Early Pregnancy Human Decidua is Enriched with Activated, Fully Differentiated and Pro-Inflammatory Gamma/Delta T Cells with Diverse TCR Repertoires

 , ,
, ,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
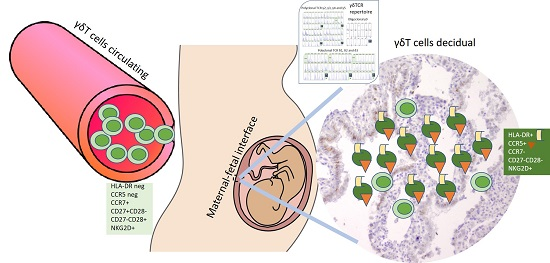
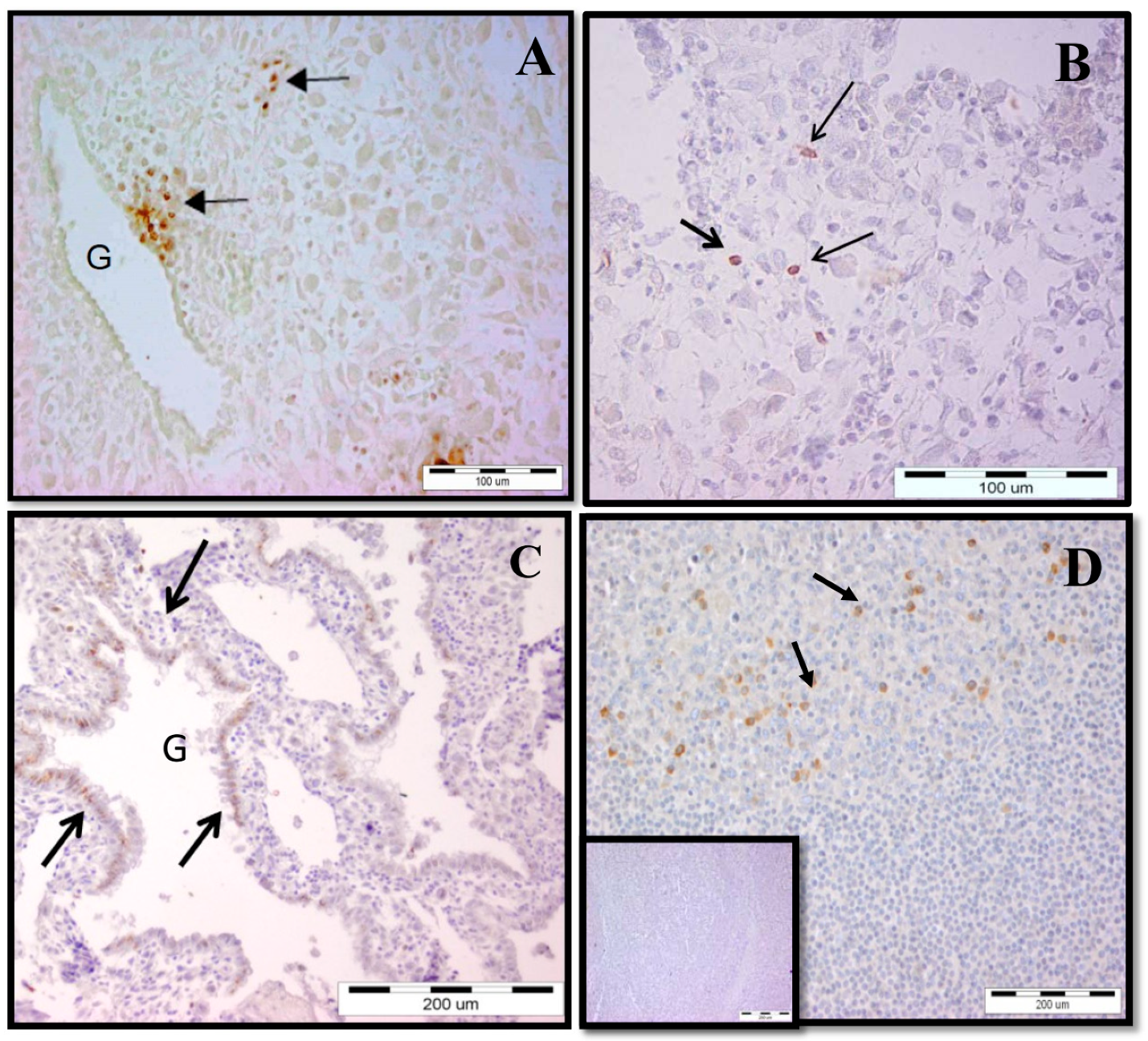
2.1. Increased γδ T-Cell Number at the MFI during Human Early Pregnancy
2.2. Accumulation of γδ T Cells at the MFI Is Restricted to the Vδ1 γδ T-Cell Subset
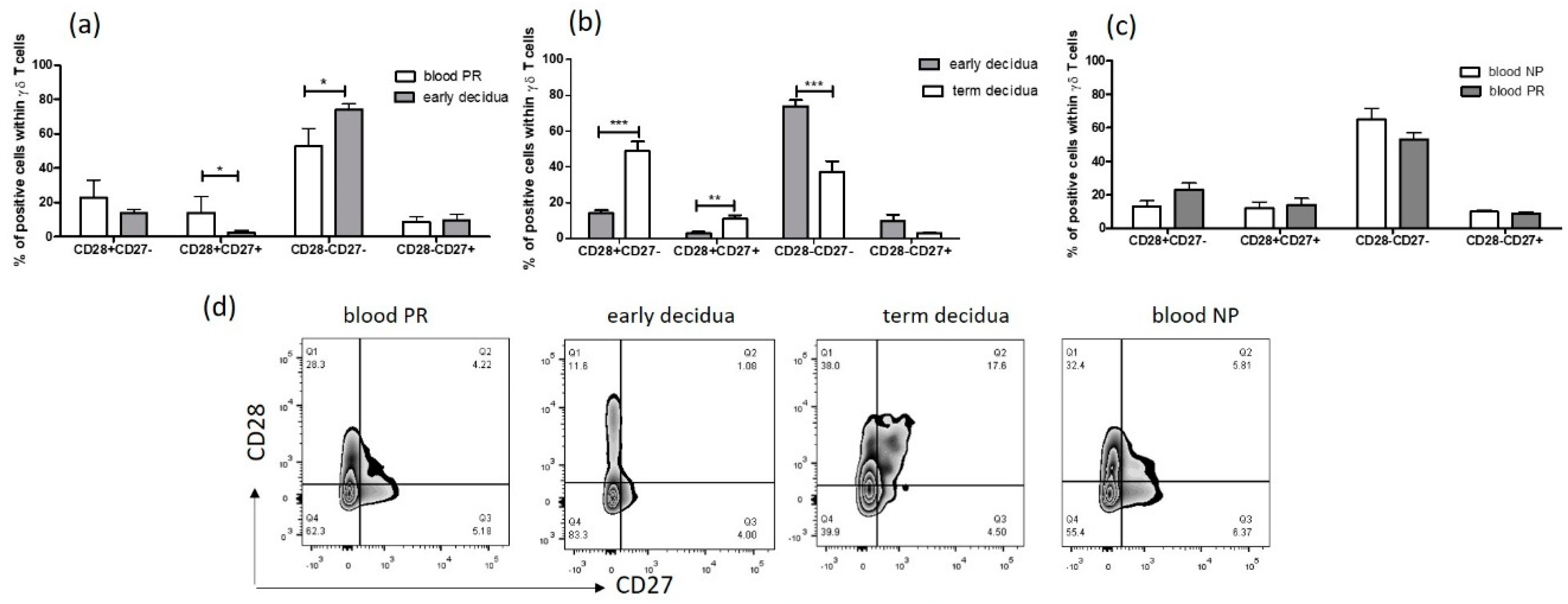
2.3. Early Pregnancy Decidua is Enriched with Activated, Terminally Differentiated and Pro-inflammatory γδ T Cells
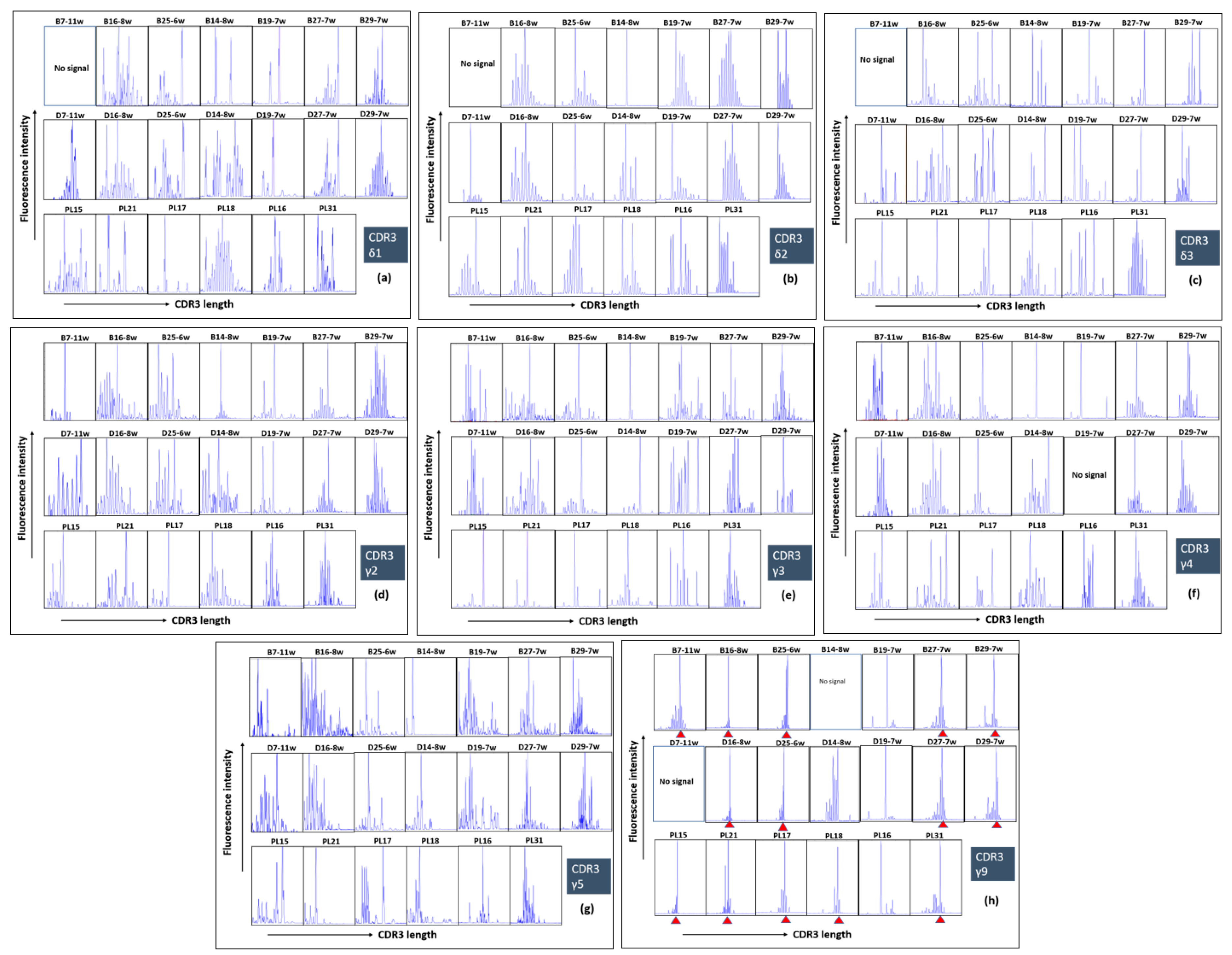
2.4. Polyclonal Profiles of All δ and γ Chains and Highly Restricted (Oligoclonal) Repertoire of γ9 Chain of γδ T Cells during Human Pregnancy
3. Discussion
4. Materials and Methods
4.1. Study Population and Samples
4.2. Mononuclear Cells Isolation (PBMC, DMC)
4.3. Cell Labelling and FACS
4.4. Immunohistochemistry (IHC)
4.5. Spectratyping
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| TCR | T cell receptor |
| MFI | Maternal–fetal interface |
| DALT | Decidua-associated lymphoid tissue |
| EVT | Extravillous cytotrophoblast |
| MHC | Major histocompatibility complex |
| CDR3 | Complementary-determining region 3 |
| NK cells | Natural killer cells |
| APC | Antigen-presenting cells |
| mAb | Monoclonal antibody |
References
- Mor, G.; Aldo, P.; Alvero, A.B. The unique immunological and microbial aspects of pregnancy. Nat. Rev. Immunol. 2017, 17, 469–482. [Google Scholar] [CrossRef] [PubMed]
- Whitelaw, P.F.; Croy, B.A. Granulated lymphocytes of pregnancy. Placenta 1996, 17, 533–543. [Google Scholar] [CrossRef]
- King, A.; Balendran, N.; Wooding, P.; Carter, N.P.; Loke, Y.W. CD3-leukocytes present in the human uterus during early placentation: Phenotypic and morphologic characterization of the CD56++ population. Dev. Immunol. 1991, 1, 169–190. [Google Scholar] [CrossRef] [PubMed]
- Chien, Y.; Konigshofer, Y. Antigen recognition by gamma delta T cells. Immunol. Rev. 2007, 215, 46–58. [Google Scholar] [CrossRef] [PubMed]
- Hayday, A.C. Gamma delta cells: A right time and a right place for a conserved third way of protection. Annu. Rev. Immunol. 2000, 18, 975–1026. [Google Scholar] [CrossRef] [PubMed]
- Holtmeier, W.; Kabelitz, D. Gamma delta T cells link innate and adaptive immune responses. Chem. Immunol. Allergy 2005, 86, 151–183. [Google Scholar] [PubMed]
- Hayday, A.C.; Spencer, J. Barrier immunity. Semin. Immunol. 2009, 21, 99–100. [Google Scholar] [CrossRef]
- Wang, L.; Kamath, A.; Das, H.; Li, L.; Bukowski, J.F. Antibacterial effect of human Vgamma2Vdelta2 T cells in vivo. J. Clin. Investig. 2001, 108, 1349–1357. [Google Scholar] [CrossRef]
