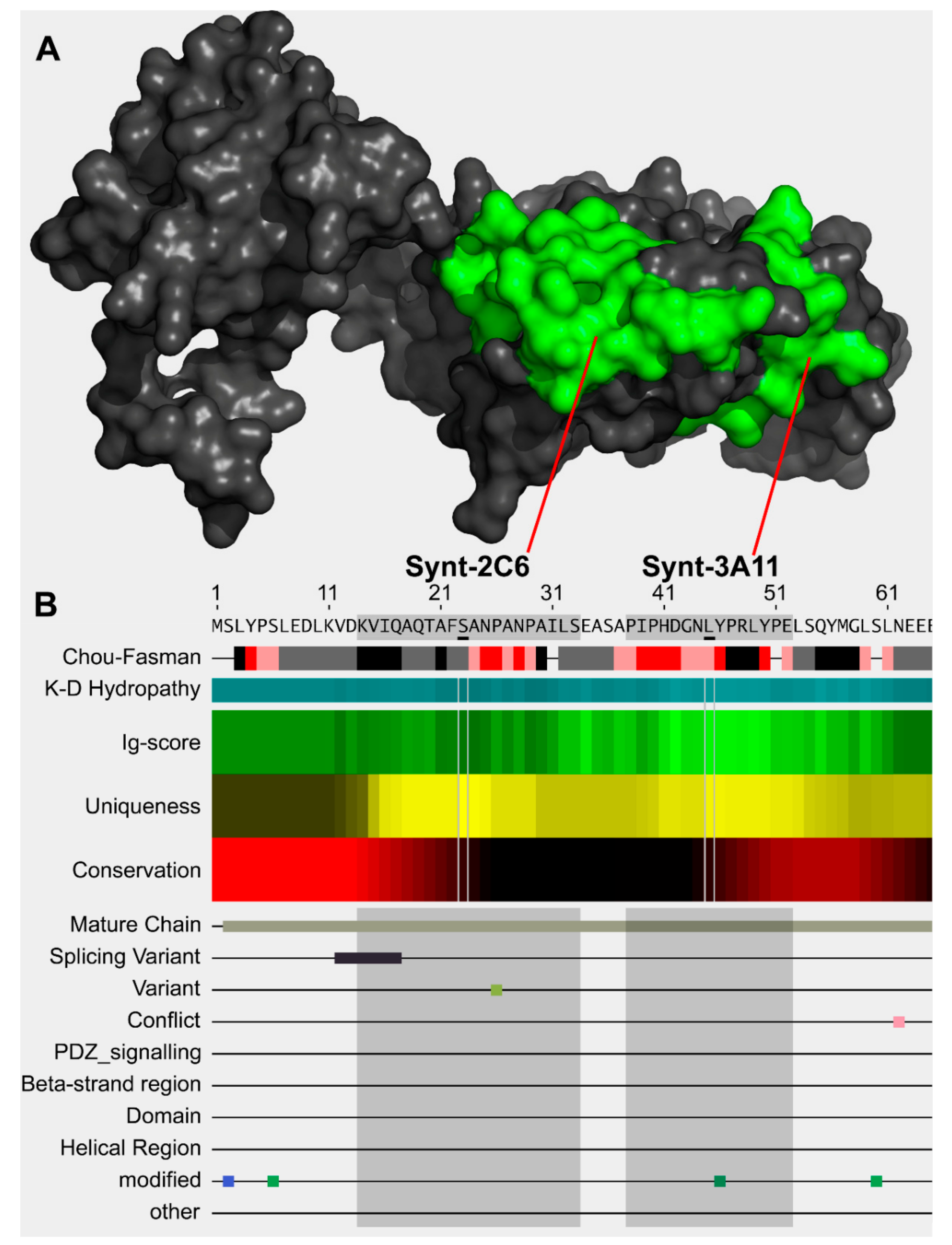
2.1. Epitope Mapping of Commercial Antibody
2.2. Antibody Design
2.3. Commercial Antibody Reagents
2.4. Cell Lines and Culture Conditions
2.5. Preparation of Cell Extracts and Conditioned Culture Media for Protein Detection
2.6. Western Blotting
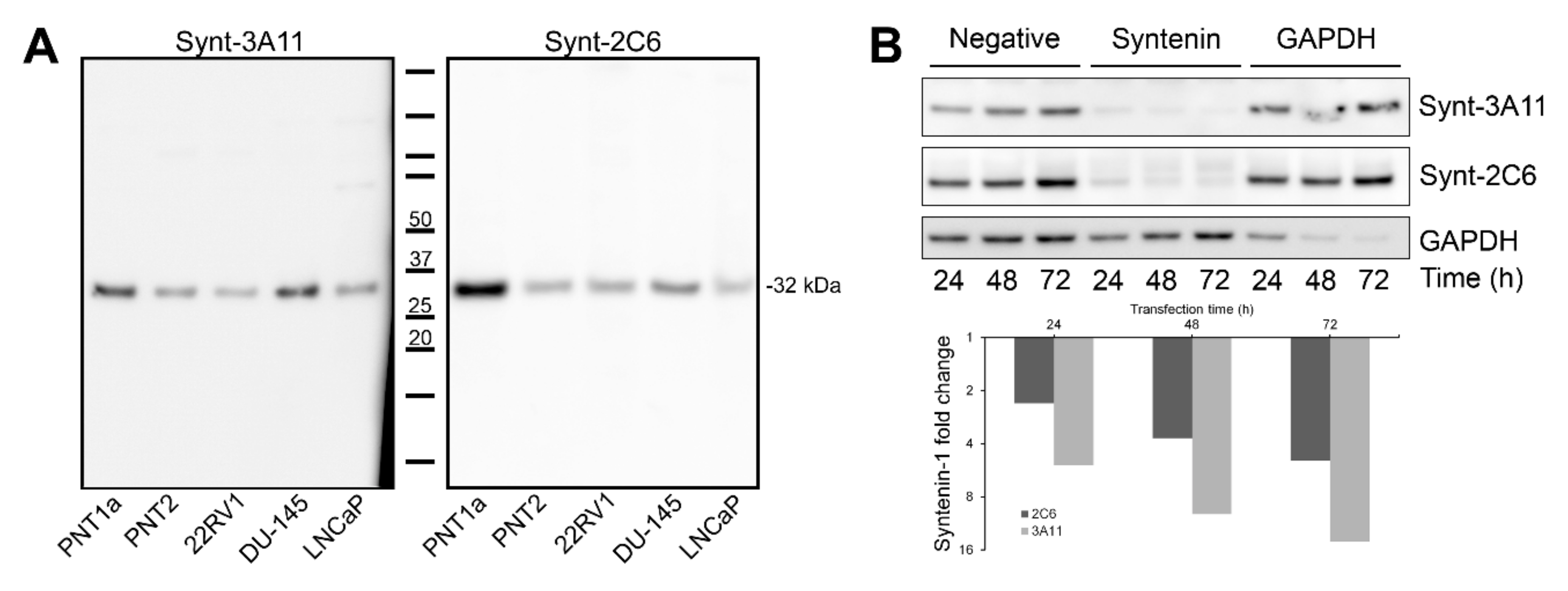
2.7. siRNA Knock-Down
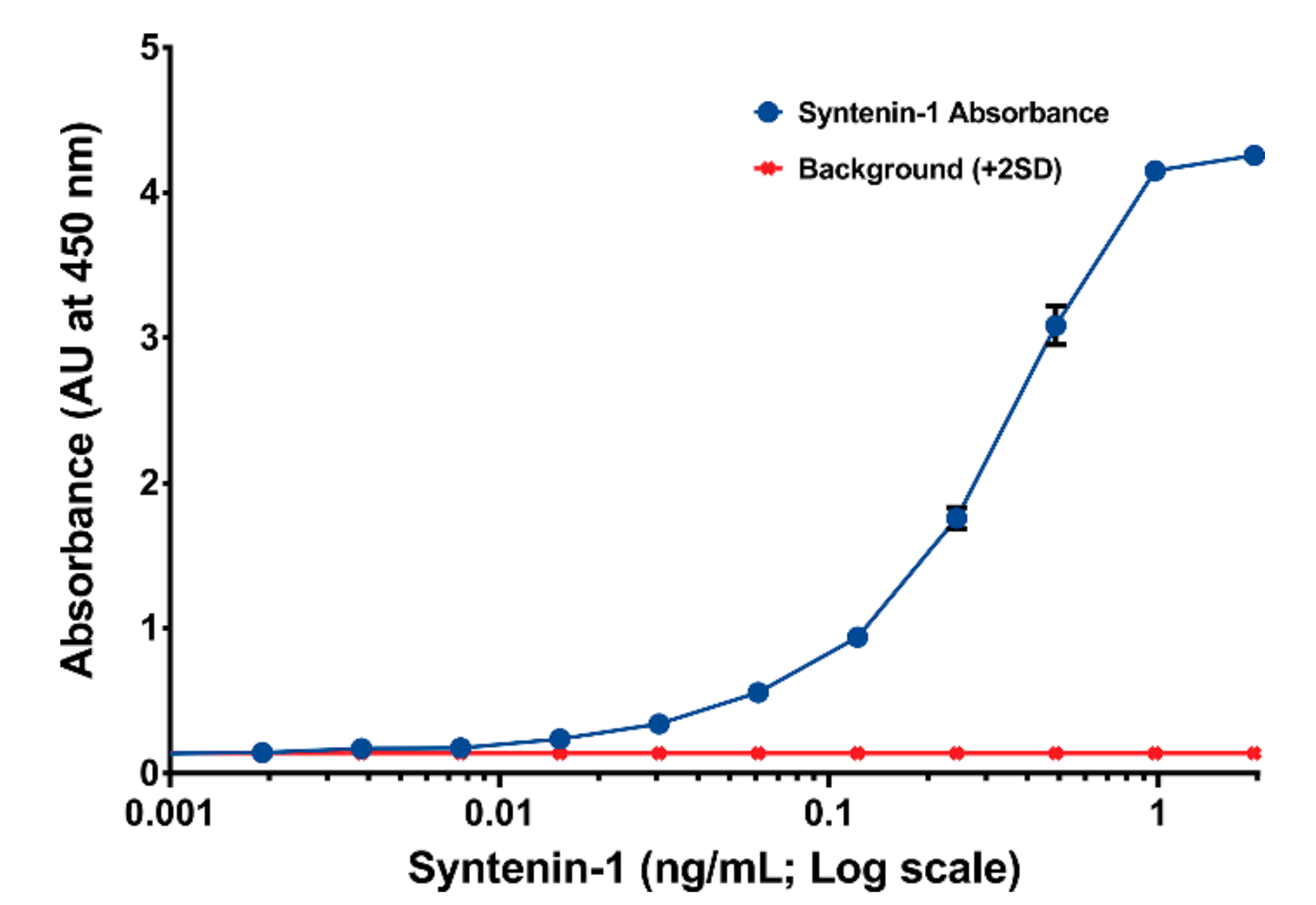
2.8. ELISA Sandwich Assay
2.9. Immunofluorescence
2.10. Nocodazole Treatment
2.11. Immunohistochemistry
2.12. Ethics
2.13. Data Availability
3. Results
3.1. Significant Epitope Cross-Reactivity for Commercial Antibodies to Syntenin-1
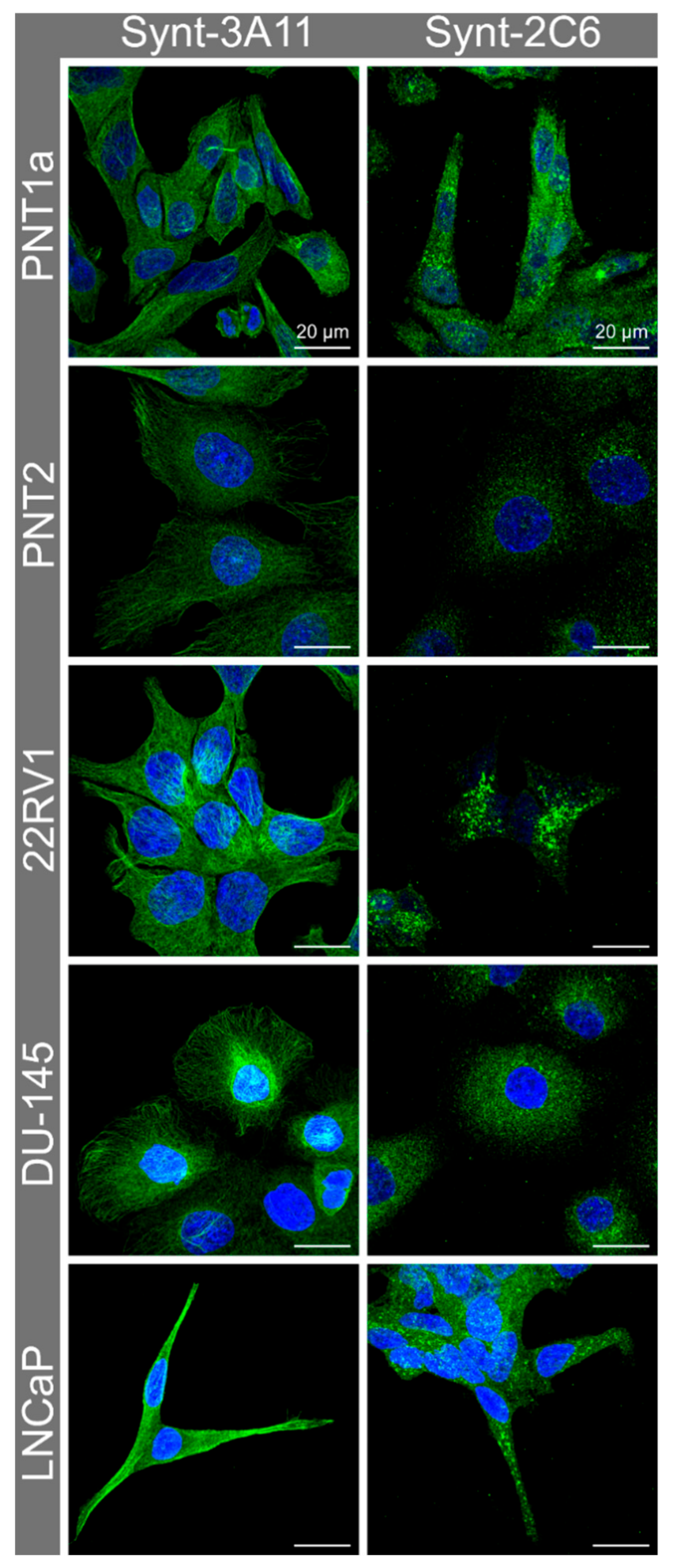
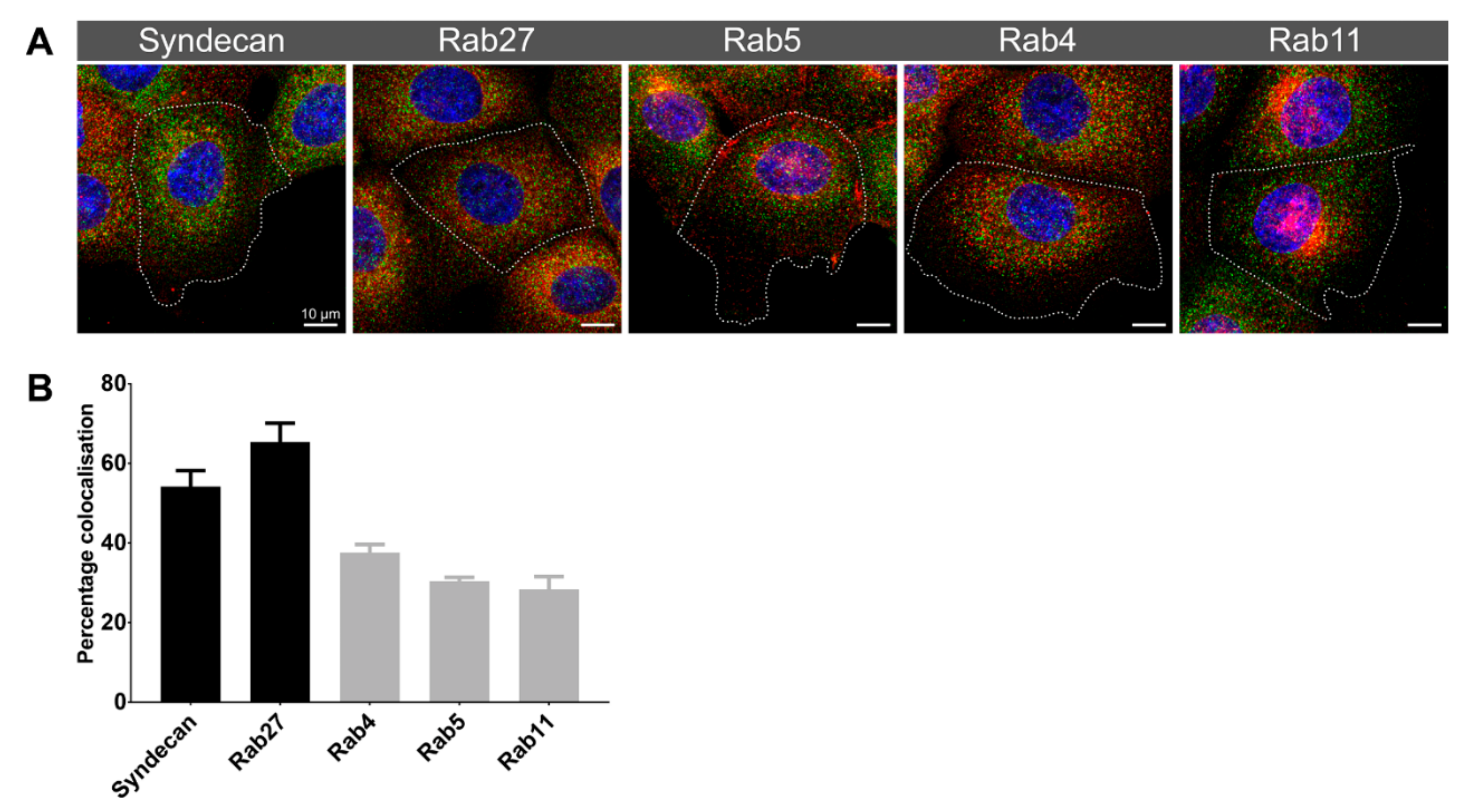
3.2. Immune Fluorescence Demonstrated a Different Cellular Distribution for the Two Syntenin-1 Mono-Specific Monoclonal Antibodies in Prostate Cells
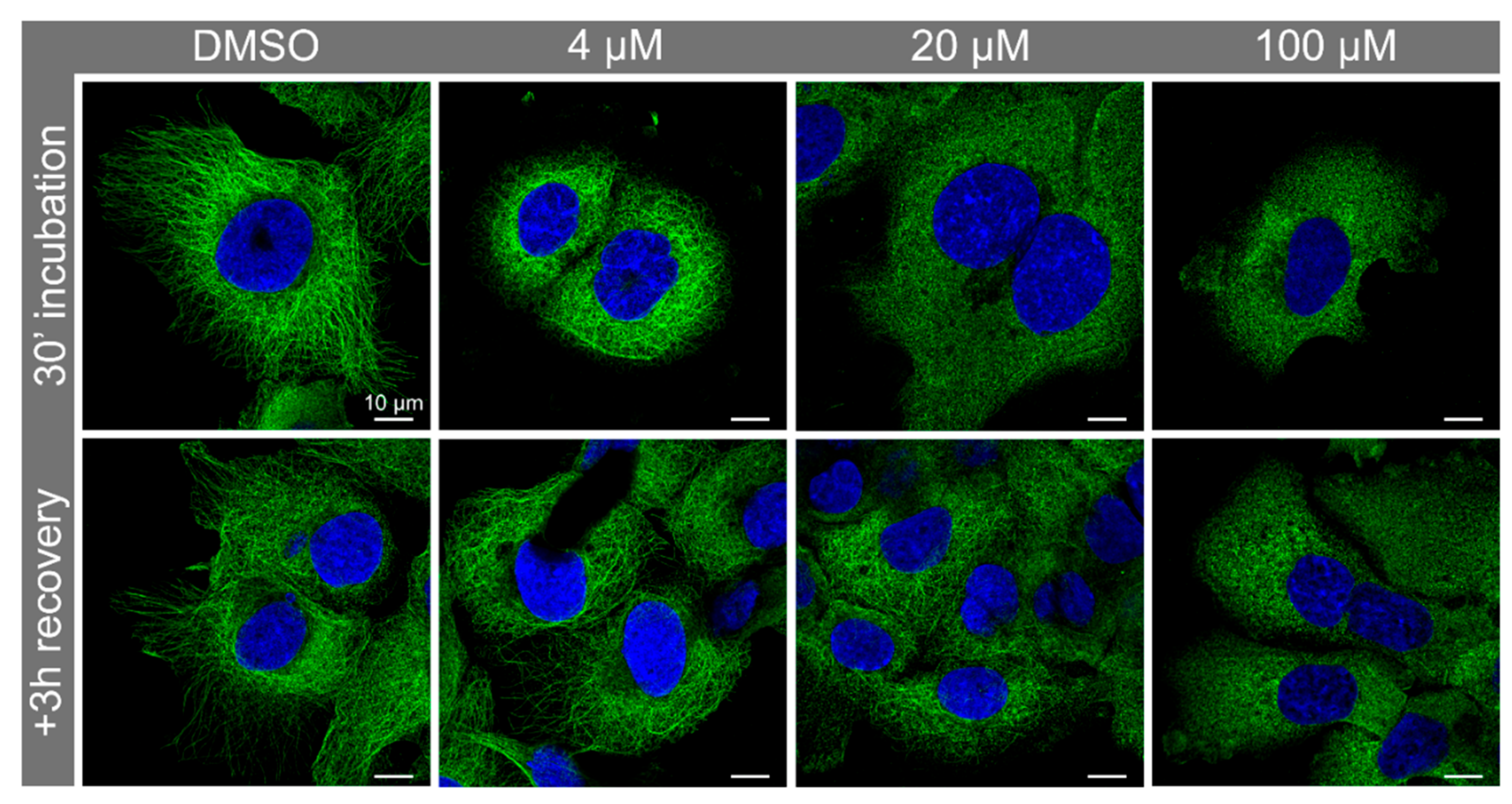
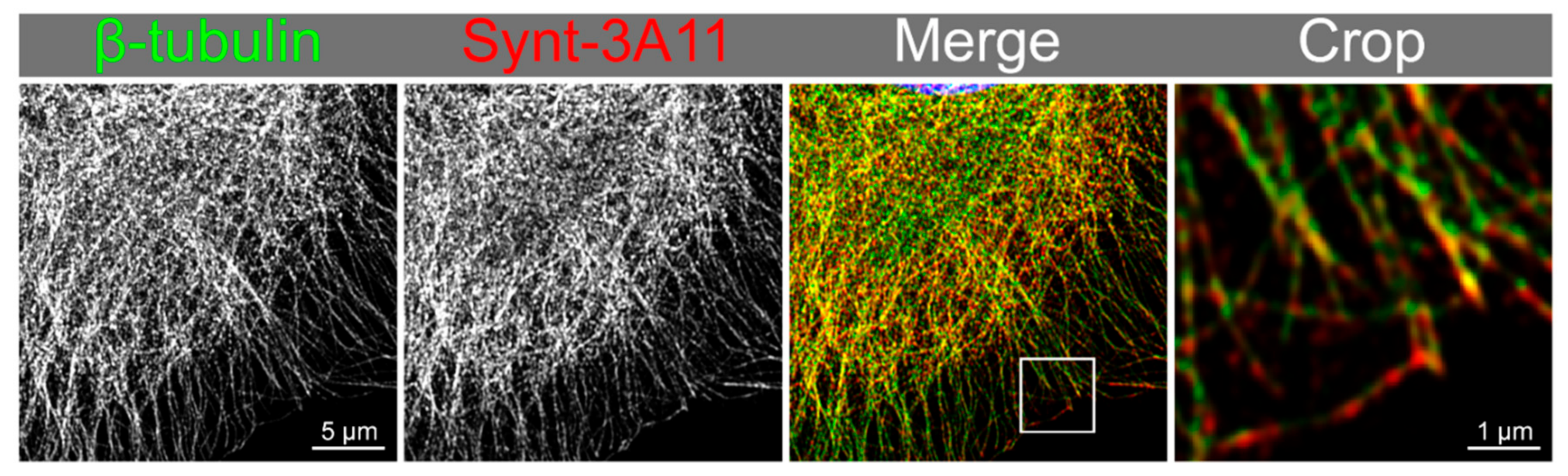
3.3. Nocodazole Treatment Confirmed that Syntenin-1 Can Associate with Microtubules
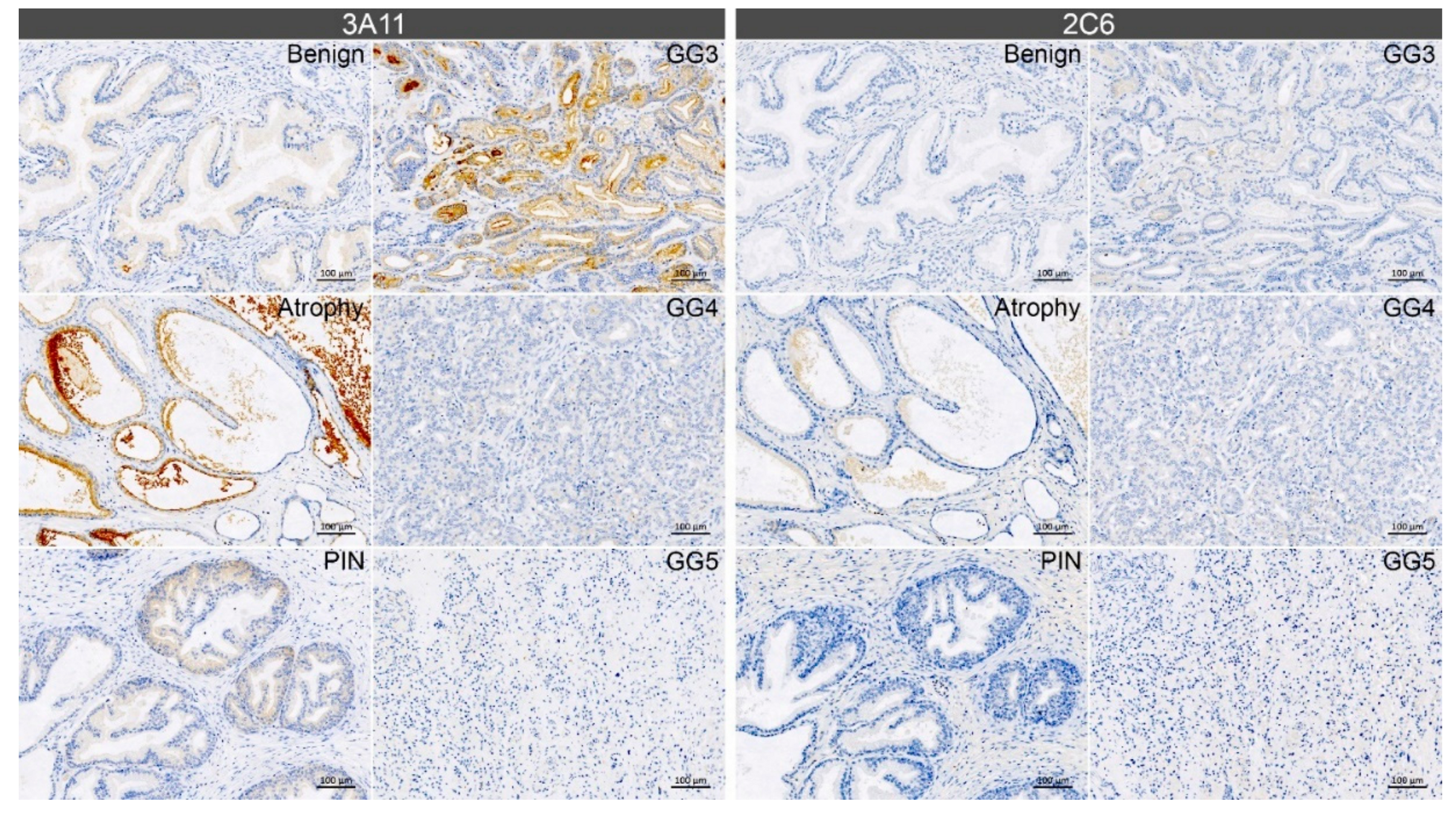
3.4. Immunohistochemistry with Synt-3A11 and Synt-2C6 Monoclonal Antibodies on Prostate Tissue
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lin, J.J.; Jiang, H.; Fisher, P.B. Melanoma differentiation associated gene-9, mda-9, is a human gamma interferon responsive gene. Gene 1998, 207, 105–110. [Google Scholar] [CrossRef]
- Philley, J.V.; Kannan, A.; Dasgupta, S. MDA-9/Syntenin Control. J. Cell Physiol. 2016, 231, 545–550. [Google Scholar] [CrossRef] [PubMed]
