IL-10 Could Play a Role in the Interrelation between Diabetes Mellitus and Osteoarthritis


 , and
, and
Abstract
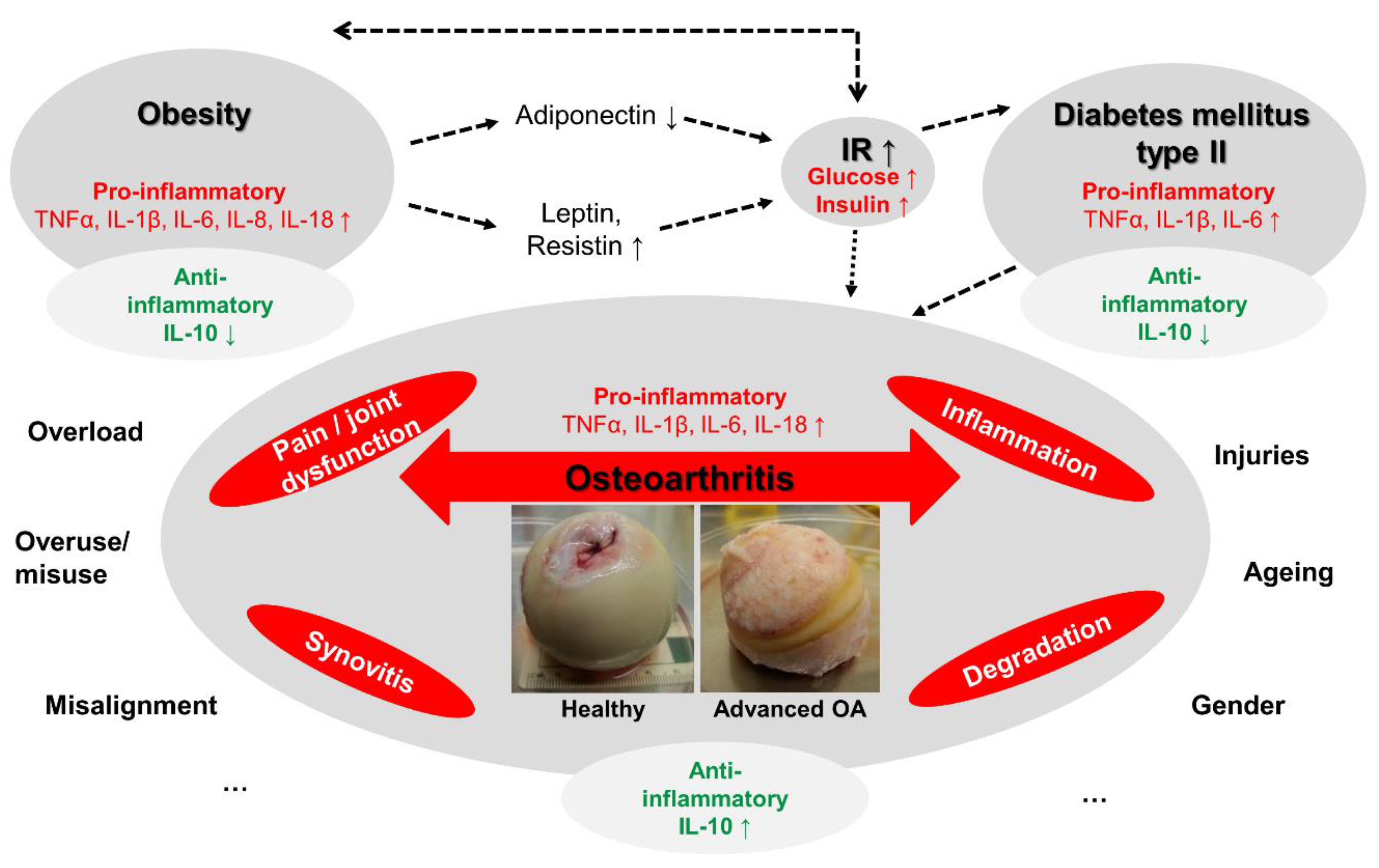
:1. Introduction
2. Results
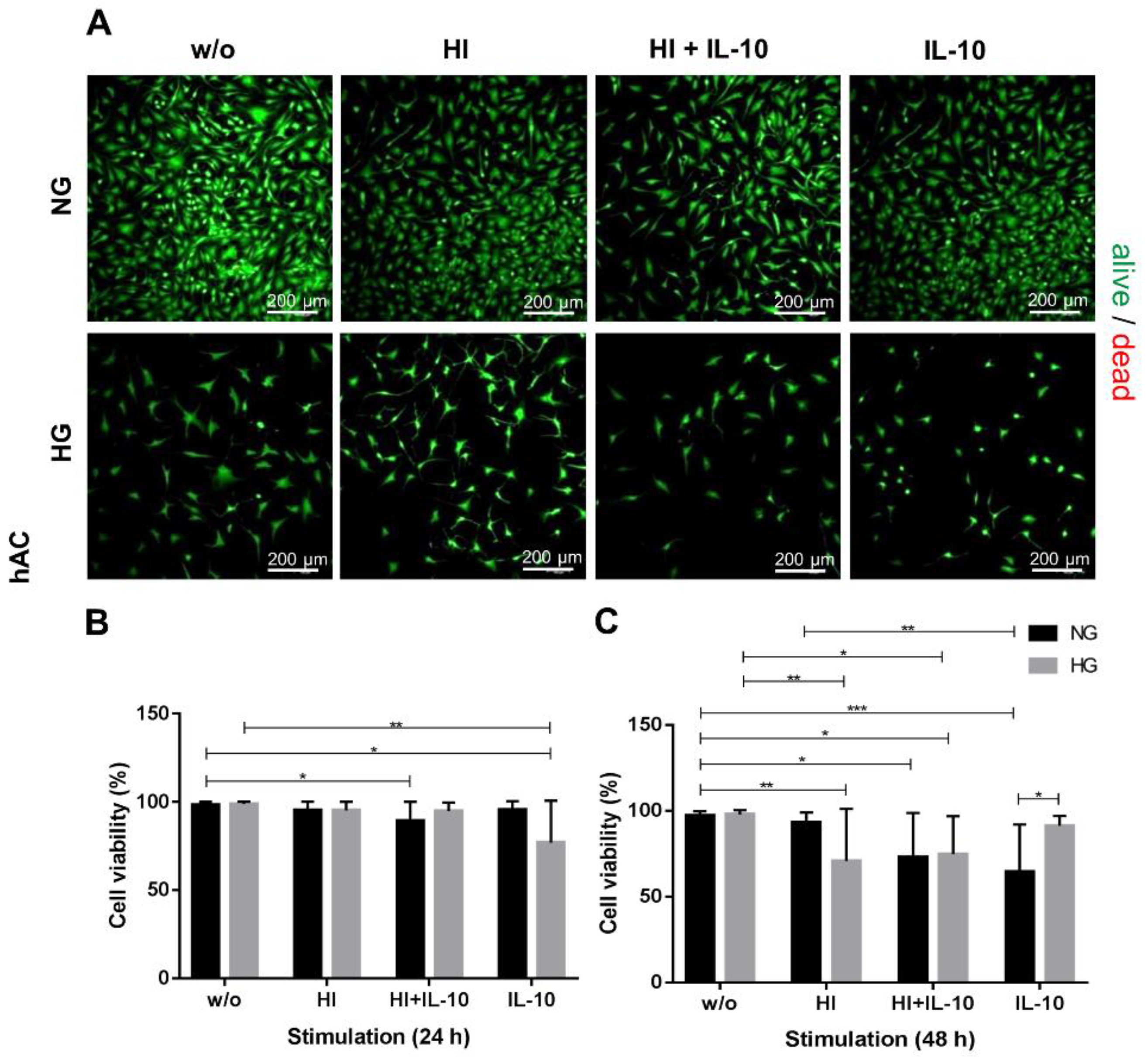
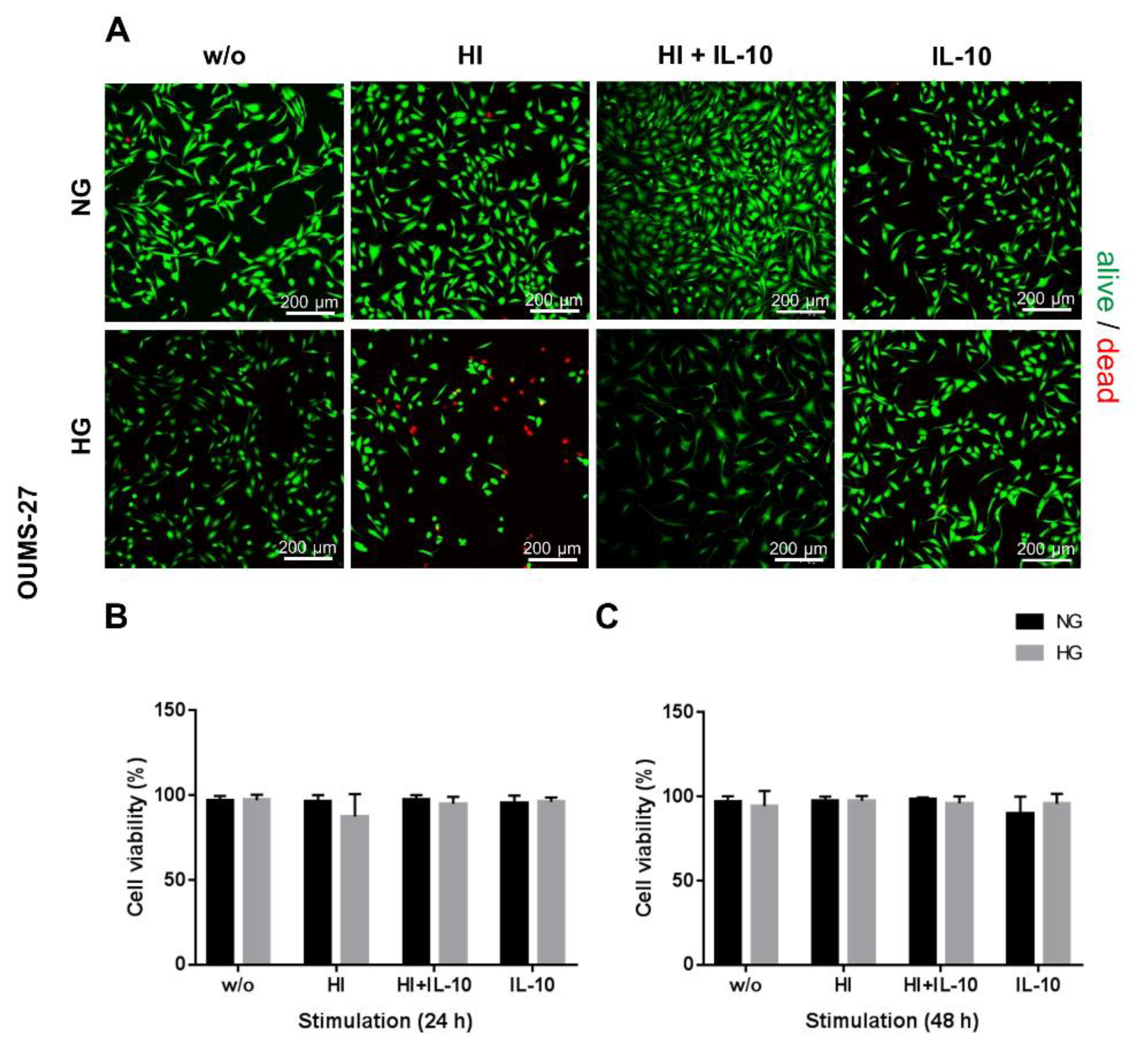
2.1. HG and HI Influence Cell Viability and Morphology
2.2. Glucose Content and Insulin Stimulation Impairs Metabolic Activity and Cell Proliferation
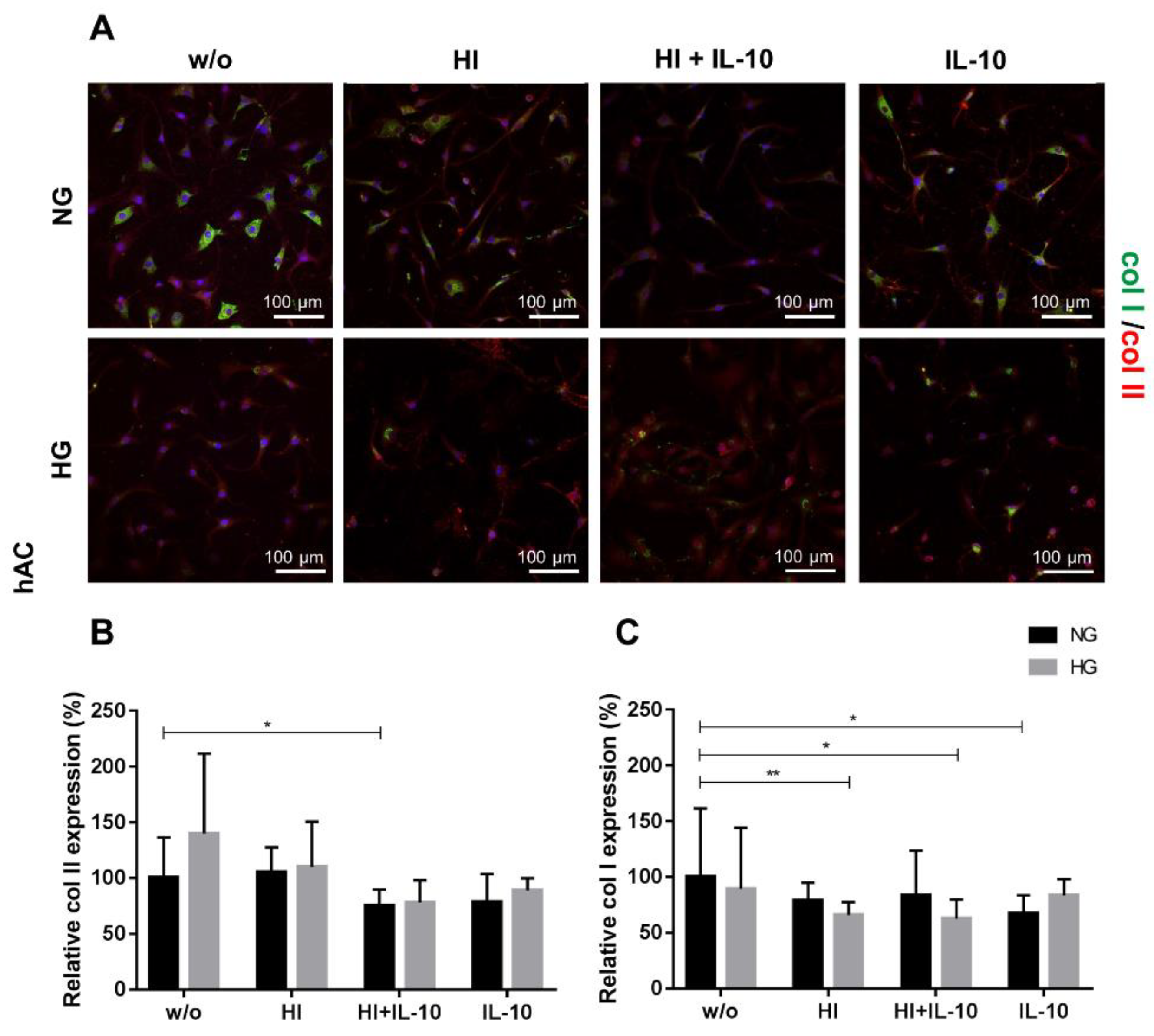
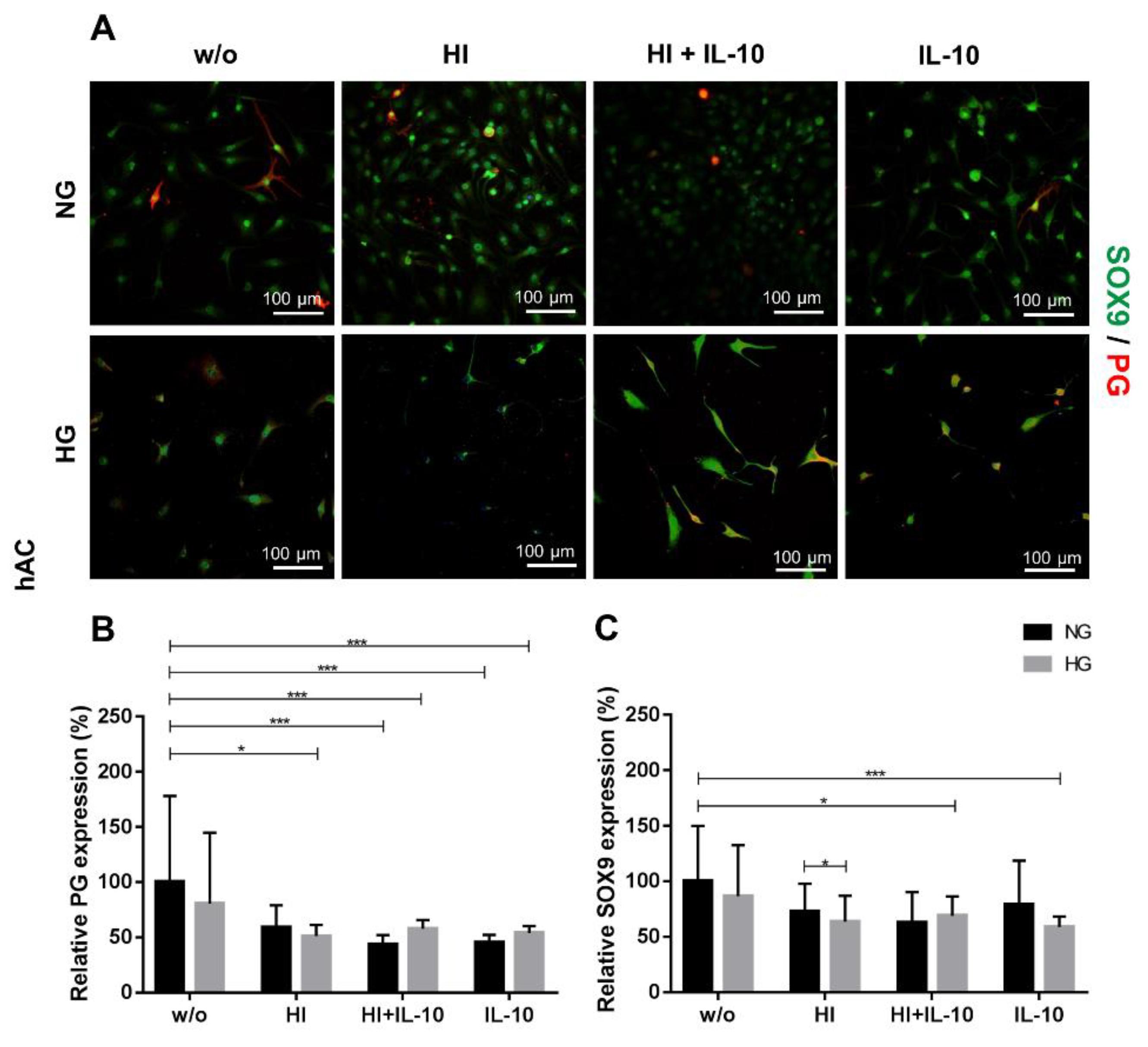
2.3. Impaired Cartilage ECM Production under Hyperglycemic and Hyperinsulinemic Conditions
3. Discussion
4. Materials and Methods
4.1. Isolation of Human Primary Articular Chondrocytes (hAC)
4.2. Expansion Culture of hAC and OUMS-27
4.3. Stimulation Culture
4.4. Cell Survival
4.5. Quantitative Assays for DNA Quantification and Assessment of Cell Metabolism
4.5.1. The CellTiter-Blue® Cell Viability Assay
4.5.2. DNA Quantification of Adherent Cells
4.6. Immunocytochemical Detection of Cartilage-Specific Markers
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| 2D | two-dimensional |
| 3D | three-dimensional |
| BMI | body mass index |
| col I/II | collagen type I/II |
| DM | diabetes mellitus |
| FBS | foetal bovine serum |
| FDA | fluorescein diacetate |
| hAC | human articular chondrocytes |
| HBSS | Hank’s balanced salt solution |
| HG | hyperglycemia |
| HI | hyperinsulinemia |
| IL | interleukin |
| MMPs | matrix metalloproteinases |
| NG | normoglycemia |
| OA | osteoarthritis |
| PBS | phosphate buffered saline |
| PFA | paraformaldehyde solution |
| PG | proteoglycan |
| PGE2 | Prostaglandin E2 |
| PI | propidium iodide |
| ROS | reactive oxygen species |
| RT | room temperature |
| SD | standard deviation |
| STAT | Signal Transducer and Activator of Transcription |
| T2DM | type 2 diabetes mellitus |
| TE | TRIS EDTA buffer |
| TBS | TRIS buffered saline |
| TBST | TRIS-buffered saline with Triton X 100 |
References
- Blom, A.B.; van Lent, P.L.; Libregts, S.; Holthuysen, A.E.; van der Kraan, P.M.; van Rooijen, N.; van den Berg, W.B. Crucial role of macrophages in matrix metalloproteinase-mediated cartilage destruction during experimental osteoarthritis: Involvement of matrix metalloproteinase 3. Arthritis Rheum. 2007, 56, 147–157. [Google Scholar] [CrossRef] [PubMed]
- Zengini, E.; Hatzikotoulas, K.; Tachmazidou, I.; Steinberg, J.; Hartwig, F.P.; Southam, L.; Hackinger, S.; Boer, C.G.; Styrkarsdottir, U.; Gilly, A.; et al. Genome-wide analyses using UK Biobank data provide insights into the genetic architecture of osteoarthritis. Nat. Genet. 2018, 50, 549–558. [Google Scholar] [CrossRef] [PubMed]