- Pennington, D.J.; Vermijlen, D.; Wise, E.L.; Clarke, S.L.; Tigelaar, R.E.; Hayday, A.C. The integration of conventional and unconventional T cells that characterizes cell-mediated responses. Adv. Immunol. 2005, 87, 27–59. [Google Scholar]
- Toulon, A.; Breton, L.; Taylor, K.R.; Tenenhaus, M.; Bhavsar, D.; Lanigan, C.; Rudolph, R.; Jameson, J.; Havran, W.L. A role for human skin–resident T cells in wound healing. J. Exp. Med. 2009, 206, 743–750. [Google Scholar] [CrossRef]
- Casorati, G.; De Libero, G.; Lanzavecchia, A.; Migone, N. Molecular analysis of human γ/δ clones from thymus and peripheral blood. J. Exp. Med. 1989, 170, 1521–1535. [Google Scholar] [CrossRef]
- Carding, S.R.; Egan, P.J. γδ T cells: Functional plasticity and heterogeneity. Nat. Rev. Immunol. 2002, 2, 336–345. [Google Scholar] [CrossRef] [PubMed]
- Mincheva-Nilsson, L.; Hammarstrom, S.; Hammarstrom, M.L. Human decidual leukocytes from early pregnancy contain high numbers of gamma delta+ cells and show selective down-regulation of alloreactivity. J. Immunol. 1992, 149, 2203–2211. [Google Scholar]
- Ditzian-Kadanoff, R.; Garon, J.; Verp, M.S.; Zilberstein, M. Gamma delta T cells in human decidua. Am. J. Obstet. Gynecol. 1993, 168, 831–836. [Google Scholar] [CrossRef]
- Vassiliadou, N.; Bulmer, J.N. Quantitative analysis of T lymphocyte subsets in pregnant and nonpregnant human endometrium. Biol. Reprod. 1996, 55, 1017–1022. [Google Scholar] [CrossRef] [PubMed]
- Polgar, B.; Barakonyi, A.; Xynos, I.; Szekeres-Bartho, J. The role of gamma/delta T cell receptor positive cells in pregnancy. Am. J. Reprod. Immunol. 1999, 41, 239–244. [Google Scholar] [CrossRef] [PubMed]
- Psarra, K.; Kapsimali, V.; Tarassi, K.; Dendrinos, S.; Athanasiadis, T.; Botsis, D.; Kreatsas, G.; Papasteriades, C. TCR gamma delta+ T lymphocytes in unexplained recurrent spontaneous abortions. Am. J. Reprod. Immunol. 2001, 45, 6–11. [Google Scholar] [CrossRef]
- Gu, Y.; Hu, Y.; Hu, K.; Liao, W.; Zheng, F.; Yu, X.; Huang, H. Rapamycin together with TGF-β1, IL-2 and IL-15 induces the generation of functional regulatory γδT cells from human peripheral blood mononuclear cells. J. Immunol. Methods 2014, 402, 82–87. [Google Scholar] [CrossRef]
- Appay, V.; Dunbar, P.R.; Callan, M.; Klenerman, P.; Gillespie, G.M.; Papagno, L.; Ogg, G.S.; King, A.; Lechner, F.; Spina, C.A.; et al. Memory CD81 T cells vary in differentiation phenotype in different persistent virus infections. Nat. Med. 2002, 8, 379–385. [Google Scholar] [CrossRef]
- Morita, C.T.; Jin, C.; Sarikonda, G.; Wang, H. Nonpeptide antigens, presentation mechanisms, and immunological memory of human Vgamma2Vdelta2 T cells: Discriminating friend from foe through the recognition of prenyl pyrophosphate antigens. Immunol. Rev. 2007, 215, 59–76. [Google Scholar] [CrossRef]
- Sallusto, F.; Lenig, D.; Forster, R.; Lipp, M.; Lanzavecchia, A. Two subsets of memory T lymphocytes with distinct homing potential and effector function. Nature 1999, 401, 708–712. [Google Scholar] [CrossRef]
