Response of Tibetan Wild Barley Genotypes to Drought Stress and Identification of Quantitative Trait Loci by Genome-Wide Association Analysis

Abstract
:1. Introduction
2. Results
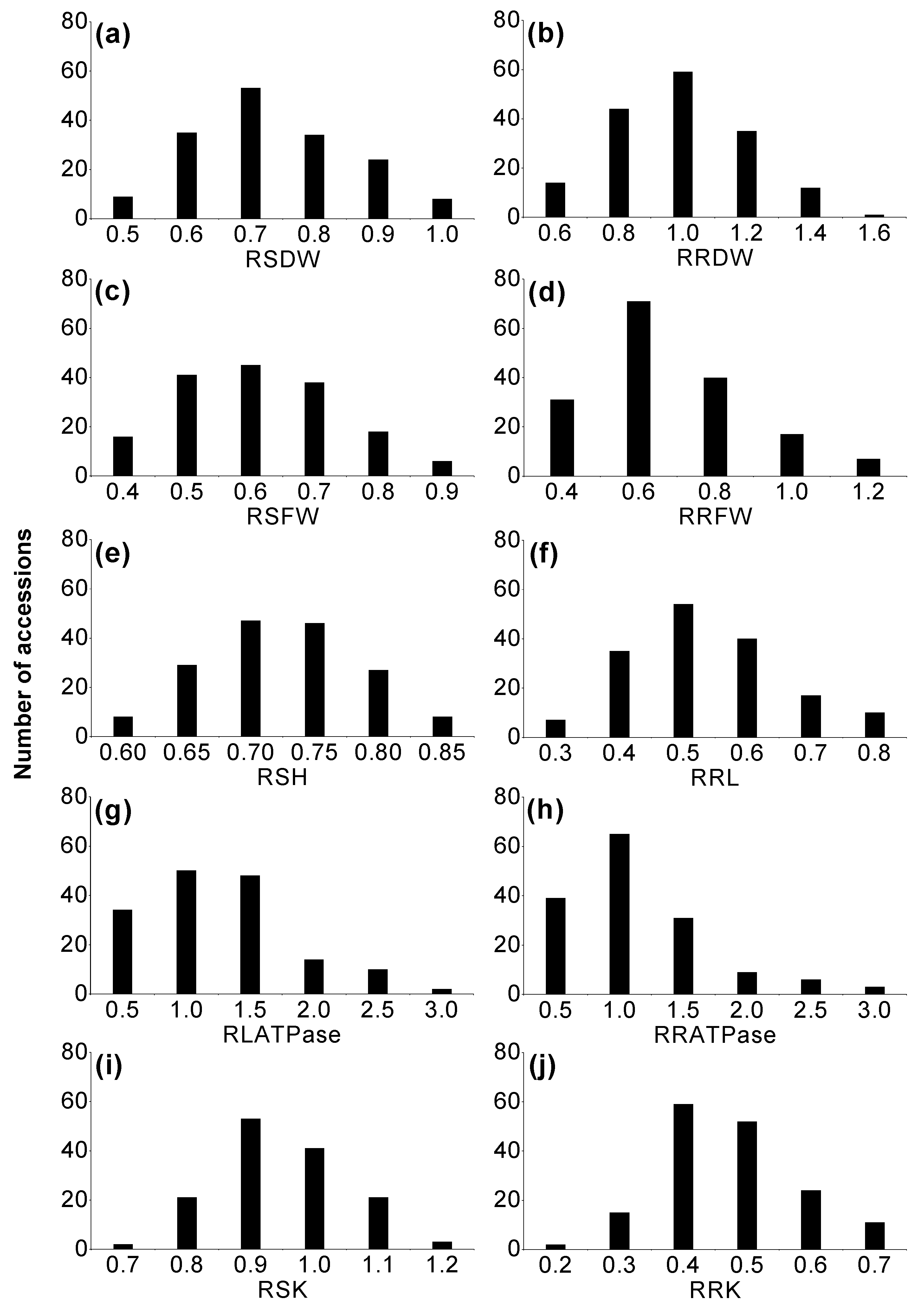
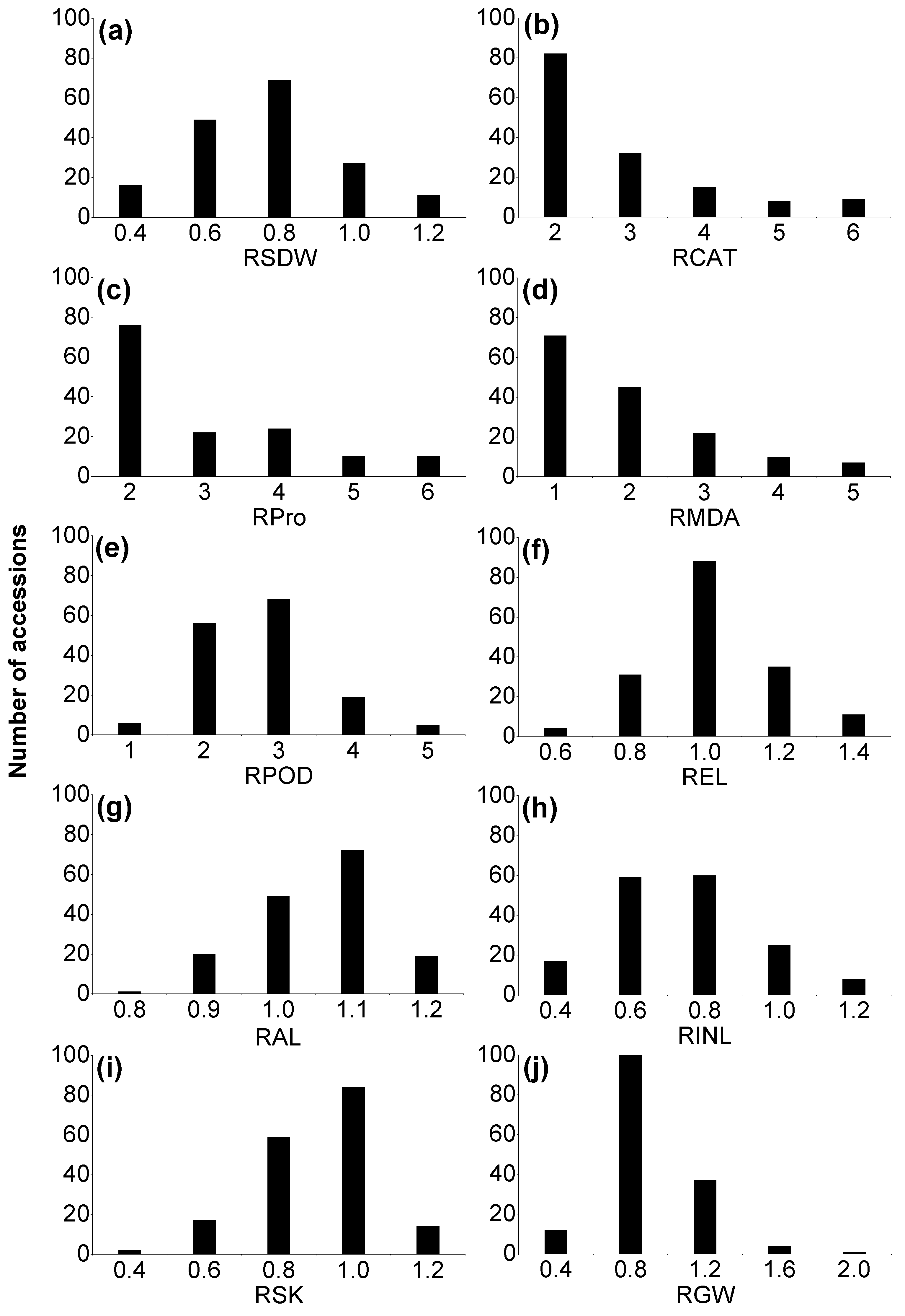
2.1. Phenotypic Variation of Morphological Traits in Wild Barley
2.2. Variation in H+K+-ATPase and Antioxidant Enzyme Activities, Malondialdehyde (MDA) and Soluble Protein Contents
2.3. Phenotypic Variation in K+ Concentration in Wild Barley
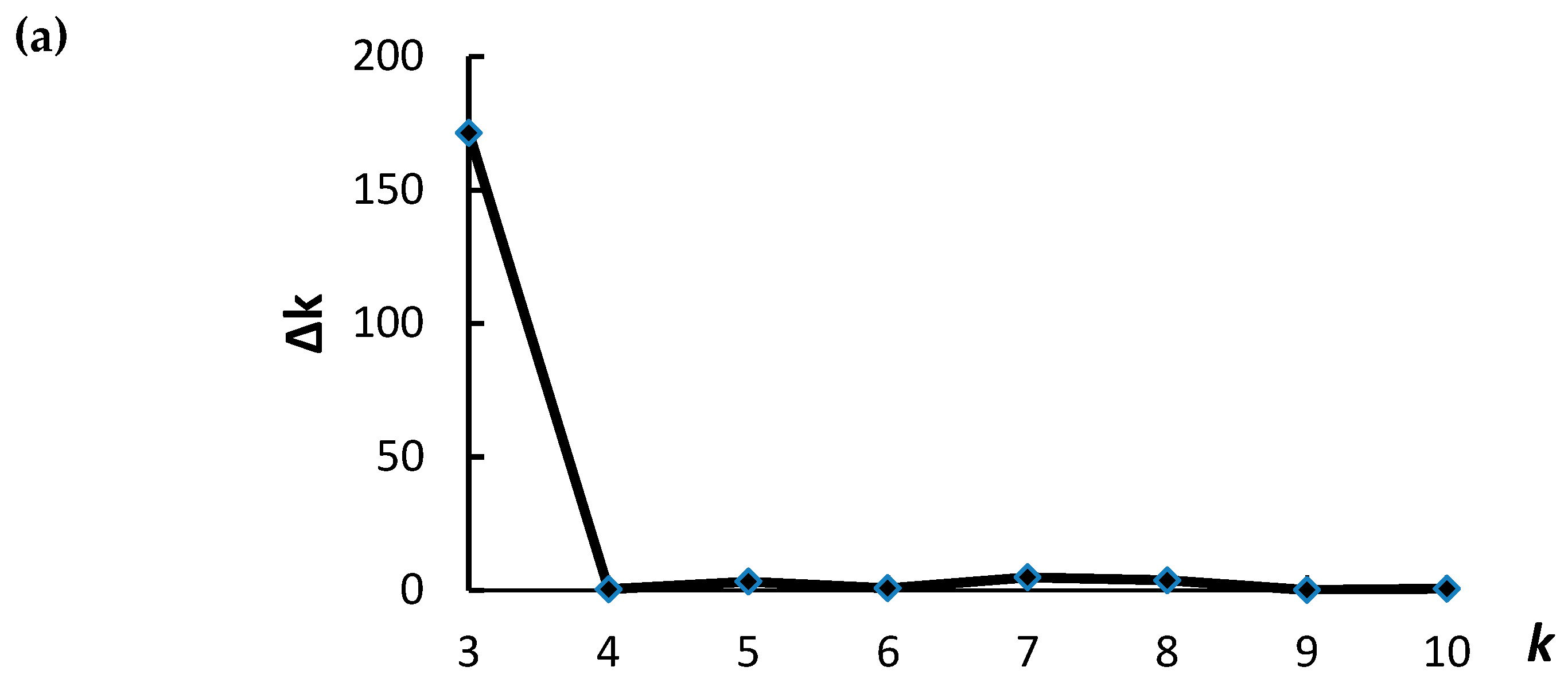
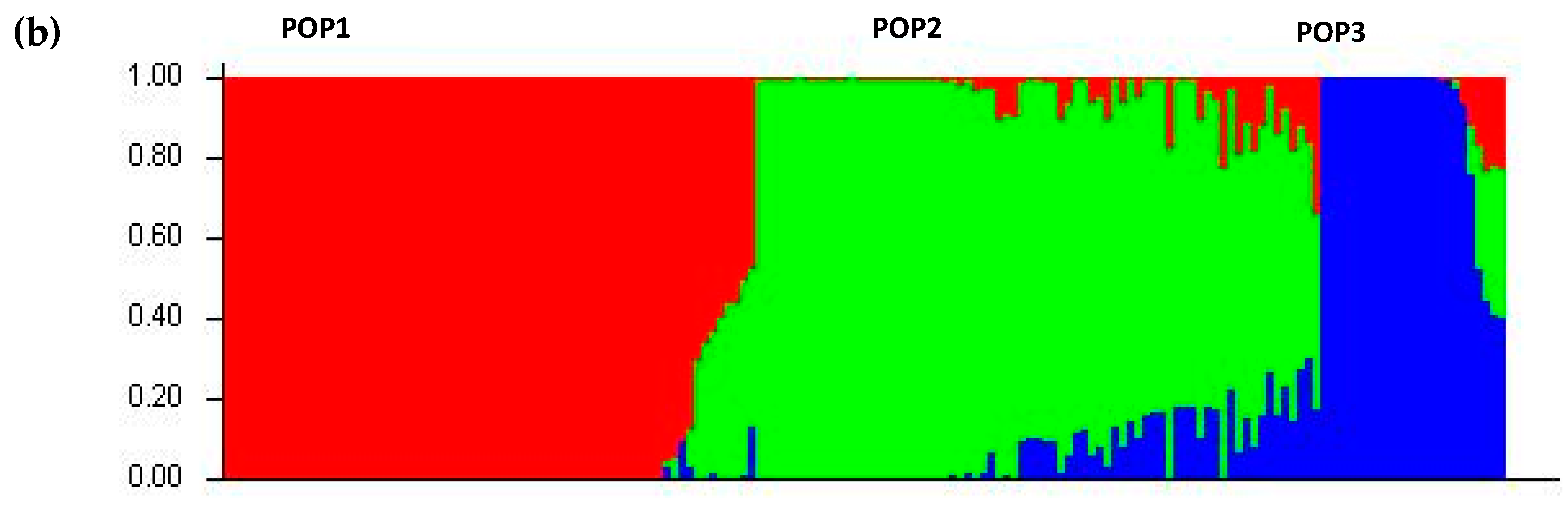
2.4. Population Structure and Linkage Disequilibrium Decay
2.5. Identification of Loci Associated with Morphological Traits
2.6. Identification of Loci Associated with H+K+-ATPase and the Antioxidant Enzyme Activities, Malondialdehyde and Soluble Protein Contents
2.7. Identification of Loci Associated with K+ Concentration
3. Discussion
3.1. Quantitative Trait Loci (QTL) Associated with Morphological Traits under Drought Stress
3.2. QTL Associated with H+K+-ATPase under Drought Stress
3.3. QTL Associated with K+ Concentration under Drought Stress
3.4. QTL Associated with Peroxidase (POD) and Catalase (CAT) Activity under Drought Stress
3.5. The Effect of Population Structure and Linkage Disequilibrium (LD) on the Association Analysis
3.6. Comparison of Genome-Wide Association (GWA) Results with Reported Meta-QTL and Exploration of Candidate Genes
4. Materials and Methods
4.1. Plant Materials
4.2. Hydroponic Experiment
4.3. Pot Experiment
4.4. Determination of H+ K+-ATPase Activity
4.5. Determination of Antioxidant Enzyme Activity, and Malondialdehyde (MDA) and Soluble Protein Contents
4.6. Determination of K+ Concentration
4.7. Population Structure and Linkage Disequilibrium (LD)
4.8. Genome-Wide (GWA) Association Analysis
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Cattivelli, L.; Rizza, F.; Badeck, F.W.; Mazzucotelli, E.; Mastrangelo, A.M.; Francia, E.; Mare, C.; Tondelli, A.; Stanca, A.M. Drought tolerance improvement in crop plants: An integrated view from breeding to genomics. Field Crops Res. 2008, 105, 1–14. [Google Scholar] [CrossRef]
- Hu, H.H.; Xiong, L.Z. Genetic Engineering and Breeding of Drought-Resistant Crops. Annu. Rev. Plant Biol. 2014, 65, 715–741. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Jin, Z.Q.; Zhao, J.; Zhang, G.P.; Wu, F.B. Physiological and biochemical responses to drought stress in cultivated and Tibetan wild barley. Plant Growth Regul. 2015, 75, 567–574. [Google Scholar] [CrossRef]
- Xue, D.W.; Zhang, X.Q.; Lu, X.L.; Chen, G.; Chen, Z.H. Molecular and Evolutionary Mechanisms of Cuticular Wax for Plant Drought Tolerance. Front. Plant Sci. 2017, 8, 621. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.H.; Chen, G.; Dai, F.; Wang, Y.Z.; Hills, A.; Ruan, Y.L.; Zhang, G.P.; Franks, P.J.; Nevo, E.; Blatt, M.R. Molecular Evolution of Grass Stomata. Trends Plant Sci. 2017, 22, 124–139. [Google Scholar] [CrossRef] [PubMed]
- Rogers, E.D.; Benfey, P.N. Regulation of plant root system architecture: Implications for crop advancement. Curr. Opin. Biotechnol. 2015, 32, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Noctor, G.; Mhamdi, A.; Foyer, C.H. The Roles of Reactive Oxygen Metabolism in Drought: Not So Cut and Dried. Plant Physiol. 2014, 164, 1636–1648. [Google Scholar] [CrossRef] [Green Version]
- Rampino, P.; Pataleo, S.; Gerardi, C.; Mita, G.; Perrotta, C. Drought stress response in wheat: Physiological and molecular analysis of resistant and sensitive genotypes. Plant Cell Environ. 2006, 29, 2143–2152. [Google Scholar] [CrossRef]
