Sequential Transcriptome Changes in the Penumbra after Ischemic Stroke


Abstract
:1. Introduction
2. Results
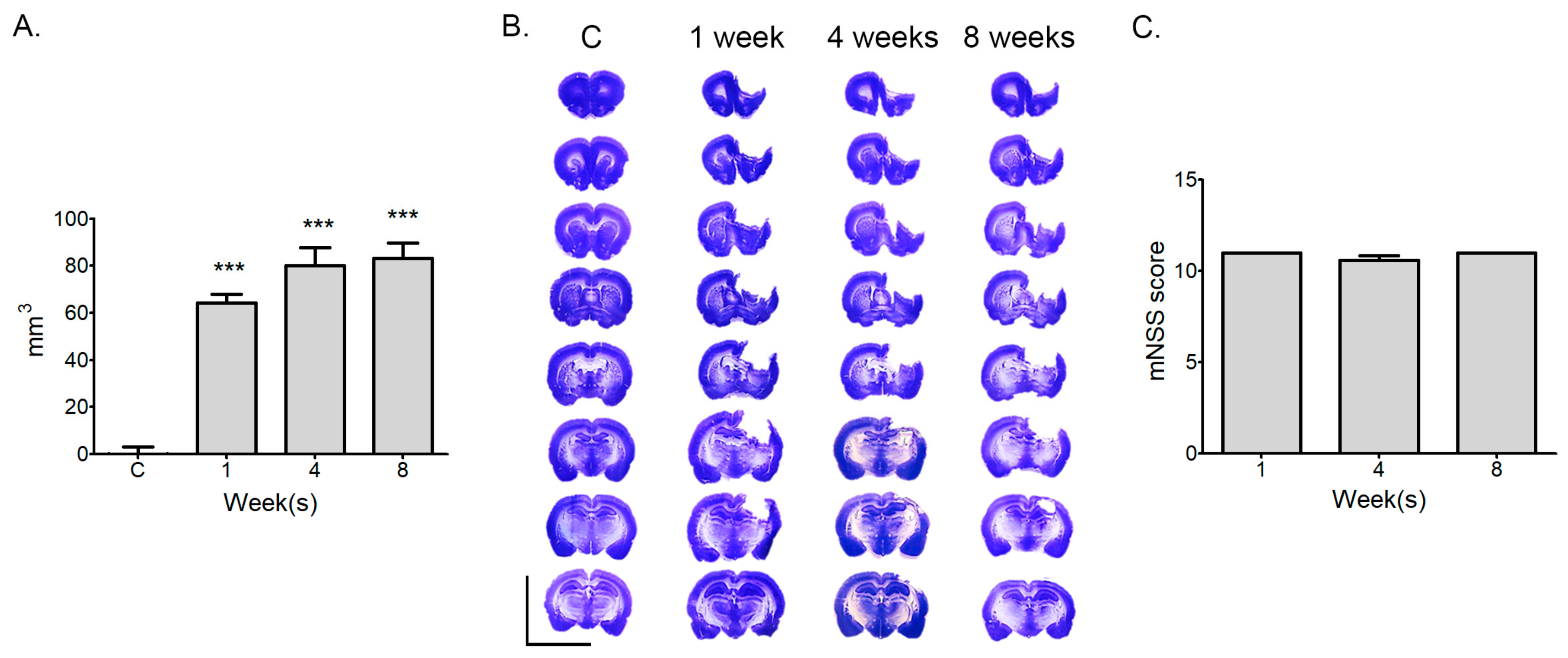
2.1. Measurement of Infarction Volume and Neurological Deficit at Various Time Points after Stroke
2.2. RNA-Seq Analysis of Gene Expression Profiles in Photothrombotic Ischemic Stroke
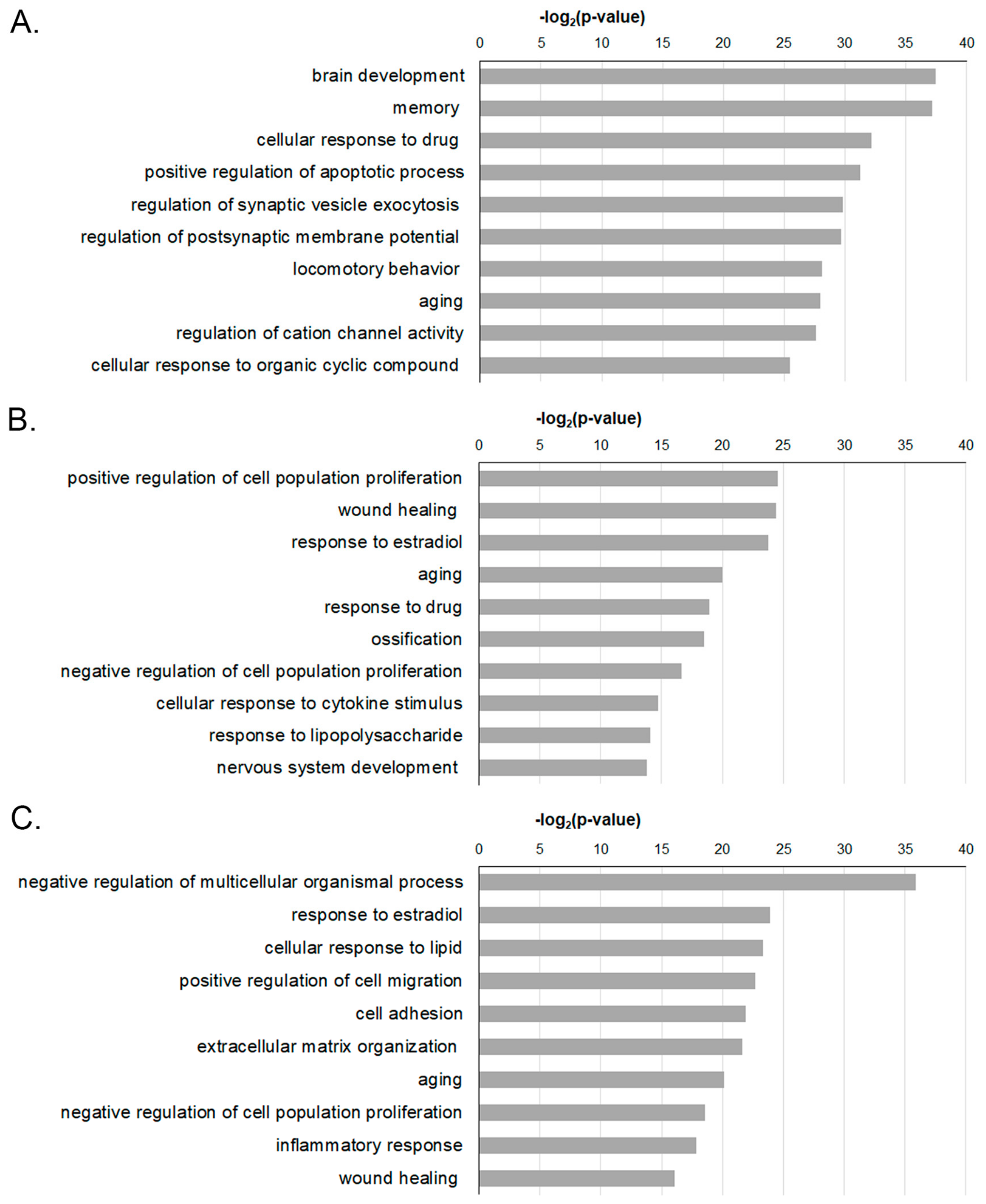
2.3. Functional Enrichment Analysis of DEGS-Temporal Changes in the GO Analysis
2.4. Analysis of the Functional Genes in the Ontology of the Biological Processes
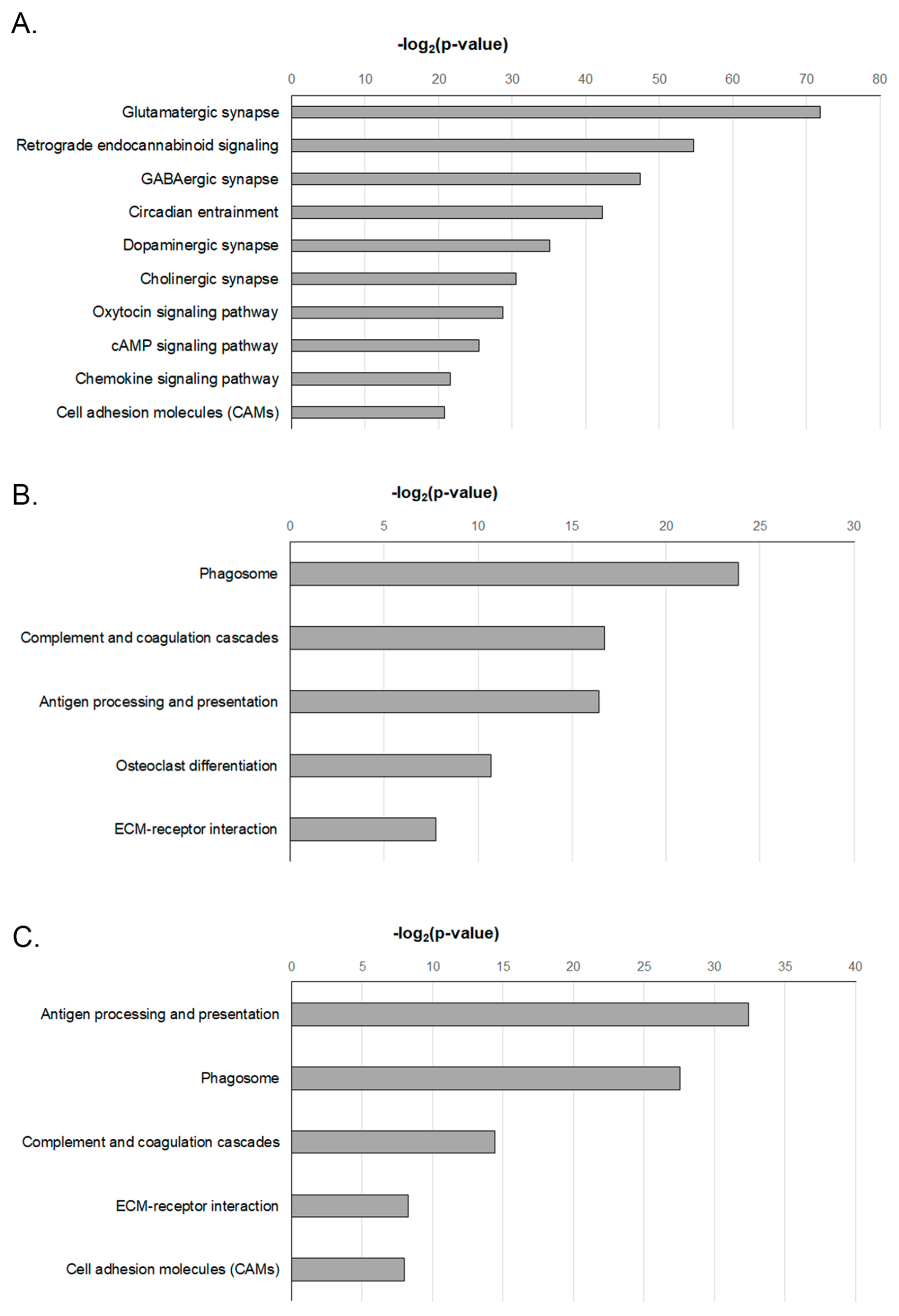
2.5. Functional Enrichment Analysis of DEGS-Temporal Changes in the KEGG Analysis
2.6. PPI Network Analysis
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Photothrombotic Stroke Model
4.3. Determination of the Infarction Volume
4.4. Modified Neurological Severity Score
4.5. RNA Extraction and RNA-Sequencing Analysis
4.6. Pathway Analysis
4.7. Gene Ontology (GO) Analysis
4.8. Identification of Key Genes from Protein–Protein Interaction (PPI) Analysis
4.9. Total RNA Extraction and RT-PCR Analysis
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| DEGs | Differentially expressed genes |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| GO | Gene ontology |
| PPI | Protein–protein interactions |
| RNA-seq | RNA-sequencing |
| ANOVA | Analysis of variance |
| h | Hour(s) |
| mNSS | Modified neurological severity score |
| SEM | Standard error of means |
| mRNA | Messenger RNA |
| ECM | Extracellular matrix |
| IGF-1 | Insulin-like growth factor |
| DAVID | Database for Annotation, Visualization and Integrated Discovery |
| GABA | Gamma aminobutyric acid |
| PSD-95 | Post-synaptic density protein 95 |
| SAP-90 | synaptic-related protein 90 |
| BDNF | Brain-derived neurotrophic factor |
| AMPA | α-Amino-3-hydroxy-5-methylisoxazole-4-propionic acid |
| NMDA | N-Methyl- d-aspartic acid or N-Methyl- d-aspartate |
| RT-PCR | Reverse transcription polymerase chain reaction |
| cAMP | 3′,5″-Cyclic adenosine monophosphate |
| GluA1 | Glutamate A1 |
| GluNMDA2A | NMDA-type glutamate receptor 2A subunit |
| JNK | c-Jun N-terminal kinases |
| Spp1 | Secreted Phosphoprotein 1 |
| FDR | False discovery rate |
| FPKM | Fragments Per Kilobase of exon per Million fragments mapped |
| GAPDH | Glyceraldehyde 3-phosphate dehydrogenase |
References
- Benjamin, E.J.; Blaha, M.J.; Chiuve, S.E.; Cushman, M.; Das, S.R.; Deo, R.; de Ferranti, S.D.; Floyd, J.; Fornage, M.; Gillespie, C.; et al. Heart Disease and Stroke Statistics-2017 Update: A Report From the American Heart Association. Circulation 2017, 135, e146–e603. [Google Scholar] [CrossRef]
- Cramer, S.C. Treatments to Promote Neural Repair after Stroke. J. Stroke 2018, 20, 57–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emberson, J.; Lees, K.R.; Lyden, P.; Blackwell, L.; Albers, G.; Bluhmki, E.; Brott, T.; Cohen, G.; Davis, S.; Donnan, G.; et al. Effect of treatment delay, age, and stroke severity on the effects of intravenous thrombolysis with alteplase for acute ischaemic stroke: A meta-analysis of individual patient data from randomised trials. Lancet 2014, 384, 1929–1935. [Google Scholar] [CrossRef] [Green Version]
