Molecular Identification and Disease Management of Date Palm Sudden Decline Syndrome in the United Arab Emirates

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
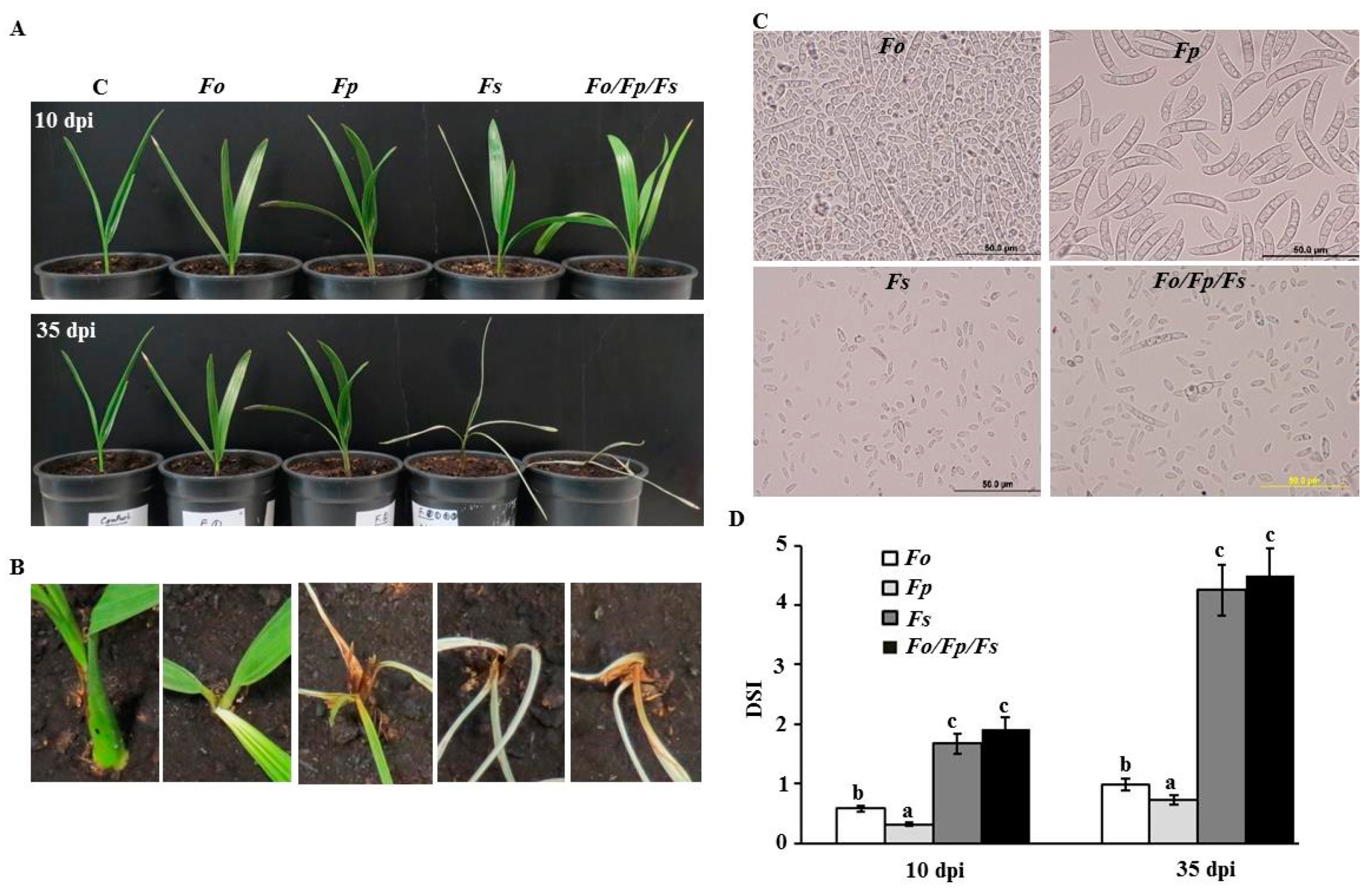
2.1. Symptoms of SDS on Date Palm
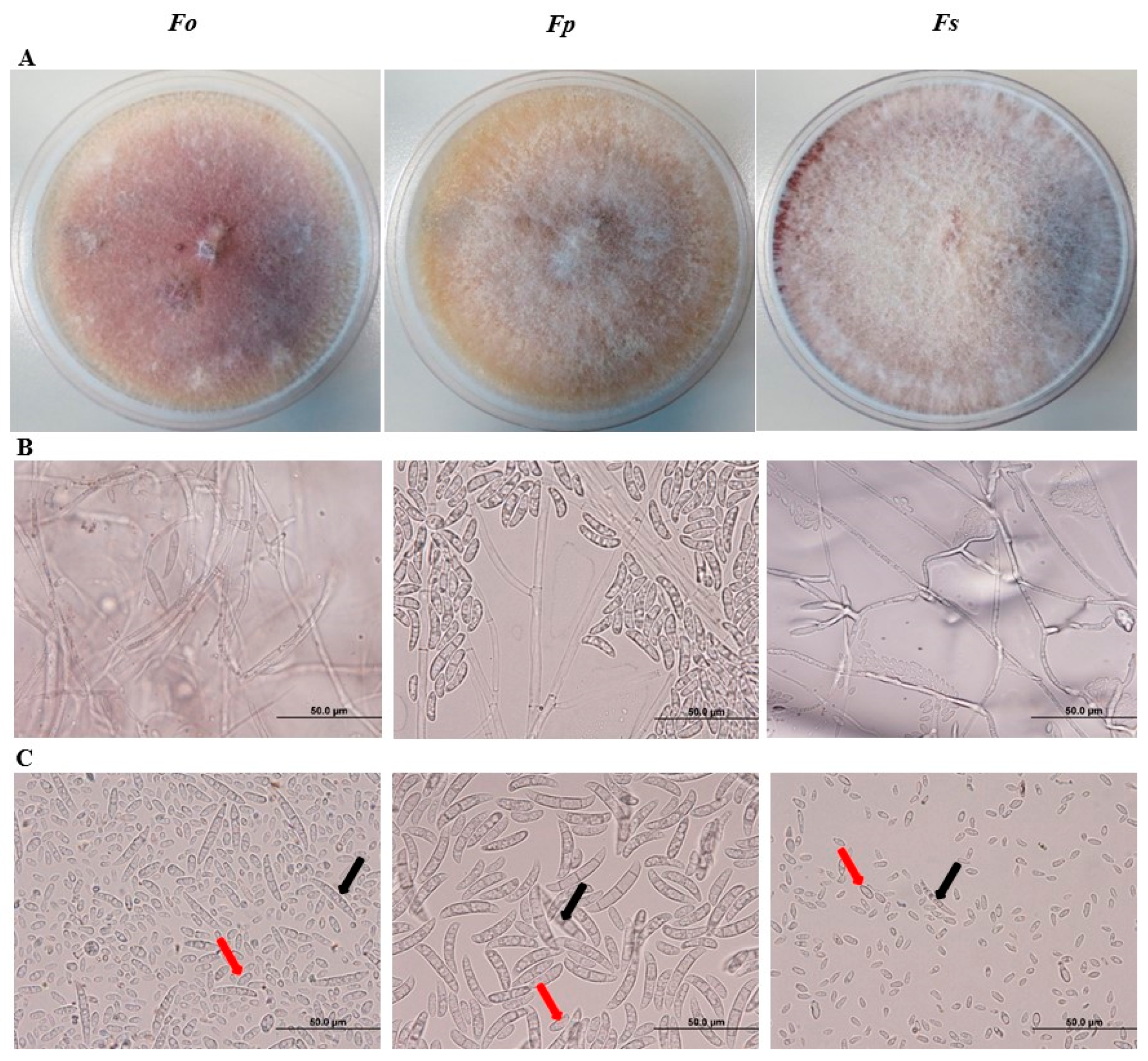
2.2. Cultural and Morphological Identification of Fusaium spp. Associated with SDS
2.3. Molecular and Phylogenetic Classification of Fusarium spp.
2.4. Pathogenicity Tests of Fusarium spp. on Date Palm Seedlings
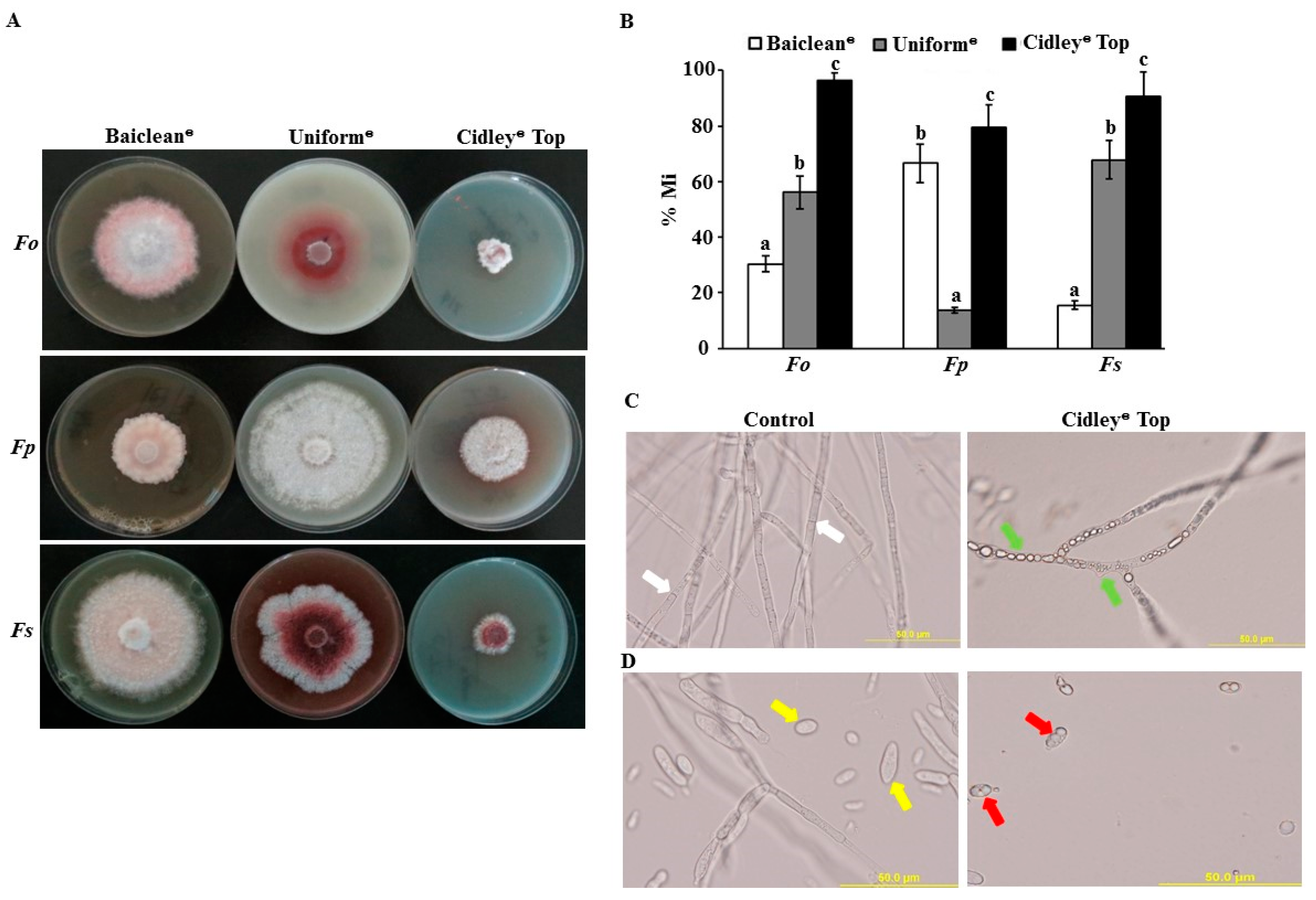
2.5. In Vitro Evaluation of Selected Fungicides against Fusarium spp.
2.6. Effect of Cidely® Top on Date Palm Seedlings Infected with F. solani
3. Discussion
4. Materials and Methods
4.1. Isolation, Purification, and Culture Maintenance
4.2. DNA Isolation, PCR, and Sequencing
4.3. Phylogenetic Analysis
4.4. Disease Assays and Pathogenicity Tests
4.5. Evaluation of Fungicides against F. solani
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Hamad, I.; AbdElgawad, H.; Al Jaouni, S.; Zinta, G.; Asard, H.; Hassan, S.; Hegab, M.; Hagagy, N.; Selim, S. Metabolic analysis of various date palm fruit (Phoenix dactylifera L.) cultivars from Saudi Arabia to assess their nutritional quality. Molecules 2015, 20, 13620–13641. [Google Scholar] [CrossRef] [PubMed]
- Food and Agriculture Organization of the United Nations (FAO). Date Palm Production; FAOSTAT Database; FAO: Rome, Italy, 2016. [Google Scholar]
- Abdullah, S.K.; Lopez Lorca, L.V.; Jansson, H.B. Diseases of date palms (Phoenix dactylifera L.). Basrah J. Date Palm Res. 2010, 9, 1–44. [Google Scholar]
- Saeed, E.E.; Sham, A.; El-Tarabily, K.A.; Abu Elsamen, F.; Iratni, R.; AbuQamar, S.F. Chemical control of dieback disease on date palm caused by the fungal pathogen, Thielaviopsis punctulata, in United Arab Emirates. Plant Dis. 2016, 100, 2370–2376. [Google Scholar] [CrossRef] [PubMed]