- Ellis, R.P.; Forster, B.P.; Robinson, D.; Handley, L.L.; Gordon, D.C.; Russell, J.R.; Powell, W. Wild barley: A source of genes for crop improvement in the 21st century? J. Exp. Bot. 2000, 51, 9–17. [Google Scholar] [CrossRef]
- He, X.Y.; Zeng, J.B.; Cao, F.B.; Ahmed, I.M.; Zhang, G.; Vincze, E.; Wu, F.B. HvEXPB7, a novel β-expansin gene revealed by the root hair transcriptome of Tibetan wild barley, improves root hair growth under drought stress. J. Exp. Bot. 2015, 66, 7405–7419. [Google Scholar] [CrossRef]
- Ahmed, I.M.; Nadira, U.A.; Bibi, N.; Cao, F.; He, X.; Zhang, G.; Wu, F.B. Secondary metabolism and antioxidants are involved in the tolerance to drought and salinity, separately and combined, in Tibetan wild barley. Environ. Exp. Bot. 2015, 111, 1–12. [Google Scholar] [CrossRef]
- Zhang, M.; Mao, W.H.; Zhang, G.P.; Wu, F.B. Development and Characterization of Polymorphic EST-SSR and Genomic SSR Markers for Tibetan Annual Wild Barley. PLoS ONE 2014, 9, e94881. [Google Scholar] [CrossRef] [PubMed]
- Gawenda, I.; Thorwarth, P.; Guenther, T.; Ordon, F.; Schmid, K.J. Genome-wide association studies in elite varieties of German winter barley using single-marker and haplotype-based methods. Plant Breed. 2015, 134, 28–39. [Google Scholar] [CrossRef]
- Wang, Q.X.; Zhao, H.; Jiang, J.P.; Xu, J.Y.; Xie, W.B.; Fu, X.K.; Liu, C.; He, Y.Q.; Wang, G.W. Genetic Architecture of Natural Variation in Rice Nonphotochemical Quenching Capacity Revealed by Genome-Wide Association Study. Front. Plant Sci. 2017, 8, 1773. [Google Scholar] [CrossRef] [PubMed]
- Hu, G.H.; Li, Z.; Lu, Y.C.; Li, C.X.; Gong, S.C.; Yan, S.Q.; Li, G.L.; Wang, M.Q.; Ren, H.L.; Guan, H.T.; et al. Genome-wide association study Identified multiple Genetic Loci on Chilling Resistance During Germination in Maize. Sci. Rep. 2017, 7, 10840. [Google Scholar] [CrossRef] [PubMed]
- Kadam, S.; Singh, K.; Shukla, S.; Goel, S.; Vikram, P.; Pawar, V.; Gaikwad, K.; Khanna-Chopra, R.; Singh, N. Genomic associations for drought tolerance on the short arm of wheat chromosome 4B. Funct. Integr. Genomic. 2012, 12, 447–464. [Google Scholar] [CrossRef] [PubMed]
- Varshney, R.K.; Paulo, M.J.; Grando, S.; van Eeuwijk, F.A.; Keizer, L.C.P.; Guo, P.; Ceccarelli, S.; Kilian, A.; Baum, M.; Graner, A. Genome wide association analyses for drought tolerance related traits in barley (Hordeum vulgare L.). Field Crops Res. 2012, 126, 171–180. [Google Scholar] [CrossRef]
- Wang, R.; Leng, Y.Q.; Ali, S.; Wang, M.N.; Zhong, S.B. Genome-wide association mapping of spot blotch resistance to three different pathotypes of Cochliobolus sativus in the USDA barley core collection. Mol. Breed. 2017, 37, 44. [Google Scholar] [CrossRef]
- Li, W.T.; Liu, C.J.; Liu, Y.X.; Pu, Z.E.; Dai, S.F.; Wang, J.R.; Lan, X.J.; Zheng, Y.L.; Wei, Y.M. Meta-analysis of QTL associated with tolerance to abiotic stresses in barley. Euphytica 2013, 189, 31–49. [Google Scholar] [CrossRef]
- Honsdorf, N.; March, T.J.; Hecht, A.; Eglinton, J.; Pillen, K. Evaluation of juvenile drought stress tolerance and genotyping by sequencing with wild barley introgression lines. Mol. Breed. 2014, 34, 1475–1495. [Google Scholar] [CrossRef]
- Hu, X.; Zuo, J.; Wang, J.; Liu, L.; Sun, G.; Li, C.; Ren, X.; Sun, D. Multi-Locus Genome-Wide Association Studies for 14 Main Agronomic Traits in Barley. Front. Plant Sci. 2018, 9, 1683. [Google Scholar] [CrossRef] [PubMed]
