IRES Trans-Acting Factors, Key Actors of the Stress Response

 and
and
Abstract
:1. Introduction
2. The Viral IRESs
3. The Cellular IRESs
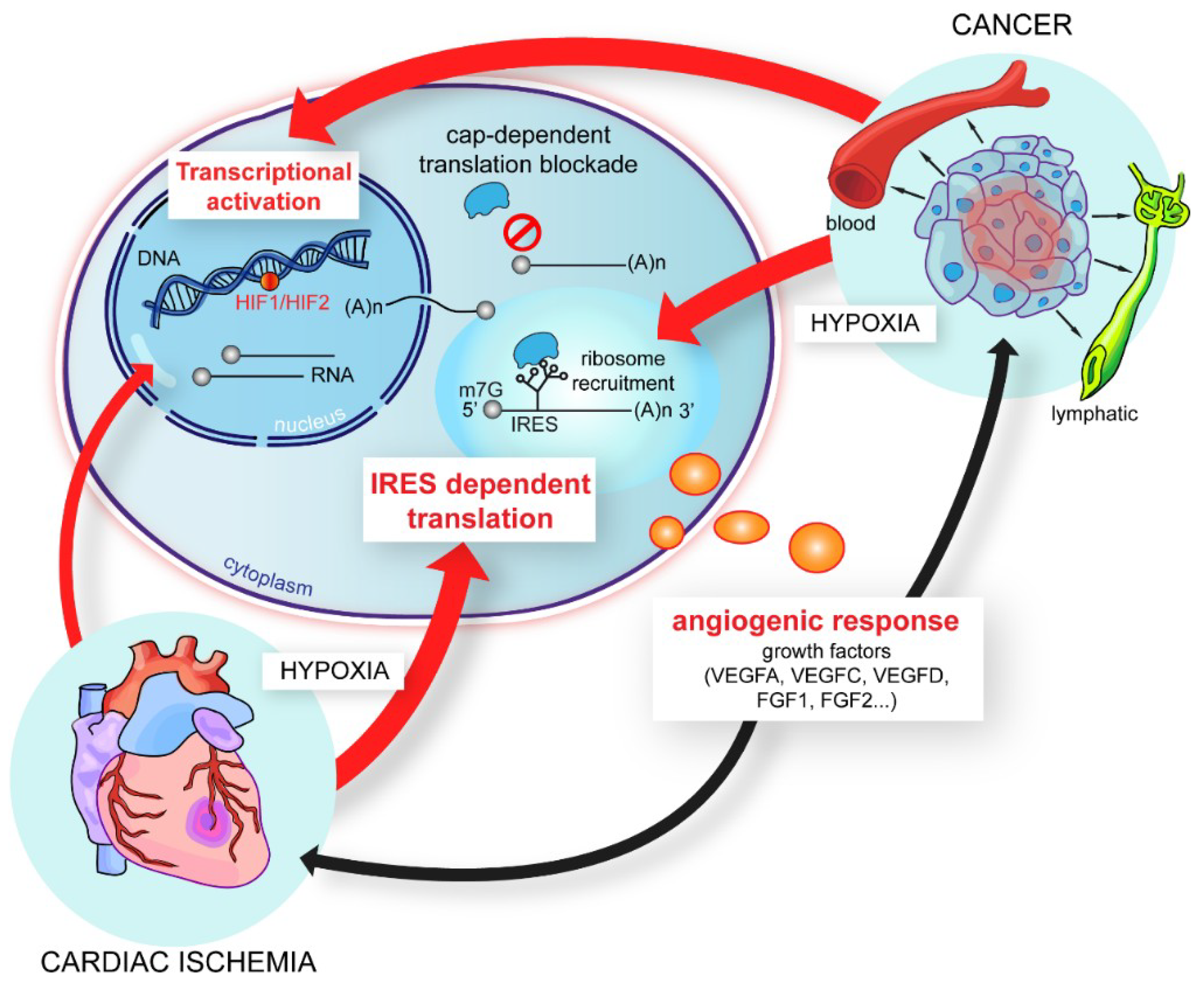
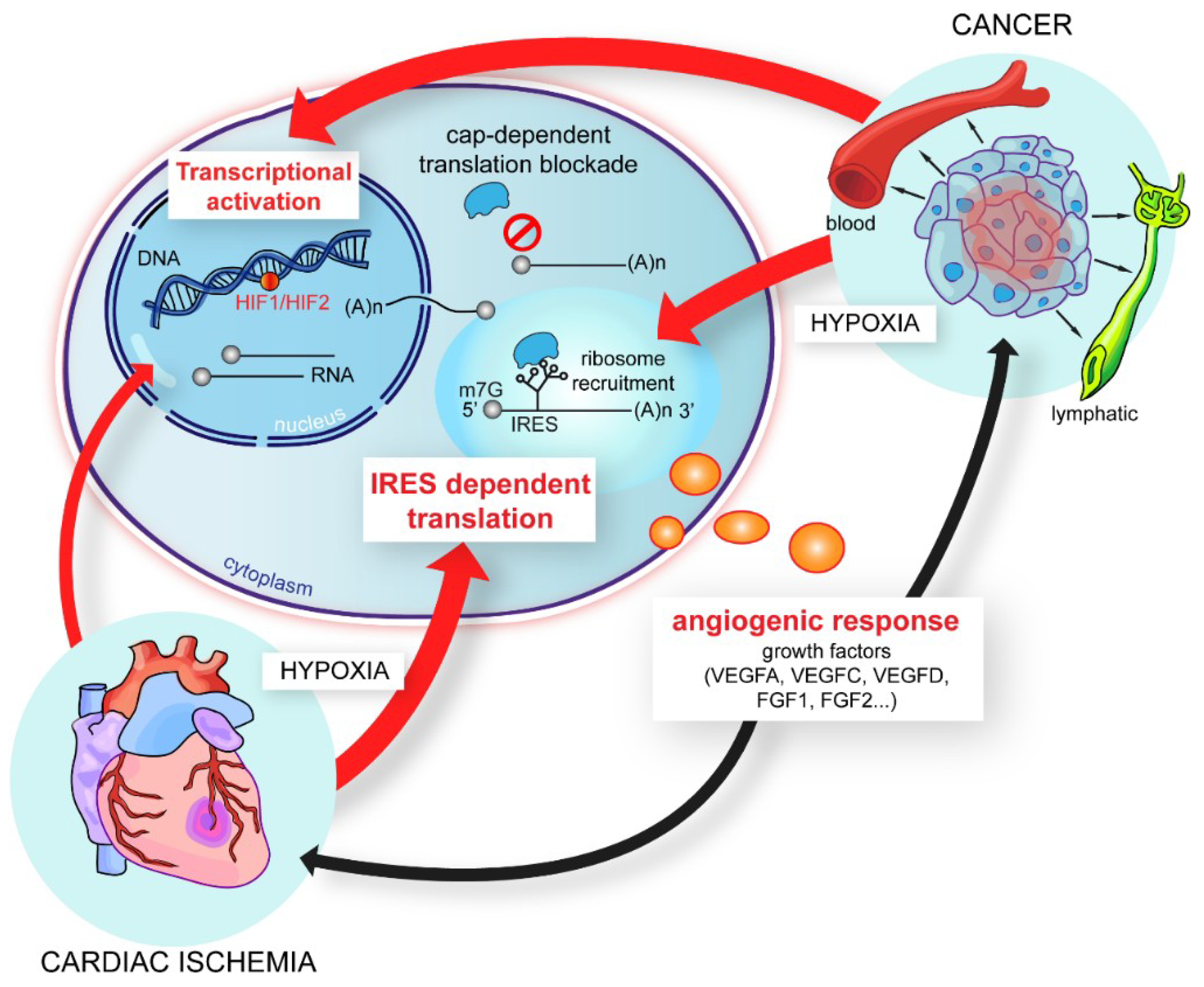
4. IRES-Dependent Translation, a Pivotal Mechanism in the Stress Response
5. IRES Trans-Acting Factors, Key Regulators of Cellular IRESs
6. Multifunctional ITAFs: How are They Assigned to the Translational Function?
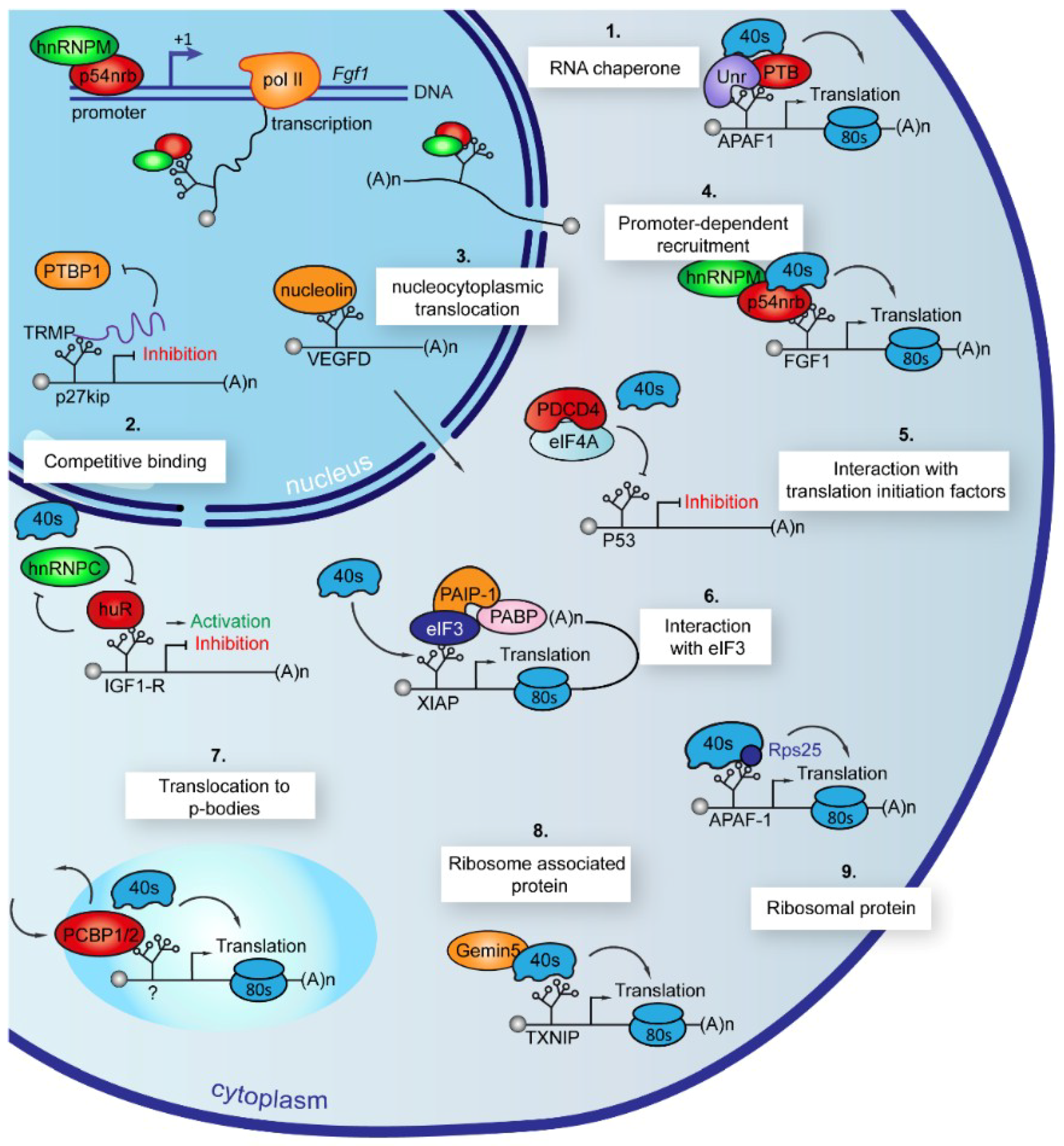
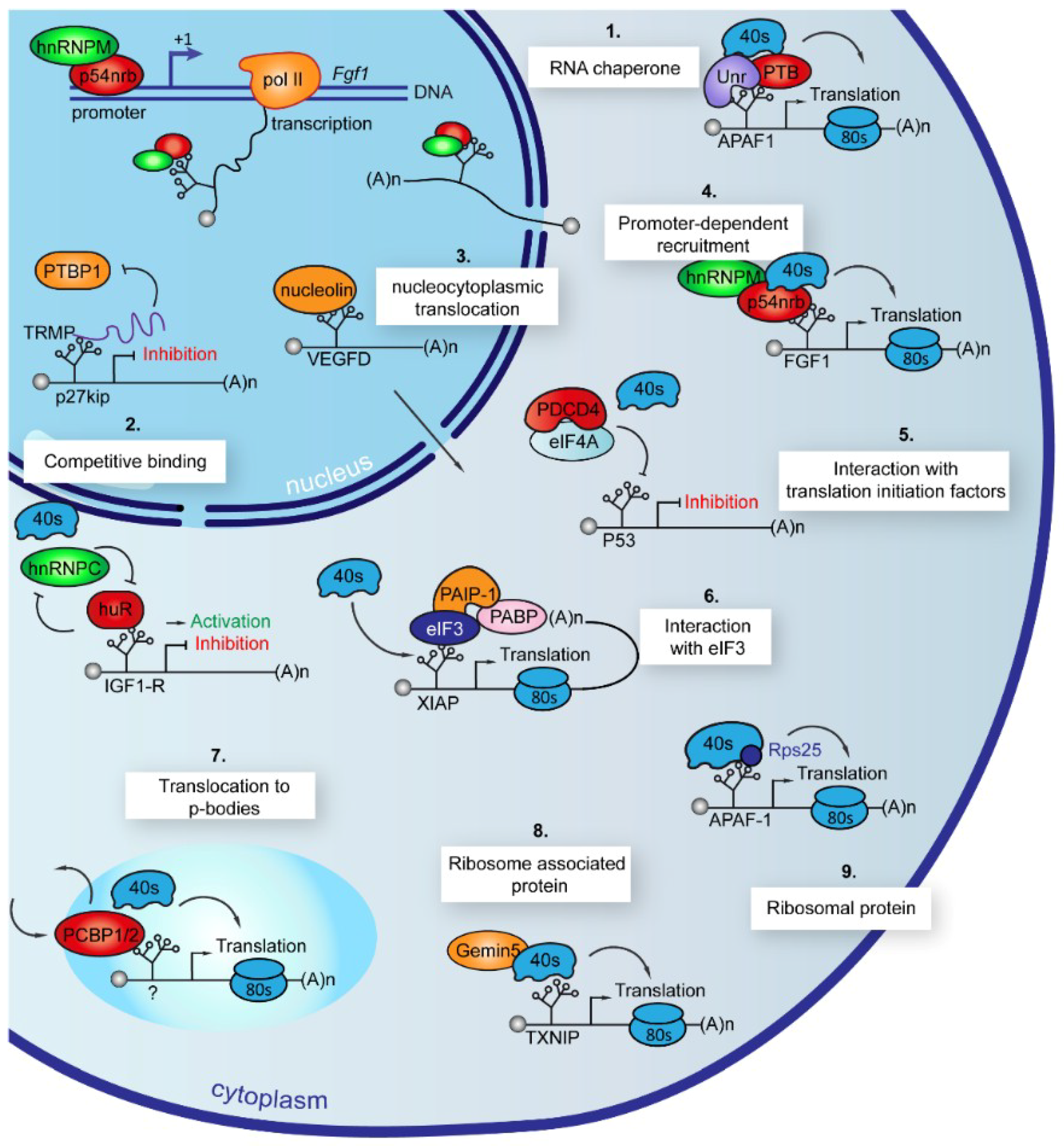
7. ITAFs Use Different Mechanisms of Action to Control IRES-Dependent Translation
7.1. Chaperones
7.2. Competitive Binding
7.3. Nucleocytoplasmic Translocation
7.4. Promoter-Dependent Recruitment
7.5. Interaction with Translation Initiation Factors or with 4E-BP
7.6. ITAF Role of eIFs, eEFs and 4E-BP
7.7. Translocation to Stress Granules and P-Bodies
7.8. Association to Ribosome
7.9. Ribosome Inherent Constituent
8. Discussion
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| 4E-BP | 4E binding protein |
| AANAT | Arylalkylamine N-acyltransferase |
| ADAR1 | Adenosine deaminase RNA specific 1 |
| APAF1 | Apoptotic peptidase activating factor1 |
| APP | Amyloid precursor protein |
| ATF4 | Activating transcription factor 4 |
| BAG1 | Bcl-2-associated athanogene 1 |
| BiP | Immunoglobulin heavy-chain binding protein |
| CDK1 | Cyclin-dependent kinase 1 |
| Chmp2A | Charged multivesicular body protein 2A |
| CrPV | Cricket paralysis virus |
| Cryo-EM | Cryogenic electron microscopy |
| CSDE1 | Cold shock domain containing E1 |
| CSFV | Classical swine fever virus |
| CUGBP1 | CUG triplet repeat RNA-binding protein 1 |
| DAP5 | Death-associated protein 5 |
| DCV | Drosophila C virus |
| eIF | Eukaryotic initiation factor |
| ELAV | Embryonic lethal abnormal vision |
| ELAV1 | ELAV-like protein 1 |
| ELG | Elongatus |
| EMCV | Encephalomyocarditis virus |
| ER | Endoplasmic reticulum |
| ER-α | Estrogen receptor alpha |
| FBP3 | Far-upstream element binding protein 3 |
| FGF | Fibroblast growth factor |
| FMDV | Foot-and-mouth disease virus |
| FMRP | Fragile X mental retardation protein |
| FUS | Fused in sarcoma |
| G3BP1 | Ras GTPase SH3 stress granule assembly factor 1 |
| GCN2 | General control non-derepressible 2 |
| GCN4 | General control non-derepressible 4 |
| GRSF1 | G-rich RNA sequence binding factor 1 |
| HCV | Hepatitis C virus |
| HIAP2 | Human inhibitor of apoptosis 2 |
| HIF | Hypoxia-inducible factor |
| hnRNP | Heterogeneous nuclear ribonucleoprotein |
| Hox | Homeobox |
| HRI | Haem-regulated inhibitor kinase |
| IGF1R | Insulin growth factor-like receptor |
| IGR | Intergenic region |
| IR | Insulin receptor |
| IRES | Internal ribosome entry site |
| IRF2 | Interferon regulatory factor 2 |
| ITAF | IRES trans-acting factor |
| LEF1 | Lymphoid enhancer binding factor 1 |
| LucR | Renilla luciferase |
| mTOR | Mechanistic target of rapamycin |
| NAT-1 | N-acetyltransferase 1 |
| NRF | NFKB repressing factor |
| NSAP | Nephritis strain-associated protein |
| P-bodies | Processing bodies |
| PABP | PolyA-binding protein |
| PCBP | poly r(C) binding protein |
| PDCD4 | Programmed cell death protein 4 |
| PERK | PKR-like endoplasmic reticulum kinase |
| PINK1 | PTEN-induced putative kinase-1 |
| PKR | Protein kinase RNA |
| PRMT5 | Protein arginine methyltransferase |
| PSF | PTB-associated splicing factor |
| PTB | Pyrimidine tract binding protein |
| PV | Poliovirus |
| RBM4 | RNA-binding motif protein 4 |
| RHA | RNA helicase A |
| RRBP1 | Ribosome binding protein 1 |
| SHMT-1 | Serine hydroxymethyltransferase 1 |
| Smac | Second mitochondrial activator of caspase |
| SMN | Survival of motor neurone |
| SREBP-1 | Sterol regulatory element-binding protein 1 |
| TCP80 | Translational control protein 80 |
| TRMP | TP53-regulated modulator of p27 |
| TXNIP | Thioredoxin-interacting protein |
| Unr | Upstream of N-ras |
| uORF | Upstream open reading frame |
| UTR | Untranslated region |
| VASH-1 | Vasohibin 1 |
| VEGF | Vascular endothelial growth factor |
| XIAP | X-linked inhibitor of apoptosis |
References
- Kozak, M. Inability of circular mRNA to attach to eukaryotic ribosomes. Nature 1979, 280, 82–85. [Google Scholar] [CrossRef] [PubMed]
- Kozak, M. The scanning model for translation: An update. J. Cell Biol. 1989, 108, 229–241. [Google Scholar] [CrossRef] [PubMed]
- Jang, S.K.; Krausslich, H.G.; Nicklin, M.J.; Duke, G.M.; Palmenberg, A.C.; Wimmer, E. A segment of the 5′ nontranslated region of encephalomyocarditis virus RNA directs internal entry of ribosomes during in vitro translation. J. Virol. 1988, 62, 2636–2643. [Google Scholar] [PubMed]
- Pelletier, J.; Sonenberg, N. Internal initiation of translation of eukaryotic mRNA directed by a sequence derived from poliovirus RNA. Nature 1988, 334, 320–325. [Google Scholar] [CrossRef] [PubMed]
- Balvay, L.; Lopez Lastra, M.; Sargueil, B.; Darlix, J.L.; Ohlmann, T. Translational control of retroviruses. Nat. Rev. Microbiol. 2007, 5, 128–140. [Google Scholar] [CrossRef]
- Berlioz, C.; Darlix, J.L. An internal ribosomal entry mechanism promotes translation of murine leukemia virus gag polyprotein precursors. J. Virol. 1995, 69, 2214–2222. [Google Scholar] [PubMed]
- Jackson, R.J. The current status of vertebrate cellular mRNA IRESs. Cold Spring Harb. Perspect. Biol. 2013, 5, a011569. [Google Scholar] [CrossRef]
- Kwan, T.; Thompson, S.R. Noncanonical Translation Initiation in Eukaryotes. Cold Spring Harb. Perspect. Biol. 2018. [Google Scholar] [CrossRef]
- Tsukiyama-Kohara, K.; Iizuka, N.; Kohara, M.; Nomoto, A. Internal ribosome entry site within hepatitis C virus RNA. J. Virol. 1992, 66, 1476–1483. [Google Scholar]
- Vagner, S.; Waysbort, A.; Marenda, M.; Gensac, M.C.; Amalric, F.; Prats, A.C. Alternative translation initiation of the Moloney murine leukemia virus mRNA controlled by internal ribosome entry involving the p57/PTB splicing factor. J. Biol. Chem. 1995, 270, 20376–20383. [Google Scholar] [CrossRef]