- Glatzel, A.; Wesch, D.; Schiemann, F.; Brandt, E.; Janssen, O.; Kabelitz, D. Patterns of chemokine receptor expression on peripheral blood gd T lymphocytes: Strong expression of CCR5 is a selective feature of Vd2/Vg9 gd T cells. J. Immunol. 2002, 168, 4920–4929. [Google Scholar] [CrossRef] [PubMed]
- Sallusto, F.; Lenig, D.; Mackay, C.R.; Lanzavecchia, A. Flexible programs of chemokine receptor expression on human polarized T helper 1 and 2 lymphocytes. J. Exp. Med. 1998, 187, 875–883. [Google Scholar] [CrossRef] [PubMed]
- Loetscher, P.; Uguccioni, M.; Bordoli, L.; Baggiolini, M.; Moser, B.; Chizzolini, C.; Dayer, J.M. CCR5 is characteristic of Th1 lymphocytes. Nature 1998, 391, 344. [Google Scholar] [CrossRef] [PubMed]
- Haller, H.; Radillo, O.; Rukavina, D.; Tedesco, F.; Candussi, G.; Petrović, O.; Randić, L. An immunohistochemical study of leucocytes in human endometrium, first and third trimester basal decidua. J. Reprod. Immunol. 1993, 23, 41–49. [Google Scholar] [CrossRef]
- Fan, D.X.; Duan, J.; Li, M.Q.; Xu, B.; Li, D.J.; Jin, L.P. The decidual gamma-delta T cells up-regulate the biological functions of trophoblasts via IL-10 secretion in early human pregnancy. Clin. Immunol. 2011, 141, 284–292. [Google Scholar] [CrossRef] [PubMed]
- Tilburgs, T.; van der Mast, B.; Nagtzaam, N.; Roelen, D.; Scherjon, S.; Claas, F. Expression of NK cell receptors on decidual T cells in human pregnancy. J. Reprod. Immunol. 2009, 80, 22–32. [Google Scholar] [CrossRef] [PubMed]
- Mincheva-Nilsson, L.; Baranov, V.; Yeung, M.M.; Hammarström, S.; Hammarström, M.-L. Immunomorphologic studies of human decidua-associated lymphoid cells in normal early pregnancy. J. Immunol. 1994, 152, 2020–2032. [Google Scholar] [PubMed]
- Suzuki, T.; Hiromatsu, K.; Ando, Y.; Okamoto, T.; Tomoda, Y.; Yoshikai, Y. Regulatory role of gamma delta T cells in uterine intraepithelial lymphocytes in maternal antifetal immune response. J. Immunol. 1995, 154, 4476–4484. [Google Scholar] [PubMed]
- Fox, A.; Lee, C.; Brandon, M.R.; Meeusen, E.N. Effects of pregnancy on lymphocytes within sheep uterine interplacentomal epithelium. Am. J. Reprod. Immunol. 1998, 40, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Carson, D.D.; Bagchi, I.; Dey, S.K.; Enders, A.C.; Fazleabas, A.T.; Lessey, B.A.; Yoshinaga, K. Embryo implantation. Dev. Biol. 2000, 223, 217–237. [Google Scholar] [CrossRef] [PubMed]
- Hempstock, J.; Cindrova-Davies, T.; Jauniaux, E.; Burton, G. Endometrial glands as a source of nutrients, growth factors and cytokines during the first trimester of human pregnancy: A morphological and immunohistochemical study. Reprod. Biol. Endocrinol. 2004, 2, 58. [Google Scholar] [CrossRef] [PubMed]
