Effect of AQP Inhibition on Boar Sperm Cryotolerance Depends on the Intrinsic Freezability of the Ejaculate

 ,
,  ,
,  ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Classification of Boar Ejaculates in GFE and PFE Groups
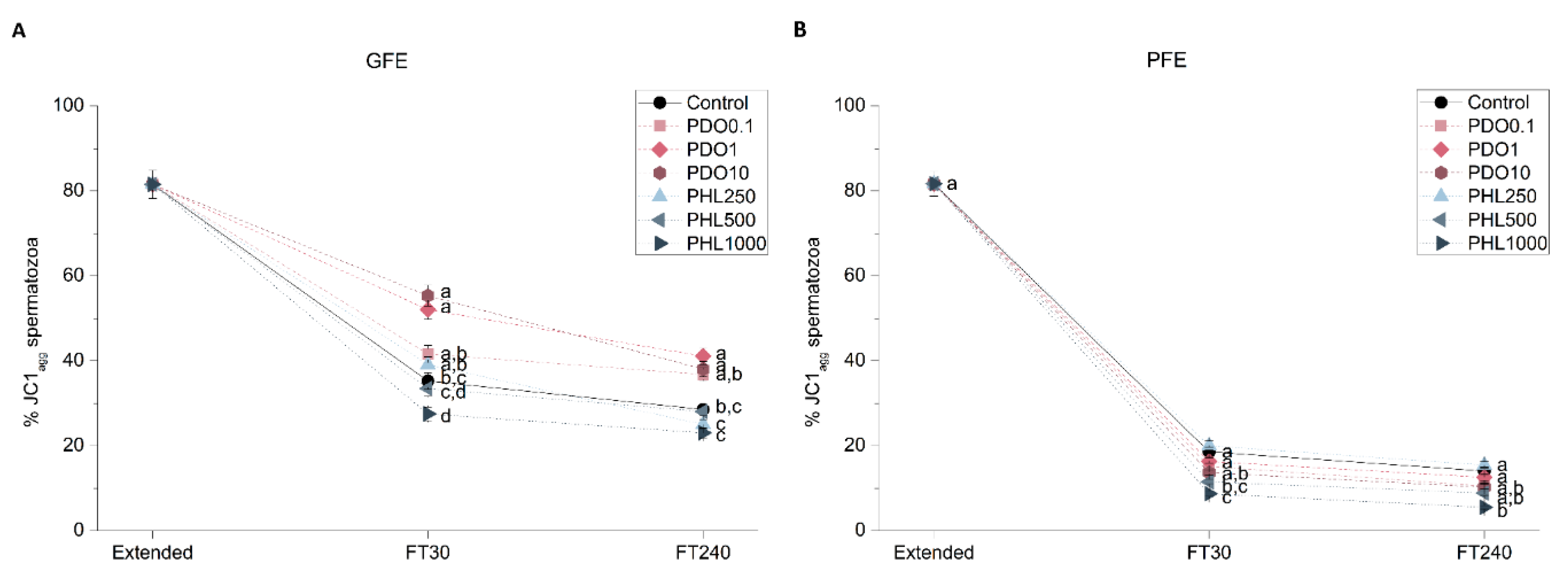
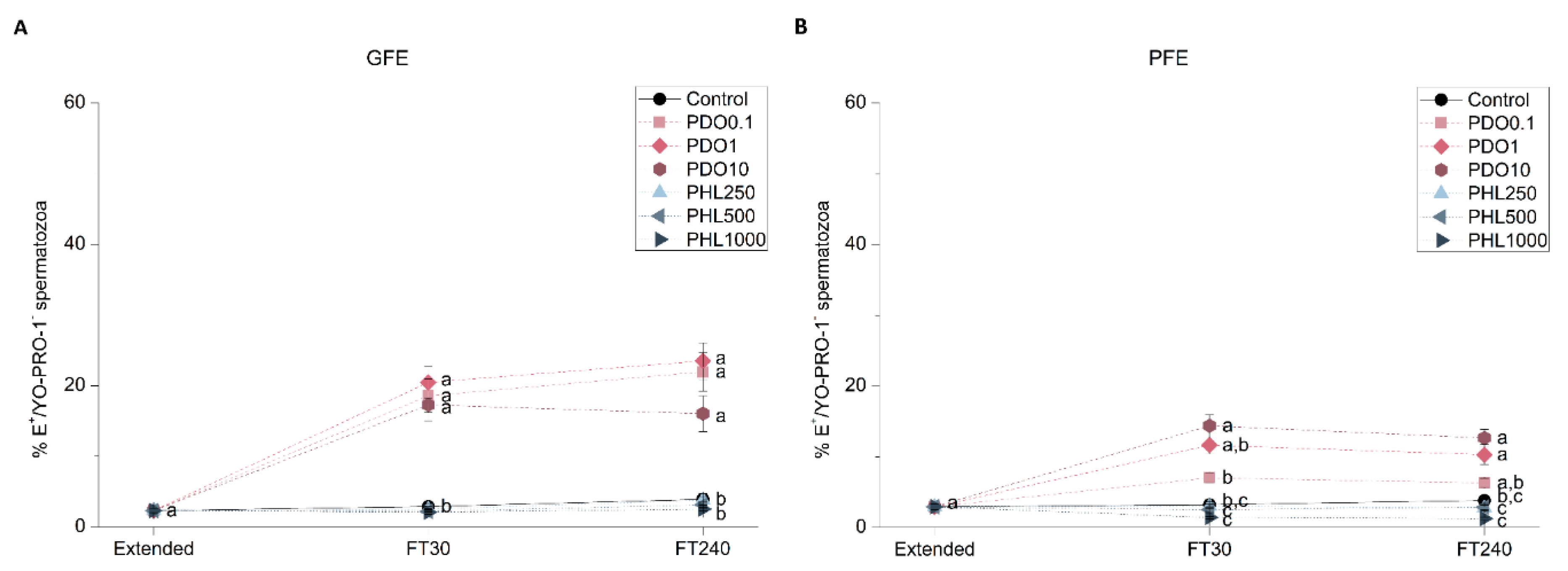
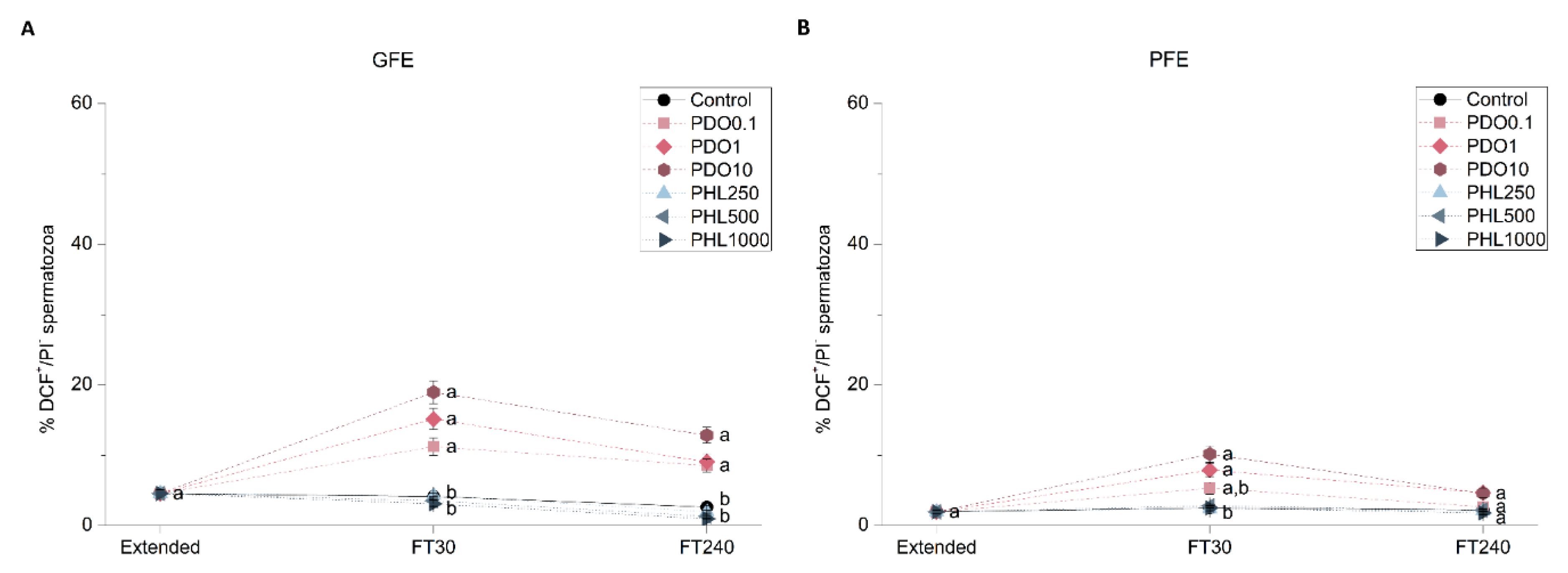
2.2. Effects of AQP Inhibition on Cryopreserved Sperm Quality Parameters
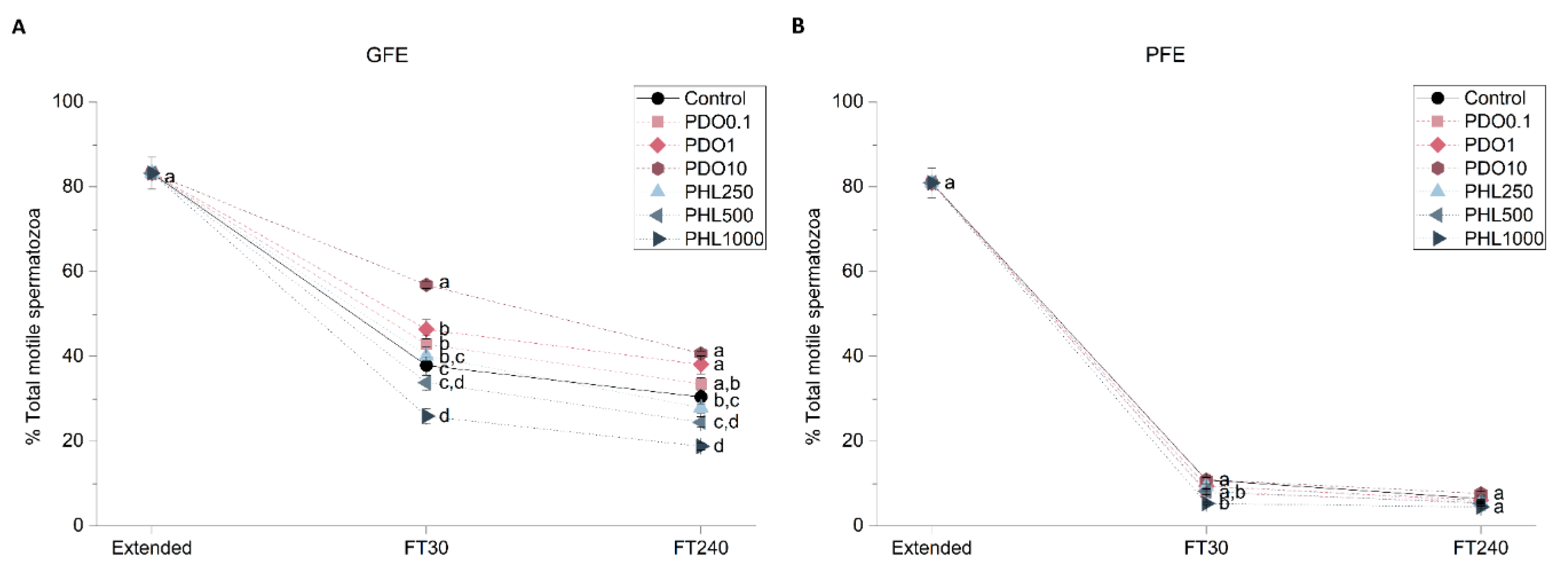
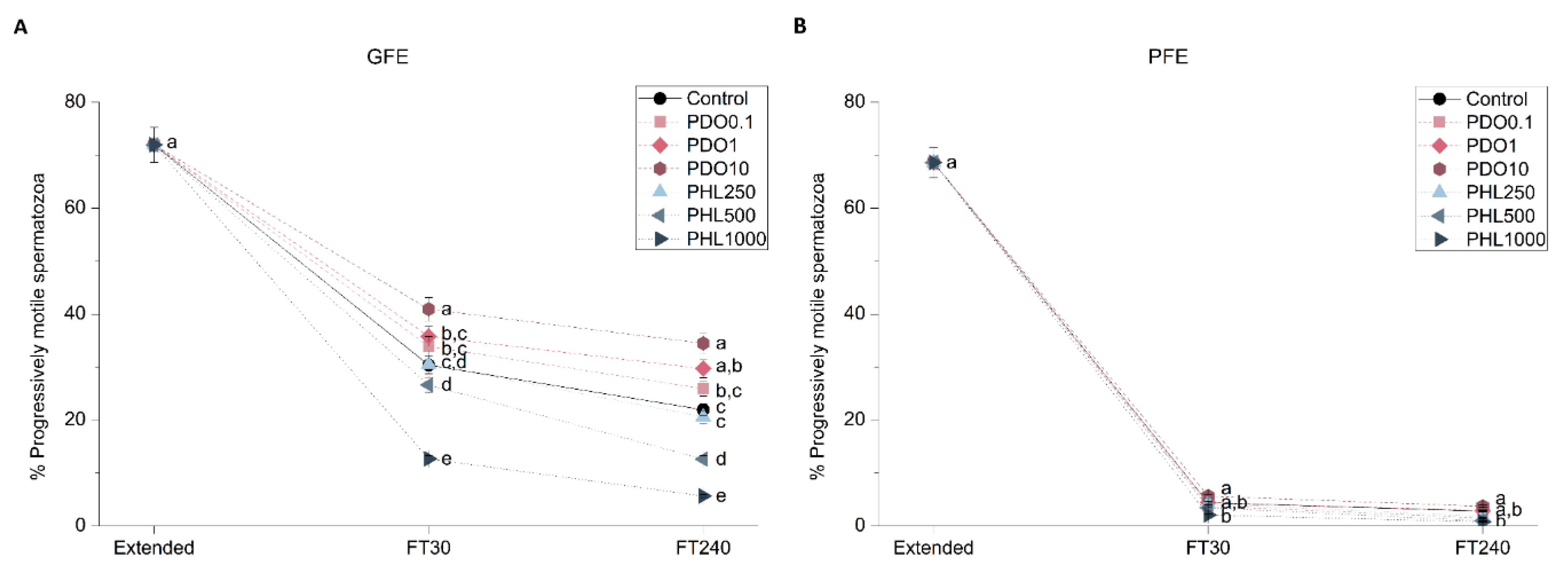
2.2.1. Sperm Motility
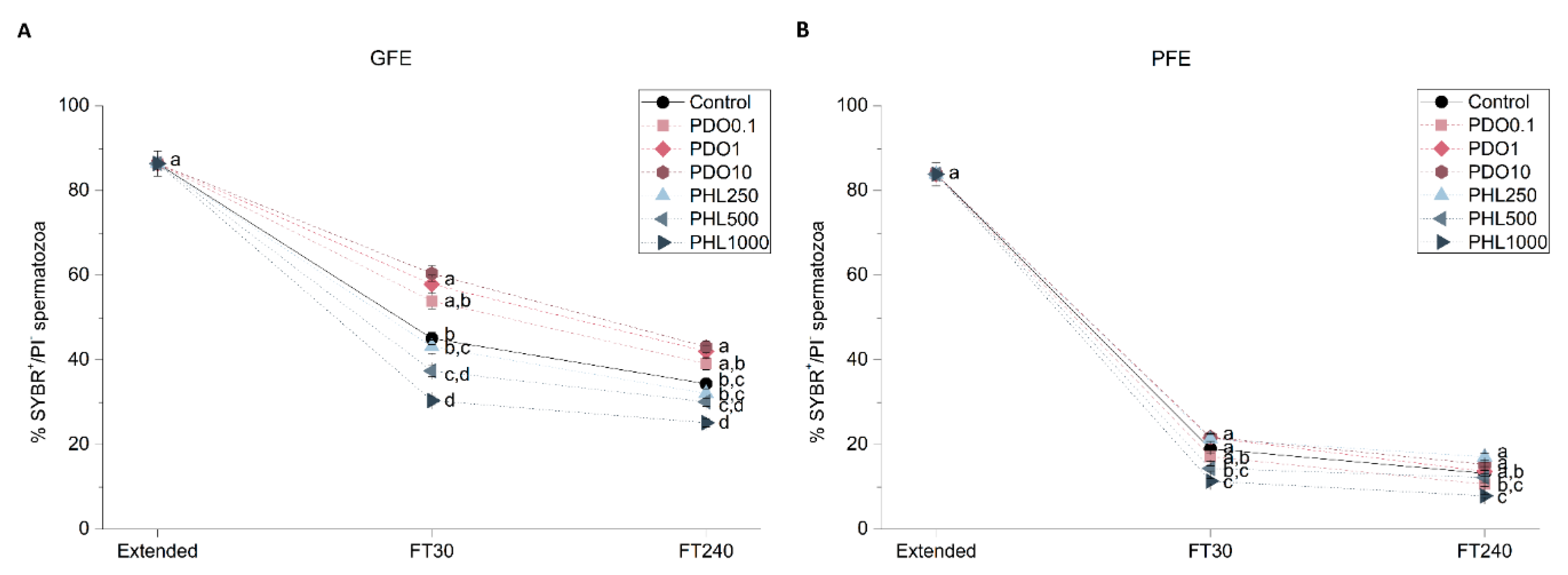
2.2.2. Sperm Viability
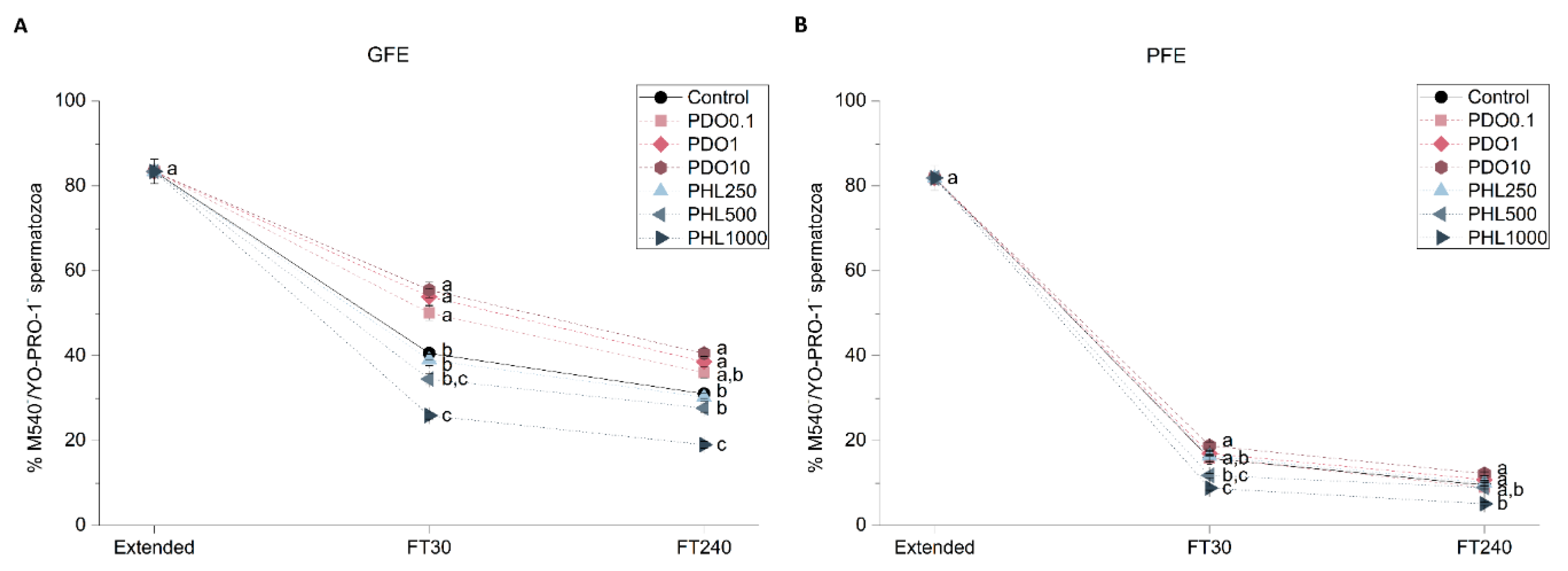
2.2.3. Sperm Membrane Lipid Disorder
2.2.4. Mitochondrial Membrane Potential (MMP)
2.2.5. Intracellular Levels of Superoxides (O2−•)
2.2.6. Intracellular Levels of Hydrogen Peroxides (H2O2)
3. Discussion
4. Materials and Methods
4.1. Boars and Ejaculates
4.2. AQP Inhibitors
4.3. Boar Sperm Cryopreservation
4.4. Sperm Motility
4.5. Flow Cytometry
4.5.1. Sperm Viability
4.5.2. Sperm Membrane Lipid Disorder
4.5.3. Mitochondrial Membrane Potential (MMP)
4.5.4. Intracellular Levels of Superoxides (O2−•)
4.5.5. Intracellular Levels of Hydrogen Peroxide (H2O2)
4.6. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AQP | Aquaporin |
| ALH | Amplitude of lateral head displacement |
| BCF | Beat cross frequency |
| BTS | Beltsville Thawing Solution |
| CASA | Computer-assisted sperm analysis |
| CPA | Cryoprotective agent |
| DCF+ | 2′,7′-dichlorofluorescein |
| E+ | Ethidium |
| EV | Electronic volume |
| FS | Forward scatter |
| GFE | Good freezability ejaculates |
| GLP | Aquaglyceroporins |
| H2DCFDA | 2′,7′-dichlorodihydrofluorescein diacetate |
| H2O2 | Hydrogen peroxide |
| HE | Hydroethidine |
| ISAC | International Society for Advancement of Cytometry |
| JC-1 | 5,5′,6,6′-tetrachloro-1,1′,3,3′tetraethyl-benzimidazolylcarbocyanine iodide |
| JC-1agg | JC-1 aggregates |
| JC-1mon | JC-1 monomers |
| LEY | β-Lactose–egg yolk–glycerol |
| LEYGO | β-Lactose–egg yolk–glycerol–Orvus ES Paste |
| LIN | Linearity |
| M540 | Merocyanine 540 |