- Krakauer, J.W.; Carmichael, S.T.; Corbett, D.; Wittenberg, G.F. Getting neurorehabilitation right: What can be learned from animal models? Neurorehabilit. Neural Repair 2012, 26, 923–931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindenberg, R.; Renga, V.; Zhu, L.L.; Betzler, F.; Alsop, D.; Schlaug, G. Structural integrity of corticospinal motor fibers predicts motor impairment in chronic stroke. Neurology 2010, 74, 280–287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, L.L.; Lindenberg, R.; Alexander, M.P.; Schlaug, G. Lesion load of the corticospinal tract predicts motor impairment in chronic stroke. Stroke 2010, 41, 910–915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, N.; Chopp, M.; Xiong, Y.; Qian, J.Y.; Lu, M.; Zhou, D.; He, L.; Liu, Z. Subacute intranasal administration of tissue plasminogen activator improves stroke recovery by inducing axonal remodeling in mice. Exp. Neurol. 2018, 304, 82–89. [Google Scholar] [CrossRef]
- Egawa, N.; Lok, J.; Washida, K.; Arai, K. Mechanisms of Axonal Damage and Repair after Central Nervous System Injury. Transl. Stroke Res. 2017, 8, 14–21. [Google Scholar] [CrossRef]
- Okabe, N.; Narita, K.; Miyamoto, O. Axonal remodeling in the corticospinal tract after stroke: How does rehabilitative training modulate it? Neural Regen. Res. 2017, 12, 185–192. [Google Scholar] [CrossRef]
- Wahl, A.S.; Buchler, U.; Brandli, A.; Brattoli, B.; Musall, S.; Kasper, H.; Ineichen, B.V.; Helmchen, F.; Ommer, B.; Schwab, M.E. Optogenetically stimulating intact rat corticospinal tract post-stroke restores motor control through regionalized functional circuit formation. Nat. Commun. 2017, 8, 1187. [Google Scholar] [CrossRef] [Green Version]
- Wiersma, A.M.; Fouad, K.; Winship, I.R. Enhancing Spinal Plasticity Amplifies the Benefits of Rehabilitative Training and Improves Recovery from Stroke. J. Neurosci. Off. J. Soc. Neurosci. 2017, 37, 10983–10997. [Google Scholar] [CrossRef] [PubMed]
- Hermann, D.M.; Chopp, M. Promoting brain remodelling and plasticity for stroke recovery: Therapeutic promise and potential pitfalls of clinical translation. Lancet Neurol. 2012, 11, 369–380. [Google Scholar] [CrossRef] [Green Version]
- Alia, C.; Spalletti, C.; Lai, S.; Panarese, A.; Lamola, G.; Bertolucci, F.; Vallone, F.; Di Garbo, A.; Chisari, C.; Micera, S. Neuroplastic changes following brain ischemia and their contribution to stroke recovery: Novel approaches in neurorehabilitation. Front. Cell. Neurosci. 2017, 11, 76. [Google Scholar] [CrossRef] [PubMed]
- Carmichael, S.T.; Archibeque, I.; Luke, L.; Nolan, T.; Momiy, J.; Li, S. Growth-associated gene expression after stroke: Evidence for a growth-promoting region in peri-infarct cortex. Exp. Neurol. 2005, 193, 291–311. [Google Scholar] [CrossRef] [PubMed]
- Blackstock, C.D.; Higashi, Y.; Sukhanov, S.; Shai, S.Y.; Stefanovic, B.; Tabony, A.M.; Yoshida, T.; Delafontaine, P. Insulin-like growth factor-1 increases synthesis of collagen type I via induction of the mRNA-binding protein LARP6 expression and binding to the 5′ stem-loop of COL1a1 and COL1a2 mRNA. J. Biol. Chem. 2014, 289, 7264–7274. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Zhan, Y.; Xu, L.; Feuerstein, G.Z.; Wang, X. Use of suppression subtractive hybridization for differential gene expression in stroke: Discovery of CD44 gene expression and localization in permanent focal stroke in rats. Stroke 2001, 32, 1020–1027. [Google Scholar] [CrossRef] [Green Version]
- Mlecnik, B.; Galon, J.; Bindea, G. Comprehensive functional analysis of large lists of genes and proteins. J. Proteom. 2018, 171, 2–10. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Zhu, Y.; Wang, S.; Zachory Wei, Z.; Jiang, M.Q.; Zhang, Y.; Pan, Y.; Tao, S.; Li, J.; Wei, L. Temporal Gene Expression Profiles after Focal Cerebral Ischemia in Mice. Aging Dis. 2018, 9, 249–261. [Google Scholar] [CrossRef] [Green Version]