- Rachmilewitz, J.; Riely, G.; Tykocinski, M.L. Placental protein 14 functions as a direct T-cell inhibitor. Cell Immunol. 1999, 191, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Moser, G.; Weiss, G.; Gauster, M.; Sundl, M.; Huppertz, B. Evidence from the very beginning: Endoglandular trophoblasts penetrate and replace uterine glands in situ and in vitro. Hum. Reprod. 2015, 30, 2747–2757. [Google Scholar] [CrossRef] [PubMed]
- Mincheva-Nilsson, L.; Kling, M.; Hammarström, S.; Nagaeva, O.; Sundqvist, K.-G.; Hammarström, M.-L.; Baranov, V. γδT cells of human early pregnancy decidua: Evidence for local proliferation, phenotypic heterogeneity, and extrathymic differentiation. J. Immunol. 1997, 159, 3266–3277. [Google Scholar] [PubMed]
- Christmas, S.E.; Brew, R.; Deniz, G.; Taylor, J.J. T-cell receptor heterogeneity of gamma delta T-cell clones from human female reproductive tissues. Immunology 1993, 78, 436–443. [Google Scholar] [PubMed]
- Heyborne, K.; Fu, Y.-X.; Nelson, A.; Farr, A.; O’Brien, B. Recognition of trophoblasts by γδT cells. J. Immunol. 1994, 153, 2918–2926. [Google Scholar] [PubMed]
- Szekeres-Bartho, J.; Barakonyi, A.; Polgar, B.; Par, G.; Faust, Z.; Palkovics, T.; Szereday, L. The role of gamma/delta T cells in progesterone-mediated immunomodulation during pregnancy: A review. Am. J. Reprod. Immunol. 1999, 42, 44–48. [Google Scholar] [CrossRef] [PubMed]
- Morita, C.T.; Parker, C.; Brenner, M.B.; Band, H. TCR usage and functional capabilities of human gamma delta T cells at birth. J. Immunol. 1994, 153, 3979–3988. [Google Scholar] [PubMed]
- Vermijlen, D.; Brouwer, M.; Donner, C.; Liesnard, C.; Tackoen, M.; Van Rysselberge, M.; Twité, N.; Goldman, M.; Marchant, A.; Willems, F. Human cytomegalovirus elicits fetal gamma delta T cell responses in utero. J. Exp. Med. 2010, 207, 807–821. [Google Scholar] [CrossRef]
- Moens, E.; Brouwer, M.; Dimova, T.; Goldman, M.; Willems, F.; Vermijlen, D. IL-23R and TCR signaling drives the generation of neonatal Vgamma9Vdelta2 T cells expressing high levels of cytotoxic mediators and producing IFN-gamma and IL-17. J. Leukoc. Biol. 2011, 89, 743–752. [Google Scholar] [CrossRef] [PubMed]
- Groh, V.; Rhinehart, R.; Secrist, H.; Bauer, S.; Grabstein, K.H.; Spies, T. Broad tumor-associated expression and recognition by tumor-derived γδT cells of MICA and MICB. Proc. Natl. Acad. Sci. USA 1999, 96, 6879–6884. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.W. Comparative biology of γδT cells. Sci. Prog. 2002, 85, 347–358. [Google Scholar] [CrossRef] [PubMed]
- Groh, V.; Steinle, A.; Bauer, S.; Spies, T. Recognition of stress-induced MHC molecules by intestinal epithelial γδT cells. Science 1998, 279, 1737–1740. [Google Scholar] [CrossRef]