- Wang, C.; Sarnow, P.; Siddiqui, A. Translation of human hepatitis C virus RNA in cultured cells is mediated by an internal ribosome-binding mechanism. J. Virol. 1993, 67, 3338–3344. [Google Scholar]
- Wilson, J.E.; Pestova, T.V.; Hellen, C.U.; Sarnow, P. Initiation of protein synthesis from the A site of the ribosome. Cell 2000, 102, 511–520. [Google Scholar] [CrossRef]
- Macejak, D.G.; Sarnow, P. Internal initiation of translation mediated by the 5′ leader of a cellular mRNA. Nature 1991, 353, 90–94. [Google Scholar] [CrossRef]
- Vagner, S.; Gensac, M.C.; Maret, A.; Bayard, F.; Amalric, F.; Prats, H.; Prats, A.C. Alternative translation of human fibroblast growth factor 2 mRNA occurs by internal entry of ribosomes. Mol. Cell Biol. 1995, 15, 35–44. [Google Scholar] [CrossRef]
- Lee, K.M.; Chen, C.J.; Shih, S.R. Regulation Mechanisms of Viral IRES-Driven Translation. Trends Microbiol. 2017, 25, 547–562. [Google Scholar] [CrossRef]
- Thompson, S.R. So you want to know if your message has an IRES? Wires RNA 2012, 3, 697–705. [Google Scholar] [Green Version]
- Wang, C.; Sarnow, P.; Siddiqui, A. A conserved helical element is essential for internal initiation of translation of hepatitis C virus RNA. J. Virol. 1994, 68, 7301–7307. [Google Scholar]
- Lu, H.H.; Wimmer, E. Poliovirus chimeras replicating under the translational control of genetic elements of hepatitis C virus reveal unusual properties of the internal ribosomal entry site of hepatitis C virus. Proc. Natl. Acad. Sci. USA 1996, 93, 1412–1417. [Google Scholar] [CrossRef]
- Reynolds, J.E.; Kaminski, A.; Carroll, A.R.; Clarke, B.E.; Rowlands, D.J.; Jackson, R.J. Internal initiation of translation of hepatitis C virus RNA: The ribosome entry site is at the authentic initiation codon. RNA 1996, 2, 867–878. [Google Scholar]
- Wang, C.; Le, S.Y.; Ali, N.; Siddiqui, A. An RNA pseudoknot is an essential structural element of the internal ribosome entry site located within the hepatitis C virus 5′ noncoding region. RNA 1995, 1, 526–537. [Google Scholar]
- Hellen, C.U.; Witherell, G.W.; Schmid, M.; Shin, S.H.; Pestova, T.V.; Gil, A.; Wimmer, E. A cytoplasmic 57-kDa protein that is required for translation of picornavirus RNA by internal ribosomal entry is identical to the nuclear pyrimidine tract-binding protein. Proc. Natl. Acad. Sci. USA 1993, 90, 7642–7646. [Google Scholar] [CrossRef]
- Jang, S.K.; Wimmer, E. Cap-independent translation of encephalomyocarditis virus RNA: Structural elements of the internal ribosomal entry site and involvement of a cellular 57-kD RNA-binding protein. Genes Dev. 1990, 4, 1560–1572. [Google Scholar] [CrossRef]
- Luz, N.; Beck, E. Interaction of a cellular 57-kilodalton protein with the internal translation initiation site of foot-and-mouth disease virus. J. Virol. 1991, 65, 6486–6494. [Google Scholar]
- Meerovitch, K.; Pelletier, J.; Sonenberg, N. A cellular protein that binds to the 5′-noncoding region of poliovirus RNA: Implications for internal translation initiation. Genes Dev. 1989, 3, 1026–1034. [Google Scholar] [CrossRef]
- Bonneau, A.M.; Sonenberg, N. Involvement of the 24-kDa cap-binding protein in regulation of protein synthesis in mitosis. J. Biol. Chem. 1987, 262, 11134–11139. [Google Scholar]
- Huang, J.T.; Schneider, R.J. Adenovirus inhibition of cellular protein synthesis involves inactivation of cap-binding protein. Cell 1991, 65, 271–280. [Google Scholar] [CrossRef]
- Lindquist, S. Varying patterns of protein synthesis in Drosophila during heat shock: Implications for regulation. Dev. Biol. 1980, 77, 463–479. [Google Scholar] [CrossRef]
- Huez, I.; Creancier, L.; Audigier, S.; Gensac, M.C.; Prats, A.C.; Prats, H. Two independent internal ribosome entry sites are involved in translation initiation of vascular endothelial growth factor mRNA. Mol. Cell Biol. 1998, 18, 6178–6190. [Google Scholar] [CrossRef]
- Morfoisse, F.; Kuchnio, A.; Frainay, C.; Gomez-Brouchet, A.; Delisle, M.B.; Marzi, S.; Helfer, A.C.; Hantelys, F.; Pujol, F.; Guillermet-Guibert, J.; et al. Hypoxia induces VEGF-C expression in metastatic tumor cells via a HIF-1alpha-independent translation-mediated mechanism. Cell Rep. 2014, 6, 155–167. [Google Scholar] [CrossRef]
- Morfoisse, F.; Tatin, F.; Hantelys, F.; Adoue, A.; Helfer, A.C.; Cassant-Sourdy, S.; Pujol, F.; Gomez-Brouchet, A.; Ligat, L.; Lopez, F.; et al. Nucleolin Promotes Heat Shock-Associated Translation of VEGF-D to Promote Tumor Lymphangiogenesis. Cancer Res. 2016, 76, 4394–4405. [Google Scholar] [CrossRef]
- Nanbru, C.; Lafon, I.; Audigier, S.; Gensac, M.C.; Vagner, S.; Huez, G.; Prats, A.C. Alternative translation of the proto-oncogene c-myc by an internal ribosome entry site. J. Biol. Chem. 1997, 272, 32061–32066. [Google Scholar] [CrossRef]
- Oh, S.K.; Scott, M.P.; Sarnow, P. Homeotic gene Antennapedia mRNA contains 5′-noncoding sequences that confer translational initiation by internal ribosome binding. Genes Dev. 1992, 6, 1643–1653. [Google Scholar] [CrossRef]
- Prats, A.C.; Prats, H. Translational control of gene expression: Role of IRESs and consequences for cell transformation and angiogenesis. Prog. Nucleic Acid Res. Mol. Biol. 2002, 72, 367–413. [Google Scholar]
- Stoneley, M.; Paulin, F.E.; Le Quesne, J.P.; Chappell, S.A.; Willis, A.E. C-Myc 5′ untranslated region contains an internal ribosome entry segment. Oncogene 1998, 16, 423–428. [Google Scholar] [CrossRef] [Green Version]
- Braunstein, S.; Karpisheva, K.; Pola, C.; Goldberg, J.; Hochman, T.; Yee, H.; Cangiarella, J.; Arju, R.; Formenti, S.C.; Schneider, R.J. A hypoxia-controlled cap-dependent to cap-independent translation switch in breast cancer. Mol. Cell 2007, 28, 501–512. [Google Scholar] [CrossRef]
- Bellodi, C.; Kopmar, N.; Ruggero, D. Deregulation of oncogene-induced senescence and p53 translational control in X-linked dyskeratosis congenita. Embo J. 2010, 29, 1865–1876. [Google Scholar] [CrossRef] [Green Version]
- Bonnal, S.; Pileur, F.; Orsini, C.; Parker, F.; Pujol, F.; Prats, A.C.; Vagner, S. Heterogeneous nuclear ribonucleoprotein A1 is a novel internal ribosome entry site trans-acting factor that modulates alternative initiation of translation of the fibroblast growth factor 2 mRNA. J. Biol. Chem. 2005, 280, 4144–4153. [Google Scholar] [CrossRef]