- Alhammadi, M.S.; Al-Shariqi, R.; Maharachchikumbura, S.; Al-Sadi, A.M. Molecular identification of fungal pathogens associated with date palm root diseases in the United Arab Emirates. J. Plant Pathol. 2018, 99, 1–7. [Google Scholar]
- Abdalla, M.Y.; AI-Rokibah, A.; Moretti, A.; Mule, G. Pathogenicity of toxigenic Fusarium proliferatum from date palm in Saudi Arabia. Plant Dis. 2000, 84, 321–324. [Google Scholar] [CrossRef]
- Armengol, J.; Moretti, A.; Perrone, G.; Vicent, A.; Bengoechea, J.A.; García-Jiménez, J. Identification, incidence and characterization of Fusarium proliferatum on ornamental palms in Spain. Eur. J. Plant Pathol. 2005, 112, 123–131. [Google Scholar] [CrossRef]
- Sarhan, A.R.T. A study on the fungi causing decline of date palm trees in middle of Iraq. In Proceedings of the Second International Conference on Date Palm, Al Ain, UAE, 25–27 March 2001; pp. 424–430. [Google Scholar]
- Mansoori, B.; Kord, H. Yellow death: A disease of date palm in Iran caused by Fusarium solani. J. Phytopathol. 2006, 154, 125–127. [Google Scholar] [CrossRef]
- Maitlo, W.A.; Markhand, G.S.; Abul-Soad, A.A.; Lodhi, A.M.; Jatoi, M.A. Chemcial control of sudden decline disease of date palm (Phoenix dactylifera L.) in Sindh, Pakistan. Pak. J. Bot. 2013, 45, 7–11. [Google Scholar]
- Sabet, K.K.; Ghanem, G.A.; Rashed, M.F.; Allam, L.A. Fungal infection of date palm tissue culture. J. Agric. Sci. Mansoura Univ. 2006, 31, 735–745. [Google Scholar]
- El Hassni, M.; El Hadrami, A.; Daayf, F.; Chérif, M.; Barka, E.; El Hadrami, I. Biological control of bayoud disease in date palm: Selection of microorganisms inhibiting the causal agent and inducing defense reactions. Environ. Exp. Bot. 2007, 59, 224–234. [Google Scholar] [CrossRef]
- Tantaoui, A.; Ouinten, M.; Geiger, J.-P.; Fernandez, D. Characterization of a single clonal lineage of Fusarium oxysporum f. sp. albedinis causing Bayoud disease of date palm in Morocco. Phytopathology 1996, 86, 787–792. [Google Scholar]