- Buga, A.M.; Margaritescu, C.; Scholz, C.J.; Radu, E.; Zelenak, C.; Popa-Wagner, A. Transcriptomics of post-stroke angiogenesis in the aged brain. Front. Aging Neurosci. 2014, 6, 44. [Google Scholar] [CrossRef]
- Li, S.; Overman, J.J.; Katsman, D.; Kozlov, S.V.; Donnelly, C.J.; Twiss, J.L.; Giger, R.J.; Coppola, G.; Geschwind, D.H.; Carmichael, S.T. An age-related sprouting transcriptome provides molecular control of axonal sprouting after stroke. Nat. Neurosci. 2010, 13, 1496. [Google Scholar] [CrossRef]
- Buga, A.M.; Sascau, M.; Pisoschi, C.; Herndon, J.G.; Kessler, C.; Popa-Wagner, A. The genomic response of the ipsilateral and contralateral cortex to stroke in aged rats. J. Cell. Mol. Med. 2008, 12, 2731–2753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ito, M.; Aswendt, M.; Lee, A.G.; Ishizaka, S.; Cao, Z.; Wang, E.H.; Levy, S.L.; Smerin, D.L.; McNab, J.A.; Zeineh, M.; et al. RNA-Sequencing Analysis Revealed a Distinct Motor Cortex Transcriptome in Spontaneously Recovered Mice After Stroke. Stroke 2018, 49, 2191–2199. [Google Scholar] [CrossRef] [PubMed]
- Choi, I.A.; Lee, C.S.; Kim, H.Y.; Choi, D.H.; Lee, J. Effect of Inhibition of DNA Methylation Combined with Task-Specific Training on Chronic Stroke Recovery. Int. J. Mol. Sci. 2018, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carmichael, S.T. Rodent models of focal stroke: Size, mechanism, and purpose. NeuroRx J. Am. Soc. Exp. Neurother. 2005, 2, 396–409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durukan, A.; Tatlisumak, T. Acute ischemic stroke: Overview of major experimental rodent models, pathophysiology, and therapy of focal cerebral ischemia. Pharmacol. Biochem. Behav. 2007, 87, 179–197. [Google Scholar] [CrossRef]
- Murphy, T.H.; Corbett, D. Plasticity during stroke recovery: From synapse to behaviour. Nat. Rev. Neurosci. 2009, 10, 861–872. [Google Scholar] [CrossRef]
- Trompet, S.; Jukema, W.; Mooijaart, S.P.; Ford, I.; Stott, D.J.; Westendorp, R.G.; de Craen, A.J. Genetic variation in galectin-3 gene associates with cognitive function at old age. Neurobiol. Aging 2012, 33, e2231–e2232, e2239. [Google Scholar] [CrossRef] [Green Version]
- Lalancette-Hebert, M.; Gowing, G.; Simard, A.; Weng, Y.C.; Kriz, J. Selective ablation of proliferating microglial cells exacerbates ischemic injury in the brain. J. Neurosci. 2007, 27, 2596–2605. [Google Scholar] [CrossRef] [Green Version]
- Lewitt, M.S.; Boyd, G.W. The Role of Insulin-Like Growth Factors and Insulin-Like Growth Factor-Binding Proteins in the Nervous System. Biochem. Insights 2019, 12, 1178626419842176. [Google Scholar] [CrossRef] [Green Version]
- Doverhag, C.; Hedtjarn, M.; Poirier, F.; Mallard, C.; Hagberg, H.; Karlsson, A.; Savman, K. Galectin-3 contributes to neonatal hypoxic-ischemic brain injury. Neurobiol. Dis. 2010, 38, 36–46. [Google Scholar] [CrossRef]
- Doyle, K.P.; Yang, T.; Lessov, N.S.; Ciesielski, T.M.; Stevens, S.L.; Simon, R.P.; King, J.S.; Stenzel-Poore, M. Nasal Administration of Osteopontin Peptide Mimetics Confers Neuroprotection in Stroke. J. Cereb. Blood Flow Metab. 2008, 28, 1235–1248. [Google Scholar] [CrossRef] [PubMed]
