Circadian Regulation of Alternative Splicing of Drought-Associated CIPK Genes in Dendrobium catenatum (Orchidaceae)


Abstract
:1. Introduction
2. Results
2.1. Genome-Wide Identification of CIPK Gene Family Members in D. catenatum
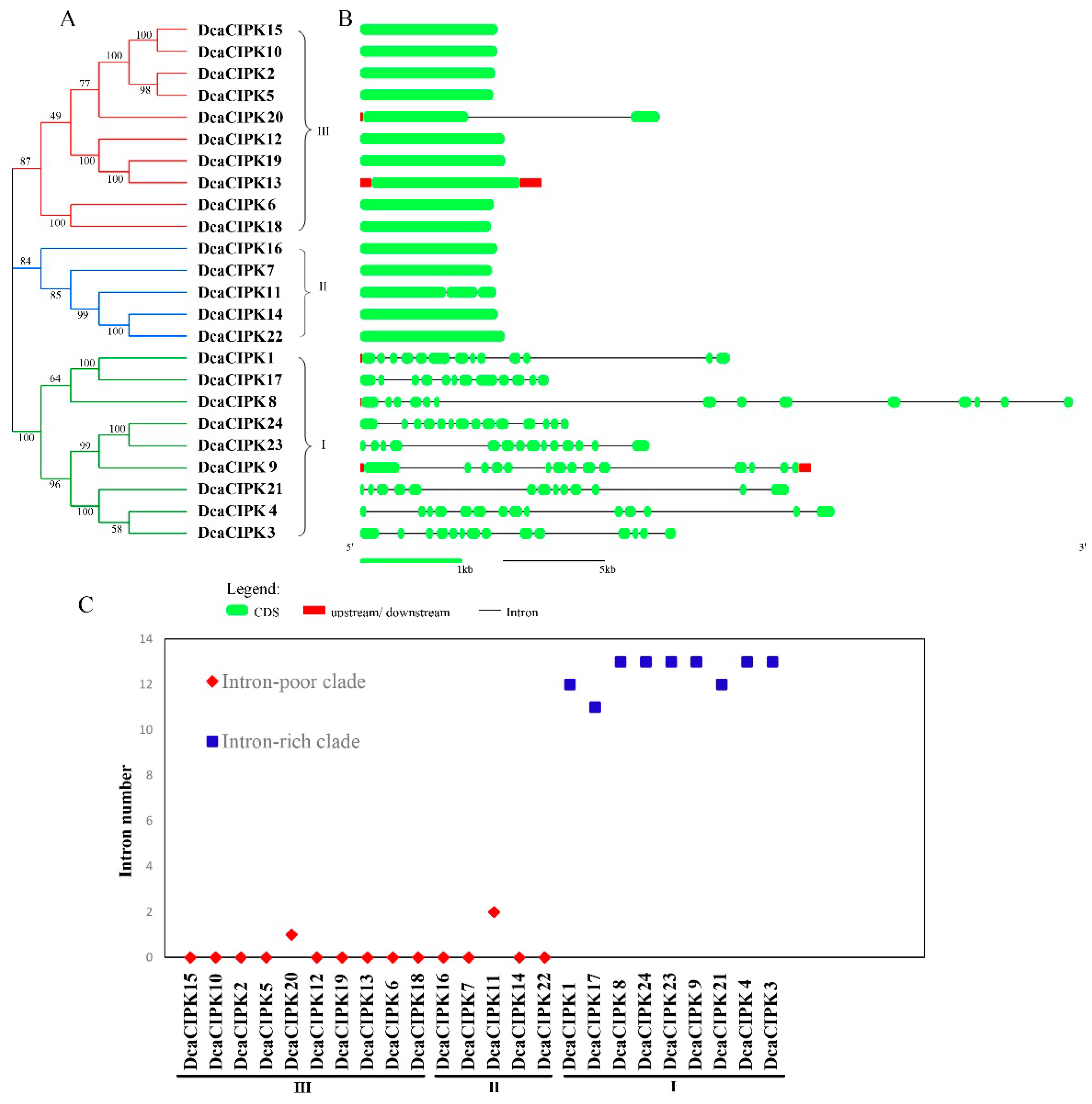
2.2. Phylogenetic Analysis and Structure of the D. catenatum CIPK Gene Family
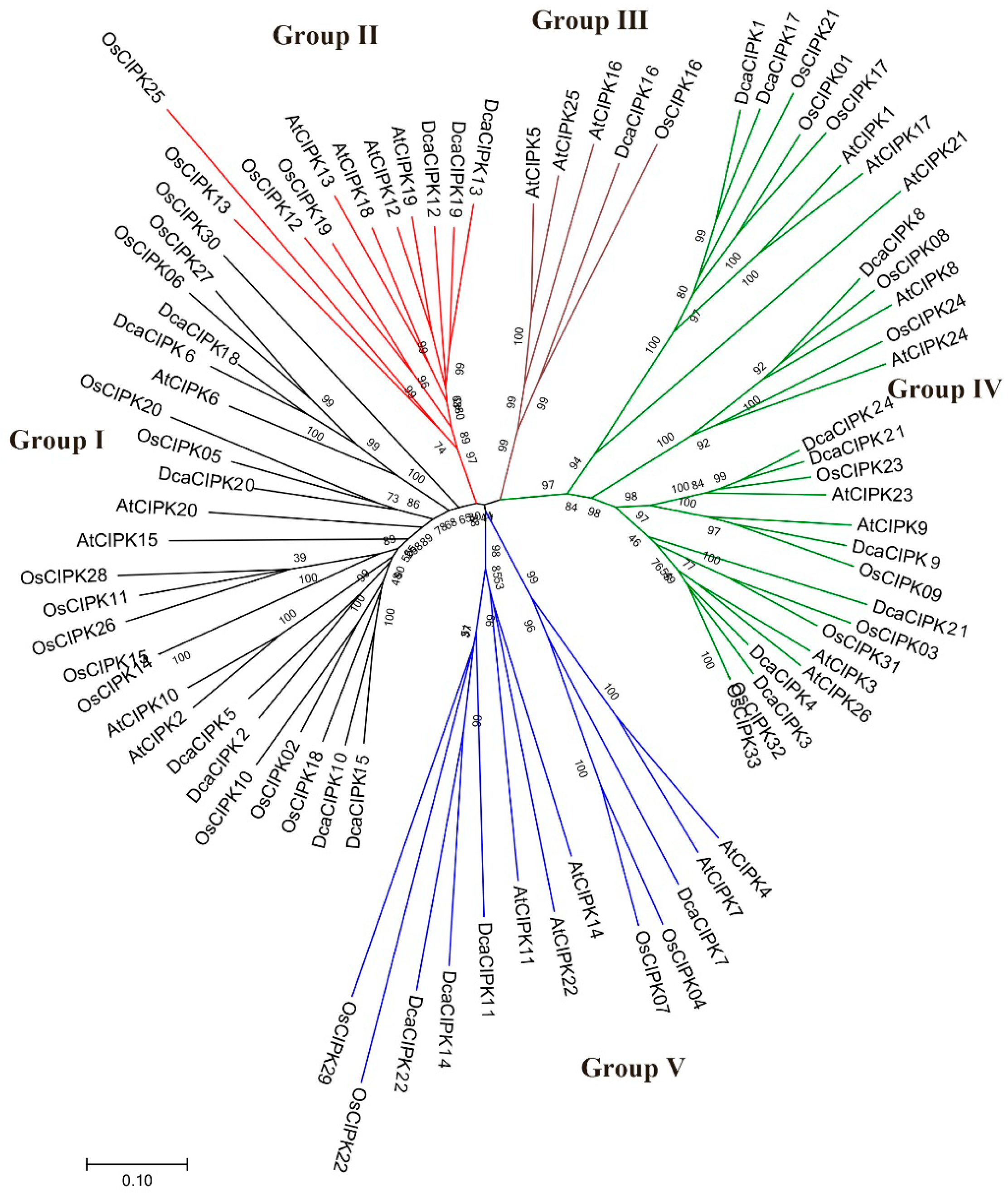
2.3. Phylogenetic Analysis of CIPK in Plants
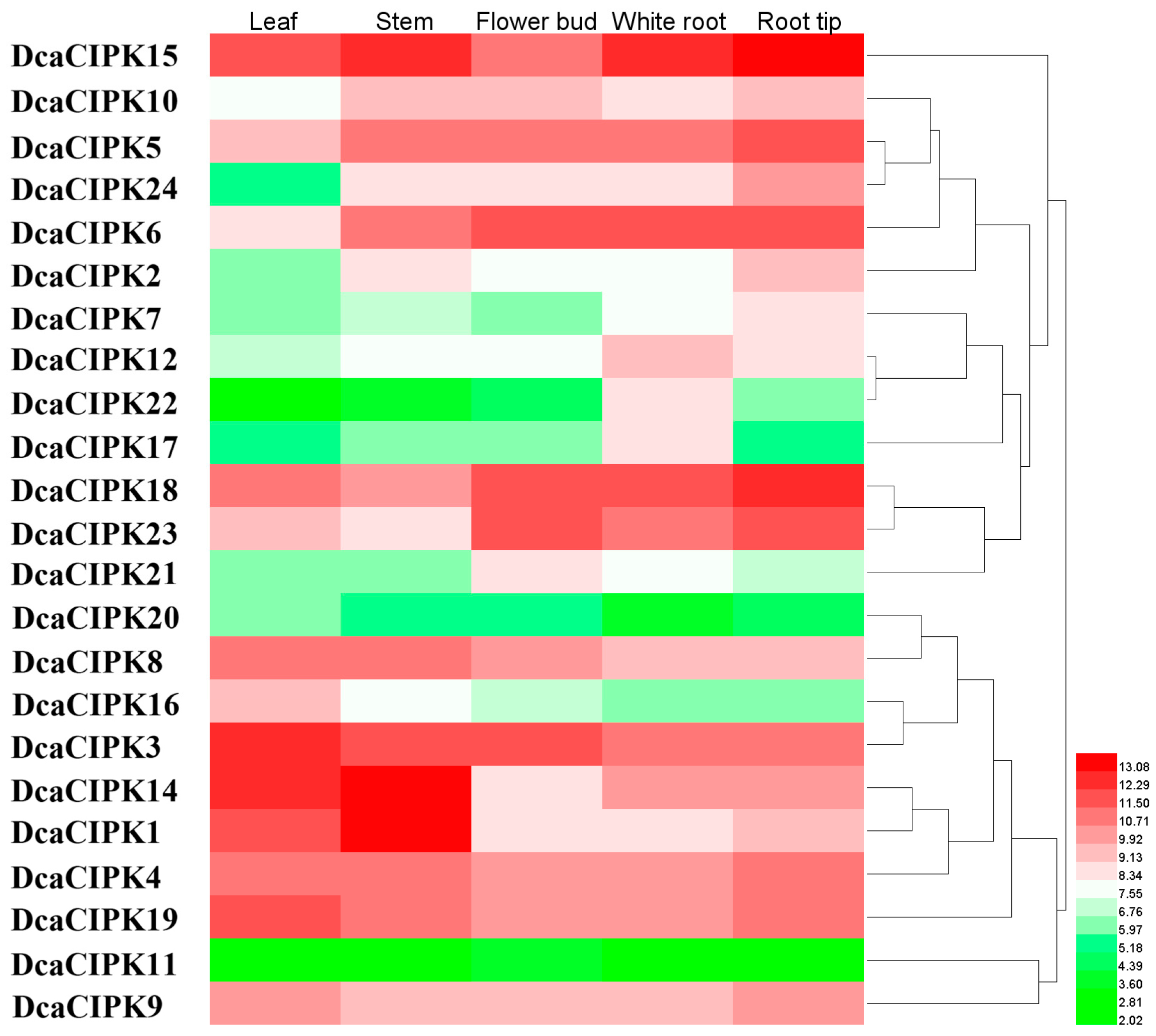
2.4. Expression of D. catenatum CIPK Genes in Different Plant Tissues
2.5. Expression of D. catenatum CIPK Genes under Abiotic Stress
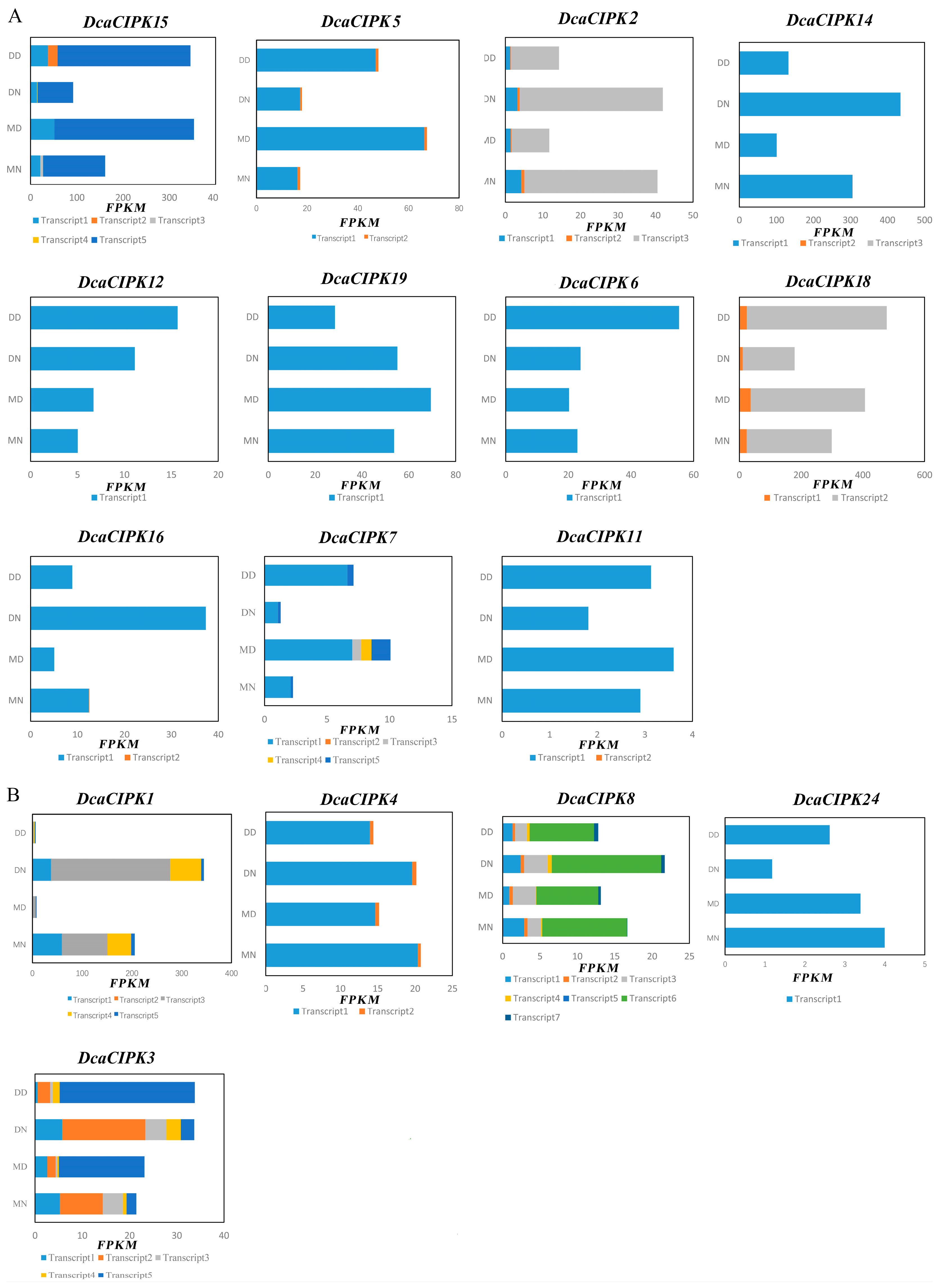
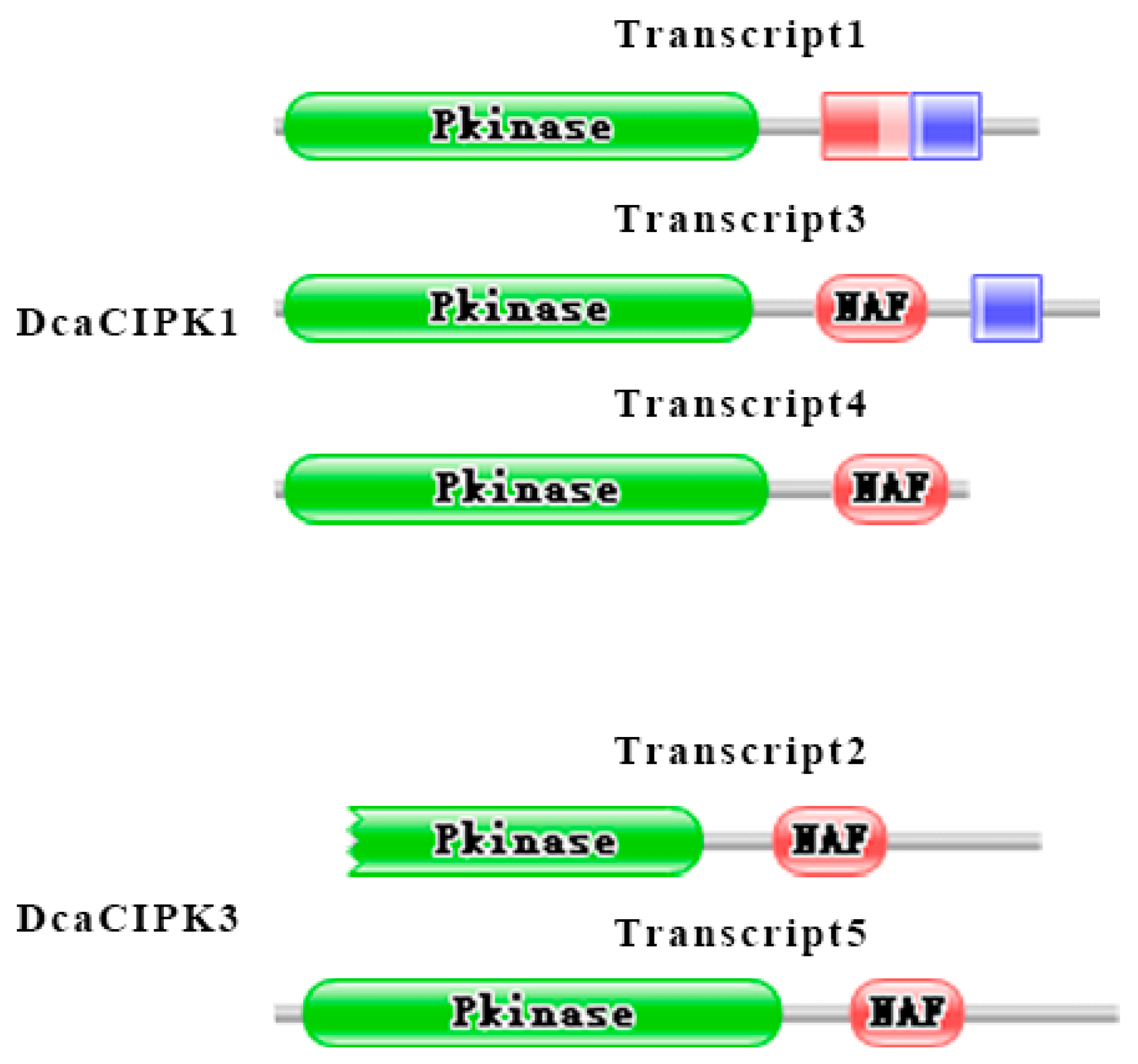
2.6. Alternative Splicing Analysis of CIPK Members under Drought Stress
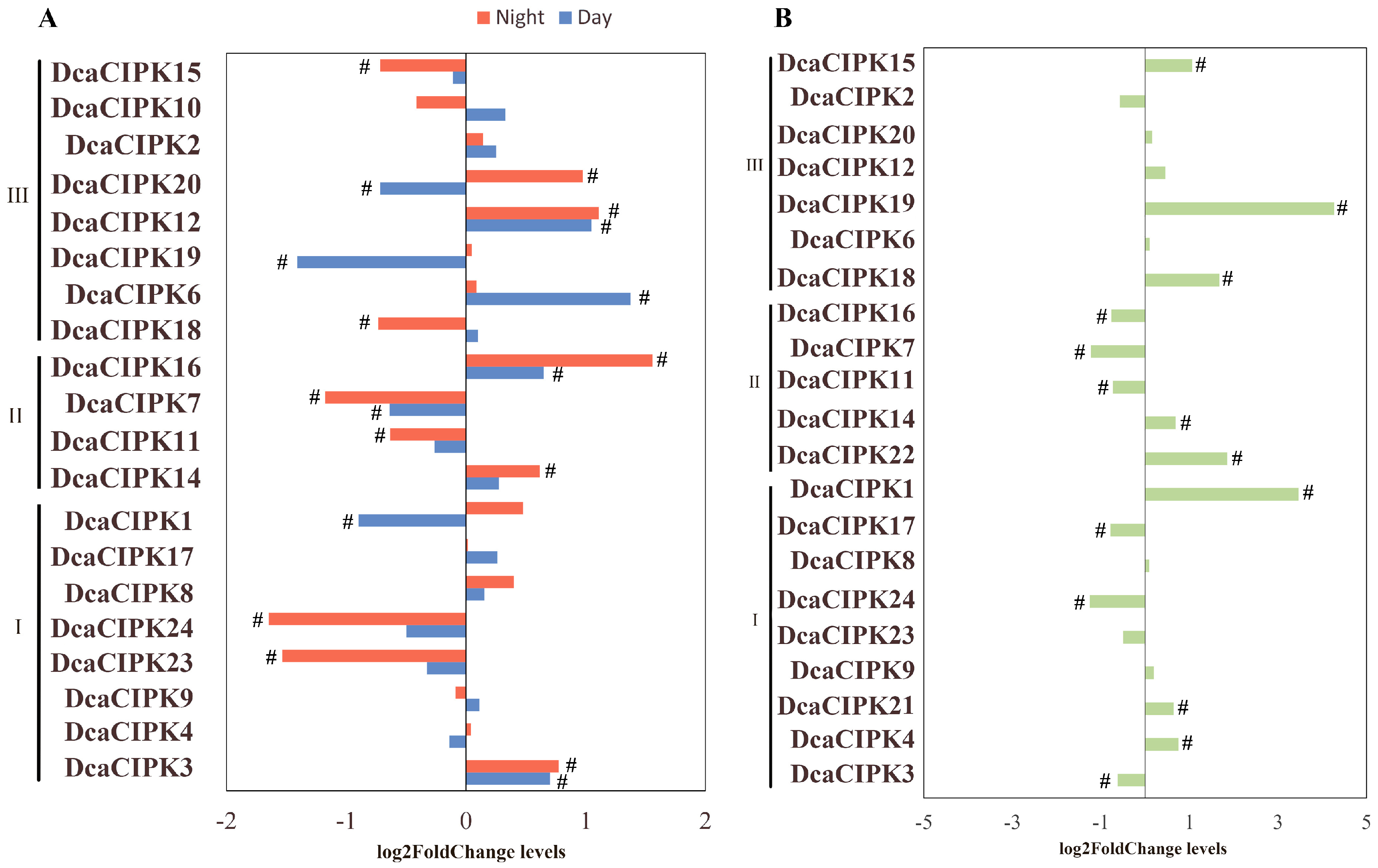
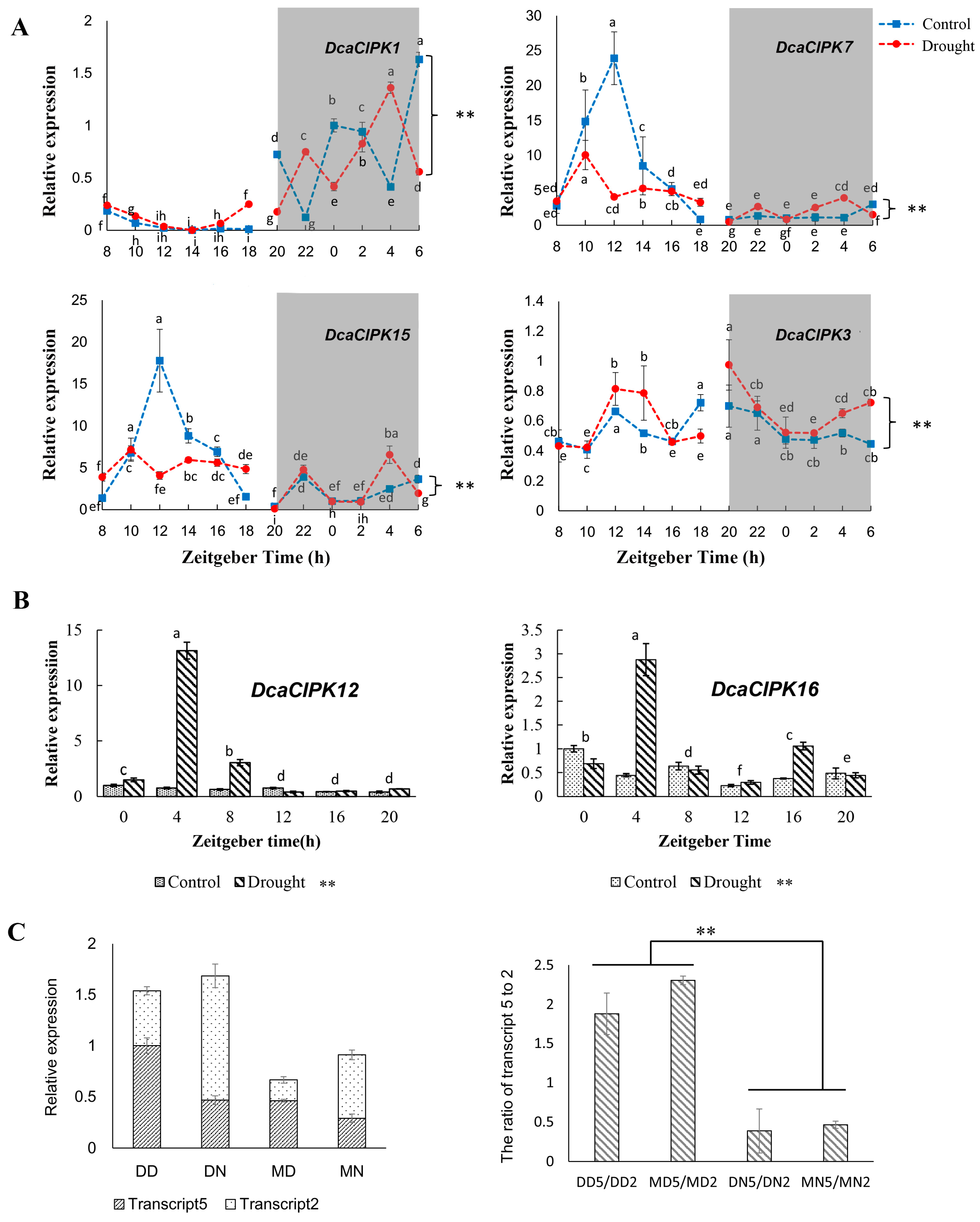
2.7. Effect of Time-of-Day and Drought Stress on Selected Genes mRNA Expression
3. Discussion
3.1. Phylogenetics and Structure of DcaCIPK Genes
3.2. DcaCIPK Gene Expression and Functions in Drought Stress
3.3. Circadian Rhythm and Drought Stress Both Influence Alternative Splicing of CIPK Members
4. Materials and Methods
4.1. Genome-Wide Identification of the CIPK Gene Family in D. catenatum
4.2. Multiple Protein Sequence Alignment and Phylogenetic Tree Construction
4.3. Exon–Intron Structure Analysis and Identification of Conserved Motifs
4.4. Transcriptome Analysis of D. catenatum CIPK Gene Expression
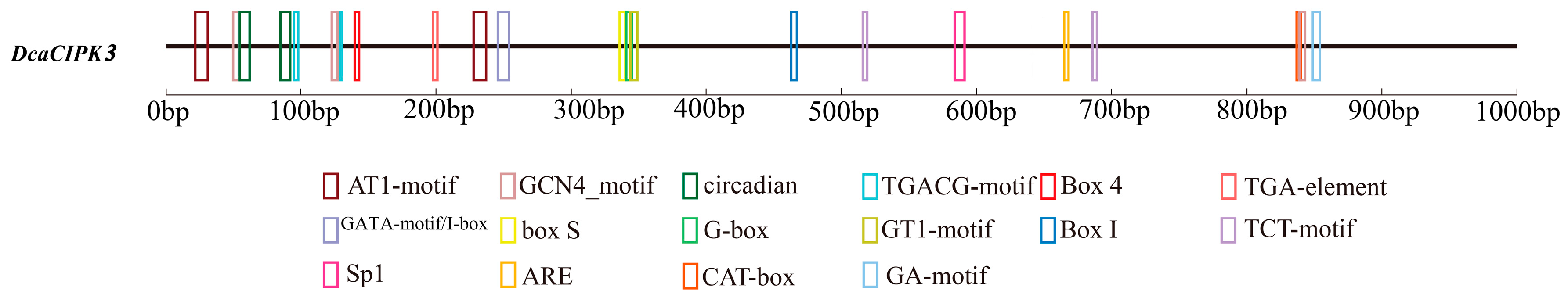
4.5. Alternative Splicing Analysis and Cis-Acting Regulatory Element Prediction
4.6. Plant Material, Growth Conditions, and Experimental Treatments
4.7. RNA Extraction
4.8. Quantitative Real-Time PCR
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Khanna-Chopra, R.; Singh, K. Drought resistance in crops: Physiological and genetic basis of traits for crop productivity. In Stress Responses in Plants; Tripathi, B.N., Müller, M., Eds.; Springer: New York, NY, USA, 2015; pp. 267–292. [Google Scholar]
- Zhang, G.Q.; Xu, Q.; Bian, C.; Tsai, W.C.; Yeh, C.M.; Liu, K.W.; Yoshida, K.; Zhang, L.S.; Chang, S.B.; Chen, F.; et al. The Dendrobium catenatum Lindl. genome sequence provides insights into polysaccharide synthase, floral development and adaptive evolution. Sci. Rep. 2016, 6, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Zotz, G.; Winkler, U. Aerial roots of epiphytic orchids: The velamen radicum and its role in water and nutrient uptake. Oecologia 2013, 171, 733–741. [Google Scholar] [CrossRef] [PubMed]