- Friand, V.; David, G.; Zimmermann, P. Syntenin and syndecan in the biogenesis of exosomes. Biol. Cell. 2015, 107, 331–341. [Google Scholar] [CrossRef] [PubMed]
- Johnson, I.R.; Parkinson-Lawrence, E.J.; Butler, L.M.; Brooks, D.A. Prostate cell lines as models for biomarker discovery: Performance of current markers and the search for new biomarkers. The Prostate 2014, 74, 547–560. [Google Scholar] [CrossRef]
- Johnson, I.R.; Parkinson-Lawrence, E.J.; Keegan, H.; Spillane, C.D.; Barry-O’Crowley, J.; Watson, W.R.; Selemidis, S.; Butler, L.M.; O’Leary, J.J.; Brooks, D.A. Endosomal gene expression: a new indicator for prostate cancer patient prognosis? Oncotarget 2015, 6, 37919–37929. [Google Scholar] [CrossRef]
- Johnson, I.R.; Parkinson-Lawrence, E.J.; Shandala, T.; Weigert, R.; Butler, L.M.; Brooks, D.A. Altered endosome biogenesis in prostate cancer has biomarker potential. Mol. cancer Res.: MCR 2014, 12, 1851–1862. [Google Scholar] [CrossRef]
- Kashyap, R.; Roucourt, B.; Lembo, F.; Fares, J.; Carcavilla, A.M.; Restouin, A.; Zimmermann, P.; Ghossoub, R. Syntenin controls migration, growth, proliferation, and cell cycle progression in cancer cells. Front. Pharmacol. 2015, 6, 241. [Google Scholar] [CrossRef]
- Cui, L.; Cheng, S.; Liu, X.; Messadi, D.; Yang, Y.; Hu, S. Syntenin-1 is a promoter and prognostic marker of head and neck squamous cell carcinoma invasion and metastasis. Oncotarget 2016, 7, 82634–82647. [Google Scholar] [CrossRef]
- Zhong, D.; Ran, J.; Zhang, X.; Tan, Y.; Chen, G.; Tang, W.; Li, X.; Wang, B. Syntenin is expressed in human gliomas and may correlate with tumor migration. Arch. Med. Sci. 2015, 11, 1303–1307. [Google Scholar] [CrossRef]