- Meller, R.; Stevens, S.L.; Minami, M.; Cameron, J.A.; King, S.; Rosenzweig, H.; Doyle, K.; Lessov, N.S.; Simon, R.P.; Stenzel-Poore, M.P. Neuroprotection by osteopontin in stroke. J. Cereb. Blood Flow Metab. Off. J. Int. Soc. Cereb. Blood Flow Metab. 2005, 25, 217–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gliem, M.; Krammes, K.; Liaw, L.; van Rooijen, N.; Hartung, H.P.; Jander, S. Macrophage-derived osteopontin induces reactive astrocyte polarization and promotes re-establishment of the blood brain barrier after ischemic stroke. Glia 2015, 63, 2198–2207. [Google Scholar] [CrossRef] [PubMed]
- Fisher, M.; Feuerstein, G.; Howells, D.W.; Hurn, P.D.; Kent, T.A.; Savitz, S.I.; Lo, E.H.; Group, S. Update of the stroke therapy academic industry roundtable preclinical recommendations. Stroke 2009, 40, 2244–2250. [Google Scholar] [CrossRef] [PubMed]
- Appelros, P.; Stegmayr, B.; Terent, A. Sex differences in stroke epidemiology: A systematic review. Stroke 2009, 40, 1082–1090. [Google Scholar] [CrossRef]
- Herson, P.S.; Traystman, R.J. Animal models of stroke: Translational potential at present and in 2050. Future Neurol. 2014, 9, 541–551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toung, T.K.; Hurn, P.D.; Traystman, R.J.; Sieber, F.E. Estrogen decreases infarct size after temporary focal ischemia in a genetic model of type 1 diabetes mellitus. Stroke 2000, 31, 2701–2706. [Google Scholar] [CrossRef] [Green Version]
- Vannucci, S.J.; Willing, L.B.; Goto, S.; Alkayed, N.J.; Brucklacher, R.M.; Wood, T.L.; Towfighi, J.; Hurn, P.D.; Simpson, I.A. Experimental stroke in the female diabetic, db/db, mouse. J. Cereb. Blood Flow Metab. 2001, 21, 52–60. [Google Scholar] [CrossRef] [Green Version]
- Choi, D.H.; Ahn, J.H.; Choi, I.A.; Kim, J.H.; Kim, B.R.; Lee, J. Effect of task-specific training on Eph/ephrin expression after stroke. BMB Rep. 2016, 49, 635–640. [Google Scholar] [CrossRef] [Green Version]
- Bodian, D.L.; Solomon, B.D.; Khromykh, A.; Thach, D.C.; Iyer, R.K.; Link, K.; Baker, R.L.; Baveja, R.; Vockley, J.G.; Niederhuber, J.E. Diagnosis of an imprinted-gene syndrome by a novel bioinformatics analysis of whole-genome sequences from a family trio. Mol. Genet. Genom. Med. 2014, 2, 530–538. [Google Scholar] [CrossRef]
- Choi, N.; Choi, J.; Kim, J.H.; Jang, Y.; Yeo, J.H.; Kang, J.; Song, S.Y.; Lee, J.; Sung, J.H. Generation of trichogenic adipose-derived stem cells by expression of three factors. J. Dermatol. Sci. 2018, 92, 18–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.-C.; Mendell, J.T.; Salzberg, S.L. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 33, 290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pertea, M.; Kim, D.; Pertea, G.M.; Leek, J.T.; Salzberg, S.L. Transcript-level expression analysis of RNA-seq experiments with HISAT, StringTie and Ballgown. Nat. Protoc. 2016, 11, 1650. [Google Scholar] [CrossRef]
- Sohn, M.; Na, H.Y.; Ryu, S.H.; Choi, W.; In, H.; Shin, H.S.; Park, J.S.; Shim, D.; Shin, S.J.; Park, C.G. Two Distinct Subsets Are Identified from the Peritoneal Myeloid Mononuclear Cells Expressing both CD11c and CD115. Immune Netw. 2019, 19, e15. [Google Scholar] [CrossRef]
- Lee, S.E.; Kang, S.G.; Choi, M.J.; Jung, S.B.; Ryu, M.J.; Chung, H.K.; Chang, J.Y.; Kim, Y.K.; Lee, J.H.; Kim, K.S.; et al. Growth Differentiation Factor 15 Mediates Systemic Glucose Regulatory Action of T-Helper Type 2 Cytokines. Diabetes 2017, 66, 2774–2788. [Google Scholar] [CrossRef] [Green Version]