- Bauer, S.; Groh, V.; Wu, J.; Steinle, A.; Phillips, J.H.; Lanier, L.L.; Spies, T. Activation of NK cells and T cells by NKG2D, a receptor for stress-inducible MICA. Science 1999, 285, 727–729. [Google Scholar] [CrossRef] [PubMed]
- Mincheva-Nilsson, L.; Nagaeva, O.; Chen, T.; Stendahl, U.; Antsiferova, J.; Mogren, I.; Hernestål, J.; Baranov, V. Placenta derived soluble MHC class I chain-related molecules down regulate NKG2D receptor on peripheral blood mononuclear cells during human pregnancy: A possible novel immune escape mechanism for fetal survival. J. Immunol. 2006, 176, 3585–3592. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Groh, V.; Spies, T. T cell antigen receptor engagement and specificity in the recognition of stress-inducible MHC class I-related chains by human epithelial gd T cells. J. Immunol. 2002, 169, 1236–1240. [Google Scholar] [CrossRef] [PubMed]
- Bessoles, S.; Ni, M.; Garcia-Jimenez, S.; Sanchez, F.; Lafont, V. Role of NKG2D and its ligands in the anti-infectious activity of Vg9Vd2 T cells against intracellular bacteria. Eur. J. Immunol. 2011, 41, 1619–1628. [Google Scholar] [CrossRef]
- Szereday, L.; Barakonyi, A.; Miko, E.; Varga, P.; Szekeres-Bartho, J. γ/δ T-cell subsets, NKG2A expression and apoptosis of Vδ2+ t cells in pregnant women with or without risk of premature pregnancy termination. Am. J. Reprod. Immunol. 2003, 50, 490–496. [Google Scholar] [CrossRef]
- Red-Horse, K.; Drake, P.; Gunn, M.D.; Fisher, S.J. Chemokine ligand and receptor expression in the pregnant uterus: Reciprocal patterns in complementary cell subsets suggest functional roles. Am. J. Pathol. 2001, 159, 2199–2213. [Google Scholar] [CrossRef]
- Tilburgs, T.; Schonkeren, D.; Eikmans, M.; Nagtzaam, N.M.; Datema, G.; Swings, G.M.; Prins, F.; van Lith, J.M.; van der Mast, B.J.; Roelen, D.L.; et al. Human decidual tissue contains differentiated CD8+ effector-memory t cells with unique properties. J. Immunol. 2010, 185, 4470–4477. [Google Scholar] [CrossRef]
- Saito, S.; Nishikawa, K.; Morii, T.; Narita, N.; Enomoto, M.; Ito, A.; Ichijo, M. A study of CD45RO, CD45RA and CD29 antigen expression on human decidual T cells in an early stage of pregnancy. Immunol. Lett. 1994, 40, 193–197. [Google Scholar] [CrossRef]
- Tilburgs, T.; Strominger, J. CD8+ effector T cells at the fetal-maternal interface, balancing fetal tolerance and antiviral immunity. Am. J. Reprod. Immunol. 2013, 69, 395–407. [Google Scholar] [CrossRef]
- Slukvin, I.I.; Merkulova, A.A.; Vodyanik, M.A.; Chernyshov, V.P. Differential expression of CD45RA and CD45RO molecules on human decidual and peripheral blood lymphocytes at early stage of pregnancy. Am. J. Reprod. Immunol. 1996, 35, 16–22. [Google Scholar] [PubMed]
- Koizumi, H.; Liu, C.; Zheng, L.M.; Joad, S.V.; Bayne, N.K.; Holoshitz, J.; Young, J.D. Expression of perforin and serine esterases by human gamma delta T cells. J. Exp. Med. 1991, 173, 499–502. [Google Scholar] [CrossRef] [PubMed]