- Yang, Y.; Hong, Q.; Shi, P.; Liu, Z.; Luo, J.; Shao, Z. Elevated expression of syntenin in breast cancer is correlated with lymph node metastasis and poor patient survival. Breast Cancer Res. 2013, 15, R50. [Google Scholar] [CrossRef]
- Baietti, M.F.; Zhang, Z.; Mortier, E.; Melchior, A.; Degeest, G.; Geeraerts, A.; Ivarsson, Y.; Depoortere, F.; Coomans, C.; Vermeiren, E.; et al. Syndecan-syntenin-ALIX regulates the biogenesis of exosomes. Nat. Cell. Biol. 2012, 14, 677–685. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, P.; Tomatis, D.; Rosas, M.; Grootjans, J.; Leenaerts, I.; Degeest, G.; Reekmans, G.; Coomans, C.; David, G. Characterization of syntenin, a syndecan-binding PDZ protein, as a component of cell adhesion sites and microfilaments. Mol. Biol. Cell. 2001, 12, 339–350. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.K.; Pan, S.H.; Chang, Y.L.; Hung, P.F.; Kao, S.H.; Wang, W.L.; Lin, C.W.; Yang, S.C.; Liang, C.H.; Wu, C.T.; et al. MDA-9/Syntenin-Slug transcriptional complex promote epithelial-mesenchymal transition and invasion/metastasis in lung adenocarcinoma. Oncotarget 2016, 7, 386–401. [Google Scholar] [CrossRef] [PubMed]
- Egea-Jimenez, A.L.; Gallardo, R.; Garcia-Pino, A.; Ivarsson, Y.; Wawrzyniak, A.M.; Kashyap, R.; Loris, R.; Schymkowitz, J.; Rousseau, F.; Zimmermann, P. Frizzled 7 and PIP2 binding by syntenin PDZ2 domain supports Frizzled 7 trafficking and signalling. Nat. Commun. 2016, 7, 12101. [Google Scholar] [CrossRef]