- Herrera, A. Crassulacean acid metabolism and fitness under water deficit stress: If not for carbon gain, what is facultative CAM good for? Ann. Bot. 2009, 103, 645–653. [Google Scholar] [CrossRef] [PubMed]
- Haider, M.S.; Barnes, J.D.; Cushman, J.C.; Borland, A.M. A CAM-and starch-deficient mutant of the facultative CAM species Mesembryanthemum crystallinum reconciles sink demands by repartitioning carbon during acclimation to salinity. J. Exp. Bot. 2012, 63, 1985–1996. [Google Scholar] [CrossRef] [PubMed]
- Winter, K.; Holtum, J.A.M. Facultative crassulacean acid metabolism (CAM) plants: Powerful tools for unravelling the functional elements of CAM photosynthesis. J. Exp. Bot. 2014, 65, 3425–3441. [Google Scholar] [CrossRef] [PubMed]
- Celenza, J.; Carlson, M. A yeast gene that is essential for release from glucose repression encodes a protein kinase. Science 1986, 233, 1175–1180. [Google Scholar] [CrossRef] [PubMed]
- Gancedo, J.M. Yeast carbon catabolite repression. Microbiol. Mol. Biol. Rev. 1998, 62, 334–361. [Google Scholar]
- Honigberg, S.M.; Lee, R.H. Snf1 kinase connects nutritional pathways controlling meiosis in Saccharomyces cerevisiae. Mol. Cell. Biol. 1998, 18, 4548–4555. [Google Scholar] [CrossRef]
- Zhu, J.-K. Abiotic stress signaling and responses in plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef]
- Hardie, D.G. Plant protein serine threonine kinases: Classification and functions. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1999, 50, 97–131. [Google Scholar] [CrossRef]
- Hrabak, E.M.; Chan, C.W.M.; Gribskov, M.; Harper, J.F.; Choi, J.H.; Halford, N.; Luan, S.; Nimmo, H.G.; Sussman, M.R.; Thomas, M.; et al. The Arabidopsis CDPK-SnRK superfamily of protein kinases. Plant Physiol. 2015, 132, 666–680. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Li, H.D.; Chen, L.Q.; Wang, Y.; Liu, L.L.; He, L.; Wu, W.H. A protein kinase, interacting with two calcineurin B-like proteins, regulates K+Transporter AKT1 in Arabidopsis. Cell 2006, 125, 1347–1360. [Google Scholar] [CrossRef] [PubMed]
- Drerup, M.M.; Schlücking, K.; Hashimoto, K.; Manishankar, P.; Steinhorst, L.; Kuchitsu, K.; Kudla, J. The calcineurin B-like calcium sensors CBL1 and CBL9 together with their interacting protein kinase CIPK26 regulate the Arabidopsis NADPH oxidase RBOHF. Mol. Plant 2013, 6, 559–569. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Yang, X.; Wang, L.; Zhu, L.; Zhou, T.; Deng, J.; Zhang, X. Molecular cloning and functional characterization of a novel cotton CBL-interacting protein kinase gene (GhCIPK6) reveals its involvement in multiple abiotic stress tolerance in transgenic plants. Biochem. Biophys. Res. Commun. 2013, 435, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Kong, Z.; Omo-Ikerodah, E.; Xu, W.; Li, Q.; Xue, Y. Calcineurin B-like interacting protein kinase OsCIPK23 functions in pollination and drought stress responses in rice (Oryza sativa L.). J. Genet. Genom. 2008, 35, 531–543. [Google Scholar] [CrossRef]
- Kolukisaoglu, U. Calcium sensors and their interacting protein kinases: Genomics of the Arabidopsis and rice CBL-CIPK signaling networks. Plant Physiol. 2004, 134, 43–58. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Xia, X.; Yin, W.; Zhang, H. Comparative genomic analysis of CIPK gene family in Arabidopsis and Populus. Plant Growth Regul. 2007, 52, 101–110. [Google Scholar] [CrossRef]
- Chen, X.; Gu, Z.; Xin, D.; Hao, L.; Liu, C.; Huang, J.; Ma, B.; Zhang, H. Identification and characterization of putative CIPK genes in maize. J. Genet. Genom. 2011, 38, 77–87. [Google Scholar] [CrossRef]
- Kanwar, P.; Sanyal, S.K.; Tokas, I.; Yadav, A.K.; Pandey, A.; Kapoor, S.; Pandey, G.K. Comprehensive structural, interaction and expression analysis of CBL and CIPK complement during abiotic stresses and development in rice. Cell Calcium 2014, 56, 81–95. [Google Scholar] [CrossRef]
- Zhu, K.; Chen, F.; Liu, J.; Chen, X.; Hewezi, T.; Cheng, Z.M.M. Evolution of an intron-poor cluster of the CIPK gene family and expression in response to drought stress in soybean. Sci. Rep. 2016, 6, 1–12. [Google Scholar] [CrossRef]
- Zhang, H.; Yang, B.; Liu, W.-Z.; Li, H.; Wang, L.; Wang, B.; Deng, M.; Liang, W.; Deyholos, M.K.; Jiang, Y.-Q. Identification and characterization of CBL and CIPK gene families in canola (Brassica napus L.). BMC Plant Biol. 2014, 14, 8. [Google Scholar] [CrossRef]
- Ye, C.-Y.; Xia, X.; Yin, W. Evolutionary analysis of CBL-interacting protein kinase gene family in plants. Plant Growth Regul. 2013, 71, 49–56. [Google Scholar] [CrossRef]
- He, C.; Yu, Z.; Teixeira Da Silva, J.A.; Zhang, J.; Liu, X.; Wang, X.; Zhang, X.; Zeng, S.; Wu, K.; Tan, J.; et al. DoGMP1 from Dendrobium officinale contributes to mannose content of water-soluble polysaccharides and plays a role in salt stress response. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef]