- Davey, M.; Willcox, C.; Joyce, S.; Ladell, K.; Kasatskaya, S.; McLaren, J.; Hunter, S.; Salim, M.; Mohammed, F.; Price, D.; Chudakov, D.; Willcox, B. Clonal selection in the human Vd1 T cell repertoire indicates gd TCR-dependent adaptive immune surveillance. Nat. Commun. 2017, 8, 14760. [Google Scholar]
- Hunter, S.; Willcox, C.; Davey, M.C.; Kasatskaya, S.A.; Jeffery, H.C.; Chudakov, D.M.; Oo, Y.H.; Willcox, B.E. Human liver infiltrating γδ T cells are composed of clonally expanded circulating and tissue-resident populations. J. Hepatol. 2018, 69, 654–665. [Google Scholar] [CrossRef]
- Kallikourdis, M.; Andersen, K.; Welch, K.; Betz, A. Alloantigen-enhanced accumulation of CCR5+ ‘effector’ regulatory T cells in the gravid uterus. Proc. Natl. Acad. Sci. USA 2007, 104, 594–599. [Google Scholar] [CrossRef]
- Carlino, C.; Stabile, H.; Morrone, S.; Bulla, R.; Soriani, A.; Agostinis, C.; Bossi, F.; Mocci, C.; Sarazani, F.; Tedesco, F.; et al. Recruitment of circulating NK cells through decidual tissues: A possible mechanism controlling NK cell accumulation in the uterus during early pregnancy. Blood 2008, 111, 3108–3115. [Google Scholar] [CrossRef]
- Santoni, A.; Carlino, C.; Stabile, H.; Gismondi, A. Mechanisms underlying recruitment and accumulation of decidual NK cells in uterus during pregnancy. Am. J. Reprod. Immunol. 2008, 59, 417–424. [Google Scholar] [CrossRef]
- Kitaya, K.; Nakayama, T.; Okubo, T.; Kuroboshi, H.; Fushiki, S.; Honjo, H. Expression of Macrophage Inflammatory Protein-1beta in human endometrium: Its role in endometrial recruitment of natural killer cells. J. Clin. Endocrinol. Metab. 2003, 88, 1809–1814. [Google Scholar] [CrossRef] [PubMed]
- Nelson, P.; Krensky, A. Chemokines, chemokines receptors and allograft rejection. Immunity 2001, 14, 377–386. [Google Scholar] [CrossRef]
- Ramhorst, R.; Patel, R.; Corigliano, A.; Etchepareborda, J.J.; Fainboim, L.; Schust, D. Induction of maternal tolerance to fetal alloantigens by RANTES production. Am. J. Reprod. Immunol. 2006, 56, 302–311. [Google Scholar] [CrossRef]
- Kemp, M.W. Preterm birth, intrauterine infection, and fetal inflammation. Front. Immunol. 2014, 5, 574. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Mossman, T.R.; Guilbert, T.; Tuntipopipat, S.; Wegmann, T.G. Synthesis of T helper 2-type cytokines at the maternal-fetal interface. J. Immunol. 1993, 151, 4562–4573. [Google Scholar] [PubMed]
- Fallon, P.G.; Jolin, H.E.; Smith, P.; Emson, C.L.; Townsend, M.J.; Fallon, R.; Smith, P.; McKenzie, A.N. Il-4 induces characteristic Th2 responses even in the combined absence of IL-5, IL-9, and IL-13. Immunity 2002, 17, 7–17. [Google Scholar] [CrossRef]
- Rugeles, M.T.; Shearer, G. Alloantigen recognition in utero: Dual advantage for the fetus? Trends Immunol. 2004, 7, 348–352. [Google Scholar] [CrossRef]