- Courties, A.; Sellam, J. Osteoarthritis and type 2 diabetes mellitus: What are the links? Diabetes Res. Clin. Pract. 2016, 122, 198–206. [Google Scholar] [CrossRef] [PubMed]
- Livshits, G.; Zhai, G.; Hart, D.J.; Kato, B.S.; Wang, H.; Williams, F.M.; Spector, T.D. Interleukin-6 is a significant predictor of radiographic knee osteoarthritis: The Chingford Study. Arthritis Rheum. 2009, 60, 2037–2045. [Google Scholar] [CrossRef] [PubMed]
- Pearson, M.J.; Herndler-Brandstetter, D.; Tariq, M.A.; Nicholson, T.A.; Philp, A.M.; Smith, H.L.; Davis, E.T.; Jones, S.W.; Lord, J.M. IL-6 secretion in osteoarthritis patients is mediated by chondrocyte-synovial fibroblast cross-talk and is enhanced by obesity. Sci. Rep. 2017, 7, 3451. [Google Scholar] [CrossRef] [PubMed]
- Poole, A.R. Osteoarthritis as a whole joint disease. HSS J. 2012, 8, 4–6. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Benjamin Ma, C.; Link, T.M.; Castillo, D.D.; Blumenkrantz, G.; Lozano, J.; Carballido-Gamio, J.; Ries, M.; Majumdar, S. In vivo T(1rho) and T(2) mapping of articular cartilage in osteoarthritis of the knee using 3 T MRI. Osteoarthr. Cartil. 2007, 15, 789–797. [Google Scholar] [CrossRef]
- Mabey, T.; Honsawek, S. Cytokines as biochemical markers for knee osteoarthritis. World J. Orthop. 2015, 6, 95–105. [Google Scholar] [CrossRef]
- Goldring, M.B. Chondrogenesis, chondrocyte differentiation, and articular cartilage metabolism in health and osteoarthritis. Ther. Adv. Musculoskelet. Dis. 2012, 4, 269–285. [Google Scholar] [CrossRef] [Green Version]
- Clockaerts, S.; Bastiaansen-Jenniskens, Y.M.; Feijt, C.; De Clerck, L.; Verhaar, J.A.; Zuurmond, A.M.; Stojanovic-Susulic, V.; Somville, J.; Kloppenburg, M.; van Osch, G.J. Cytokine production by infrapatellar fat pad can be stimulated by interleukin 1beta and inhibited by peroxisome proliferator activated receptor alpha agonist. Ann. Rheum. Dis. 2012, 71, 1012–1018. [Google Scholar] [CrossRef]
- Loeser, R.F.; Goldring, S.R.; Scanzello, C.R.; Goldring, M.B. Osteoarthritis: A disease of the joint as an organ. Arthritis Rheum. 2012, 64, 1697–1707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Botha-Scheepers, S.; Watt, I.; Slagboom, E.; de Craen, A.J.; Meulenbelt, I.; Rosendaal, F.R.; Breedveld, F.C.; Huizinga, T.W.; Kloppenburg, M. Innate production of tumour necrosis factor alpha and interleukin 10 is associated with radiological progression of knee osteoarthritis. Ann. Rheum. Dis. 2008, 67, 1165–1169. [Google Scholar] [CrossRef] [PubMed]