- Jabbari, M.; Fakheri, B.A.; Aghnoum, R.; Mahdi Nezhad, N.; Ataei, R. GWAS analysis in spring barley (Hordeum vulgare L.) for morphological traits exposed to drought. PLoS ONE 2018, 13, e0204952. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Sun, H.Y.; Dai, H.X.; Zhang, G.P.; Wu, F.B. Difference in response to drought stress among Tibet wild barley genotypes. Euphytica 2010, 172, 395–403. [Google Scholar] [CrossRef]
- Hoad, S.P.; Russell, G.; Lucas, M.E.; Bingham, I.J. The management of wheat, barley, and oat root systems. Adv. Agron. 2001, 74, 193–246. [Google Scholar]
- Zhan, A.; Schneider, H.; Lynch, J.P. Reduced Lateral Root Branching Density Improves Drought Tolerance in Maize. Plant Physiol. 2015, 168, 1603–1615. [Google Scholar] [CrossRef] [PubMed]
- Lambers, H.; Atkin, O.K.; Millenaar, F.F. Respiratory patterns in roots in relation to their functioning. In Plant Roots; Waisel, Y., Eshel, A., Kafkafi, U., Eds.; Marcel Dekker: New York, NY, USA, 2002; pp. 521–552. [Google Scholar]
- Sehgal, A.; Sita, K.; Siddique, K.H.M.; Kumar, R.; Bhogireddy, S.; Varshney, R.K.; HanumanthaRao, B.; Nair, R.M.; Prasad, P.V.V.; Nayyar, H. Drought or/and Heat-Stress Effects on Seed Filling in Food Crops: Impacts on Functional Biochemistry, Seed Yields, and Nutritional Quality. Front. Plant Sci. 2018, 9, 1705. [Google Scholar] [CrossRef] [PubMed]
- Duffus, C.M.; Cochrane, M.P. Formation of the barley grain-morphology, physiology, and biochemistry. In Barley: Chemistry and technology; MacGregor, A.W., Bhatty, R.S., Eds.; American Association of Cereal Chemists: St. Paul, MN, USA, 1993; pp. 31–72. [Google Scholar]
- Abebe, T.; Wise, R.P.; Skadsen, R.W. Comparative Transcriptional Profiling Established the Awn as the Major Photosynthetic Organ of the Barley Spike While the Lemma and the Palea Primarily Protect the Seed. Plant Genome 2009, 2, 247–259. [Google Scholar] [CrossRef] [Green Version]
- Araus, J.L.; Brown, H.R.; Febrero, A.; Bort, J.; Serret, M.D. Ear photosynthesis, carbon isotope discrimination and the contribution of respiratory CO2 to differences in grain mass in durum-wheat. Plant Cell Environ. 1993, 16, 383–392. [Google Scholar] [CrossRef]
- Tambussi, E.A.; Bort, J.; Jose Guiamet, J.; Nogues, S.; Luis Araus, J. The photosynthetic role of ears in C(3) cereals: Metabolism, water use efficiency and contribution to grain yield. Crit. Rev. Plant Sci. 2007, 26, 1–16. [Google Scholar] [CrossRef]
- Hein, J.A.; Sherrard, M.E.; Manfredi, K.P.; Abebe, T. The fifth leaf and spike organs of barley (Hordeum vulgare L.) display different physiological and metabolic responses to drought stress. BMC Plant Biol. 2016, 16. [Google Scholar] [CrossRef]
- Wang, Z.; Gu, Y.J.; Gao, Y.Z. Structure and photosynthetic characteristics of awns of wheat and barley. Acta Bot. Sin. 1993, 35, 921–928. [Google Scholar]
- Palmgren, M.G. Plant plasma membrane H+-ATPases: Powerhouses for nutrient uptake. Annu. Rev. Plant Physiol. Plant Mol. Biol. 2001, 52, 817–845. [Google Scholar] [CrossRef] [PubMed]
- Briskin, D.P. The plasma-membrane H+-ATPase of higher-plant cells-biochemistry and transport function. Biochim. Biophys. Acta 1990, 1019, 95–109. [Google Scholar] [CrossRef]
- Gong, D.S.; Xiong, Y.C.; Ma, B.L.; Wang, T.M.; Ge, J.P.; Qin, X.L.; Li, P.F.; Kong, H.Y.; Li, Z.Z.; Li, F.M. Early activation of plasma membrane H+-ATPase and its relation to drought adaptation in two contrasting oat (Avena sativa L.) genotypes. Environ. Exp. Bot. 2010, 69, 1–8. [Google Scholar] [CrossRef]