- Shabani, F.; Kumar, L. Risk levels of invasive Fusarium oxysporum f. sp. in areas suitable for date palm (Phoenix dactylifera) cultivation under various climate change projections. PLoS ONE 2013, 8, e83404. [Google Scholar] [CrossRef]
- Okungbowa, F.I.; Shittu, H.O. Fusarium wilts: An overview. Environ. Res. J. 2012, 6, 83–102. [Google Scholar]
- Saremi, H.; Okhovvat, S.M.; Ashrafi, S.J. Wilting of date palm branches by Fusarium oxysporum in south of Iran and its control managements with soil solarization method. Commun. Agric. Appl. Biol. Sci. 2007, 72, 831–837. [Google Scholar]
- Bokhary, H.A. Seed-borne fungi of date-palm, Phoenix dactylifera L. from Saudi Arabia. Saudi J. Biol. Sci. 2010, 17, 327–329. [Google Scholar] [CrossRef]
- Maitlo, W.A.; Markhand, G.S.; Abul-Soad, A.A.; Lodhi, A.M.; Jatoi, M.A. Fungi associated with sudden decline disease of date palm (Phoenix dactylifera L.) and its incidence at Khairpur, Pakistan. Pak. J. Bot. 2016, 26, 65–71. [Google Scholar]
- Abul-Soad, A.A.; Mahdi, S.M.; Markhand, G.S. Date Palm Status and Perspective in Pakistan. In Date Palm Genetic Resources and Utilization; Al-Khayri, J., Jain, S., Johnson, D., Eds.; Springer: Dordrecht, The Netherlands, 2015; pp. 153–205. [Google Scholar]
- El Kinany, S.; Faggroud, M.; Ouahmane, L.; El Hilali, R.; Haggoud, A.; Bouamri, R. Effect of organic fertilizer and commercial arbuscular mycorrhizal fungi on the growth of micropropagated date palm cv. Feggouss. J. Saudi Soc. Agric. Sci. 2018. [Google Scholar] [CrossRef]
- Salim, H.A.; Hassan, K.A.; Ishak, H.S.; Hussein, A.A.; Gab, A. Control of wilt disease (Sudden Decline Syndrome) on date palms in Iraq. Am. Multidis. Int. Res. J. 2015, 2, 29–33. [Google Scholar]
- Abul-Soad, A.A.; Maitlo, W.A.; Markhand, G.S.; Mahdi, S.M. Date palm wilt disease (sudden decline syndrome) in Pakistan, symptoms and remedy. Blessed Tree 2011, 3, 38–43. [Google Scholar]