- Chung, H.K.; Ryu, D.; Kim, K.S.; Chang, J.Y.; Kim, Y.K.; Yi, H.S.; Kang, S.G.; Choi, M.J.; Lee, S.E.; Jung, S.B.; et al. Growth differentiation factor 15 is a myomitokine governing systemic energy homeostasis. J. Cell Biol. 2017, 216, 149–165. [Google Scholar] [CrossRef]
- Choi, D.H.; Kim, J.H.; Seo, J.H.; Lee, J.; Choi, W.S.; Kim, Y.S. Matrix metalloproteinase-3 causes dopaminergic neuronal death through Nox1-regenerated oxidative stress. PLoS ONE 2014, 9, e115954. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 1 week post stroke | |||||
| Gene_ID | Gene_Symbol of Rats | Description | Fold Change | log2(Fc) | Volume |
| 50665 | Tmsb10 | thymosin, beta 10 | 2.98 | 1.57 | 10.03 |
| 29292 | Ftl1 | ferritin light chain 1 | 7.36 | 2.88 | 9.96 |
| 64529 | Ctsb | cathepsin B | 7.31 | 2.87 | 9.46 |
| 171293 | Ctsd | cathepsin D | 12.36 | 3.63 | 9.29 |
| 24223 | B2m | beta-2 microglobulin | 3.28 | 1.71 | 8.77 |
| 24547 | Mbp | myelin basic protein | −22.65 | −3.32 | 10.88 |
| 25742 | S100b | S100 calcium binding protein B | −6.88 | −2.32 | 9.76 |
| 287005 | Camk2n1 | calcium/calmodulin-dependent protein kinase II inhibitor 1 | −48.01 | −4.04 | 9.39 |
| 24191 | Aldoc | aldolase, fructose-bisphosphate C | −9.07 | −2.80 | 9.03 |
| 171114 | Ndrg2 | NDRG family member 2 | −7.55 | −2.50 | 8.72 |
| 4 week post stroke | |||||
| Gene_ID | Gene_Symbol of Rats | Description | Fold Change | log2(Fc) | Volume |
| 25526 | Ptgds | prostaglandin D2 synthase | 13.45 | 1.58 | 9.77 |
| 24387 | Gfap | glial fibrillary acidic protein | 8.65 | 1.73 | 8.36 |
| 80841 | Fabp7 | fatty acid binding protein 7 | 4.86 | 2.88 | 7.00 |
| 286898 | Npc2 | NPC intracellular cholesterol transporter 2 | 2.84 | 2.92 | 6.86 |
| 94270 | Nnat | neuronatin | 4.69 | 2.90 | 6.71 |
| 362220 | Lamp5 | lysosomal-associated membrane protein family, member 5 | −2.88 | −1.77 | 6.73 |
| 24587 | Nefh | neurofilament, heavy polypeptide | −3.72 | −1.41 | 5.54 |
| 315611 | Scn4b | sodium voltage-gated channel beta subunit 4 | −3.82 | −1.85 | 4.57 |
| 108348108 | LOC108348108 | heat shock 70 kDa protein 1A | −5.71 | −2.57 | 3.89 |
| 363168 | Lyzl4 | lysozyme-like 4 | −3.89 | −1.60 | 3.42 |
| 8 week post stroke | |||||
| Gene_ID | Gene_Symbol of Rats | Description | Fold Change | log2(Fc) | Volume |
| 25526 | Ptgds | prostaglandin D2 synthase | 10.31 | 3.33 | 10.35 |
| 24387 | Gfap | glial fibrillary acidic protein | 6.08 | 2.44 | 8.86 |
| 81818 | Vim | vimentin | 10.17 | 3.13 | 6.96 |
| 25211 | Lyz2 | lysozyme 2 | 5.49 | 2.35 | 6.32 |
| 56646 | Lgals1 | galectin 1 | 4.47 | 2.05 | 5.98 |
| 24587 | Nefh | neurofilament, heavy polypeptide | −3.17 | −1.64 | 5.41 |
| 314627 | Plk5 | polo-like kinase 5 | −2.60 | −1.37 | 3.46 |
| 315564 | Robo3 | roundabout guidance receptor 3 | −3.49 | −1.72 | 3.42 |
| 117023 | Kcns1 | potassium voltage-gated channel, modifier subfamily S, member 1 | −3.57 | −1.81 | 2.99 |
| 24805 | Syt2 | synaptotagmin 2 | −5.61 | −2.39 | 2.13 |
| Time | 1 Week | 4 Weeks | 8 Weeks | Time | 1 Week | 4 Weeks | 8 Weeks | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| GO | Brain Development (234 Genes) | Positive Regulation of Cell Population Proliferation (57 Genes) | Negative Regulation of Multicellular Organismal Process (73 Genes) | GO | Regulation of Postsynaptic Membrane Potential (58 Genes) | Ossification (24 Genes) | Extracellular Matrix Organization (25 Genes) | ||||||
| Gene | Fc | Gene | Fc | Gene | Fc | Gene | Fc | Gene | Fc | Gene | Fc | ||
| 1 | Tp53 | 2.75 | Igf1 | 2.35 | Cd44 | 6.13 | 1 | Gria1 | −4.58 | Igf1 | 2.35 | Col1a1 | 13.87 |