- Zhang, J.; He, C.; Wu, K.; Teixeira da Silva, J.A.; Zeng, S.; Zhang, X.; Yu, Z.; Xia, H.; Duan, J. Transcriptome analysis of Dendrobium officinale and its application to the identification of genes associated with polysaccharide synthesis. Front. Plant Sci. 2016, 7, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Kleist, T.J.; Spencley, A.L.; Luan, S. Comparative phylogenomics of the CBL-CIPK calcium-decoding network in the moss Physcomitrella, Arabidopsis, and other green lineages. Front. Plant Sci. 2014, 5, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.; Miao, Z.; Ren, C.; Yuan, R.; Tang, Y.; Zhang, X.; Han, Z.; Ma, C. Evolution of intron-poor clades and expression patterns of the glycosyltransferase family 47. Planta 2017, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.C.; Lan, W.-Z.; Kim, B.-G.; Li, L.; Cheong, Y.H.; Pandey, G.K.; Lu, G.; Buchanan, B.B.; Luan, S. A protein phosphorylation/dephosphorylation network regulates a plant potassium channel. Proc. Natl. Acad. Sci. USA 2007, 104, 15959–15964. [Google Scholar] [CrossRef]
- Kim, K.-N.; Cheong, Y.H.; Grant, J.J.; Pandey, G.K.; Luan, S. CIPK3, a calcium sensor–associated protein kinase that regulates abscisic acid and cold signal transduction in Arabidopsis. Plant Cell 2003, 15, 411–423. [Google Scholar] [CrossRef]
- Xiong, L. The Arabidopsis LOS5/ABA3 locus encodes a molybdenum cofactor sulfurase and modulates cold stress- and osmotic stress-responsive gene expression. Plant Cell Online 2001, 13, 2063–2083. [Google Scholar] [CrossRef]
- Wu, Z.-G.; Jiang, W.; Chen, S.-L.; Mantri, N.; Tao, Z.-M.; Jiang, C.-X. Insights from the cold transcriptome and metabolome of Dendrobium officinale: Global reprogramming of metabolic and gene regulation networks during cold acclimation. Front. Plant Sci. 2016, 7, 1–16. [Google Scholar] [CrossRef]
- Keunen, E.; Peshev, D.; Vangronsveld, J.; Van Den Ende, W.; Cuypers, A. Plant sugars are crucial players in the oxidative challenge during abiotic stress: Extending the traditional concept. Plant Cell Environ. 2013, 36, 1242–1255. [Google Scholar] [CrossRef] [PubMed]
- Nishizawa, A.; Yabuta, Y.; Shigeoka, S. Galactinol and raffinose constitute a novel function to protect plants from oxidative damage. Plant Physiol. 2008, 147, 1251–1263. [Google Scholar] [CrossRef] [PubMed]