- Tae, N.; Lee, S.; Kim, O.; Park, J.; Na, S.; Lee, J.H. Syntenin promotes VEGF-induced VEGFR2 endocytosis and angiogenesis by increasing ephrin-B2 function in endothelial cells. Oncotarget 2017, 8, 38886–38901. [Google Scholar] [CrossRef]
- Zimmermann, P.; Zhang, Z.; Degeest, G.; Mortier, E.; Leenaerts, I.; Coomans, C.; Schulz, J.; N’Kuli, F.; Courtoy, P.J.; David, G. Syndecan recycling [corrected] is controlled by syntenin-PIP2 interaction and Arf6. Dev. Cell. 2005, 9, 377–388. [Google Scholar] [CrossRef]
- Ghossoub, R.; Lembo, F.; Rubio, A.; Gaillard, C.B.; Bouchet, J.; Vitale, N.; Slavik, J.; Machala, M.; Zimmermann, P. Syntenin-ALIX exosome biogenesis and budding into multivesicular bodies are controlled by ARF6 and PLD2. Nat. Commun. 2014, 5, 3477. [Google Scholar] [CrossRef]
- Gangemi, R.; Mirisola, V.; Barisione, G.; Fabbi, M.; Brizzolara, A.; Lanza, F.; Mosci, C.; Salvi, S.; Gualco, M.; Truini, M.; et al. Mda-9/syntenin is expressed in uveal melanoma and correlates with metastatic progression. PLoS ONE 2012, 7, e29989. [Google Scholar] [CrossRef]
- Boukerche, H.; Aissaoui, H.; Prevost, C.; Hirbec, H.; Das, S.K.; Su, Z.Z.; Sarkar, D.; Fisher, P.B. Src kinase activation is mandatory for MDA-9/syntenin-mediated activation of nuclear factor-kappaB. Oncogene 2010, 29, 3054–3066. [Google Scholar] [CrossRef]
- Tudor, C.; te Riet, J.; Eich, C.; Harkes, R.; Smisdom, N.; Bouhuijzen Wenger, J.; Ameloot, M.; Holt, M.; Kanger, J.S.; Figdor, C.G.; et al. Syntenin-1 and ezrin proteins link activated leukocyte cell adhesion molecule to the actin cytoskeleton. J. Biol. Chem. 2014, 289, 13445–13460. [Google Scholar] [CrossRef]