- Razdan, V.; Sabitha, M. Integrated disease management: Concepts and practices. In Integrated Pest Management: Innovation-Development Process; Peshin, R., Dhawan, A.K., Eds.; Springer: Dordrecht, The Netherlands, 2009; pp. 369–389. [Google Scholar]
- Patel, P.N.; Prasad, N.; Mathur, R.L.; Mathur, B.L. Fusarium wilt of cumin. Curr. Sci. 1957, 26, 181–182. [Google Scholar]
- Nirenberg, H. Unterscuchungen ϋber die morphologische and biologische. Differenzierung in der Fusarium-Section Liseola. Mitteilungen der Biologischen Budesanstalt fϋr Land- und Forstwirtschaft 1976, 169, 1–117. [Google Scholar]
- Saccardo, P.A. Fungi Veneti novi vel critici v. mycologiae Veneti addendi. Series XII. Michelia 1881, 2, 241–301. [Google Scholar]
- Alves, A.; Crous, P.W.; Correia, A.; Phillips, A.J.L. Morphological and molecular data reveal cryptic speciation in Lasiodiplodia theobromae. Fungal Divers. 2008, 28, 1–13. [Google Scholar]
- Fernandez, D.; Ouinten, M.; Tantaoui, A.; Geiger, J.P.; Daboussi, M.J.; Langin, T. Fot 1 insertions in the Fusarium oxysporum f. sp. albedinis genome provide diagnostic PCR targets for detection of the date palm pathogen. Appl. Environ. Microbiol. 1998, 64, 633–636. [Google Scholar]
- Watanabe, M. Molecular phylogeny and identification of Fusarium species based on nucleotide sequences. Mycotoxins 2013, 63, 133–142. [Google Scholar] [CrossRef]
- Saleh, A.A.; Sharafaddin, A.H.; El-Komy, M.H.; Ibrahim, Y.E.; Hamad, Y.K.; Molan, Y.Y. Fusarium species associated with date palm in Saudi Arabia. Eur. J. Plant Pathol. 2017, 148, 367–377. [Google Scholar] [CrossRef]
- O’Callaghan, M. Microbial inoculation of seed for improved crop performance: Issues and opportunities. Appl. Microbiol. Biotechnol. 2016, 100, 5729–5746. [Google Scholar] [CrossRef]