- Carballo, C.B.; Coelho, T.R.P.; de Holanda Afonso, R.C.; Faria, J.C.O.; Alves, T.; Monte, S.M.; Ventura Matioszek, G.M.; Moura-Neto, V.; Brito, J.M. Osteoarthritic Synovial Fluid and TGF-beta1 Induce Interleukin-18 in Articular Chondrocytes. Cartilage 2018. [Google Scholar] [CrossRef]
- Wojdasiewicz, P.; Poniatowski, L.A.; Szukiewicz, D. The role of inflammatory and anti-inflammatory cytokines in the pathogenesis of osteoarthritis. Mediat. Inflamm. 2014, 2014, 561459. [Google Scholar] [CrossRef] [PubMed]
- Dan, H.; Liu, W.; Wang, J.; Wang, Z.; Wu, R.; Chen, Q.; Zeng, X.; Zhou, Y. Elevated IL-10 concentrations in serum and saliva from patients with oral lichen planus. Quintessence Int. 2011, 42, 157–163. [Google Scholar] [PubMed]
- Adams, S.B., Jr.; Nettles, D.L.; Jones, L.C.; Miller, S.D.; Guyton, G.P.; Schon, L.C. Inflammatory cytokines and cellular metabolites as synovial fluid biomarkers of posttraumatic ankle arthritis. Foot Ankle Int. 2014, 35, 1241–1249. [Google Scholar] [CrossRef]
- Iannone, F.; Lapadula, G. Obesity and inflammation—Targets for OA therapy. Curr. Drug Targets. 2010, 11, 586–598. [Google Scholar] [CrossRef]
- Papalia, R.; Vadala, G.; Torre, G.; Perna, M.; Saccone, L.; Cannata, F.; Denaro, V. The cytokinome in osteoarthritis, a new paradigm in diagnosis and prognosis of cartilage disease. J. Biol. Regul. Homeost. Agents 2016, 30, 77–83. [Google Scholar]
- Bigoni, M.; Turati, M.; Gandolla, M.; Sacerdote, P.; Piatti, M.; Castelnuovo, A.; Franchi, S.; Gorla, M.; Munegato, D.; Gaddi, D.; et al. Effects of ACL Reconstructive Surgery on Temporal Variations of Cytokine Levels in Synovial Fluid. Mediat. Inflamm. 2016, 2016, 8243601. [Google Scholar] [CrossRef]
- Suyasa, I.K.; Kawiyana, I.K.; Bakta, I.M.; Widiana, I.G. Interleukin-6 and ratio of plasma interleukin-6/interleukin-10 as risk factors of symptomatic lumbar osteoarthritis. World J. Orthop. 2017, 8, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Schulze-Tanzil, G.; Zreiqat, H.; Sabat, R.; Kohl, B.; Halder, A.; Muller, R.D.; John, T. Interleukin-10 and articular cartilage: Experimental therapeutical approaches in cartilage disorders. Curr. Gene Ther. 2009, 9, 306–315. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, S.; Mrosewski, I.; Silawal, S.; Schulze-Tanzil, G. The interrelation of osteoarthritis and diabetes mellitus: Considering the potential role of interleukin-10 and in vitro models for further analysis. Inflamm. Res. 2018, 67, 285–300. [Google Scholar] [CrossRef] [PubMed]
- Behrendt, P.; Preusse-Prange, A.; Kluter, T.; Haake, M.; Rolauffs, B.; Grodzinsky, A.J.; Lippross, S.; Kurz, B. IL-10 reduces apoptosis and extracellular matrix degradation after injurious compression of mature articular cartilage. Osteoarthr. Cartil. 2016, 24, 1981–1988. [Google Scholar] [CrossRef] [PubMed]