- Simpson, K.; Browning, M. Antibodies That Work Again and Again and Again. Methods Mol. Biol. 2017, 1554, 41–59. [Google Scholar] [CrossRef] [PubMed]
- Ellinger, I.; Chatuphonprasert, W.; Reiter, M.; Voss, A.; Kemper, J.; Straka, E.; Scheinast, M.; Zeisler, H.; Salzer, H.; Gundacker, C. Don’t trust an(t)ybody - Pitfalls during investigation of candidate proteins for methylmercury transport at the placental interface. Placenta 2016, 43, 13–16. [Google Scholar] [CrossRef] [PubMed]
- Couchman, J.R. Commercial antibodies: the good, bad, and really ugly. J. Histochem. Cytochem. 2009, 57, 7–8. [Google Scholar] [CrossRef] [PubMed]
- Pisitkun, T.; Hoffert, J.D.; Saeed, F.; Knepper, M.A. NHLBI-AbDesigner: an online tool for design of peptide-directed antibodies. Am. J. Physiol. Cell Physiol. 2012, 302, C154–C164. [Google Scholar] [CrossRef]
- Kelley, L.A.; Mezulis, S.; Yates, C.M.; Wass, M.N.; Sternberg, M.J.E. The Phyre2 web portal for protein modeling, prediction and analysis. Nat. Protoc. 2015, 10, 845–858. [Google Scholar] [CrossRef]
- Grootjans, J.J.; Zimmermann, P.; Reekmans, G.; Smets, A.; Degeest, G.; Durr, J.; David, G. Syntenin, a PDZ protein that binds syndecan cytoplasmic domains. Proc. Natl. Acad Sci. USA 1997, 94, 13683–13688. [Google Scholar] [CrossRef]
- Benjamin, D.C.; Perdue, S.S. Site-Directed Mutagenesis in Epitope Mapping. Methods 1996, 9, 508–515. [Google Scholar] [CrossRef]
- Dougan, D.A.; Malby, R.L.; Gruen, L.C.; Kortt, A.A.; Hudson, P.J. Effects of substitutions in the binding surface of an antibody on antigen affinity. Protein Eng. 1998, 11, 65–74. [Google Scholar] [CrossRef]