- Farani, T.F.; Gentile, A.; Tavares, R.G.; Ribeiro, C.; Menossi, M. Characterization of a protein-protein interaction network of the CBL-interacting protein kinase 8 from sugarcane. Genet. Mol. Res. 2015, 14, 483–491. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, C.; Felix, J.M.; Menossi, M. Sugar signaling on the regulation of the CBL/CIPK network in sugarcane contrasting sugar amount varieties. In Proceedings of the 56° Brazilian Genetics Conference, Guarujá, Brazil, 2010. [Google Scholar]
- Lee, E.-J.; Iai, H.; Sano, H.; Koizumi, N. Sugar responsible and tissue specific expression of a gene encoding AtCIPK14, an Arabidopsis CBL-interacting protein kinase. Biosci. Biotechnol. Biochem. 2005, 69, 242–245. [Google Scholar] [CrossRef]
- Sanyal, S.K.; Kanwar, P.; Samtani, H.; Kaur, K.; Jha, S.K.; Pandey, G.K. Alternative splicing of CIPK3 results in distinct target selection to propagate ABA signaling in Arabidopsis. Front. Plant Sci. 2017, 8, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Kelemen, O.; Convertini, P.; Zhang, Z.; Wen, Y.; Shen, M.; Falaleeva, M.; Stamm, S. Function of alternative splicing. Gene 2013, 514, 1–30. [Google Scholar] [CrossRef]
- Finn, R.D.; Bateman, A.; Clements, J.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Heger, A.; Hetherington, K.; Holm, L.; Mistry, J.; et al. Pfam: The protein families database. Nucleic Acids Res. 2014, 42, 222–230. [Google Scholar] [CrossRef] [PubMed]
- Eddy, S.R. Profile hidden markov models. Bioinformatics 1999, 14, 755–763. [Google Scholar] [CrossRef]
- Letunic, I. SMART 4.0: Towards genomic data integration. Nucleic Acids Res. 2004, 32, 142–144. [Google Scholar] [CrossRef]
- Gasteiger, E.; Hoogland, C.; Gattiker, A.; Duvaud, S.; Wilkins, M.R.; Appel, R.D.; Bairoch, A. Protein identification and analysis tools on the ExPASy server. Methods Mol. Biol. 1999, 52, 571–608. [Google Scholar]
- Bjellqvist, B.; Hughes, G.J.; Pasquali, C.; Paquet, N.; Ravier, F.; Sanchez, J.; Frutiger, S.; Hochstrasser, D.F. The focusing positions of polypeptides in immobilized pH gradients can be predicted from their amino acid sequences. Electrophoresis 1993, 14, 1023–1031. [Google Scholar] [CrossRef] [PubMed]
- Celis, J.E. Reference points for comparisons of two-dimensional maps of proteins from different human cell types defined in a pH scale where isoelectric points correlate with polypeptide compositions. Electrophoresis 1994, 15, 529–539. [Google Scholar] [CrossRef]