- Behrendt, P.; Feldheim, M.; Preusse-Prange, A.; Weitkamp, J.T.; Haake, M.; Eglin, D.; Rolauffs, B.; Fay, J.; Seekamp, A.; Grodzinsky, A.J.; et al. Chondrogenic potential of IL-10 in mechanically injured cartilage and cellularized collagen ACI grafts. Osteoarthr. Cartil. 2018, 26, 264–275. [Google Scholar] [CrossRef] [PubMed]
- King, L.K.; March, L.; Anandacoomarasamy, A. Obesity & osteoarthritis. Indian J. Med. Res. 2013, 138, 185–193. [Google Scholar] [PubMed]
- Schett, G.; Kleyer, A.; Perricone, C.; Sahinbegovic, E.; Iagnocco, A.; Zwerina, J.; Lorenzini, R.; Aschenbrenner, F.; Berenbaum, F.; D’Agostino, M.A.; et al. Diabetes is an independent predictor for severe osteoarthritis: Results from a longitudinal cohort study. Diabetes Care 2013, 36, 403–409. [Google Scholar] [CrossRef] [PubMed]
- Petersen, A.M.; Pedersen, B.K. The anti-inflammatory effect of exercise. J. Appl. Physiol. (1985) 2005, 98, 1154–1162. [Google Scholar] [CrossRef] [Green Version]
- Coggon, D.; Reading, I.; Croft, P.; McLaren, M.; Barrett, D.; Cooper, C. Knee osteoarthritis and obesity. Int. J. Obes. Relat. Metab. Disord. 2001, 25, 622–627. [Google Scholar] [CrossRef] [Green Version]
- Hotamisligil, G.S.; Shargill, N.S.; Spiegelman, B.M. Adipose expression of tumor necrosis factor-alpha: Direct role in obesity-linked insulin resistance. Science 1993, 259, 87–91. [Google Scholar] [CrossRef]
- Rosa, S.C.; Goncalves, J.; Judas, F.; Mobasheri, A.; Lopes, C.; Mendes, A.F. Impaired glucose transporter-1 degradation and increased glucose transport and oxidative stress in response to high glucose in chondrocytes from osteoarthritic versus normal human cartilage. Arthritis Res. Ther. 2009, 11, R80. [Google Scholar] [CrossRef] [Green Version]
- Sophia Fox, A.J.; Bedi, A.; Rodeo, S.A. The basic science of articular cartilage: Structure, composition, and function. Sports Health 2009, 1, 461–468. [Google Scholar] [CrossRef]
- Maroudas, A. Distribution and diffusion of solutes in articular cartilage. Biophys. J. 1970, 10, 365–379. [Google Scholar] [CrossRef]
- Faryna, A.; Goldenberg, K. Joint Fluid. In Clinical Methods: The History, Physical, and Laboratory Examinations; Walker, H.K., Hall, W.D., Hurst, J.W., Eds.; Butterworth Publishers: Boston, MA, USA, 1990. [Google Scholar]
- Akyol, S.; Comertoglu, I.; Firat, R.; Cakmak, O.; Yukselten, Y.; Erden, G.; Ugurcu, V.; Demircan, K. Effect of insulin on the mRNA expression of procollagen N-proteinases in chondrosarcoma OUMS-27 cells. Oncol. Lett. 2015, 10, 1091–1096. [Google Scholar] [CrossRef] [Green Version]