| 2 | Casp3 | 2.40 | Ptprc | 2.64 | Timp1 | 6.30 | 2 | Grin2a | −3.68 | Spp1 | 10.97 | Col3a1 | 13.73 |
| 3 | Rhoa | 2.25 | Timp1 | 7.38 | Ccl2 | 2.66 | 3 | Grm5 | −3.18 | Mmp2 | 2.17 | Lum | 2.92 |
| 4 | Gnai1 | -2.84 | Cxcl12 | 2.13 | Tgfb1 | 2.05 | 4 | Gabra1 | −26.42 | Col1a1 | 15.05 | Postn | −2.40 |
| 5 | Syt1 | -20.35 | Mmp2 | 2.17 | Spp1 | 9.09 | 5 | Gabrg2 | −12.33 | Sparc | 2.12 | Fbln1 | 6.40 |
| 6 | Grin2a | -3.68 | Tgfb1 | 2.09 | Col3a1 | 13.73 | 6 | Grin2b | −2.39 | Igf2 | 25.06 | Mmp14 | 6.88 |
| 7 | Ccr5 | 3.14 | Lgals3 | 36.53 | Vim | 10.17 | 7 | Gabrb2 | −6.06 | Tgfb1 | 2.09 | Fmod | 7.47 |
| 8 | Myc | 5.49 | Rac2 | 2.59 | Dcn | 6.11 | 8 | Gria2 | −8.97 | Igfbp5 | 2.65 | Tgfb1 | 2.05 |
| 9 | Aif1 | 10.36 | Hmox1 | 2.39 | Cd74 | 15.20 | 9 | Grm1 | −2.47 | Ctsk | 3.21 | Lgals3 | 21.69 |
| 10 | Gnao1 | -5.73 | Igf2 | 25.06 | Anxa1 | 10.74 | 10 | Grik1 | −3.15 | Twist1 | 2.44 | Col18a1 | 3.03 |
| GO | Memory (76 Genes) | Wound Healing (32 Genes) | Response to Estradiol (27 Genes) | GO | Locomotory Behavior (91 Genes) | Negative Regulation of Cell Population Proliferation (41 genes) | Aging (34 Genes) | ||||||
| Gene | Fc | Gene | Fc | Gene | Fc | Gene | Fc | Gene | Fc | Gene | Fc | ||
| 1 | Bdnf | −3.08 | Timp1 | 7.38 | Ccl2 | 2.66 | 1 | Gad1 | −3.54 | Igf1 | 2.35 | Ccl2 | 2.66 |
| 2 | Gria1 | −4.58 | Igf1 | 2.35 | Col1a1 | 13.87 | 2 | Scn1a | −9.28 | Cd44 | 7.93 | Tgfb1 | 2.05 |
| 3 | Grin2b | −2.39 | Sparc | 2.12 | Igf2 | 28.16 | 3 | Snap25 | −226.77 | Tgfb1 | 2.09 | Timp1 | 6.30 |
| 4 | Grin2a | −3.68 | Tgfb1 | 2.09 | Tgfb1 | 2.05 | 4 | Ppp1r1b | −4.62 | Sparc | 2.12 | Icam1 | 2.10 |
| 5 | Slc17a7 | −134.52 | Col1a1 | 15.05 | Vim | 10.17 | 5 | Adora2a | −2.65 | Tgfbr2 | 2.58 | Clu | 2.29 |
| 6 | Cnr1 | −3.65 | Dcn | 4.81 | Nqo1 | 2.16 | 6 | Mapk8 | −2.18 | Lgals3 | 36.53 | Gpx1 | 2.09 |
| 7 | Egr1 | −3.21 | Cd44 | 7.93 | Anxa1 | 10.74 | 7 | Gnao1 | −5.73 | Hmox1 | 2.39 | Nqo1 | 2.16 |
| 8 | Htr2a | −3.24 | Igf2 | 25.06 | Igfbp3 | 5.34 | 8 | Cacna1a | −5.10 | Bmp7 | 5.42 | Itgb2 | 2.36 |
| 9 | Snap25 | −226.77 | Col3a1 | 18.13 | Cd4 | 2.38 | 9 | Grm5 | −3.18 | Timp2 | 2.29 | Serping1 | 16.63 |
| 10 | Cck | −98.24 | Pf4 | 2.86 | Pdgfra | 2.33 | 10 | Grm1 | −2.47 | Gpc3 | 4.84 | Mbp | −3.01 |
| GO | Cellular Response to Drug (140 Genes) | Response to Estradiol (27 Genes) | Cellular Response to Lipid (43 Genes) | GO | Aging (138 Genes) | Cellular Response to Cytokine Stimulus (42 Genes) | Negative Regulation of Cell Population Proliferation (42 Genes) | ||||||
| Gene | Fc | Gene | Fc | Gene | Fc | Gene | Fc | Gene | Fc | Gene | Fc | ||
| 1 | Tp53 | 2.75 | Igf1 | 2.35 | Ccl2 | 2.66 | 1 | Tp53 | 2.75 | Cxcl12 | 2.13 | Cd44 | 6.13 |
| 2 | Mapk8 | −2.18 | Col1a1 | 15.05 | Icam1 | 2.10 | 2 | Igf1 | 52.19 | Cd44 | 7.93 | Lgals3 | 21.69 |
| 3 | Ccl2 | 22.32 | Igf2 | 25.06 | Cd40 | 2.28 | 3 | Stat3 | 2.55 | Mmp2 | 2.17 | Ccl2 | 2.66 |
| 4 | Myc | 5.49 | Mmp2 | 2.17 | Cxcl13 | 2.31 | 4 | Agt | −18.17 | Spp1 | 10.97 | Tgfb1 | 2.05 |
| 5 | Icam1 | 6.54 | Tgfb1 | 2.09 | Tgfb1 | 2.05 | 5 | Mmp9 | 17.34 | Col1a1 | 15.05 | Tgfbr2 | 2.19 |
| 6 | Igf1 | 52.19 | Vim | 13.22 | Il18 | 2.36 | 6 | Bdnf | −3.08 | Ifitm1 | 34.44 | Aif1 | 2.16 |
| 7 | Hmox1 | 52.12 | Bmp7 | 5.42 | Spp1 | 9.09 | 7 | Icam1 | 6.54 | Cd74 | 10.83 | Fcgr2b | 2.12 |
| 8 | Rhoa | 2.25 | Igfbp3 | 2.12 | Col1a1 | 13.87 | 8 | Ccl2 | 22.32 | Ifitm3 | 4.49 | Gpc3 | 3.97 |
| 9 | Il18 | 10.22 | Pdgfra | 2.66 | Cxcl16 | 6.41 | 9 | Fos | −2.45 | Gbp2 | 4.47 | Tspo | 2.79 |