- Gasteiger, E.; Gattiker, A.; Hoogland, C.; Ivanyi, I.; Appel, R.D.; Bairoch, A. ExPASy: The proteomics server for in-depth protein knowledge and analysis. Nucleic Acids Res. 2003, 31, 3784–3788. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: A multiple sequence alignment method with reduced time and space complexity. BMC Bioinform. 2004, 5, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Saitou, N.; Nei, M. The neighbour-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Jin, J.; Guo, A.Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef] [PubMed]
- Bailey, T.L.; Johnson, J.; Grant, C.E.; Noble, W.S. The MEME Suite. Nucleic Acids Res. 2015, 43, W39–W49. [Google Scholar] [CrossRef] [PubMed]
- Wan, X.; Zou, L.; Zheng, B.; Tian, Y.; Wang, Y. Transcriptomic profiling for prolonged drought in Dendrobium catenatum. Sci. Data 2018, 5, 180233. [Google Scholar] [CrossRef]
- Zhang, G.Q.; Liu, K.W.; Li, Z.; Lohaus, R.; Hsiao, Y.Y.; Niu, S.C.; Wang, J.Y.; Lin, Y.C.; Xu, Q.; Chen, L.J.; et al. The Apostasia genome and the evolution of orchids. Nature 2017, 549, 379–383. [Google Scholar] [CrossRef]
- Zhang, M.; Sun, H.; Fei, Z.; Zhan, F.; Gong, X.; Gao, S. Fastq_clean: An optimized pipeline to clean the Illumina sequencing data with quality control. 2014 IEEE Int. Conf. Bioinform. Biomed. 2014, 44–48. [Google Scholar] [CrossRef]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed]
- Pertea, M.; Kim, D.; Pertea, G.M.; Leek, J.T.; Salzberg, S.L. Transcript-level expression analysis of RNA-seq experiments with HISAT, StringTie and Ballgown. Nat. Protoc. 2016, 11, 1650–1667. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R.; 1000 genome project data processing subgroup. The sequence alignment/map (SAM) format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [PubMed]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq-A Python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef] [PubMed]
- Anders, S.; McCarthy, D.J.; Chen, Y.S.; Okoniewski, M.; Smyth, G.K.; Huber, W.; Robinson, M.D. Count-based differential expression analysis of RNA sequencing data using R and Bioconductor. Nat. Protoc. 2013, 8, 1765–1786. [Google Scholar] [CrossRef] [PubMed]
- Deng, W.; Wang, Y.; Liu, Z.; Cheng, H.; Xue, Y.; Min, M. HemI: A Toolkit for Illustrating Heatmaps. PLoS ONE 2014, 9, 9–13. [Google Scholar] [CrossRef]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.; Mendell, J.T.; Salzberg, S.L. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef]
- R Foundation for Statistical Computing. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2015. [Google Scholar]
- Frazee, A.C.; Pertea, G.; Jaffe, A.E.; Langmead, B.; Salzberg, S.L.; Leek, J.T. Ballgown bridges the gap between transcriptome assembly and expression analysis. Nat. Biotechnol. 2015, 33, 243–246. [Google Scholar] [CrossRef]