- Bradbury, A.; Pluckthun, A. Reproducibility: Standardize antibodies used in research. Nature 2015, 518, 27–29. [Google Scholar] [CrossRef]
- Andersson, S.; Sundberg, M.; Pristovsek, N.; Ibrahim, A.; Jonsson, P.; Katona, B.; Clausson, C.M.; Zieba, A.; Ramstrom, M.; Soderberg, O.; et al. Insufficient antibody validation challenges oestrogen receptor beta research. Nat. Commun. 2017, 8, 15840. [Google Scholar] [CrossRef]
- Sala-Valdes, M.; Gordon-Alonso, M.; Tejera, E.; Ibanez, A.; Cabrero, J.R.; Ursa, A.; Mittelbrunn, M.; Lozano, F.; Sanchez-Madrid, F.; Yanez-Mo, M. Association of syntenin-1 with M-RIP polarizes Rac-1 activation during chemotaxis and immune interactions. J. Cell. Sci. 2012, 125, 1235–1246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.J. Dynamic regulation of the microtubule and actin cytoskeleton in zebrafish epiboly. Biochem. Biophys Res. Commun. 2014, 452, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Brockstedt, U.; Dobra, K.; Nurminen, M.; Hjerpe, A. Immunoreactivity to cell surface syndecans in cytoplasm and nucleus: tubulin-dependent rearrangements. Exp. Cell Res. 2002, 274, 235–245. [Google Scholar] [CrossRef] [PubMed]
- Johnson, T.R.; Khandrika, L.; Kumar, B.; Venezia, S.; Koul, S.; Chandhoke, R.; Maroni, P.; Donohue, R.; Meacham, R.B.; Koul, H.K. Focal adhesion kinase controls aggressive phenotype of androgen-independent prostate cancer. Mol. Cancer Res. 2008, 6, 1639–1648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwangbo, C.; Kim, J.; Lee, J.J.; Lee, J.H. Activation