- Kunisada, T.; Miyazaki, M.; Mihara, K.; Gao, C.; Kawai, A.; Inoue, H.; Namba, M. A new human chondrosarcoma cell line (OUMS-27) that maintains chondrocytic differentiation. Int. J. Cancer 1998, 77, 854–859. [Google Scholar] [CrossRef] [Green Version]
- Cakmak, O.; Comertoglu, I.; Firat, R.; Erdemli, H.K.; Kursunlu, S.F.; Akyol, S.; Ugurcu, V.; Altuntas, A.; Adam, B.; Demircan, K. The Investigation of ADAMTS16 in Insulin-Induced Human Chondrosarcoma Cells. Cancer Biother. Radiopharm. 2015, 30, 255–260. [Google Scholar] [CrossRef] [Green Version]
- Schulze-Tanzil, G. Activation and dedifferentiation of chondrocytes: Implications in cartilage injury and repair. Ann. Anat. 2009, 191, 325–338. [Google Scholar] [CrossRef]
- Kanbara, K.; Otsuki, Y.; Watanabe, M.; Yokoe, S.; Mori, Y.; Asahi, M.; Neo, M. GABAB receptor regulates proliferation in the high-grade chondrosarcoma cell line OUMS-27 via apoptotic pathways. BMC Cancer 2018, 18, 263. [Google Scholar] [CrossRef]
- Fukumoto, S.; Kanbara, K.; Neo, M. Synergistic anti-proliferative effects of mTOR and MEK inhibitors in high-grade chondrosarcoma cell line OUMS-27. Acta Histochem. 2018, 120, 142–150. [Google Scholar] [CrossRef]
- Lu, Y.; Li, F.; Xu, T.; Sun, J. miRNA-497 Negatively Regulates the Growth and Motility of Chondrosarcoma Cells by Targeting Cdc25A. Oncol. Res. 2016, 23, 155–163. [Google Scholar] [CrossRef]
- Kurita, T.; Yamamura, H.; Suzuki, Y.; Giles, W.R.; Imaizumi, Y. The ClC-7 Chloride Channel Is Downregulated by Hypoosmotic Stress in Human Chondrocytes. Mol. Pharmacol. 2015, 88, 113–120. [Google Scholar] [CrossRef]
- Inayama, M.; Suzuki, Y.; Yamada, S.; Kurita, T.; Yamamura, H.; Ohya, S.; Giles, W.R.; Imaizumi, Y. Orai1-Orai2 complex is involved in store-operated calcium entry in chondrocyte cell lines. Cell Calcium 2015, 57, 337–347. [Google Scholar] [CrossRef]
- Altuntas, A.; Halacli, S.O.; Cakmak, O.; Erden, G.; Akyol, S.; Ugurcu, V.; Hirohata, S.; Demircan, K. Interleukin-1beta induced nuclear factor-kappaB binds to a disintegrin-like and metalloproteinase with thrombospondin type 1 motif 9 promoter in human chondrosarcoma cells. Mol. Med. Rep. 2015, 12, 595–600. [Google Scholar] [CrossRef]
- Suzuki, Y.; Ohya, S.; Yamamura, H.; Giles, W.R.; Imaizumi, Y. A New Splice Variant of Large Conductance Ca2+-activated K+ (BK) Channel alpha Subunit Alters Human Chondrocyte Function. J. Biol. Chem. 2016, 291, 24247–24260. [Google Scholar] [CrossRef]
- Masutani, T.; Tanaka, Y.T.; Kojima, H.; Tsuboi, M.; Hara, A.; Niwa, M. Cynaropicrin is dual regulator for both degradation factors and synthesis factors in the cartilage metabolism. Life Sci. 2016, 158, 70–77. [Google Scholar] [CrossRef]