| MMP | Mitochondrial membrane potential |
| O2−• | Superoxide |
| PDO | 1,3-propanediol |
| PfAQP | Plasmodium falciparum aquaporin |
| PFE | Poor freezability ejaculates |
| PHL | Phloretin |
| PI | Propidium iodide |
| PMOT | Progressive motility |
| ROS | Reactive oxygen species |
| SEM | Standard error of the mean |
| SLC2A2 | Solute carrier family 2, facilitated glucose transporter member 2 |
| SS | Side scatter |
| STR | Straightness |
| TMOT | Total motility |
| VAP | Average path velocity |
| VCL | Curvilinear velocity |
| VSL | Straight line velocity |
| WOB | Motility parameter wobble |
| PMOT | Progressive motility |
| ROS | Reactive oxygen species |
| SEM | Standard error of the mean |
| SLC2A2 | Solute carrier family 2, facilitated glucose transporter member 2 |
| SS | Side scatter |
| STR | Straightness |
References
- Watson, H. Biological membranes. Essays Biochem. 2015, 59, 43–70. [Google Scholar] [CrossRef]
- Yeste, M.; Morató, R.; Rodríguez-Gil, J.E.; Bonet, S.; Prieto-Martínez, N. Aquaporins in the male reproductive tract and sperm: Functional implications and cryobiology. Reprod. Domest. Anim. 2017, 52, 12–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prieto-Martínez, N.; Vilagran, I.; Morató, R.; Rodríguez-Gil, J.E.; Yeste, M.; Bonet, S. Aquaporins 7 and 11 in boar spermatozoa: Detection, localisation and relationship with sperm quality. Reprod. Fertil. Dev. 2014, 28, 663–672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prieto-Martínez, N.; Morató, R.; Vilagran, I.; Rodríguez-Gil, J.E.; Bonet, S.; Yeste, M. Aquaporins in boar spermatozoa. Part II: Detection and localisation of aquaglyceroporin 3. Reprod. Fertil. Dev. 2015, 29, 703–711. [Google Scholar] [CrossRef] [PubMed]
- Bonilla-Correal, S.; Noto, F.; Garcia-Bonavila, E.; Rodríguez-Gil, J.E.; Yeste, M.; Miró, J. First evidence for the presence of aquaporins in stallion sperm. Reprod. Domest. Anim. 2017, 52, 61–64. [Google Scholar] [CrossRef] [Green Version]
- Chen, Q.; Peng, H.; Lei, L.; Zhang, Y.; Kuang, H.; Cao, Y.; Shi, Q.; Ma, T.; Duan, E. Aquaporin 3 is a sperm water channel essential for postcopulatory sperm osmoadaptation and migration. Cell Res. 2011, 21, 922–933. [Google Scholar] [CrossRef]