- Zaid, A.; de Wet, P.F.; Djerbi, M.; Oihabi, A. Chapter XII: Diseases and pests of date palm. In Date Palm Cultivation; Zaid, A., Ed.; Plant Production and Protection Paper No. 156; FAO: Rome, Italy, 2002. [Google Scholar]
- Saeed, E.E.; Sham, A.; AbuZarqa, A.; Al Shurafa, K.; Al Naqbi, T.S.; Iratni, R.; El-Tarabily, K.A.; AbuQamar, S.F. Detection and management of mango dieback disease in the United Arab Emirates. Int. J. Mol. Sci. 2017, 18, 2086. [Google Scholar] [CrossRef]
- El-Deeb, H.M.; Lashin, S.M.; Arab, Y.A. Distribution and pathogenesis of date palm fungi in Egypt. Acta Hortic. 2007, 736, 421–429. [Google Scholar] [CrossRef]
- Abdullah, S.K.; Asensio, L.; Monfort, E.; Gomez-Vidal, S.; Salinas, J.; Lopez Lorca, L.V.; Jansson, H.B. Incidence of the two date palm pathogens, Thielaviopsis paradoxa and T. punctulata in soil from date palm plantations in Elx, south-east Spain. J. Plant Prot. Res. 2009, 49, 276–279. [Google Scholar]
- Hameed, M.A. Inflorescence rot disease of date palm caused by Fusarium proliferatum in Southern Iraq. Afr. J. Biotechnol. 2012, 11, 8616–8621. [Google Scholar]
- Rashed, M.F.; Abdel Hafeez, N.E. Decline of date palm trees in Egypt. In Proceedings of the Second International Conference on Date Palm, Al Ain, UAE, 25–27 March 2001; pp. 401–407. [Google Scholar]
- Killian, C.; Maire, R. Le bayoud maladie du dattier. Bull. Soc. Hist. Na. Afr. 1930, 21, 89–101. [Google Scholar]
- Djerbi, M. Bayoud disease in North Africa, history, distribution, diagnosis and control. Date Palm J. 1982, 1, 153–197. [Google Scholar]
- Nawade, B.; Talaviya, J.R.; Vyas, U.M.; Jadeja, K.B.; Golakiya, B.A. Diversity analysis among Fusarium oxysporum F. sp. cumini isolates using ISSR markers, spore morphology and pathogenicity. Int. J. Curr. Microbiol. App. Sci. 2017, 6, 79–87. [Google Scholar]
- Adame-García, G.; Rodríguez-Guerra, R.; Iglesias-Andreu, L.G.; Ramos-Prado, J.M.; Luna-Rodríguez, M. Molecular identification and pathogenic variation of Fusarium species isolated from Vanilla planifolia in Papantla Mexico. Bot. Sci. 2015, 93, 669–678. [Google Scholar] [CrossRef]