- Martineau, Y.; Le Bec, C.; Monbrun, L.; Allo, V.; Chiu, I.M.; Danos, O.; Moine, H.; Prats, H.; Prats, A.C. Internal ribosome entry site structural motifs conserved among mammalian fibroblast growth factor 1 alternatively spliced mRNAs. Mol. Cell Biol. 2004, 24, 7622–7635. [Google Scholar] [CrossRef]
- Karginov, T.A.; Pastor, D.P.H.; Semler, B.L.; Gomez, C.M. Mammalian Polycistronic mRNAs and Disease. Trends Genet. 2017, 33, 129–142. [Google Scholar] [CrossRef] [Green Version]
- Arnaud, E.; Touriol, C.; Boutonnet, C.; Gensac, M.C.; Vagner, S.; Prats, H.; Prats, A.C. A new 34-kilodalton isoform of human fibroblast growth factor 2 is cap dependently synthesized by using a non-AUG start codon and behaves as a survival factor. Mol. Cell Biol. 1999, 19, 505–514. [Google Scholar] [CrossRef]
- Huez, I.; Bornes, S.; Bresson, D.; Creancier, L.; Prats, H. New vascular endothelial growth factor isoform generated by internal ribosome entry site-driven CUG translation initiation. Mol. Endocrinol. 2001, 15, 2197–2210. [Google Scholar] [CrossRef]
- Prats, H.; Kaghad, M.; Prats, A.C.; Klagsbrun, M.; Lelias, J.M.; Liauzun, P.; Chalon, P.; Tauber, J.P.; Amalric, F.; Smith, J.A.; et al. High molecular mass forms of basic fibroblast growth factor are initiated by alternative CUG codons. Proc. Natl. Acad. Sci. USA 1989, 86, 1836–1840. [Google Scholar] [CrossRef]
- Nanbru, C.; Prats, A.C.; Droogmans, L.; Defrance, P.; Huez, G.; Kruys, V. Translation of the human c-myc P0 tricistronic mRNA involves two independent internal ribosome entry sites. Oncogene 2001, 20, 4270–4280. [Google Scholar] [CrossRef] [Green Version]
- Xiong, Z.; Liu, E.; Yan, Y.; Silver, R.T.; Yang, F.; Chen, I.H.; Chen, Y.; Verstovsek, S.; Wang, H.; Prchal, J.; et al. An unconventional antigen translated by a novel internal ribosome entry site elicits antitumor humoral immune reactions. J. Immunol. 2006, 177, 4907–4916. [Google Scholar] [CrossRef]
- Hui, D.; Kumar, K.N.; Mach, J.R.; Srinivasan, A.; Pal, R.; Bao, X.; Agbas, A.; Hofner, G.; Wanner, K.T.; Michaelis, E.K. A rat brain bicistronic gene with an internal ribosome entry site codes for a phencyclidine-binding protein with cytotoxic activity. J. Biol. Chem. 2009, 284, 2245–2257. [Google Scholar] [CrossRef]
- Bahar Halpern, K.; Veprik, A.; Rubins, N.; Naaman, O.; Walker, M.D. GPR41 gene expression is mediated by internal ribosome entry site (IRES)-dependent translation of bicistronic mRNA encoding GPR40 and GPR41 proteins. J. Biol. Chem. 2012, 287, 20154–20163. [Google Scholar] [CrossRef]
- Holcik, M.; Lefebvre, C.; Yeh, C.L.; Chow, T.; Korneluk, R.G. A new internal-ribosome-entry-site motif potentiates XIAP-mediated cytoprotection. Nat. Cell Biol. 1999, 1, 190–192. [Google Scholar] [CrossRef]
- Coldwell, M.J.; Mitchell, S.A.; Stoneley, M.; MacFarlane, M.; Willis, A.E. Initiation of Apaf-1 translation by internal ribosome entry. Oncogene 2000, 19, 899–905. [Google Scholar] [CrossRef] [Green Version]
- Ray, P.S.; Grover, R.; Das, S. Two internal ribosome entry sites mediate the translation of p53 isoforms. Embo Rep. 2006, 7, 404–410. [Google Scholar] [CrossRef] [Green Version]
- Stoneley, M.; Chappell, S.A.; Jopling, C.L.; Dickens, M.; MacFarlane, M.; Willis, A.E. c-Myc protein synthesis is initiated from the internal ribosome entry segment during apoptosis. Mol. Cell Biol. 2000, 20, 1162–1169. [Google Scholar] [CrossRef]
- Yang, D.Q.; Halaby, M.J.; Zhang, Y. The identification of an internal ribosomal entry site in the 5′-untranslated region of p53 mRNA provides a novel mechanism for the regulation of its translation following DNA damage. Oncogene 2006, 25, 4613–4619. [Google Scholar] [CrossRef] [Green Version]
- Holcik, M.; Sonenberg, N. Translational control in stress and apoptosis. Nat. Rev. Mol. Cell Biol. 2005, 6, 318–327. [Google Scholar] [CrossRef]
- Creancier, L.; Morello, D.; Mercier, P.; Prats, A.C. Fibroblast growth factor 2 internal ribosome entry site (IRES) activity ex vivo and in transgenic mice reveals a stringent tissue-specific regulation. J. Cell Biol. 2000, 150, 275–281. [Google Scholar] [CrossRef]
- Audigier, S.; Guiramand, J.; Prado-Lourenco, L.; Conte, C.; Gonzalez-Herrera, I.G.; Cohen-Solal, C.; Recasens, M.; Prats, A.C. Potent activation of FGF-2 IRES-dependent mechanism of translation during brain development. RNA 2008, 14, 1852–1864. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Herrera, I.G.; Prado-Lourenco, L.; Pileur, F.; Conte, C.; Morin, A.; Cabon, F.; Prats, H.; Vagner, S.; Bayard, F.; Audigier, S.; et al. Testosterone regulates FGF-2 expression during testis maturation by an IRES-dependent translational mechanism. Faseb J. 2006, 20, 476–478. [Google Scholar] [CrossRef]
- Conte, C.; Ainaoui, N.; Delluc-Clavieres, A.; Khoury, M.P.; Azar, R.; Pujol, F.; Martineau, Y.; Pyronnet, S.; Prats, A.C. Fibroblast growth factor 1 induced during myogenesis by a transcription-translation coupling mechanism. Nucleic Acids Res. 2009, 37, 5267–5278. [Google Scholar] [CrossRef]
- Creancier, L.; Mercier, P.; Prats, A.C.; Morello, D. c-myc Internal ribosome entry site activity is developmentally controlled and subjected to a strong translational repression in adult transgenic mice. Mol. Cell Biol. 2001, 21, 1833–1840. [Google Scholar] [CrossRef]
- Xue, S.; Tian, S.; Fujii, K.; Kladwang, W.; Das, R.; Barna, M. RNA regulons in Hox 5′ UTRs confer ribosome specificity to gene regulation. Nature 2015, 517, 33–38. [Google Scholar] [CrossRef]
- Weingarten-Gabbay, S.; Elias-Kirma, S.; Nir, R.; Gritsenko, A.A.; Stern-Ginossar, N.; Yakhini, Z.; Weinberger, A.; Segal, E. Systematic discovery of cap-independent translation sequences in human and viral genomes. Science 2016, 351, aad4939. [Google Scholar] [CrossRef]
- Chappell, S.A.; LeQuesne, J.P.C.; Paulin, F.E.M.; deSchoolmeester, M.L.; Stoneley, M.; Soutar, R.L.; Ralston, S.H.; Helfrich, M.H.; Willis, A.E. A mutation in the c-myc-IRES leads to enhanced internal ribosome entry in multiple myeloma: A novel mechanism of oncogene de-regulation. Oncogene 2000, 19, 4437–4440. [Google Scholar] [CrossRef] [Green Version]