- Cai, S.G.; Chen, G.; Wang, Y.Y.; Huang, Y.Q.; Marchant, D.B.; Wang, Y.Z.; Yang, Q.; Dai, F.; Hills, A.; Franks, P.J.; et al. Evolutionary Conservation of ABA Signaling for Stomatal Closure. Plant Physiol. 2017, 174, 732–747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, H.J.; Chen, K.M.; Chen, G.C.; Wang, S.M.; Zhang, C.L. Drought stress stimulates p-nitrophenyl phosphate hydrolysis rate of the plasma membrane H+-ATPase from wheat leaves. Plant Growth Regul. 2003, 40, 139–145. [Google Scholar] [CrossRef]
- Ahmed, I.M.; Nadira, U.A.; Qiu, C.W.; Cao, F.B.; Zhang, G.; Holford, P.; Wu, F.B. Tolerance to Drought, Low pH and Al Combined Stress in Tibetan Wild Barley Is Associated with Improvement of ATPase and Modulation of Antioxidant Defense System. Int. J. Mol. Sci. 2018, 19, 3553. [Google Scholar] [CrossRef]
- Lebaudy, A.; Very, A.-A.; Sentenac, H. K+ channel activity in plants: Genes, regulations and functions. FEBS Lett. 2007, 581, 2357–2366. [Google Scholar] [CrossRef]
- Chen, Z.H.; Wang, Y.Z.; Wang, J.W.; Babla, M.; Zhao, C.C.; Garcia-Mata, C.; Sani, E.; Differ, C.; Mak, M.; Hills, A.; et al. Nitrate reductase mutation alters potassium nutrition as well as nitric oxide-mediated control of guard cell ion channels in Arabidopsis. New Phytol. 2016, 209, 1456–1469. [Google Scholar] [CrossRef]
- Shabala, S. Signalling by potassium: Another second messenger to add to the list? J. Exp. Bot. 2017, 68, 4003–4007. [Google Scholar] [CrossRef] [PubMed]
- Benlloch-Gonzalez, M.; Arquero, O.; Fournier, J.M.; Barranco, D.; Benlloch, M. K+ starvation inhibits water-stress-induced stomatal closure. J Plant Physiol 2008, 165, 623–630. [Google Scholar] [CrossRef] [PubMed]
- Mian, A.; Oomen, R.J.F.J.; Isayenkov, S.; Sentenac, H.; Maathuis, F.J.M.; Very, A.-A. Over-expression of an Na+- and K+-permeable HKT transporter in barley improves salt tolerance. Plant J. 2011, 68, 468–479. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.H.; Cai, S.G.; Wang, G.; Wang, F.F.; Dong, F.B.; Mak, M.; Holford, P.; Ji, J.; Salih, A.; Zhou, M.X.; et al. Halophytic NHXs confer salt tolerance by altering cytosolic and vacuolar K+ and Na+ in Arabidopsis root cell. Plant Growth Regul. 2017, 82, 333–351. [Google Scholar] [CrossRef]
- Wang, F.F.; Chen, Z.H.; Liu, X.H.; Colmer, T.D.; Zhou, M.X.; Shabala, S. Tissue-specific root ion profiling reveals essential roles of the CAX and ACA calcium transport systems in response to hypoxia in Arabidopsis. J. Exp. Bot. 2016, 67, 3747–3762. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.F.; Chen, Z.H.; Shabala, S. Hypoxia Sensing in Plants: On a Quest for Ion Channels as Putative Oxygen Sensors. Plant Cell Physiol. 2017, 58, 1126–1142. [Google Scholar] [CrossRef] [PubMed]
- Gargallo-Garriga, A.; Sardans, J.; Perez-Trujillo, M.; Oravec, M.; Urban, O.; Jentsch, A.; Kreyling, J.; Beierkuhnlein, C.; Parella, T.; Penuelas, J. Warming differentially influences the effects of drought on stoichiometry and metabolomics in shoots and roots. New Phytol. 2015, 207, 591–603. [Google Scholar] [CrossRef] [Green Version]
- Demidchik, V.; Cuin, T.A.; Svistunenko, D.; Smith, S.J.; Miller, A.J.; Shabala, S.; Sokolik, A.; Yurin, V. Arabidopsis root K+-efflux conductance activated by hydroxyl radicals: Single-channel properties, genetic basis and involvement in stress-induced cell death. J. Cell Sci. 2010, 123, 1468–1479. [Google Scholar] [CrossRef]
- Demidchik, V. Mechanisms and physiological roles of K+ efflux from root cells. J Plant Physiol 2014, 171, 696–707. [Google Scholar] [CrossRef]
- Wu, F.B.; Zhang, G.P.; Dominy, P. Four barley genotypes respond differently to cadmium: Lipid peroxidation and activities of antioxidant capacity. Environ. Exp. Bot. 2003, 50, 67–78. [Google Scholar] [CrossRef]