- Abd-Elsalam, K.A.; Youssef, K.; Almoammar, H. First morphogenetic identification of Fusarium solani isolated from orange fruit in Egypt. FYTON 2015, 84, 128–131. [Google Scholar]
- Lemanceau, P.; Bakker, P.A.H.M.; De Kogel, W.J.; Alabouvette, C.; Schippers, B. Antagonistic effect of nonpathogenic Fusarium oxysporum Fo47 and Pseudobactin 358 upon pathogenic Fusarium oxysporum f. sp. dianthi. Appl. Environ. Microbiol. 1993, 59, 74–82. [Google Scholar]
- Lamichhane, J.R.; Venturi, V. Synergisms between microbial pathogens in plant disease complexes: A growing trend. Front. Plant Sci. 2015, 6, 385. [Google Scholar] [CrossRef]
- Kuzdraliński, A.; Nowak, M.; Szczerba, H.; Dudziak, K.; Muszyńska, M.; Leśniowska-Nowak, J. The composition of Fusarium species in wheat husks and grains in south-eastern Poland. J. Integr. Agric. 2017, 16, 60345–60347. [Google Scholar] [CrossRef]
- Landschoot, S.; Waegeman, W.; Audenaert, K.; Vandepitte, J.; Baetens, J.M.; De Baets, B.; Haesaert, G. An empirical analysis of explanatory variables affecting Fusarium head blight infection and deoxynivalenol content in wheat. J. Plant Pathol. 2012, 94, 135–147. [Google Scholar]
- Saeed, E.E.; Sham, A.; Salmin, Z.; Abdelmowla, Y.; Iratni, R.; El-Tarabily, K.A.; AbuQamar, S.F. Streptomyces globosus UAE1, a potential effective biocontrol agent for black scorch disease in date palm plantations. Front. Microbiol. 2017, 8, 1455. [Google Scholar] [CrossRef]
- Iqbal, Z.; Pervez, M.A.; Ahmad, S.; Iftikhar, Y.; Yasin, M.; Nawaz, A.; Ghazanfar, M.U.; Dasti, A.A.; Saleem, A. Determination of minimum inhibitory concentrations of fungicides against fungus Fusarium mangiferae. Pak. J. Bot. 2010, 42, 3525–3532. [Google Scholar]