- Hudder, A.; Werner, R. Analysis of a Charcot-Marie-Tooth disease mutation reveals an essential internal ribosome entry site element in the connexin-32 gene. J. Biol. Chem. 2000, 275, 34586–34591. [Google Scholar] [CrossRef]
- Lambrechts, D.; Storkebaum, E.; Morimoto, M.; Del-Favero, J.; Desmet, F.; Marklund, S.L.; Wyns, S.; Thijs, V.; Andersson, J.; van Marion, I.; et al. VEGF is a modifier of amyotrophic lateral sclerosis in mice and humans and protects motoneurons against ischemic death. Nat. Genet. 2003, 34, 383–394. [Google Scholar] [CrossRef] [Green Version]
- Marcel, V.; Ghayad, S.E.; Belin, S.; Therizols, G.; Morel, A.P.; Solano-Gonzalez, E.; Vendrell, J.A.; Hacot, S.; Mertani, H.C.; Albaret, M.A.; et al. p53 acts as a safeguard of translational control by regulating fibrillarin and rRNA methylation in cancer. Cancer Cell 2013, 24, 318–330. [Google Scholar] [CrossRef]
- Ryoo, H.D.; Vasudevan, D. Two distinct nodes of translational inhibition in the Integrated Stress Response. Bmb Rep. 2017, 50, 539–545. [Google Scholar] [CrossRef] [Green Version]
- Hinnebusch, A.G. Translational regulation of GCN4 and the general amino acid control of yeast. Annu. Rev. Microbiol. 2005, 59, 407–450. [Google Scholar] [CrossRef]
- Fernandez, J.; Yaman, I.; Sarnow, P.; Snider, M.D.; Hatzoglou, M. Regulation of internal ribosomal entry site-mediated translation by phosphorylation of the translation initiation factor eIF2 alpha. J. Biol. Chem. 2002, 277, 19198–19205. [Google Scholar] [CrossRef]
- Kang, M.J.; Vasudevan, D.; Kang, K.; Kim, K.; Park, J.E.; Zhang, N.; Zeng, X.M.; Neubert, T.A.; Marr, M.T.; Ryoo, H.D. 4E-BP is a target of the GCN2 ATF4 pathway during Drosophila development and aging. J. Cell Biol. 2017, 216, 115–129. [Google Scholar] [CrossRef]
- Thakor, N.; Holcik, M. IRES-mediated translation of cellular messenger RNA operates in eIF2 alpha-independent manner during stress. Nucleic Acids Res 2012, 40, 541–552. [Google Scholar] [CrossRef]
- Holcik, M. Could the elF2 alpha-Independent Translation Be the Achilles Heel of Cancer? Front. Oncol. 2015, 5, 264. [Google Scholar] [CrossRef]
- Bornes, S.; Prado-Lourenco, L.; Bastide, A.; Zanibellato, C.; Iacovoni, J.S.; Lacazette, E.; Prats, A.C.; Touriol, C.; Prats, H. Translational induction of VEGF internal ribosome entry site elements during the early response to ischemic stress. Circ. Res. 2007, 100, 305–308. [Google Scholar] [CrossRef]
- Conte, C.; Riant, E.; Toutain, C.; Pujol, F.; Arnal, J.F.; Lenfant, F.; Prats, A.C. FGF2 translationally induced by hypoxia is involved in negative and positive feedback loops with HIF-1alpha. PLoS ONE 2008, 3, e3078. [Google Scholar] [CrossRef]
- Hantelys FGodet, A.C.; Tatin, F.; Renaud-Gabardos, E.; Pujol, F.A.I.; Ligat, L.; Henras, A.; Lacazette, E.; Parini, A.; Garmy-Susini, A. aPAC Vasohibin1, a new IRES trans-acting factor for sequential induction of angiogenic factors in hypoxia. BioRxiv 2019. [Google Scholar] [CrossRef]
- Renaud-Gabardos, E.; Tatin, F.; Hantelys, F.; Lebas, B.; Calise, D.; Kunduzova, O.; Masri, B.; Pujol, F.; Sicard, P.; Valet, P.; et al. Therapeutic Benefit and Gene Network Regulation by Combined Gene Transfer of Apelin, FGF2, and SERCA2a into Ischemic Heart. Mol. Ther. 2018, 26, 902–916. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Wang, C.F.; Zhao, K.L.; Zhang, G.; Wang, D.C.; Mei, Y.D. TRMP, a p53-inducible long noncoding RNA, regulates G1/S cell cycle progression by modulating IRES-dependent p27 translation. Cell Death Dis. 2018, 9, 886. [Google Scholar] [CrossRef] [PubMed]
- Masaki, T.; Arend, K.C.; Li, Y.; Yamane, D.; McGivern, D.R.; Kato, T.; Wakita, T.; Moorman, N.J.; Lemon, S.M. miR-122 stimulates hepatitis C virus RNA synthesis by altering the balance of viral RNAs engaged in replication versus translation. Cell Host Microbe 2015, 17, 217–228. [Google Scholar] [CrossRef] [PubMed]
- Sharathchandra, A.; Lal, R.; Khan, D.; Das, S. Annexin A2 and PSF proteins interact with p53 IRES and regulate translation of p53 mRNA. RNA Biol. 2012, 9, 1429–1439. [Google Scholar] [CrossRef] [Green Version]
- Fox, J.T.; Shin, W.K.; Caudill, M.A.; Stover, P.J. A UV-responsive internal ribosome entry site enhances serine hydroxymethyltransferase 1 expression for DNA damage repair. J. Biol. Chem. 2009, 284, 31097–31108. [Google Scholar] [CrossRef]
- Zheng, Y.; Miskimins, W.K. CUG-binding protein represses translation of p27Kip1 mRNA through its internal ribosomal entry site. RNA Biol. 2011, 8, 365–371. [Google Scholar] [CrossRef] [Green Version]
- Hanson, P.J.; Ye, X.; Qiu, Y.; Zhang, H.M.; Hemida, M.G.; Wang, F.; Lim, T.; Gu, A.; Cho, B.; Kim, H.; et al. Cleavage of DAP5 by coxsackievirus B3 2A protease facilitates viral replication and enhances apoptosis by altering translation of IRES-containing genes. Cell Death Differ. 2016, 23, 828–840. [Google Scholar] [CrossRef]
- Henis-Korenblit, S.; Shani, G.; Sines, T.; Marash, L.; Shohat, G.; Kimchi, A. The caspase-cleaved DAP5 protein supports internal ribosome entry site-mediated translation of death proteins. Proc. Natl. Acad. Sci. USA 2002, 99, 5400–5405. [Google Scholar] [CrossRef] [Green Version]
- Henis-Korenblit, S.; Strumpf, N.L.; Goldstaub, D.; Kimchi, A. A novel form of DAP5 protein accumulates in apoptotic cells as a result of caspase cleavage and internal ribosome entry site-mediated translation. Mol. Cell Biol. 2000, 20, 496–506. [Google Scholar] [CrossRef] [PubMed]
- Lewis, S.M.; Holcik, M. For IRES trans-acting factors, it is all about location. Oncogene 2008, 27, 1033–1035. [Google Scholar] [CrossRef] [PubMed]
- Liberman, N.; Gandin, V.; Svitkin, Y.V.; David, M.; Virgili, G.; Jaramillo, M.; Holcik, M.; Nagar, B.; Kimchi, A.; Sonenberg, N. DAP5 associates with eIF2beta and eIF4AI to promote Internal Ribosome Entry Site driven translation. Nucleic Acids Res. 2015, 43, 3764–3775. [Google Scholar] [CrossRef] [PubMed]