- Ranney, T.G.; Bassuk, N.L.; Whitlow, T.H. Osmotic adjustment and solute constituents in leaves and roots of water-stressed cherry (prunus) trees. J. Am. Soc. Hort. Sci. 1991, 116, 684–688. [Google Scholar] [CrossRef]
- Sharbel, T.F.; Haubold, B.; Mitchell-Olds, T. Genetic isolation by distance in Arabidopsis thaliana: Biogeography and postglacial colonization of Europe. Mol. Ecol. 2000, 9, 2109–2118. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.C.; Shabala, S.; Koutoulis, A.; Shabala, L.; Zhou, M.X. Meta-analysis of major QTL for abiotic stress tolerance in barley and implications for barley breeding. Planta 2017, 245, 283–295. [Google Scholar] [CrossRef] [PubMed]
- Min, M.K.; Jang, M.; Lee, M.; Lee, J.; Song, K.; Lee, Y.; Choi, K.Y.; Robinson, D.G.; Hwang, I. Recruitment of Arf1-GDP to Golgi by Glo3p-Type ArfGAPs Is Crucial for Golgi Maintenance and Plant Growth. Plant. Physiol. 2013, 161, 676–691. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, X.; Xu, Y.; Chong, K.; Lan, L.; Xue, Y.; Xu, Z. OsAGAP, an ARF-GAP from rice, regulates root development mediated by auxin in Arabidopsis. Plant. Cell Environ. 2005, 28, 147–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osipenkova, O.V.; Ermokhina, O.V.; Belkina, G.G.; Oleskina, Y.P.; Fattakhov, S.G.; Yurina, N.P. Effect of Melaphene on Expression of Elip1 and Elip2 Genes Encoding Chloroplast Light-Induced Stress Proteins in Barley. Appl. Biochem. Microbiol. 2008, 44, 635–641. [Google Scholar] [CrossRef]
- Danilova, M.N.; Kudryakova, N.V.; Voronin, P.Y.; Oelmueller, R.; Kusnetsov, V.V.; Kulaeva, O.N. Membrane receptors of cytokinin and their regulatory role in Arabidopsis thaliana plant response to photooxidative stress under conditions of water deficit. Russ. J. Plant. Physl. 2014, 61, 434–442. [Google Scholar] [CrossRef]
- Yabuta, S.; Ifuku, K.; Takabayashi, A.; Ishihara, S.; Ido, K.; Ishikawa, N.; Endo, T.; Sato, F. Three PsbQ-Like Proteins are Required for the Function of the Chloroplast NAD(P)H Dehydrogenase Complex in Arabidopsis. Plant. Cell Physiol. 2010, 51, 866–876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwarz, N.; Armbruster, U.; Iven, T.; Brueckle, L.; Melzer, M.; Feussner, I.; Jahns, P. Tissue-Specific Accumulation and Regulation of Zeaxanthin Epoxidase in Arabidopsis Reflect the Multiple Functions of the Enzyme in Plastids. Plant. Cell Physiol. 2015, 56, 346–357. [Google Scholar] [CrossRef] [PubMed]
- Teng, K.Q.; Li, J.Z.; Liu, L.; Han, Y.C.; Du, Y.X.; Zhang, J.; Sun, H.Z.; Zhao, Q.Z. Exogenous ABA induces drought tolerance in upland rice: The role of chloroplast and ABA biosynthesis-related gene expression on photosystem II during PEG stress. Acta Physiol. Plant. 2014, 36, 2219–2227. [Google Scholar] [CrossRef]
- Zhao, J.; Wang, X.M. Arabidopsis phospholipase D alpha 1 interacts with the heterotrimeric G-protein alpha-subunit through a motif analogous to the DRY motif in G-protein-coupled receptors. J. Biol. Chem. 2004, 279, 1794–1800. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.H.; Qin, C.B.; Zhao, J.; Wang, X.M. Phospholipase D alpha 1-derived phosphatidic acid interacts with ABI1 phosphatase 2C and regulates abscisic acid signaling. Proc. Natl. Acad. Sci. USA 2004, 101, 9508–9513. [Google Scholar] [CrossRef] [PubMed]
- Penfield, S.; Clements, S.; Bailey, K.J.; Gilday, A.D.; Leegood, R.C.; Gray, J.E.; Graham, I.A. Expression and manipulation of PHOSPHOENOLPYRUVATE CARBOXYKINASE 1 identifies a role for malate metabolism in stomatal closure. Plant J. 2012, 69, 679–688. [Google Scholar] [CrossRef] [PubMed]