- Koizumi, H.; Arito, M.; Endo, W.; Kurokawa, M.S.; Okamoto, K.; Omoteyama, K.; Suematsu, N.; Beppu, M.; Kato, T. Effects of tofacitinib on nucleic acid metabolism in human articular chondrocytes. Mod. Rheumatol. 2015, 25, 522–527. [Google Scholar] [CrossRef]
- Wang, Y.; Lou, S. Direct protective effect of interleukin-10 on articular chondrocytes in vitro. Chin. Med. J. (Engl.) 2001, 114, 723–725. [Google Scholar]
- John, T.; Muller, R.D.; Oberholzer, A.; Zreiqat, H.; Kohl, B.; Ertel, W.; Hostmann, A.; Tschoeke, S.K.; Schulze-Tanzil, G. Interleukin-10 modulates pro-apoptotic effects of TNF-alpha in human articular chondrocytes in vitro. Cytokine 2007, 40, 226–234. [Google Scholar] [CrossRef]
- Radons, J.; Falk, W.; Schubert, T.E. Interleukin-10 does not affect IL-1-induced interleukin-6 and metalloproteinase production in human chondrosarcoma cells, SW1353. Int. J. Mol. Med. 2006, 17, 377–383. [Google Scholar] [CrossRef]
- Huang, T.C.; Chang, W.T.; Hu, Y.C.; Hsieh, B.S.; Cheng, H.L.; Yen, J.H.; Chiu, P.R.; Chang, K.L. Zinc Protects Articular Chondrocytes through Changes in Nrf2-Mediated Antioxidants, Cytokines and Matrix Metalloproteinases. Nutrients 2018, 10, 471. [Google Scholar] [CrossRef]
- Iannone, F.; De Bari, C.; Dell’Accio, F.; Covelli, M.; Cantatore, F.P.; Patella, V.; Lo Bianco, G.; Lapadula, G. Interleukin-10 and interleukin-10 receptor in human osteoarthritic and healthy chondrocytes. Clin. Exp. Rheumatol. 2001, 19, 139–145. [Google Scholar]
- Muller, R.D.; John, T.; Kohl, B.; Oberholzer, A.; Gust, T.; Hostmann, A.; Hellmuth, M.; Laface, D.; Hutchins, B.; Laube, G.; et al. IL-10 overexpression differentially affects cartilage matrix gene expression in response to TNF-alpha in human articular chondrocytes in vitro. Cytokine 2008, 44, 377–385. [Google Scholar] [CrossRef] [PubMed]
- Ortved, K.F.; Begum, L.; Stefanovski, D.; Nixon, A.J. AAV-mediated Overexpression of IL-10 Mitigates the Inflammatory Cascade in Stimulated Equine Chondrocyte Pellets. Curr. Gene Ther. 2018, 18, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.J.; Chan, D.C.; Lan, K.C.; Wang, C.C.; Chen, C.M.; Chao, S.C.; Tsai, K.S.; Yang, R.S.; Liu, S.H. PPARgamma is involved in the hyperglycemia-induced inflammatory responses and collagen degradation in human chondrocytes and diabetic mouse cartilages. J. Orthop. Res. 2015, 33, 373–381. [Google Scholar] [CrossRef] [PubMed]
- Van Vulpen, L.F.D.; Popov-Celeketic, J.; van Meegeren, M.E.R.; Coeleveld, K.; van Laar, J.M.; Hack, C.E.; Schutgens, R.E.G.; Mastbergen, S.C.; Lafeber, F. A fusion protein of interleukin-4 and interleukin-10 protects against blood-induced cartilage damage in vitro and in vivo. J. Thromb. Haemost. 2017, 15, 1788–1798. [Google Scholar] [CrossRef] [PubMed]