- Liberman, N.; Marash, L.; Kimchi, A. The translation initiation factor DAP5 is a regulator of cell survival during mitosis. Cell Cycle 2009, 8, 204–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marash, L.; Liberman, N.; Henis-Korenblit, S.; Sivan, G.; Reem, E.; Elroy-Stein, O.; Kimchi, A. DAP5 promotes cap-independent translation of Bcl-2 and CDK1 to facilitate cell survival during mitosis. Mol Cell. 2008, 30, 447–459. [Google Scholar] [CrossRef]
- Nevins, T.A.; Harder, Z.M.; Korneluk, R.G.; Holcik, M. Distinct regulation of internal ribosome entry site-mediated translation following cellular stress is mediated by apoptotic fragments of eIF4G translation initiation factor family members eIF4GI and p97/DAP5/NAT1. J. Biol. Chem. 2003, 278, 3572–3579. [Google Scholar] [CrossRef]
- Warnakulasuriyarachchi, D.; Cerquozzi, S.; Cheung, H.H.; Holcik, M. Translational induction of the inhibitor of apoptosis protein HIAP2 during endoplasmic reticulum stress attenuates cell death and is mediated via an inducible internal ribosome entry site element. J. Biol. Chem. 2004, 279, 17148–17157. [Google Scholar] [CrossRef]
- Weingarten-Gabbay, S.; Khan, D.; Liberman, N.; Yoffe, Y.; Bialik, S.; Das, S.; Oren, M.; Kimchi, A. The translation initiation factor DAP5 promotes IRES-driven translation of p53 mRNA. Oncogene 2014, 33, 611–618. [Google Scholar] [CrossRef]
- Lampe, S.; Kunze, M.; Scholz, A.; Brauss, T.F.; Winslow, S.; Simm, S.; Keller, M.; Heidler, J.; Wittig, I.; Brune, B.; et al. Identification of the TXNIP IRES and characterization of the impact of regulatory IRES trans-acting factors. Biochim. Biophys. Acta Gene Regul. Mech. 2018, 1861, 147–157. [Google Scholar] [CrossRef]
- Tsai, B.P.; Wang, X.; Huang, L.; Waterman, M.L. Quantitative profiling of in vivo-assembled RNA-protein complexes using a novel integrated proteomic approach. Mol. Cell Proteomics 2011, 10, M110 007385. [Google Scholar] [CrossRef]
- Cobbold, L.C.; Spriggs, K.A.; Haines, S.J.; Dobbyn, H.C.; Hayes, C.; de Moor, C.H.; Lilley, K.S.; Bushell, M.; Willis, A.E. Identification of internal ribosome entry segment (IRES)-trans-acting factors for the Myc family of IRESs. Mol. Cell Biol. 2008, 28, 40–49. [Google Scholar] [CrossRef]
- Woeller, C.F.; Fox, J.T.; Perry, C.; Stover, P.J. A ferritin-responsive internal ribosome entry site regulates folate metabolism. J. Biol. Chem. 2007, 282, 29927–29935. [Google Scholar] [CrossRef]
- Malbert-Colas, L.; Ponnuswamy, A.; Olivares-Illana, V.; Tournillon, A.S.; Naski, N.; Fahraeus, R. HDMX folds the nascent p53 mRNA following activation by the ATM kinase. Mol. Cell 2014, 54, 500–511. [Google Scholar] [CrossRef] [PubMed]
- Cammas, A.; Pileur, F.; Bonnal, S.; Lewis, S.M.; Leveque, N.; Holcik, M.; Vagner, S. Cytoplasmic relocalization of heterogeneous nuclear ribonucleoprotein A1 controls translation initiation of specific mRNAs. Mol. Biol. Cell 2007, 18, 5048–5059. [Google Scholar] [CrossRef] [PubMed]
- Jo, O.D.; Martin, J.; Bernath, A.; Masri, J.; Lichtenstein, A.; Gera, J. Heterogeneous nuclear ribonucleoprotein A1 regulates cyclin D1 and c-myc internal ribosome entry site function through Akt signaling. (vol 283, pg 23274, 2008). J. Biol. Chem. 2008, 283, 31268. [Google Scholar] [CrossRef]
- Kim, H.J.; Lee, H.R.; Seo, J.Y.; Ryu, H.G.; Lee, K.H.; Kim, D.Y.; Kim, K.T. Heterogeneous nuclear ribonucleoprotein A1 regulates rhythmic synthesis of mouse Nfil3 protein via IRES-mediated translation. Sci. Rep. 2017, 7, 42882. [Google Scholar] [CrossRef] [Green Version]
- Kunze, M.M.; Benz, F.; Brauss, T.F.; Lampe, S.; Weigand, J.E.; Braun, J.; Richter, F.M.; Wittig, I.; Brune, B.; Schmid, T. sST2 translation is regulated by FGF2 via an hnRNP A1-mediated IRES-dependent mechanism. Biochim. Biophys. Acta 2016, 1859, 848–859. [Google Scholar] [CrossRef]
- Lewis, S.M.; Veyrier, A.; Hosszu Ungureanu, N.; Bonnal, S.; Vagner, S.; Holcik, M. Subcellular relocalization of a trans-acting factor regulates XIAP IRES-dependent translation. Mol. Biol. Cell 2007, 18, 1302–1311. [Google Scholar] [CrossRef] [PubMed]
- Roy, R.; Durie, D.; Li, H.; Liu, B.Q.; Skehel, J.M.; Mauri, F.; Cuorvo, L.V.; Barbareschi, M.; Guo, L.; Holcik, M.; et al. hnRNPA1 couples nuclear export and translation of specific mRNAs downstream of FGF-2/S6K2 signalling. Nucleic Acids Res. 2014, 42, 12483–12497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, Y.; Frost, P.J.; Hoang, B.Q.; Benavides, A.; Sharma, S.; Gera, J.F.; Lichtenstein, A.K. IL-6-induced stimulation of c-myc translation in multiple myeloma cells is mediated by myc internal ribosome entry site function and the RNA-binding protein, hnRNP A1. Cancer Res. 2008, 68, 10215–10222. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Yang, Y.; Hoang, B.; Bardeleben, C.; Holmes, B.; Gera, J.; Lichtenstein, A. Therapeutic potential of targeting IRES-dependent c-myc translation in multiple myeloma cells during ER stress. Oncogene 2016, 35, 1015–1024. [Google Scholar] [CrossRef] [PubMed]
- Siculella, L.; Tocci, R.; Rochira, A.; Testini, M.; Gnoni, A.; Damiano, F. Lipid accumulation stimulates the cap-independent translation of SREBP-1a mRNA by promoting hnRNP A1 binding to its 5′-UTR in a cellular model of hepatic steatosis. Biochim. Biophys. Acta 2016, 1861, 471–481. [Google Scholar] [CrossRef] [PubMed]
- Christian, K.J.; Lang, M.A.; Raffalli-Mathieu, F. Interaction of heterogeneous nuclear ribonucleoprotein C1/C2 with a novel cis-regulatory element within p53 mRNA as a response to cytostatic drug treatment. Mol. Pharm. 2008, 73, 1558–1567. [Google Scholar] [CrossRef] [PubMed]
- Holcik, M.; Gordon, B.W.; Korneluk, R.G. The internal ribosome entry site-mediated translation of antiapoptotic protein XIAP is modulated by the heterogeneous nuclear ribonucleoproteins C1 and C2. Mol. Cell Biol. 2003, 23, 280–288. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Paek, K.Y.; Choi, K.; Kim, T.D.; Hahm, B.; Kim, K.T.; Jang, S.K. Heterogeneous nuclear ribonucleoprotein C modulates translation of c-myc mRNA in a cell cycle phase-dependent manner. Mol. Cell Biol. 2003, 23, 708–720. [Google Scholar] [CrossRef] [PubMed]