- Robinson, D.; Handley, L.L.; Scrimgeour, C.M.; Gordon, D.C.; Forster, B.P.; Ellis, R.P. Using stable isotope natural abundances (delta N-15 and delta C-13) to integrate the stress responses of wild barley (Hordeum spontaneum C. Koch.) genotypes. J. Exp. Bot. 2000, 51, 41–50. [Google Scholar] [PubMed]
- Zhang, X.Z. The measurement and mechanism of lipid peroxidation and SOD, POD and CAT activities in biological system. In Research methodology of crop physiology; Zhang, X.Z., Ed.; Agriculture Press: Beijing, China, 1992; pp. 208–211. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Cai, S.G.; Wu, D.Z.; Jabeen, Z.; Huang, Y.Q.; Huang, Y.C.; Zhang, G.P. Genome-Wide Association Analysis of Aluminum Tolerance in Cultivated and Tibetan Wild Barley. PLoS ONE 2013, 8, e69776. [Google Scholar] [CrossRef] [PubMed]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [PubMed]
- Bradbury, P.J.; Zhang, Z.W.; Kroon, D.E.; Casstevens, T.M.; Ramdoss, Y.; Buckler, E.S. TASSEL: Software for association mapping of complex traits in diverse samples. Bioinformatics 2007, 23, 2633–2635. [Google Scholar] [CrossRef] [PubMed]
- Hardy, O.J.; Vekemans, X. SPAGEDi: A versatile computer program to analyse spatial genetic structure at the individual or population levels. Mol. Ecol. Notes 2002, 2, 618–620. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experiments | Traits | Minimum | Maximum | Average | CV (%) | Between Genotypes |
|---|---|---|---|---|---|---|
| Hydroponic | RSDW | 0.42 | 0.98 | 0.69 | 18.0 | ** |
| RRDW | 0.42 | 1.48 | 0.90 | 23.6 | ** | |
| RSFW | 0.26 | 0.96 | 0.57 | 23.3 | ** | |
| RRFW | 0.27 | 1.16 | 0.57 | 34.2 | ** | |
| RSH | 0.57 | 0.84 | 0.71 | 8.4 | ** | |
| RRL | 0.26 | 0.79 | 0.49 | 24.7 | ** | |
| RLATPase | 0.16 | 3.36 | 1.06 | 61.8 | ** | |
| RRATPase | 0.07 | 3.56 | 0.86 | 75.9 | ** | |
| RSK | 0.69 | 1.14 | 0.91 | 10.7 | ** | |
| RRK | 0.21 | 0.68 | 0.43 | 23.8 | ** | |
| Pot | RSDW | 0.20 | 1.14 | 0.66 | 29.9 | ** |
| RPOD | 0.30 | 4.46 | 2.18 | 36.6 | ** | |
| RCAT | 0.34 | 6.99 | 2.02 | 74.7 | ** | |
| RMDA | 0.20 | 4.51 | 1.48 | 71.6 | ** | |
| RPro | 0.71 | 6.37 | 2.28 | 60.7 | ** | |
| REL | 0.52 | 1.38 | 0.93 | 17.1 | ** | |
| RAL | 0.78 | 1.20 | 1.01 | 8.4 | ** | |
| RINL | 0.29 | 1.33 | 0.65 | 30.8 | ** | |
| RSK | 0.37 | 1.13 | 0.80 | 18.6 | ** | |
| RGW | 0.10 | 1.77 | 0.71 | 37.4 | ** |
| RSDW | RRDW | RSFW | RRFW | RLATPase | RRATPase | RSH | RRL | RSK | RRK | |
|---|---|---|---|---|---|---|---|---|---|---|
| RSDW | 1.000 | |||||||||
| RRDW | 0.479 ** | 1.000 | ||||||||
| RSFW | 0.896 ** | 0.375 ** | 1.000 | |||||||
| RRFW | 0.400 ** | 0.730 ** | 0.362 ** | 1.000 | ||||||
| RLATPase | 0.110 | 0.168 * | −0.033 | 0.142 | 1.000 | |||||
| RRATPase | 0.067 | 0.090 | 0.031 | −0.023 | 0.554 ** | 1.000 | ||||
| RSH | 0.440 ** | 0.189 * | 0.548 ** | 0.303 ** | −0.034 | 0.058 | 1.000 | |||
| RRL | 0.155 * | −0.059 | −0.095 | −0.200 | −0.280 ** | 0.019 | 0.006 | 1.000 | ||
| RSK | −0.409 ** | 0.029 | −0.078 | 0.072 | −0.117 | −0.077 | −0.093 | 0.085 | 1.000 | |
| RRK | 0.067 | 0.171 * | 0.185 * | 0.303 ** | 0.066 | 0.024 | 0.170 * | -0.007 | −0.010 | 1.000 |
| RSDW | RSK | RPOD | RCAT | RMDA | RPro | RAL | RINL | REL | RGW | |
|---|---|---|---|---|---|---|---|---|---|---|
| RSDW | 1.000 | |||||||||
| RSK | −0.112 | 1.000 | ||||||||
| RPOD | −0.012 | 0.210 ** | 1.000 | |||||||
| RCAT | 0.067 | 0.051 | −0.066 | 1.000 | ||||||
| RMDA | −0.073 | 0.259 ** | −0.075 | −0.032 | 1.000 | |||||
| RPro | 0.163 * | 0.126 | −0.133 | 0.410 ** | 0.384 ** | 1.000 | ||||
| RAL | 0.122 | −0.050 | −0.135 | −0.036 | 0.039 | 0.012 | 1.000 | |||
| RINL | 0.081 | −0.056 | −0.187 * | 0.005 | 0.157 * | 0.223 ** | 0.329 ** | 1.000 | ||