- Yeung, C.H.; Callies, C.; Rojek, A.; Nielsen, S.; Cooper, T.G. Aquaporin Isoforms Involved in Physiological Volume Regulation of Murine Spermatozoa. Biol. Reprod. 2009, 80, 350–357. [Google Scholar] [CrossRef]
- Yeung, C.H.; Cooper, T.G. Aquaporin AQP11 in the testis: Molecular identity and association with the processing of residual cytoplasm of elongated spermatids. Reproduction 2010, 139, 209–216. [Google Scholar] [CrossRef] [Green Version]
- Yeung, C.H.; Callies, C.; Tüttelmann, F.; Kliesch, S.; Cooper, T.G. Aquaporins in the human testis and spermatozoa - identification, involvement in sperm volume regulation and clinical relevance. Int. J. Androl. 2010, 33, 629–641. [Google Scholar] [CrossRef]
- Laforenza, U.; Pellavio, G.; Marchetti, A.; Omes, C.; Todaro, F.; Gastaldi, G. Aquaporin-Mediated Water and Hydrogen Peroxide Transport Is Involved in Normal Human Spermatozoa Functioning. Int. J. Mol. Sci. 2017, 18, 66. [Google Scholar] [CrossRef] [Green Version]
- Prieto-Martínez, N.; Morató, R.; Muiño, R.; Hidalgo, C.O.; Rodríguez-Gil, J.E.; Bonet, S.; Yeste, M. Aquaglyceroporins 3 and 7 in bull spermatozoa: Identification, localisation and their relationship with sperm cryotolerance. Reprod. Fertil. Dev. 2016, 29, 1249–1259. [Google Scholar] [CrossRef] [PubMed]
- Morató, R.; Prieto-Martínez, N.; Muiño, R.; Hidalgo, C.O.; Rodríguez-Gil, J.E.; Bonet, S.; Yeste, M. Aquaporin 11 is related to cryotolerance and fertilising ability of frozen–thawed bull spermatozoa. Reprod. Fertil. Dev. 2018, 30, 1099–1108. [Google Scholar] [CrossRef] [PubMed]
- Vicente-Carrillo, A.; Ekwall, H.; Alvarez-Rodriguez, M.; Rodriguez-Martinez, H. Membrane Stress During Thawing Elicits Redistribution of Aquaporin 7 But Not of Aquaporin 9 in Boar Spermatozoa. Reprod. Domest. Anim. 2016, 51, 665–679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, H.F.; He, R.H.; Sun, C.C.; Zhang, Y.; Meng, Q.X.; Ma, Y.Y. Function of aquaporins in female and male reproductive systems. Hum. Reprod. Update 2006, 12, 785–795. [Google Scholar] [CrossRef] [Green Version]