- Meng, Z.; Jackson, N.L.; Choi, H.; King, P.H.; Emanuel, P.D.; Blume, S.W. Alterations in RNA-binding activities of IRES-regulatory proteins as a mechanism for physiological variability and pathological dysregulation of IGF-IR translational control in human breast tumor cells. J. Cell Physiol. 2008, 217, 172–183. [Google Scholar] [CrossRef]
- Schepens, B.; Tinton, S.A.; Bruynooghe, Y.; Parthoens, E.; Haegman, M.; Beyaert, R.; Cornelis, S. A role for hnRNP C1/C2 and Unr in internal initiation of translation during mitosis. Embo J. 2007, 26, 158–169. [Google Scholar] [CrossRef]
- Omnus, D.J.; Mehrtens, S.; Ritter, B.; Resch, K.; Yamada, M.; Frank, R.; Nourbakhsh, M.; Reboll, M.R. JKTBP1 is involved in stabilization and IRES-dependent translation of NRF mRNAs by binding to 5′ and 3′ untranslated regions. J. Mol. Biol. 2011, 407, 492–504. [Google Scholar] [CrossRef]
- Reboll, M.R.; Oumard, A.; Gazdag, A.C.; Renger, I.; Ritter, B.; Schwarzer, M.; Hauser, H.; Wood, M.; Yamada, M.; Resch, K.; et al. NRF IRES activity is mediated by RNA binding protein JKTBP1 and a 14-nt RNA element. RNA 2007, 13, 1328–1340. [Google Scholar] [CrossRef] [Green Version]
- Dobbyn, H.C.; Hill, K.; Hamilton, T.L.; Spriggs, K.A.; Pickering, B.M.; Coldwell, M.J.; de Moor, C.H.; Bushell, M.; Willis, A.E. Regulation of BAG-1 IRES-mediated translation following chemotoxic stress. Oncogene 2008, 27, 1167–1174. [Google Scholar] [CrossRef]
- Evans, J.R.; Mitchell, S.A.; Spriggs, K.A.; Ostrowski, J.; Bomsztyk, K.; Ostarek, D.; Willis, A.E. Members of the poly (rC) binding protein family stimulate the activity of the c-myc internal ribosome entry segment in vitro and in vivo. Oncogene 2003, 22, 8012–8020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pickering, B.M.; Mitchell, S.A.; Evans, J.R.; Willis, A.E. Polypyrimidine tract binding protein and poly r(C) binding protein 1 interact with the BAG-1 IRES and stimulate its activity in vitro and in vivo. Nucleic Acids Res. 2003, 31, 639–646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Notari, M.; Neviani, P.; Santhanam, R.; Blaser, B.W.; Chang, J.S.; Galietta, A.; Willis, A.E.; Roy, D.C.; Caligiuri, M.A.; Marcucci, G.; et al. A MAPK/HNRPK pathway controls BCR/ABL oncogenic potential by regulating MYC mRNA translation. Blood 2006, 107, 2507–2516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Majumder, M.; Yaman, I.; Gaccioli, F.; Zeenko, V.V.; Wang, C.; Caprara, M.G.; Venema, R.C.; Komar, A.A.; Snider, M.D.; Hatzoglou, M. The hnRNA-binding proteins hnRNP L and PTB are required for efficient translation of the Cat-1 arginine/lysine transporter mRNA during amino acid starvation. Mol. Cell Biol. 2009, 29, 2899–2912. [Google Scholar] [CrossRef] [PubMed]
- Peddigari, S.; Li, P.W.; Rabe, J.L.; Martin, S.L. hnRNPL and nucleolin bind LINE-1 RNA and function as host factors to modulate retrotransposition. Nucleic Acids Res. 2013, 41, 575–585. [Google Scholar] [CrossRef]
- Seo, J.Y.; Kim, D.Y.; Kim, S.H.; Kim, H.J.; Ryu, H.G.; Lee, J.; Lee, K.H.; Kim, K.T. Heterogeneous nuclear ribonucleoprotein (hnRNP) L promotes DNA damage-induced cell apoptosis by enhancing the translation of p53. Oncotarget 2017, 8, 51108–51122. [Google Scholar] [CrossRef]
- Ainaoui, N.; Hantelys, F.; Renaud-Gabardos, E.; Bunel, M.; Lopez, F.; Pujol, F.; Planes, R.; Bahraoui, E.; Pichereaux, C.; Burlet-Schiltz, O.; et al. Promoter-Dependent Translation Controlled by p54nrb and hnRNPM during Myoblast Differentiation. PLoS ONE 2015, 10, e0136466. [Google Scholar] [CrossRef]
- Cho, S.; Park, S.M.; Kim, T.D.; Kim, J.H.; Kim, K.T.; Jang, S.K. BiP internal ribosomal entry site activity is controlled by heat-induced interaction of NSAP1. Mol. Cell Biol. 2007, 27, 368–383. [Google Scholar] [CrossRef]
- Choi, J.H.; Kim, S.H.; Jeong, Y.H.; Kim, S.W.; Min, K.T.; Kim, K.T. hnRNP Q regulates IRES-mediated fmr1 translation in neurons. Mol. Cell Biol. 2018. [Google Scholar] [CrossRef]
- Kim, D.Y.; Kim, W.; Lee, K.H.; Kim, S.H.; Lee, H.R.; Kim, H.J.; Jung, Y.; Choi, J.H.; Kim, K.T. hnRNP Q regulates translation of p53 in normal and stress conditions. Cell Death Differ. 2013, 20, 226–234. [Google Scholar] [CrossRef]
- Kim, D.Y.; Woo, K.C.; Lee, K.H.; Kim, T.D.; Kim, K.T. hnRNP Q and PTB modulate the circadian oscillation of mouse Rev-erb alpha via IRES-mediated translation. Nucleic Acids Res. 2010, 38, 7068–7078. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.D.; Woo, K.C.; Cho, S.; Ha, D.C.; Jang, S.K.; Kim, K.T. Rhythmic control of AANAT translation by hnRNP Q in circadian melatonin production. Genes Dev. 2007, 21, 797–810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.H.; Woo, K.C.; Kim, D.Y.; Kim, T.D.; Shin, J.; Park, S.M.; Jang, S.K.; Kim, K.T. Rhythmic interaction between Period1 mRNA and hnRNP Q leads to circadian time-dependent translation. Mol. Cell Biol. 2012, 32, 717–728. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.R.; Kim, T.D.; Kim, H.J.; Jung, Y.; Lee, D.; Lee, K.H.; Kim, D.Y.; Woo, K.C.; Kim, K.T. Heterogeneous ribonucleoprotein R regulates arylalkylamine N-acetyltransferase synthesis via internal ribosomal entry site-mediated translation in a circadian manner. J. Pineal Res. 2015, 59, 518–529. [Google Scholar] [CrossRef] [PubMed]
- Badawi, A.; Biyanee, A.; Nasrullah, U.; Winslow, S.; Schmid, T.; Pfeilschifter, J.; Eberhardt, W. Inhibition of IRES-dependent translation of caspase-2 by HuR confers chemotherapeutic drug resistance in colon carcinoma cells. Oncotarget 2018, 9, 18367–18385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durie, D.; Hatzoglou, M.; Chakraborty, P.; Holcik, M. HuR controls mitochondrial morphology through the regulation of BclxL translation. Translation 2013, 1, e23980. [Google Scholar] [CrossRef]
- Durie, D.; Lewis, S.M.; Liwak, U.; Kisilewicz, M.; Gorospe, M.; Holcik, M. RNA-binding protein HuR mediates cytoprotection through stimulation of XIAP translation. Oncogene 2011, 30, 1460–1469. [Google Scholar] [CrossRef]
- Kullmann, M.; Gopfert, U.; Siewe, B.; Hengst, L. ELAV/Hu proteins inhibit p27 translation via an IRES element in the p27 5′UTR. Genes Dev. 2002, 16, 3087–3099. [Google Scholar] [CrossRef] [PubMed]