| REL | 0.118 | −0.058 | −0.255 ** | 0.027 | 0.129 | 0.153 | 0.515 ** | 0.551 ** | 1.000 | |
| RGW | 0.202 ** | −0.138 | −0.224 ** | −0.059 | 0.126 | 0.172 * | 0.396 ** | 0.289 ** | 0.554 ** | 1.000 |
| Traits | Marker | Chromosome | Distance (cM) | −log10 (p) | r2 (%) |
|---|---|---|---|---|---|
| RRDW | bPb-9005 | 1H | 67.88 | 4.72 | 13.0 |
| bPb-4481 | 2H | 18.81 | 4.37 | 13.1 | |
| bPb-0827 | 2H | 86.41 | 4.34 | 14.2 | |
| bPb-0775 | 2H | 140.87 | 4.34 | 13.0 | |
| bPb-2203 | 3H | 35.93 | 5.16 | 15.6 | |
| bPb-8013 | 4H | 86.69 | 3.37 | 13.3 | |
| bPb-6096 | 4H | 96.78 | 3.33 | 9.8 | |
| bPb-3491 | 7H | 100.50 | 4.44 | 9.9 | |
| bPb-6384 | 7H | 100.50 | 4.53 | 13.6 | |
| bPb-3506 | 7H | 101.20 | 4.53 | 13.6 | |
| bPb-8037 | 7H | 160.68 | 4.40 | 13.2 | |
| RSH | bPb-1127 | 1H | 57.23 | 3.35 | 9.9 |
| bPb-9336 | 3H | 100.76 | 3.48 | 10.3 | |
| bPb-8419 | 3H | 153.55 | 3.11 | 9.1 | |
| bPb-3375 | 6H | 122.08 | 3.57 | 10.5 | |
| RL | bPb-8935 | 1H | 118.95 | 4.14 | 12.3 |
| RRL | bPb-9908 | 7H | 111.69 | 3.77 | 8.9 |
| RSDW | bPb-3653 | 2H | 108.05 | 3.05 | 9.4 |
| AL | bPb-2976 | 1H | 54.01 | 4.11 | 12.9 |
| bPb-5334 | 1H | 67.88 | 4.25 | 13.4 | |
| bPb-5444 | 2H | 26.24 | 3.38 | 10.5 | |
| bPb-4184 | 2H | 119.90 | 3.95 | 12.4 | |
| bPb-1264 | 3H | 5.97 | 3.80 | 9.6 | |
| bPb-0068 | 3H | 66.50 | 6.09 | 19.8 | |
| bPb-6611 | 4H | 60.55 | 4.27 | 13.5 | |
| bPb-9820 | 4H | 142.09 | 4.16 | 13.1 | |
| bPb-1485 | 5H | 81.35 | 4.07 | 12.8 | |
| bPb-2835 | 5H | 81.35 | 3.56 | 11.1 | |
| bPb-8022 | 5H | 101.34 | 3.33 | 10.4 | |
| bPb-8553 | 5H | 120.44 | 4.09 | 12.9 | |
| bPb-4457 | 7H | 3.02 | 4.41 | 13.9 | |
| bPb-8051 | 7H | 78.22 | 3.25 | 10.1 | |
| INL | bPb-4184 | 2H | 119.90 | 3.15 | 9.7 |
| Traits | Marker | Chromosome | Distance (cM) | −log10 (p) | r2 (%) |
|---|---|---|---|---|---|
| RRATPase | bPb-3217 | 1H | 40.53 | 3.34 | 12.4 |
| bPb-6911 | 1H | 94.90 | 4.07 | 15.3 | |
| bPb-9121 | 1H | 94.90 | 4.08 | 15.3 | |
| bPb-4144 | 1H | 94.90 | 3.06 | 11.3 | |
| bPb-4877 | 2H | 47.38 | 3.03 | 11.2 | |
| bPb-8779 | 2H | 77.41 | 3.03 | 11.2 | |
| bPb-1593 | 2H | 149.44 | 3.11 | 11.5 | |
| bPb-8255 | 2H | 149.44 | 3.85 | 14.4 | |
| bPb-6884 | 3H | 0.98 | 4.09 | 15.3 | |
| bPb-3025 | 3H | 9.88 | 3.05 | 11.3 | |
| bPb-1137 | 3H | 10.20 | 6.00 | 23.2 | |
| bPb-2548 | 3H | 48.32 | 4.68 | 17.7 | |
| bPb-9299 | 6H | 14.35 | 5.44 | 20.8 | |
| bPb-3921 | 6H | 38.16 | 4.35 | 16.4 | |
| bPb-3722 | 6H | 68.53 | 5.83 | 22.5 | |
| bPb-7399 | 7H | 94.41 | 3.13 | 11.6 | |
| RLATPase | bPb-9957 | 1H | 63.32 | 3.37 | 8.1 |
| bPb-3921 | 6H | 38.16 | 3.37 | 10.1 | |
| bPb-4369 | 6H | 74.34 | 3.11 | 9.4 | |
| RATPase | bPb-8081 | 1H | 116.46 | 3.01 | 12.7 |
| bPb-6884 | 3H | 0.98 | 4.09 | 17.5 | |
| bPb-1137 | 3H | 10.20 | 4.46 | 19.2 | |
| bPb-9299 | 6H | 14.35 | 3.17 | 13.4 | |
| bPb-3722 | 6H | 68.53 | 4.35 | 18.7 | |
| bPb-6607 | 6H | 84.64 | 3.48 | 14.8 | |
| LATPase | bPb-8779 | 2H | 77.41 | 3.06 | 9.2 |
| RSK | bPb-3382 | 1H | 66.24 | 4.73 | 14.4 |
| bPb-0689 | 2H | 157.09 | 4.72 | 14.4 | |
| bPb-3045 | 4H | 65.34 | 4.42 | 13.4 | |
| bPb-9908 | 7H | 111.69 | 4.72 | 14.4 | |
| RRK | bPb-0910 | 1H | 59.42 | 3.10 | 9.1 |
| bPb-9957 | 1H | 63.32 | 3.25 | 7.6 | |
| SK | bPb-3382 | 1H | 66.24 | 3.22 | 9.6 |
| bPb-0689 | 2H | 157.09 | 3.25 | 9.7 | |
| bPb-3045 | 4H | 65.34 | 3.49 | 10.5 | |
| bPb-9908 | 7H | 111.69 | 3.29 | 9.8 | |
| MDA | bPb-7723 | 2H | 163.34 | 4.00 | 12.8 |
| POD | bPb-8737 | 2H | 108.72 | 3.07 | 9.5 |
| RPOD | bPb-5755 | 2H | 133.29 | 3.35 | 10.6 |
| RPro | bPb-9957 | 1H | 63.32 | 6.72 | 20.2 |
| bPb-5403 | 7H | 159.05 | 3.06 | 7.8 | |