| 10 | Gnai2 | 2.31 | Igfbp2 | 14.03 | Pf4 | 3.40 | 10 | Hras | −2.11 | Cyba | 4.20 | Inhba | −2.18 |
| GO | Positive Regulation of Apoptotic Process (181 Genes) | Aging (34 Genes) | Positive Regulation of Cell Migration (39 Genes) | GO | Regulation of Cation Channel Activity (73 Genes) | Response to Lipopopysaccharide (30 Genes) | Inflammatory Response (31 Genes) | ||||||
| Gene | Fc | Gene | Fc | Gene | Fc | Gene | Fc | Gene | Fc | Gene | Fc | ||
| 1 | Tp53 | 2.75 | Igf1 | 2.35 | Cxcl12 | 2.44 | 1 | Dlg4 | −8.41 | Igf1 | 2.35 | Ccl2 | 2.66 |
| 2 | Casp3 | 2.40 | Tgfb1 | 2.09 | Ccl2 | 2.66 | 2 | Cacna1a | −5.10 | Spp1 | 10.97 | Icam1 | 2.10 |
| 3 | Mapk8 | −2.18 | Timp1 | 7.38 | Icam1 | 2.10 | 3 | Cacng8 | −9.08 | Col1a1 | 15.05 | Timp1 | 6.30 |
| 4 | Myc | 5.49 | Mmp2 | 2.17 | Cxcl13 | 2.31 | 4 | Cacng7 | −4.68 | Fos | −2.66 | Cd44 | 6.13 |
| 5 | Tlr4 | 3.46 | Fos | -2.66 | Ccl19 | 3.69 | 5 | Shank1 | −28.89 | Tgfbr2 | 2.58 | Spp1 | 9.09 |
| 6 | Ccl2 | 22.32 | Clu | 2.19 | Col1a1 | 13.87 | 6 | Rasgrf1 | −14.10 | Sparc | 2.12 | Ccl19 | 3.69 |
| 7 | Mmp9 | 17.34 | Dcn | 4.81 | Anxa1 | 10.74 | 7 | Cacnb1 | −6.68 | Tgfb1 | 2.09 | Pf4 | 3.40 |
| 8 | Casp8 | 3.39 | Serping1 | 21.82 | Cd40 | 2.28 | 8 | Cacnb3 | −4.16 | Bmp7 | 5.42 | Itgb2 | 2.36 |
| 9 | Agt | −18.17 | Vim | 13.22 | Il18 | 2.36 | 9 | Cacnb2 | −2.62 | C3 | 8.25 | Il18 | 2.36 |
| 10 | Anxa5 | 2.05 | Ucp2 | 2.75 | Aif1 | 2.16 | 10 | Cacnb4 | −4.46 | Cyba | 4.20 | Cxcl13 | 2.31 |
| GO | Regulation of Synaptic Vesicle Exocytosis (59 Genes) | Response to Drug (64 Genes) | Cell Adhesion (44 genes) | GO | Cellular Response to Organic Cyclic Compound (166 Genes) | Nervous System Development (90 Genes) | Wound Healing (28 Genes) | ||||||
| Gene | Fc | Gene | Fc | Gene | Fc | Gene | Fc | Gene | Fc | Gene | Fc | ||
| 1 | Syt1 | −20.35 | Mmp2 | 2.17 | Cd44 | 6.13 | 1 | Casp3 | 2.40 | Cd44 | 7.93 | Timp1 | 6.30 |
| 2 | Stxbp1 | −7.15 | Igf1 | 2.35 | Spp1 | 9.09 | 2 | Stat3 | 2.55 | Igf1 | 2.35 | Col1a1 | 13.87 |
| 3 | Stx1a | −23.65 | Col1a1 | 15.05 | Cxcl12 | 2.44 | 3 | Igf1 | 52.19 | Mmp2 | 2.17 | Tgfb1 | 2.05 |
| 4 | Rab3a | −23.39 | Tgfb1 | 2.09 | Icam1 | 2.10 | 4 | Myc | 5.49 | Spp1 | 10.97 | Dcn | 6.11 |
| 5 | Cplx1 | −62.03 | Fos | -2.66 | Col3a1 | 13.73 | 5 | Bdnf | -3.08 | Cxcl12 | 2.13 | Cd44 | 6.13 |
| 6 | Syt2 | −8.04 | Hmox1 | 2.39 | Lamb2 | 2.15 | 6 | Ccl2 | 22.32 | Gfap | 8.65 | Col3a1 | 13.73 |
| 7 | Vamp1 | −14.02 | Igf2 | 25.06 | Itgb4 | 10.95 | 7 | Rhoa | 2.25 | Col3a1 | 18.13 | Igf2 | 28.16 |
| 8 | Cplx2 | −18.49 | Sparc | 2.12 | Thbs2 | 3.49 | 8 | Egr1 | −3.21 | Fos | −2.66 | Postn | −2.40 |
| 9 | Syp | −44.39 | Vim | 13.22 | Tgfbr2 | 2.19 | 9 | Icam1 | 6.54 | Vim | 13.22 | Vim | 10.17 |
| 10 | Camk2a | −139.81 | Gpx1 | 2.17 | Postn | -2.40 | 10 | Actb | 2.25 | Mmp14 | 6.52 | Pf4 | 3.40 |
| 1 week post stroke | |||
| Gene_Symbol | Description | Degrees | Fc |
| Dlg4 | discs large MAGUK scaffold protein 4 | 93 | −8.41 |
| Bdnf | brain-derived neurotrophic factor | 84 | −3.08 |
| Gria1 | glutamate ionotropic receptor AMPA type subunit 1 | 81 | −4.58 |
| Rhoa | ras homolog family member A | 76 | 2.25 |
| Mapk8 | mitogen-activated protein kinase 8 | 76 | −2.18 |
| Gng3 | G protein subunit gamma 3 | 74 | −5.48 |
| Grin2a | glutamate ionotropic receptor NMDA type subunit 2A | 73 | −3.68 |
| Gnb5 | G protein subunit beta 5 | 72 | −3 |
| Ptprc | protein tyrosine phosphatase, receptor type, C | 72 | 15.69 |
| Tp53 | tumor protein p53 | 70 | 2.75 |