- Bonet, S.; Casas, I.; Holt, W.V. Boar Reproduction: Fundamentals and New Biotechnological Trends, 1st ed.; Springer: London, UK, 2013; ISBN 9783642350481. [Google Scholar]
- Yeste, M. Sperm cryopreservation update: Cryodamage, markers, and factors affecting the sperm freezability in pigs. Theriogenology 2016, 85, 47–64. [Google Scholar] [CrossRef]
- Yeste, M.; Rodríguez-Gil, J.E.; Bonet, S. Artificial insemination with frozen-thawed boar sperm. Mol. Reprod. Dev. 2017, 84, 802–813. [Google Scholar] [CrossRef] [Green Version]
- Yeste, M. Recent advances in boar sperm cryopreservation: State of the art and current perspectives. Reprod. Domest. Anim. 2015, 50, 71–79. [Google Scholar] [CrossRef]
- Kuisma, P.; Andersson, M.; Koskinen, E.; Katila, T. Fertility of frozen-thawed stallion semen cannot be predicted by the currently used laboratory methods. Acta Vet. Scand. 2006, 48, 14. [Google Scholar] [CrossRef] [Green Version]
- Peña, F.J.; Macías García, B.; Samper, J.C.; Aparicio, I.M.; Tapia, J.A.; Ortega Ferrusola, C. Dissecting the molecular damage to stallion spermatozoa: The way to improve current cryopreservation protocols? Theriogenology 2011, 76, 1177–1186. [Google Scholar] [CrossRef]
- Casas, I.; Sancho, S.; Briz, M.; Pinart, E.; Bussalleu, E.; Yeste, M.; Bonet, S. Freezability prediction of boar ejaculates assessed by functional sperm parameters and sperm proteins. Theriogenology 2009, 72, 930–948. [Google Scholar] [CrossRef]
- Yeste, M.; Estrada, E.; Casas, I.; Bonet, S.; Rodríguez-Gil, J.E. Good and bad freezability boar ejaculates differ in the integrity of nucleoprotein structure after freeze-thawing but not in ROS levels. Theriogenology 2013, 79, 929–939. [Google Scholar] [CrossRef]
- Yeste, M.; Estrada, E.; Pinart, E.; Bonet, S.; Miró, J.; Rodríguez-Gil, J.E. The improving effect of reduced glutathione on boar sperm cryotolerance is related with the intrinsic ejaculate freezability. Cryobiology 2014, 68, 251–261. [Google Scholar] [CrossRef] [PubMed]