- Abrahams, V.; Visitin, I.; Aldo, P.; Guller, S.; Romero, R.; Mor, G. A role for TLRs in the regulation of immune cell migration by first trimester trophoblast cells. J. Immunol. 2005, 176, 8096–8104. [Google Scholar] [CrossRef]
- Zeng, W.; Liu, Z.; Liu, X.; Zhang, S.; Khanniche, A.; Zheng, Y.; Ma, X.; Yu, T.; Tian, F.; Liu, X.R.; et al. Distinct transcriptional and alternative splicing signatures of decidual CD4+ T cells in early human pregnancy. Front. Immunol. 2017, 8, 682. [Google Scholar] [CrossRef]
- Saito, S.; Nishikawa, K.; Morii, T.; Narita, N.; Enomoto, M.; Ito, A.; Ichijo, M. Th17 cells and regulatory T cells: New light on pathophysiology of preeclampsia. Immunol. Cell Biol. 2010, 88, 615–617. [Google Scholar] [CrossRef]
- Erkers, T.; Stikvoort, A.; Uhlin, M. Lymphocytes in placental tissues: Immune regulation and translational possibilities for immunotherapy. Stem Cells Int. 2017. [Google Scholar] [CrossRef] [PubMed]
- Otun, H.A.; Lash, G.; Innes, B.A.; Bulmer, J.N.; Naruse, K.; Hannon, T.; Searle, R.F.; Robson, S.C. Effect of Tumour Necrosis Factor-α in combination with Interferon-γ on first trimester extravillous trophoblast invasion. J. Reprod. Immunol. 2011, 88, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Bauer, S.; Pollheimer, J.; Hartmann, J.; Husslein, P.; Aplin, J.D.; Knöfler, M. Tumor Necrosis Factor-alpha inhibits trophoblast migration through elevation of plasminogen activator inhibitor-1 in first-trimester villous explant cultures. J. Clin. Endocrinol. Metab. 2004, 89, 812–822. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Dutz, J.; MacCalman, C.; Yong, P.; Tan, P.; Dadelszen, P. Decidual NK cells alter in vitro first trimester extravillous cytotrophoblast migration: A role for ifn-γ. J. Immunol. 2006, 177, 8522–8530. [Google Scholar] [CrossRef] [PubMed]
- Itohara, S.; Farra, A.G.; Lafaille, J.J.; Bonneville, M.; Takagaki, Y.; Haas, W.; Tonegawa, S. Homing of a gd thymocyte subset with homogeneous T-cell receptors to mucosal epithelia. Nature 1990, 343, 754. [Google Scholar] [CrossRef] [PubMed]
- Nandi, D.; Allison, J.P. Phenotypic analysis and gd T cell receptor repertoire of murine T cells associated with the vaginal epithelium. J. Immunol 1991, 147, 1773–1778. [Google Scholar] [PubMed]
- Bukowski, J.F.; Morita, C.T.; Brenner, M.B. Human γδ T cells recognize alkylamines derived from microbes, edible plants, and tea: Implications for innate immunity. Immunity 1999, 11, 57–65. [Google Scholar] [CrossRef]
- Dimova, T.; Brouwer, M.; Gosselin, F.; Tassignon, J.; Leo, O.; Donner, C.; Marchant, A.; Vermijlen, D. Effector Vγ9Vδ2 T cells dominate the human fetal γδ T-cell repertoire. Proc. Natl. Acad. Sci. USA 2015, 112, E556–E565. [Google Scholar] [CrossRef]