of the integrin effector kinase focal adhesion kinase in cancer cells is regulated by crosstalk between protein kinase Calpha and the PDZ adapter protein mda-9/Syntenin. Cancer Res. 2010, 70, 1645–1655. [Google Scholar] [CrossRef] [Green Version]
- DeRita, R.M.; Zerlanko, B.; Singh, A.; Lu, H.; Iozzo, R.V.; Benovic, J.L.; Languino, L.R. c-Src, Insulin-Like Growth Factor I Receptor, G-Protein-Coupled Receptor Kinases and Focal Adhesion Kinase are Enriched Into Prostate Cancer Cell Exosomes. J. Cell Biochem. 2017, 118, 66–73. [Google Scholar] [CrossRef] [Green Version]
- Michel, S.; Deleage, G.; Charrier, J.P.; Passagot, J.; Battail-Poirot, N.; Sibai, G.; Jolivet, M.; Jolivet-Reynaud, C. Anti-free prostate-specific antigen monoclonal antibody epitopes defined by mimotopes and molecular modeling. Clin. Chem. 1999, 45, 638–650. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clone | Epitope | aa Range | Cross-Reactivity ≥ 6 aa |
|---|---|---|---|
| Synt-2C6 | KVIQAQTAFSANPANPAILS | 14–33 | 0 |
| Synt-3A11 | PIPHDGNLYPRLYPE | 38–52 | 0 |
| Sequence Range | Matches ≥ 6 aa | Potential Cross-Reactivity | |
|---|---|---|---|
| ab154940 | 1–45 | 37 | ANDR, CE042, CENPM, CF211, CLIP2, CTL1, DESP, EPHX4, FNTB, FOXO4, GLSK, HERC5, HERC6, KAT2A, KC1A, KC1AL, KC1D, KC1E, KC1G1, KC1G2, KC1G3, LUZP1, M3K2, MOSC1, NPHP4, NTAL, PO6F1, Q71TU5, Q8IWC0, Q9BZG5, SDCB2, SLN13, TFDP2, TITIN, TTC28, UBE3C, ZCC18 |
| ab53552 | 6–19 | 11 | CF211, CLIP2, DESP, FNTB, HERC5, HERC6, LUZP1, SDCB2, TFDP2, TITIN, UBE3C |