- Vilagran, I.; Yeste, M.; Sancho, S.; Casas, I.; Rivera del Álamo, M.M.; Bonet, S. Relationship of sperm small heat-shock protein 10 and voltage-dependent anion channel 2 with semen freezability in boars. Theriogenology 2014, 82, 418–426. [Google Scholar] [CrossRef]
- Prieto-Martínez, N.; Vilagran, I.; Morató, R.; Rivera del Álamo, M.M.; Rodríguez-Gil, J.E.; Bonet, S.; Yeste, M. Relationship of aquaporins 3 (AQP3), 7 (AQP7), and 11 (AQP11) with boar sperm resilience to withstand freeze-thawing procedures. Andrology 2017, 5, 1153–1164. [Google Scholar] [CrossRef] [Green Version]
- Delgado-Bermúdez, A.; Llavanera, M.; Fernández-Bastit, L.; Recuero, S.; Mateo, Y.; Bonet, S.; Barranco, I.; Fernández-Fuertes, B.; Yeste, M. Aquaglyceroporins but not orthodox aquaporins are involved in the cryotolerance of pig spermatozoa. J. Anim. Sci. Biotechnol. 2019, in press. [Google Scholar]
- Delgado-Bermúdez, A.; Noto, F.; Bonilla-Correal, S.; Garcia-Bonavila, E.; Catalán, J.; Papas, M.; Bonet, S.; Miró, J.; Yeste, M. Cryotolerance of stallion spermatozoa relies on GLPs rather than orthodox AQPs. Biology (Basel) 2019, 8, 85. [Google Scholar]
- Yu, L.; Rodriguez, R.A.; Chen, L.L.; Chen, L.Y.; Perry, G.; McHardy, S.F.; Yeh, C.K. 1,3-Propanediol Binds Deep Inside the Channel To Inhibit Water Permeation Through Aquaporins. Protein Sci. 2016, 25, 433–441. [Google Scholar] [CrossRef] [Green Version]
- Yu, L.; Villarreal, O.D.; Chen, L.L.; Chen, L.Y. 1,3-Propanediol binds inside the water-conducting pore of aquaporin 4: Does this efficacious inhibitor have sufficient potency? J. Syst. Integr. Neurosci. 2016, 2, 91–98. [Google Scholar] [CrossRef] [Green Version]
- Tsukaguchi, H.; Shayakul, C.; Berger, U.V.; Mackenzie, B.; Devidas, S.; Guggino, W.B.; van Hoek, A.N.; Hediger, M.A. Molecular Characterization of a Broad Selectivity Neutral Solute Channel. J. Biol. Chem. 1998, 273, 24737–24743. [Google Scholar] [CrossRef] [Green Version]
- Cheung, K.H.; Leung, C.T.; Leung, G.P.H.; Wong, P.Y.D. Synergistic Effects of Cystic Fibrosis Transmembrane Conductance Regulator and Aquaporin-9 in the Rat Epididymis. Biol. Reprod. 2003, 68, 1505–1510. [Google Scholar] [CrossRef] [Green Version]