- Itoh, K.; Hirohata, S. The role of IL-10 in human B cell activation, proliferation, and differentiation. J. Immunol. 1995, 154, 4341–4350. [Google Scholar] [PubMed]
- Stephanou, A.; Brar, B.K.; Knight, R.A.; Latchman, D.S. Opposing actions of STAT-1 and STAT-3 on the Bcl-2 and Bcl-x promoters. Cell Death Differ. 2000, 7, 329–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stephanou, A.; Latchman, D.S. Opposing actions of STAT-1 and STAT-3. Growth Factors 2005, 23, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Zheng, F.; Shen, J.; Cheng, S.; Yu, P.; You, M.; Cai, X. Inhibition of osteoarthritis chondrocyte proliferation by IL-10 via modulating NF-κB and related mechanisms. Int. J. Clin. Exp. Med. 2017, 10, 11688–11695. [Google Scholar]
- Mobasheri, A.; Vannucci, S.J.; Bondy, C.A.; Carter, S.D.; Innes, J.F.; Arteaga, M.F.; Trujillo, E.; Ferraz, I.; Shakibaei, M.; Martin-Vasallo, P. Glucose transport and metabolism in chondrocytes: A key to understanding chondrogenesis, skeletal development and cartilage degradation in osteoarthritis. Histol. Histopathol. 2002, 17, 1239–1267. [Google Scholar] [PubMed]
- Laiguillon, M.C.; Courties, A.; Houard, X.; Auclair, M.; Sautet, A.; Capeau, J.; Feve, B.; Berenbaum, F.; Sellam, J. Characterization of diabetic osteoarthritic cartilage and role of high glucose environment on chondrocyte activation: Toward pathophysiological delineation of diabetes mellitus-related osteoarthritis. Osteoarthr. Cartil. 2015, 23, 1513–1522. [Google Scholar] [CrossRef] [PubMed]
- Heywood, H.K.; Nalesso, G.; Lee, D.A.; Dell’accio, F. Culture expansion in low-glucose conditions preserves chondrocyte differentiation and enhances their subsequent capacity to form cartilage tissue in three-dimensional culture. Biores. Open Access 2014, 3, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Stevens, R.L.; Nissley, S.P.; Kimura, J.H.; Rechler, M.M.; Caplan, A.I.; Hascall, V.C. Effects of insulin and multiplication-stimulating activity on proteoglycan biosynthesis in chondrocytes from the Swarm rat chondrosarcoma. J. Biol. Chem. 1981, 256, 2045–2052. [Google Scholar] [PubMed]
- Rosa, S.C.; Rufino, A.T.; Judas, F.; Tenreiro, C.; Lopes, M.C.; Mendes, A.F. Expression and function of the insulin receptor in normal and osteoarthritic human chondrocytes: Modulation of anabolic gene expression, glucose transport and GLUT-1 content by insulin. Osteoarthr. Cartil. 2011, 19, 719–727. [Google Scholar] [CrossRef]