- Bhanumathi, A.; Ravishankar, R.V. Leaf blight of Azadirachta indica and its in vitro management. Afr. J. Agric. Res. 2007, 2, 53–543. [Google Scholar]
- Aktar, M.W.; Sengupta, D.; Chowdhury, A. Impact of pesticides use in agriculture: Their benefits and hazards. Interdiscip. Toxicol. 2009, 2, 1–12. [Google Scholar] [CrossRef]
- Ganesan, K.; Raza, S.K.; Vijayaraghavan, R. Chemical warfare agents. J. Pharm. Bioallied Sci. 2010, 2, 166–178. [Google Scholar] [CrossRef]
- Kamil, F.H.; Saeed, E.E.; El-Tarabily, K.A.; AbuQamar, S.F. Biological control of mango dieback disease caused by Lasiodiplodia theobromae using streptomycete and non-streptomycete actinobacteria in the United Arab Emirates. Front Microbiol. 2018, 9, 829. [Google Scholar] [CrossRef]
- AbuQamar, S.F.; Moustafa, K.; Tran, L.S. Mechanisms and strategies of plant defense against Botrytis cinerea. Crit. Rev. Biotechnol. 2017, 37, 263–275. [Google Scholar] [CrossRef]
- Weems, J.D.; Haudenshield, J.S.; Bond, J.P.; Hartman, G.L.; Ames, K.A.; Bradley, C.A. Effect of fungicide seed treatments on Fusarium virguliforme infection of soybean and development of sudden death syndrome. Can. J. Plant Pathol. 2015, 37, 435–447. [Google Scholar] [CrossRef]
- Kirsop, B.E.; Doyle, A. Maintenance of Microorganisms and Cultured Cells, a Manual of Laboratory Methods, 2nd ed.; Academic Press: London, UK, 1991. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplificatin and direstc sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc. 1990, 91, 315–322. [Google Scholar]
- Schoch, C.L.; Seifert, K.A.; Huhndorf, S.; Robert, V.; Spouge, J.L.; Levesque, C.A.; Chen, W. Fungal Barcoding Consortium. Nuclear ribosomal internal transcribed spacer (ITS) region as a universal DNA barcode marker for Fungi. Proc. Natl. Acad. Sci. USA 2012, 109, 6241–6246. [Google Scholar] [CrossRef]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–555. [Google Scholar] [CrossRef]