- Rezk, B.M.; Haenen, G.R.M.M.; van der Vijgh, W.J.F.; Bast, A. The antioxidant activity of phloretin: The disclosure of a new antioxidant pharmacophore in flavonoids. Biochem. Biophys. Res. Commun. 2002, 295, 9–13. [Google Scholar] [CrossRef]
- Barreca, D.; Currò, M.; Bellocco, E.; Ficarra, S.; Laganà, G.; Tellone, E.; Giunta, M.L.; Visalli, G.; Caccamo, D.; Galtieri, A.; et al. Neuroprotective effects of phloretin and its glycosylated derivative on rotenone-induced toxicity in human SH-SY5Y neuronal-like cells. BioFactors 2017, 43, 549–557. [Google Scholar] [CrossRef] [PubMed]
- Przybylo, M.; Procek, J.; Hof, M.; Langner, M. The alteration of lipid bilayer dynamics by phloretin and 6-ketocholestanol. Chem. Phys. Lipids 2014, 178, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Pohl, P.; Rokitskaya, T.I.; Pohl, E.E.; Saparov, S.M. Permeation of phloretin across bilayer lipid membranes monitored by dipole potential and microelectrode measurements. Biochim. Biophys. Acta-Biomembr. 1997, 1323, 163–172. [Google Scholar] [CrossRef] [Green Version]
- Wacker, S.J.; Aponte-Santamaría, C.; Kjellbom, P.; Nielsen, S.; De Groot, B.L.; Rützler, M. The identification of novel, high affinity AQP9 inhibitors in an intracellular binding site. Mol. Membr. Biol. 2013, 30, 246–260. [Google Scholar] [CrossRef] [Green Version]
- Fan, H.; Morishima, S.; Kida, H.; Okada, Y. Phloretin differentially inhibits volume-sensitive and cyclic AMP-activated, but not Ca-activated, Cl- channels. Br. J. Pharmacol. 2001, 133, 1096–1106. [Google Scholar] [CrossRef] [Green Version]
- Bucci, D.; Isani, G.; Spinaci, M.; Tamanini, C.; Mari, G.; Zambelli, D.; Galeati, G. Comparative Immunolocalization of GLUTs 1, 2, 3 and 5 in Boar, Stallion and Dog Spermatozoa. Reprod. Domest. Anim. 2010, 45, 315–322. [Google Scholar] [CrossRef]
- Lopes, C.A.; Alves, A.M.; Jewgenow, K.; Báo, S.N.; de Figueiredo, J.R. Cryopreservation of canine ovarian cortex using DMSO or 1,3-propanediol. Theriogenology 2016, 86, 1165–1174. [Google Scholar] [CrossRef]