- Delfau, M.H.; Hance, A.; Lecossier, D.; Vilmer, E.; Grandchamp, B. Restricted diversity of Vgamma9-JP rearrangements in unstimulated human gamma/delta T lymphocytes. Eur. J. Immunol. 1992, 22, 2437–2443. [Google Scholar] [CrossRef]
- Sherwood, A.M.; Desmarais, C.; Livingston, R.J.; Andriesen, J.; Haussler, M.; Carlson, C.S.; Robins, H. Deep sequencing of the human TCRγ and TCRβ repertoires suggests that TCRβ rearranges after αβ and γδ T cell commitment. Sci. Transl. Med. 2011, 3, 90ra61. [Google Scholar] [CrossRef]
- McVay, L.D.; Carding, S. Extrathymic origin of human gamma delta T cells during fetal development. J. Immunol. 1996, 157, 2873–2882. [Google Scholar] [PubMed]
- Ryan, P.; Sumaria, N.; Holland, C.; Bradford, C.; Izotova, N.; Grandjean, C.; Jawad, A.; Bergmeier, L.; Pennington, D. Heterogeneous yet stable Vδ2(+) T-cell profiles define distinct cytotoxic effector potentials in healthy human individuals. Proc. Natl. Acad. Sci. USA 2016, 113, 14378–14383. [Google Scholar] [CrossRef] [PubMed]
- Davey, M.S.; Willcox, C.; Hunter, S.; Oo, Y.H.; Willcox, B.E. Vδ2 + T cells—Two subsets for the price of one. Front. Immunol. 2018, 9, 2106. [Google Scholar] [CrossRef] [PubMed]
- Nagaeva, O.; Mincheva-Nilsson, L.M. Dominant IL-10 and TGF-β mRNA expression in γδ T cells of human early pregnancy decidua suggests immunoregulatory potential. Am. J. Reprod. Immunol. 2002, 48, 9–17. [Google Scholar] [CrossRef]
- Barakonyi, A.; Kovacs, K.; Miko, E.; Szereday, L.; Varga, P.; Szekeres-Bartho, J. Recognition of nonclassical HLA class I antigens by gd T cells during pregnancy. J. Immunol. 2002, 168, 2683–2688. [Google Scholar] [CrossRef] [PubMed]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Terzieva, A.; Dimitrova, V.; Djerov, L.; Dimitrova, P.; Zapryanova, S.; Hristova, I.; Vangelov, I.; Dimova, T. Early Pregnancy Human Decidua is Enriched with Activated, Fully Differentiated and Pro-Inflammatory Gamma/Delta T Cells with Diverse TCR Repertoires. Int. J. Mol. Sci. 2019, 20, 687. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20030687
Terzieva A, Dimitrova V, Djerov L, Dimitrova P, Zapryanova S, Hristova I, Vangelov I, Dimova T. Early Pregnancy Human Decidua is Enriched with Activated, Fully Differentiated and Pro-Inflammatory Gamma/Delta T Cells with Diverse TCR Repertoires. International Journal of Molecular Sciences. 2019; 20(3):687. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20030687
Chicago/Turabian StyleTerzieva, Antonia, Violeta Dimitrova, Lyubomir Djerov, Petya Dimitrova, Silvina Zapryanova, Iana Hristova, Ivaylo Vangelov, and Tanya Dimova. 2019. "Early Pregnancy Human Decidua is Enriched with Activated, Fully Differentiated and Pro-Inflammatory Gamma/Delta T Cells with Diverse TCR Repertoires" International Journal of Molecular Sciences 20, no. 3: 687. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20030687