- Orbach, M.J.; Porro, E.B.; Yanofsky, C. Cloning and characterization of the gene for β-tubulin from a benomyl-resistant mutant of Neurospora crassa and its use as a dominant selectable marker. Mol. Cell. Biol. 1986, 6, 2452–2461. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef]
- Elhassan, A.M. Isolation, identification and characterization of Fusarium oxysporum, the causal agent of Fusarium wilt disease of date palm Phoeniex dactylifera L. in Northern State Sudan. Int. J. Curr. Microbiol. App. Sci. 2016, 5, 381–386. [Google Scholar] [CrossRef]
- Molan, Y.Y.; Al-Obeed, R.S.; Harhash, M.M.; El-Husseini, S. Decline of date-palm offshoots with Chalara paradoxa in Riyadh region. J. King Saud Univ. 2004, 16, 79–86. [Google Scholar]
- SAS Institute. The SAS System for Windows; Release 9.0; SAS Institute: Cary, NC, USA, 2002. [Google Scholar]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alwahshi, K.J.; Saeed, E.E.; Sham, A.; Alblooshi, A.A.; Alblooshi, M.M.; El-Tarabily, K.A.; AbuQamar, S.F. Molecular Identification and Disease Management of Date Palm Sudden Decline Syndrome in the United Arab Emirates. Int. J. Mol. Sci. 2019, 20, 923. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20040923
Alwahshi KJ, Saeed EE, Sham A, Alblooshi AA, Alblooshi MM, El-Tarabily KA, AbuQamar SF. Molecular Identification and Disease Management of Date Palm Sudden Decline Syndrome in the United Arab Emirates. International Journal of Molecular Sciences. 2019; 20(4):923. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20040923
Chicago/Turabian StyleAlwahshi, Khawla J., Esam Eldin Saeed, Arjun Sham, Aisha A. Alblooshi, Marwa M. Alblooshi, Khaled A. El-Tarabily, and Synan F. AbuQamar. 2019. "Molecular Identification and Disease Management of Date Palm Sudden Decline Syndrome in the United Arab Emirates" International Journal of Molecular Sciences 20, no. 4: 923. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20040923