| 4 weeks post stroke | |||
| Gene_Symbol | Description | Degrees | Fc |
| Cd44 | CD44 molecule (Indian blood group) | 17 | 7.93 |
| C1qb | complement component 1, q subcomponent, B chain | 15 | 2.88 |
| Fcgr2b | Fc fragment of IgG, low affinity IIb, receptor | 14 | 2.78 |
| Spp1 | secreted phosphoprotein 1 | 12 | 10.97 |
| Cd74 | CD74 molecule | 12 | 10.83 |
| C4a | complement component 4A (Rodgers blood group) | 12 | 7.5 |
| C1qa | complement component 1, q subcomponent, A chain | 12 | 2.72 |
| C3 | complement component 3 | 10 | 8.25 |
| Cd14 | CD14 molecule | 10 | 2.22 |
| Itgb2 | integrin subunit beta 2 | 10 | 3.42 |
| 8 weeks post stroke | |||
| Gene_Symbol | Description | Degrees | Fc |
| Cd44 | CD44 molecule (Indian blood group) | 14 | 6.13 |
| Fcgr2b | Fc fragment of IgG, low affinity IIb, receptor | 13 | 2.12 |
| C1qb | complement component 1, q subcomponent, B chain | 13 | 2.39 |
| Cd74 | CD74 molecule | 12 | 15.2 |
| C1qc | complement component 1, q subcomponent, C chain | 11 | 2.68 |
| Cd14 | CD14 molecule | 10 | 2.58 |
| C4a | complement component 4A (Rodgers blood group) | 10 | 5.42 |
| Spp1 | secreted phosphoprotein 1 | 10 | 9.09 |
| RT1-A2 | RT1 class Ia, locus A2 | 9 | 3.53 |
| Itgb2 | integrin subunit beta 2 | 9 | 2.36 |
| Gene Symbol of Rats | Forward Primer | Reverse Primer |
|---|---|---|
| Dlg4 | ATGCCTACCTGAGTGACAGC | CCCAGCAAGGATGAAGGAGA |
| Bdnf | AGGTTCGAGAGGTCTGACGA | GCTGTGACCCACTCGCTAAT |
| Gria1 | AAGCACGTGGGCTACTCCTA | GACGACGCTCACTCCAATGT |
| Rhoa | TCCATCGACAGCCCTGATAG | CTTTTCTTCCCGCGTCTAGC |
| Mapk8 | CTACAACCAACAGTAAGGAC | GTTTCCACTCCTCTATTGTG |
| Gng3 | GACCCCCGTTAACAGCACTA | GAAGTGGGCACAGGAGTGAT |
| Grin2a | TGGTGATGGTGAGATGGAGG | GTGTACCCCATGGATGCAAC |
| Gnb5 | GTCTGTCGCTATGCACACCA | AGCATCTCCACTGGGGTAGT |
| Ptprc | GATTGCCGATGAGGGTAGAC | CATCAACTGTCTCATCCCGG |
| Tp53 | GCCCATCCTTACCATCATCA | GCACGGGCATCCTTTAATTC |
| CD44 | GACAGAAACAGCACCAGTGC | CTTGGATGGTTGTTGTGGGC |
| C1qb | TGATGGCAAACCAGGCACTC | CCTTTTCGAAGCGAATGGCC |
| Fcgr2b | TGGGAGTGATTTCTGACTGG | GCTACAATCGTCAATACCGG |
| Spp1 | CAGGAGTCCGATGAGGCTAT | CCTCATGGCTGTGAAACTCG |
| Cd74 | CAGGCCACCACTGCTTACTT | TGTGCTTCAGATTCTCCGGG |
| C4a | GCCCAGCAAGTATCAGTGCC | CAGTCAGGGTAGGGGCCAAA |
| C1qa | CTCAGCTATTCGGCAGAACC | CCTTCTCAATCCACACCTCG |
| C3 | CTTCATGGACTGCTGCAACT | TCTGCCACACAGATCCCTTT |
| Cd14 | TTTCTTGCAAACAGGTCGGC | AGCAGTATCCCGCAGTGAAT |
| Itgb2 | ACACCCATCCCGAGAAGCTG | CCATCGTTGGGGGTCAGGAT |
| C1qc | GATGGGCATGATGGACTTCA | GTGTGTTGTAATCCCCCTGA |
| RT1-A2 | CCAGGACATGGAGCTTGTGG | CTGGAGCAGGGGTGTAGTCA |
| GAPDH | ATCACCATCTTCCAGGAGCG | GATGGCATGGACTGTGGTCA |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, I.-A.; Yun, J.H.; Kim, J.-H.; Kim, H.Y.; Choi, D.-H.; Lee, J. Sequential Transcriptome Changes in the Penumbra after Ischemic Stroke. Int. J. Mol. Sci. 2019, 20, 6349. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20246349
Choi I-A, Yun JH, Kim J-H, Kim HY, Choi D-H, Lee J. Sequential Transcriptome Changes in the Penumbra after Ischemic Stroke. International Journal of Molecular Sciences. 2019; 20(24):6349. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20246349
Chicago/Turabian StyleChoi, In-Ae, Ji Hee Yun, Ji-Hye Kim, Hahn Young Kim, Dong-Hee Choi, and Jongmin Lee. 2019. "Sequential Transcriptome Changes in the Penumbra after Ischemic Stroke" International Journal of Molecular Sciences 20, no. 24: 6349. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20246349