- Meng, Z.; King, P.H.; Nabors, L.B.; Jackson, N.L.; Chen, C.Y.; Emanuel, P.D.; Blume, S.W. The ELAV RNA-stability factor HuR binds the 5′-untranslated region of the human IGF-IR transcript and differentially represses cap-dependent and IRES-mediated translation. Nucleic Acids Res. 2005, 33, 2962–2979. [Google Scholar] [CrossRef]
- Yeh, C.H.; Hung, L.Y.; Hsu, C.; Le, S.Y.; Lee, P.T.; Liao, W.L.; Lin, Y.T.; Chang, W.C.; Tseng, J.T. RNA-binding protein HuR interacts with thrombomodulin 5′untranslated region and represses internal ribosome entry site-mediated translation under IL-1 beta treatment. Mol. Biol. Cell 2008, 19, 3812–3822. [Google Scholar] [CrossRef]
- Gao, W.; Li, Q.; Zhu, R.; Jin, J. La Autoantigen Induces Ribosome Binding Protein 1 (RRBP1) Expression through Internal Ribosome Entry Site (IRES)-Mediated Translation during Cellular Stress Condition. Int. J. Mol. Sci. 2016, 17, 1174. [Google Scholar] [CrossRef] [PubMed]
- Holcik, M.; Korneluk, R.G. Functional characterization of the X-linked inhibitor of apoptosis (XIAP) internal ribosome entry site element: Role of La autoantigen in XIAP translation. Mol. Cell Biol. 2000, 20, 4648–4657. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.K.; Back, S.H.; Rho, J.; Lee, S.H.; Jang, S.K. La autoantigen enhances translation of BiP mRNA. Nucleic Acids Res. 2001, 29, 5009–5016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, L.; Zhu, N.; Zhang, H.; Durden, D.L.; Feng, Y.; Zhou, M. Regulation of XIAP translation and induction by MDM2 following irradiation. Cancer Cell 2009, 15, 363–375. [Google Scholar] [CrossRef]
- Faye, M.D.; Graber, T.E.; Liu, P.; Thakor, N.; Baird, S.D.; Durie, D.; Holcik, M. Nucleotide composition of cellular internal ribosome entry sites defines dependence on NF45 and predicts a posttranscriptional mitotic regulon. Mol. Cell Biol. 2013, 33, 307–318. [Google Scholar] [CrossRef]
- Spriggs, K.A.; Cobbold, L.C.; Ridley, S.H.; Coldwell, M.; Bottley, A.; Bushell, M.; Willis, A.E.; Siddle, K. The human insulin receptor mRNA contains a functional internal ribosome entry segment. Nucleic Acids Res. 2009, 37, 5881–5893. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Guo, K.; Kastan, M.B. Interactions of nucleolin and ribosomal protein L26 (RPL26) in translational control of human p53 mRNA. J. Biol. Chem. 2012, 287, 16467–16476. [Google Scholar] [CrossRef]
- Takagi, M.; Absalon, M.J.; McLure, K.G.; Kastan, M.B. Regulation of p53 translation and induction after DNA damage by ribosomal protein L26 and nucleolin. Cell 2005, 123, 49–63. [Google Scholar] [CrossRef]
- Shen, W.; Liang, X.H.; Sun, H.; De Hoyos, C.L.; Crooke, S.T. Depletion of NEAT1 lncRNA attenuates nucleolar stress by releasing sequestered P54nrb and PSF to facilitate c-Myc translation. PLoS ONE 2017, 12, e0173494. [Google Scholar] [CrossRef]
- Liwak, U.; Thakor, N.; Jordan, L.E.; Roy, R.; Lewis, S.M.; Pardo, O.E.; Seckl, M.; Holcik, M. Tumor Suppressor PDCD4 Represses Internal Ribosome Entry Site-Mediated Translation of Antiapoptotic Proteins and Is Regulated by S6 Kinase 2. Mol. Cell Biol. 2012, 32, 1818–1829. [Google Scholar] [CrossRef] [Green Version]
- Olson, C.M.; Donovan, M.R.; Spellberg, M.J.; Marr, M.T., 2nd. The insulin receptor cellular IRES confers resistance to eIF4A inhibition. Elife 2013, 2, e00542. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Yang, H.S. The role of Pdcd4 in tumour suppression and protein translation. Biol. Cell 2018. [Google Scholar] [CrossRef] [PubMed]
- Wedeken, L.; Singh, P.; Klempnauer, K.H. Tumor suppressor protein Pdcd4 inhibits translation of p53 mRNA. J. Biol. Chem. 2011, 286, 42855–42862. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.; Kim, J.H.; Back, S.H.; Jang, S.K. Polypyrimidine tract-binding protein enhances the internal ribosomal entry site-dependent translation of p27Kip1 mRNA and modulates transition from G1 to S phase. Mol. Cell Biol. 2005, 25, 1283–1297. [Google Scholar] [CrossRef] [PubMed]
- Cobbold, L.C.; Wilson, L.A.; Sawicka, K.; King, H.A.; Kondrashov, A.V.; Spriggs, K.A.; Bushell, M.; Willis, A.E. Upregulated c-myc expression in multiple myeloma by internal ribosome entry results from increased interactions with and expression of PTB-1 and YB-1. Oncogene 2010, 29, 2884–2891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cornelis, S.; Tinton, S.A.; Schepens, B.; Bruynooghe, Y.; Beyaert, R. UNR translation can be driven by an IRES element that is negatively regulated by polypyrimidine tract binding protein. Nucleic Acids Res. 2005, 33, 3095–3108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhar, D.; Venkataramana, M.; Ponnuswamy, A.; Das, S. Role of polypyrimidine tract binding protein in mediating internal initiation of translation of interferon regulatory factor 2 RNA. PLoS ONE 2009, 4, e7049. [Google Scholar] [CrossRef]
- Grover, R.; Ray, P.S.; Das, S. Polypyrimidine tract binding protein regulates IRES-mediated translation of p53 isoforms. Cell Cycle 2008, 7, 2189–2198. [Google Scholar] [CrossRef] [Green Version]
- Ismail, R.; Ul Hussain, M. The up regulation of phosphofructokinase1 (PFK1) protein during chemically induced hypoxia is mediated by the hypoxia-responsive internal ribosome entry site (IRES) element, present in its 5′untranslated region. Biochimie 2017, 139, 38–45. [Google Scholar] [CrossRef]
- Kim, Y.K.; Hahm, B.; Jang, S.K. Polypyrimidine tract-binding protein inhibits translation of bip mRNA. J. Mol. Biol. 2000, 304, 119–133. [Google Scholar] [CrossRef]
- Mitchell, S.A.; Spriggs, K.A.; Coldwell, M.J.; Jackson, R.J.; Willis, A.E. The Apaf-1 internal ribosome entry segment attains the correct structural conformation for function via interactions with PTB and unr. Mol. Cell 2003, 11, 757–771. [Google Scholar] [CrossRef]