- Widiasih, D.; Yeung, C.H.; Junaidi, A.; Cooper, T.G. Multistep and single-step treatment of human spermatozoa with cryoprotectants. Fertil. Steril. 2009, 92, 382–389. [Google Scholar] [CrossRef]
- Miller, E.W.; Dickinson, B.C.; Chang, C.J. Aquaporin-3 mediates hydrogen peroxide uptake to regulate downstream intracellular signaling. Proc. Natl. Acad. Sci. 2010, 107, 15681–15686. [Google Scholar] [CrossRef] [Green Version]
- Aitken, R.J. Reactive oxygen species as mediators of sperm capacitation and pathological damage. Mol. Reprod. Dev. 2017, 84, 1039–1052. [Google Scholar] [CrossRef] [PubMed]
- Casas, I.; Sancho, S.; Briz, M.; Pinart, E.; Bussalleu, E.; Yeste, M.; Bonet, S. Fertility after post-cervical artificial insemination with cryopreserved sperm from boar ejaculates of good and poor freezability. Anim. Reprod. Sci. 2010, 118, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Ramió, L.; Rivera, M.M.; Ramírez, A.; Concha, I.I.; Peña, A.; Rigau, T.; Rodríguez-Gil, J.E. Dynamics of motile-sperm subpopulation structure in boar ejaculates subjected to “in vitro” capacitation and further “in vitro” acrosome reaction. Theriogenology 2008, 69, 501–512. [Google Scholar] [CrossRef] [PubMed]
- Verstegen, J.; Iguer-Ouada, M.; Onclin, K. Computer assisted semen analyzers in andrology research and veterinary practice. Theriogenology 2002, 57, 149–179. [Google Scholar] [CrossRef]
- Maes, D.G.D.; Mateusen, B.; Rijsselaere, T.; de Vliegher, S.; van Soom, A.; de Kruif, A. Motility characteristics of boar spermatozoa after addition of prostaglandin F2a. Theriogenology 2003, 60, 1435–1443. [Google Scholar] [CrossRef]
- Petrunkina, A.M.; Waberski, D.; Bollwein, H.; Sieme, H. Identifying non-sperm particles during flow cytometric physiological assessment: A simple approach. Theriogenology 2010, 73, 995–1000. [Google Scholar] [CrossRef]
- Garner, D.L.; Johnson, L.A. Viability assessment of mammalian sperm using SYBR-14 and propidium iodide. Biol. Reprod. 1995, 53, 276–284. [Google Scholar] [CrossRef]
- Rathi, R.; Colenbrander, B.; Bevers, M.M.; Gadella, B.M. Evaluation of in vitro capacitation of stallion spermatozoa. Biol. Reprod. 2001, 65, 462–470. [Google Scholar] [CrossRef] [Green Version]