- Lescot, M. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Marcoulides, G.A.; Marcoulides, L.D. SAS (Statistical Analysis System); California State University: Fullerton, CA, USA, 2003. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Gene ID | Accession ID in NCBI | Locus | Gene Length (bp) | Amino Acid Length (aa) | PI | Mw (KDa) | Exons | CDS Length (bp) |
|---|---|---|---|---|---|---|---|---|---|
| DcaCIPK1 | Dca001684 | XP_020700642.1 | Dcat_scaffold_83:1213297-1226000 | 12,704 | 452 | 7.94 | 51.17 | 13 | 1359 |
| DcaCIPK2 | Dca021183 | XP_020683888.1 | Dcat_scaffold_6544:136242-137564 | 1323 | 440 | 9.16 | 50.23 | 1 | 1323 |
| DcaCIPK3 | Dca019377 | XP_020701813.1 | Dcat_scaffold_8095:113831-123888 | 10,058 | 451 | 8.7 | 51.47 | 14 | 1356 |
| DcaCIPK4 | Dca004523 | XP_020703034.1 | Dcat_scaffold_4350:650622-668668 | 18,047 | 434 | 8.32 | 49.36 | 14 | 1305 |
| DcaCIPK5 | Novl001335 | XP_020700988.1 | Dcat_scaffold_787:157699-159003 | 1305 | 432 | 9.18 | 49.55 | 1 | 1299 |
| DcaCIPK6 | Dca007455 | XP_020673946.1 | Dcat_scaffold_3166:141500-142810 | 1311 | 436 | 9.1 | 48.19 | 1 | 1311 |
| DcaCIPK7 | Dca005524 | XP_020676722.1 | Dcat_scaffold_1341:459825-461114 | 1290 | 429 | 9.32 | 48.2 | 1 | 1290 |
| DcaCIPK8 | Dca002402 | XP_020676379.1 | Dcat_scaffold_2414:350624-380208 | 29,585 | 449 | 8.01 | 51.09 | 14 | 1350 |
| DcaCIPK9 | Dca000041 | XP_020685625.1 | Dcat_scaffold_358:1253684-1269205 | 15,522 | 502 | 9.24 | 57.14 | 14 | 1509 |
| DcaCIPK10 | Dca024760 | XP_020681762.1 | Dcat_scaffold_45917:29214-30560 | 1347 | 448 | 9.07 | 50.97 | 1 | 1347 |
| DcaCIPK11 | Dca000484 | XP_020701016.1 | Dcat_scaffold_787:369843-371213 | 1371 | 443 | 8.71 | 49.58 | 3 | 1332 |
| DcaCIPK12 | Dca017399 | XP_020676379.1 | Dcat_scaffold_2131:315817-317232 | 1416 | 471 | 8.79 | 53.46 | 1 | 1416 |
| DcaCIPK13 | Novl001612 | XP_020695665.1 | Dcat_scaffold_937523:77342-78802 | 1847 | 486 | 8.01 | 54.74 | 1 | 1461 |
| DcaCIPK14 | Dca010961 | XP_020686786.1 | Dcat_scaffold_19096:431277-432626 | 1350 | 449 | 8.28 | 50.5 | 1 | 1350 |
| DcaCIPK15 | Dca024725 | XP_020705240.1 | Dcat_scaffold_18826:50805-52154 | 1350 | 449 | 9.25 | 50.96 | 1 | 1350 |
| DcaCIPK16 | Dca007453 | XP_020673947.1 | Dcat_scaffold_3166:100617-101960 | 1344 | 447 | 8.59 | 49.78 | 1 | 1344 |
| DcaCIPK17 | Dca018676 | XP_020695665.1 | Dcat_scaffold_937621:2927-7179 | 4253 | 417 | 7.57 | 47.27 | 12 | 1250 |