| ab62530 | 109–158 | 29 | AKA11, B3A3, CAN7, CG063, CK093, CNTP1, DYH3, DYH7, DYST, EAA3, FAT1, GEMI4, HAUS6, MACF1, MACF4, MTCH1, MYO15, PAPL, PDE12, Q5TDC2, Q5VY60, RGS22, SC16A, SDCB2, SV2A, TBCD, VEGFC, VP13B, WDR35 |
| Epitope Reactivity | Range | Matches ≥6 aa | Protein Cross-Reactivity |
|---|---|---|---|
| HDGNLYPRLYPELSQYMGLS | 41–60 | 4 | CQ047, DYST, EPG5, SDCB2 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Johnson, I.R.D.; Sorvina, A.; Logan, J.M.; Moore, C.R.; Heatlie, J.K.; Parkinson-Lawrence, E.J.; Selemidis, S.; O’Leary, J.J.; Butler, L.M.; Brooks, D.A. A Paradigm in Immunochemistry, Revealed by Monoclonal Antibodies to Spatially Distinct Epitopes on Syntenin-1. Int. J. Mol. Sci. 2019, 20, 6035. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20236035
Johnson IRD, Sorvina A, Logan JM, Moore CR, Heatlie JK, Parkinson-Lawrence EJ, Selemidis S, O’Leary JJ, Butler LM, Brooks DA. A Paradigm in Immunochemistry, Revealed by Monoclonal Antibodies to Spatially Distinct Epitopes on Syntenin-1. International Journal of Molecular Sciences. 2019; 20(23):6035. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20236035
Chicago/Turabian StyleJohnson, Ian R. D., Alexandra Sorvina, Jessica M. Logan, Courtney R. Moore, Jessica K. Heatlie, Emma J. Parkinson-Lawrence, Stavros Selemidis, John J. O’Leary, Lisa M. Butler, and Douglas A. Brooks. 2019. "A Paradigm in Immunochemistry, Revealed by Monoclonal Antibodies to Spatially Distinct Epitopes on Syntenin-1" International Journal of Molecular Sciences 20, no. 23: 6035. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20236035