- Phornphutkul, C.; Wu, K.Y.; Gruppuso, P.A. The role of insulin in chondrogenesis. Mol. Cell. Endocrinol. 2006, 249, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Claassen, H.; Schluter, M.; Schunke, M.; Kurz, B. Influence of 17beta-estradiol and insulin on type II collagen and protein synthesis of articular chondrocytes. Bone 2006, 39, 310–317. [Google Scholar] [CrossRef]
- Ribeiro, M.; Lopez de Figueroa, P.; Blanco, F.J.; Mendes, A.F.; Carames, B. Insulin decreases autophagy and leads to cartilage degradation. Osteoarthr. Cartil. 2016, 24, 731–739. [Google Scholar] [CrossRef]
- Otsu, K.; Geary, E.S.; Stevens, R.L. Aberrant regulation of the metabolism of the insulin receptor in Swarm rat chondrosarcoma chondrocytes. Biochem. J. 1988, 254, 203–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Without Stimulation (w/o) | HI | HI+IL-10 (Co-Stimulation) | IL-10 |
|---|---|---|---|
| NG (Control Group) | NG + 10 µg mL−1 insulin | NG + 10 µg mL−1 insulin +10 ng mL−1 IL-10 | NG + 10 ng mL−1 IL-10 |
| HG | HG + 10 µg mL−1 insulin | HG + 10 µg mL−1 insulin + 10 ng mL−1 IL-10 | HG + 10 ng mL−1 IL-10 |
| Target | Primary Antibody | Dilution | Secondary Antibody | Dilution |
|---|---|---|---|---|
| col I | goat anti human, Biozol, Eching, Germany | 1:50 | donkey anti goat, Alexa Fluor 555, ThermoFisher Scientific, Darmstadt, Germany | 1:200 |
| col II | rabbit anti human, Acris, Herford, Germany | 1:50 | donkey anti rabbit, Alexa Fluor 488, ThermoFisher Scientific, Darmstadt, Germany | 1:200 |
| PG | mouse anti human, Merck, Darmstadt, Germany | 1:70 | donkey anti mouse Cy3, Dianova, Hamburg, Germany | 1:200 |
| SOX9 | rabbit anti human, Merck, Darmstadt, Germany | 1:100 | donkey anti rabbit, Alexa Fluor 488, Dianova, Hamburg, Germany | 1:200 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silawal, S.; Willauschus, M.; Schulze-Tanzil, G.; Gögele, C.; Geßlein, M.; Schwarz, S. IL-10 Could Play a Role in the Interrelation between Diabetes Mellitus and Osteoarthritis. Int. J. Mol. Sci. 2019, 20, 768. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20030768
Silawal S, Willauschus M, Schulze-Tanzil G, Gögele C, Geßlein M, Schwarz S. IL-10 Could Play a Role in the Interrelation between Diabetes Mellitus and Osteoarthritis. International Journal of Molecular Sciences. 2019; 20(3):768. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20030768
Chicago/Turabian StyleSilawal, Sandeep, Maximilian Willauschus, Gundula Schulze-Tanzil, Clemens Gögele, Markus Geßlein, and Silke Schwarz. 2019. "IL-10 Could Play a Role in the Interrelation between Diabetes Mellitus and Osteoarthritis" International Journal of Molecular Sciences 20, no. 3: 768. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20030768