| DcaCIPK18 | Dca003241 | XP_020675333.1 | Dcat_scaffold_9707:650080-651360 | 1281 | 426 | 9.04 | 47.15 | 1 | 1281 |
| DcaCIPK19 | Dca021168 | XP_020702232.1 | Dcat_scaffold_1902:144820-146241 | 1422 | 473 | 7.99 | 53.5 | 1 | 1422 |
| DcaCIPK20 | Dca003305 | XP_020702013.1 | Dcat_scaffold_3734:1066698-1076027 | 9330 | 446 | 9.37 | 49.92 | 2 | 1341 |
| DcaCIPK21 | Dca016061 | XP_020688235.1 | Dcat_scaffold_114:141330-157531 | 16,202 | 401 | 8.78 | 45.15 | 13 | 1206 |
| DcaCIPK22 | Dca021931 | XP_020680456.1 | Dcat_scaffold_8676:215192-216610 | 1419 | 472 | 9.3 | 52.82 | 1 | 1419 |
| DcaCIPK23 | Dca012740 | XP_020697021.1 | Dcat_scaffold_2959:424009-432987 | 8979 | 432 | 8.94 | 48.82 | 14 | 1299 |
| DcaCIPK24 | Dca025565 | XP_020688792.1 | Dcat_scaffold_11189:59465-64362 | 4898 | 442 | 8.78 | 49.93 | 14 | 1329 |
| Dataset | Tissue | Treatment | Collected Time | BioSample Accessions in NCBI | Sources |
|---|---|---|---|---|---|
| Dataset A | Leaf | Drought stress | Day (9:00 AM) | SAMN08512102–SAMN08512105 | Wan et al. [51] |
| Night (9:00 PM) | SAMN08512110–SAMN08512113 | ||||
| Control | Day | SAMN08512106–SAMN08512109 | |||
| Night | SAMN08512114–SAMN08512117 | ||||
| Dataset B | Leaf | Cold stress | Day | SAMN04534730–SAMN04534732 | Wu et al. [31] |
| Control | Day | SAMN04534727–SAMN04534729 | |||
| Dataset C | Flower buds | n.a. | Day | SAMN05908201 | Zhang et al. [52] |
| Leaf | n.a. | Day | SAMN05912851 | ||
| Green root tip | n.a. | Day | SAMN05908239 | ||
| White root | n.a. | Day | SAMN05908241 | ||
| Stem | n.a. | Day | SAMN05908200 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wan, X.; Zou, L.-H.; Zheng, B.-Q.; Wang, Y. Circadian Regulation of Alternative Splicing of Drought-Associated CIPK Genes in Dendrobium catenatum (Orchidaceae). Int. J. Mol. Sci. 2019, 20, 688. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20030688
Wan X, Zou L-H, Zheng B-Q, Wang Y. Circadian Regulation of Alternative Splicing of Drought-Associated CIPK Genes in Dendrobium catenatum (Orchidaceae). International Journal of Molecular Sciences. 2019; 20(3):688. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20030688
Chicago/Turabian StyleWan, Xiao, Long-Hai Zou, Bao-Qiang Zheng, and Yan Wang. 2019. "Circadian Regulation of Alternative Splicing of Drought-Associated CIPK Genes in Dendrobium catenatum (Orchidaceae)" International Journal of Molecular Sciences 20, no. 3: 688. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20030688