| bPb-6701 | 7H | 159.19 | 3.06 | 7.8 | |
| SK (soil) | bPb-1447 | 7H | 78.22 | 3.21 | 9.4 |
| bPb-8112 | 1H | 140.85 | 3.14 | 9.2 | |
| RCAT | bPb-0994 | 2H | 113.25 | 3.43 | 9.3 |
| bPb-4601 | 2H | 157.00 | 5.69 | 16.1 | |
| bPb-1857 | 3H | 0.98 | 3.43 | 9.4 | |
| bPb-9599 | 3H | 149.85 | 3.40 | 9.3 | |
| bPb-0347 | 3H | 175.25 | 4.89 | 13.6 | |
| bPb-7069 | 3H | 178.60 | 8.66 | 25.7 | |
| bPb-7063 | 3H | 178.60 | 8.66 | 25.7 | |
| bPb-8580 | 5H | 8.49 | 4.98 | 13.9 | |
| bPb-3792 | 5H | 45.58 | 5.07 | 14.2 | |
| bPb-5584 | 5H | 54.10 | 5.28 | 14.8 | |
| bPb-2304 | 6H | 136.06 | 8.89 | 26.4 | |
| bPb-7146 | 6H | 137.76 | 4.40 | 12.2 | |
| bPb-1621 | 6H | 137.76 | 5.17 | 14.5 | |
| bPb-0783 | 7H | 160.68 | 4.71 | 13.1 |
| Trait | Marker | Chromosome | cM | Gene | cM | Description |
|---|---|---|---|---|---|---|
| RRDW | bPb-9005 | 1H | 54.39 | MLOC_59277 | 52.51 | Lateral root primordium (LRP) protein-related |
| bPb-3491 | 7H | 89.14 | MLOC_74469 | 89.14 | Lateral root primordium (LRP) protein-related | |
| RRATPase | bpb-4144 | 1H | 87.87 | MLOC_13794 | 90.3 | ATP-binding cassette transporter |
| MLOC_58493 | 90.3 | ABC (ATP-binding) family transporter | ||||
| bpb-3722 | 6H | 54.89 | MLOC_68594 | 52.83 | Potassium transporter | |
| MLOC_46910 | 53.6 | AAA-type ATPase family protein | ||||
| MLOC_57716 | 53.61 | ATPase 7B; Copper-transporting ATPase 2 | ||||
| MLOC_10111 | 53.9 | Potassium channel | ||||
| MLOC_72084 | 54.82 | ADP, ATP carrier protein 1 | ||||
| MLOC_34488 | 54.89 | ABC transporter ATP-binding protein | ||||
| MLOC_5994 | 55.03 | ATP-binding cassette transporter | ||||
| bpb-7399 | 7H | 84.56 | MLOC_52035 | 84.57 | ATP-binding cassette transporter | |
| MLOC_13204 | 89.14 | Potassium transporter | ||||
| RRK RLATPase | bpb-9957 | 1H | 47.83 | MLOC_37216 | 46.6 | ABC transporter ATP-binding protein |
| MLOC_18422 | 47.83 | Phospholipid-transporting ATPase 3 | ||||
| RRK | bPb-0910 | 1H | 46.81 | AK366430 | 48.09 | K Homology type 1 |
| MLOC_5191.1 | 52.48 | K Homology type 1 subgroup | ||||
| POD | bPb-8737 | 2H | 98.52 | MLOC_25978 | 98.52 | Peroxidase 60, putative |
| RPOD | bPb-5755 | 2H | 119.76 | MLOC_66485 | 119.33 | Peroxidase 16 |
| MLOC_15491 | 124.77 | Peroxidase | ||||
| RCAT | bPb-4601 | 2H | 144.12 | MLOC_65477 | 146.53 | Peroxidase 10 |
| MLOC_54893 | 146.92 | Peroxidase 12 | ||||
| bPb-2304 | 6H | 117.55 | MLOC_55199 | 117.55 | Peroxidase superfamily protein | |
| RPro | bPb-5403 | 7H | 138.22 | MLOC_78556 | 140.86 | High mobility group protein |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, M.; Fu, M.-M.; Qiu, C.-W.; Cao, F.; Chen, Z.-H.; Zhang, G.; Wu, F. Response of Tibetan Wild Barley Genotypes to Drought Stress and Identification of Quantitative Trait Loci by Genome-Wide Association Analysis. Int. J. Mol. Sci. 2019, 20, 791. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20030791
Zhang M, Fu M-M, Qiu C-W, Cao F, Chen Z-H, Zhang G, Wu F. Response of Tibetan Wild Barley Genotypes to Drought Stress and Identification of Quantitative Trait Loci by Genome-Wide Association Analysis. International Journal of Molecular Sciences. 2019; 20(3):791. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20030791
Chicago/Turabian StyleZhang, Mian, Man-Man Fu, Cheng-Wei Qiu, Fangbin Cao, Zhong-Hua Chen, Guoping Zhang, and Feibo Wu. 2019. "Response of Tibetan Wild Barley Genotypes to Drought Stress and Identification of Quantitative Trait Loci by Genome-Wide Association Analysis" International Journal of Molecular Sciences 20, no. 3: 791. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20030791