- Yeste, M.; Estrada, E.; Rivera Del Álamo, M.M.; Bonet, S.; Rigau, T.; Rodríguez-Gil, J.E. The increase in phosphorylation levels of serine residues of protein HSP70 during holding time at 17 °C is concomitant with a higher cryotolerance of boar spermatozoa. PLoS ONE 2014, 9, e90887. [Google Scholar] [CrossRef] [Green Version]
- Ortega-Ferrusola, C.; Sotillo-Galán, Y.; Varela-Fernández, E.; Gallardo-Bolaños, J.M.; Muriel, A.; González-Fernández, L.; Tapia, J.A.; Peña, F.J. Detection of “Apoptosis-Like” Changes During the Cryopreservation Process in Equine Sperm. J. Androl. 2008, 29, 213–221. [Google Scholar] [CrossRef]
- Guthrie, H.D.; Welch, G.R. Determination of intracellular reactive oxygen species and high mitochondrial membrane potential in Percoll-treated viable boar sperm using fluorescence-activated flow cytometry. J. Anim. Sci. 2006, 84, 2089–2100. [Google Scholar] [CrossRef] [Green Version]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Delgado-Bermúdez, A.; Llavanera, M.; Recuero, S.; Mateo-Otero, Y.; Bonet, S.; Barranco, I.; Fernandez-Fuertes, B.; Yeste, M. Effect of AQP Inhibition on Boar Sperm Cryotolerance Depends on the Intrinsic Freezability of the Ejaculate. Int. J. Mol. Sci. 2019, 20, 6255. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20246255
Delgado-Bermúdez A, Llavanera M, Recuero S, Mateo-Otero Y, Bonet S, Barranco I, Fernandez-Fuertes B, Yeste M. Effect of AQP Inhibition on Boar Sperm Cryotolerance Depends on the Intrinsic Freezability of the Ejaculate. International Journal of Molecular Sciences. 2019; 20(24):6255. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20246255
Chicago/Turabian StyleDelgado-Bermúdez, Ariadna, Marc Llavanera, Sandra Recuero, Yentel Mateo-Otero, Sergi Bonet, Isabel Barranco, Beatriz Fernandez-Fuertes, and Marc Yeste. 2019. "Effect of AQP Inhibition on Boar Sperm Cryotolerance Depends on the Intrinsic Freezability of the Ejaculate" International Journal of Molecular Sciences 20, no. 24: 6255. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20246255