- Romanelli, M.G.; Diani, E.; Lievens, P.M.J. New Insights into Functional Roles of the Polypyrimidine Tract-Binding Protein. Int. J. Mol. Sci. 2013, 14, 22906–22932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schepens, B.; Tinton, S.A.; Bruynooghe, Y.; Beyaert, R.; Cornelis, S. The polypyrimidine tract-binding protein stimulates HIF-1alpha IRES-mediated translation during hypoxia. Nucleic Acids Res. 2005, 33, 6884–6894. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Hu, P.; Lin, X.; Han, W.; Zhu, L.; Tan, X.; Ye, F.; Wang, G.; Wu, F.; Yin, B.; et al. PTBP1 induces ADAR1 p110 isoform expression through IRES-like dependent translation control and influences cell proliferation in gliomas. Cell Mol. Life Sci. 2015, 72, 4383–4397. [Google Scholar] [CrossRef] [PubMed]
- Halaby, M.J.; Harris, B.R.; Miskimins, W.K.; Cleary, M.P.; Yang, D.Q. Deregulation of Internal Ribosome Entry Site-Mediated p53 Translation in Cancer Cells with Defective p53 Response to DNA Damage. Mol. Cell Biol. 2015, 35, 4006–4017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, D.; Chattopadhyay, S.; Das, S. Influence of metabolic stress on translation of p53 isoforms. Mol. Cell Oncol. 2016, 3, e1039689. [Google Scholar] [CrossRef] [PubMed]
- Bisio, A.; Latorre, E.; Andreotti, V.; Bressac-de Paillerets, B.; Harland, M.; Scarra, G.B.; Ghiorzo, P.; Spitale, R.C.; Provenzani, A.; Inga, A. The 5′-untranslated region of p16INK4a melanoma tumor suppressor acts as a cellular IRES, controlling mRNA translation under hypoxia through YBX1 binding. Oncotarget 2015, 6, 39980–39994. [Google Scholar] [CrossRef]
- Marr, M.T., 2nd; D’Alessio, J.A.; Puig, O.; Tjian, R. IRES-mediated functional coupling of transcription and translation amplifies insulin receptor feedback. Genes Dev. 2007, 21, 175–183. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Pehar, M.; Liu, Y.; Bhattacharyya, A.; Zhang, S.C.; O’Riordan, K.J.; Burger, C.; D’Adamio, L.; Puglielli, L. The amyloid precursor protein (APP) intracellular domain regulates translation of p44, a short isoform of p53, through an IRES-dependent mechanism. Neurobiol. Aging 2015, 36, 2725–2736. [Google Scholar] [CrossRef] [Green Version]
- Miura, P.; Coriati, A.; Belanger, G.; De Repentigny, Y.; Lee, J.; Kothary, R.; Holcik, M.; Jasmin, B.J. The utrophin A 5′-UTR drives cap-independent translation exclusively in skeletal muscles of transgenic mice and interacts with eEF1A2. Hum. Mol. Genet. 2010, 19, 1211–1220. [Google Scholar] [CrossRef]
- Spriggs, K.A.; Cobbold, L.C.; Jopling, C.L.; Cooper, R.E.; Wilson, L.A.; Stoneley, M.; Coldwell, M.J.; Poncet, D.; Shen, Y.C.; Morley, S.J.; et al. Canonical initiation factor requirements of the Myc family of internal ribosome entry segments. Mol. Cell Biol. 2009, 29, 1565–1574. [Google Scholar] [CrossRef] [PubMed]
- Thakor, N.; Smith, M.D.; Roberts, L.; Faye, M.D.; Patel, H.; Wieden, H.J.; Cate, J.H.D.; Holcik, M. Cellular mRNA recruits the ribosome via eIF3-PABP bridge to initiate internal translation. RNA Biol. 2017, 14, 553–567. [Google Scholar] [CrossRef]
- Morrison, J.K.; Friday, A.J.; Henderson, M.A.; Hao, E.; Keiper, B.D. Induction of cap-independent BiP (hsp-3) and Bcl-2 (ced-9) translation in response to eIF4G (IFG-1) depletion in C. elegans. Translation 2014, 2, e28935. [Google Scholar] [CrossRef]
- Horos, R.; Ijspeert, H.; Pospisilova, D.; Sendtner, R.; Andrieu-Soler, C.; Taskesen, E.; Nieradka, A.; Cmejla, R.; Sendtner, M.; Touw, I.P.; et al. Ribosomal deficiencies in Diamond-Blackfan anemia impair translation of transcripts essential for differentiation of murine and human erythroblasts. Blood 2012, 119, 262–272. [Google Scholar] [CrossRef]
- Hertz, M.I.; Landry, D.M.; Willis, A.E.; Luo, G.X.; Thompson, S.R. Ribosomal Protein S25 Dependency Reveals a Common Mechanism for Diverse Internal Ribosome Entry Sites and Ribosome Shunting. Mol. Cell Biol. 2013, 33, 1016–1026. [Google Scholar] [CrossRef] [PubMed]
- Shi, Z.; Fujii, K.; Kovary, K.M.; Genuth, N.R.; Rost, H.L.; Teruel, M.N.; Barna, M. Heterogeneous Ribosomes Preferentially Translate Distinct Subpools of mRNAs Genome-wide. Mol. Cell 2017, 67, 71–83 e77. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Zhang, M.; Wu, Q.; Peng, J.; Ruan, Y.; Gu, J. Hepsin inhibits CDK11p58 IRES activity by suppressing unr expression and eIF-2alpha phosphorylation in prostate cancer. Cell Signal 2015, 27, 789–797. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.; Wadlington, N.L.; Chen, L.; Zhuang, X.; Brorson, J.R.; Kang, U.J. Loss of PINK1 attenuates HIF-1alpha induction by preventing 4E-BP1-dependent switch in protein translation under hypoxia. J. Neurosci. 2014, 34, 3079–3089. [Google Scholar] [CrossRef] [PubMed]
- Vaklavas, C.; Zinn, K.R.; Samuel, S.L.; Meng, Z.; Grizzle, W.E.; Choi, H.; Blume, S.W. Translational control of the undifferentiated phenotype in ERpositive breast tumor cells: Cytoplasmic localization of ERalpha and impact of IRES inhibition. Oncol. Rep. 2018, 39, 2482–2498. [Google Scholar] [PubMed]
- Kampen, K.R.; Sulima, S.O.; Verbelen, B.; Girardi, T.; Vereecke, S.; Rinaldi, G.; Verbeeck, J.; Op de Beeck, J.; Uyttebroeck, A.; Meijerink, J.P.P.; et al. The ribosomal RPL10 R98S mutation drives IRES-dependent BCL-2 translation in T-ALL. Leukemia 2018. [Google Scholar] [CrossRef]
- Kolupaeva, V.G.; Hellen, C.U.; Shatsky, I.N. Structural analysis of the interaction of the pyrimidine tract-binding protein with the internal ribosomal entry site of encephalomyocarditis virus and foot-and-mouth disease virus RNAs. RNA 1996, 2, 1199–1212. [Google Scholar] [PubMed]
- Kafasla, P.; Morgner, N.; Poyry, T.A.; Curry, S.; Robinson, C.V.; Jackson, R.J. Polypyrimidine tract binding protein stabilizes the encephalomyocarditis virus IRES structure via binding multiple sites in a unique orientation. Mol. Cell 2009, 34, 556–568. [Google Scholar] [CrossRef] [PubMed]
- Carvajal, F.; Vallejos, M.; Walters, B.; Contreras, N.; Hertz, M.I.; Olivares, E.; Caceres, C.J.; Pino, K.; Letelier, A.; Thompson, S.R.; et al. Structural domains within the HIV-1 mRNA and the ribosomal protein S25 influence cap-independent translation initiation. Febs J. 2016, 283, 2508–2527. [Google Scholar] [CrossRef] [Green Version]
- Olivares, E.; Landry, D.M.; Caceres, C.J.; Pino, K.; Rossi, F.; Navarrete, C.; Huidobro-Toro, J.P.; Thompson, S.R.; Lopez-Lastra, M. The 5′ Untranslated Region of the Human T-Cell Lymphotropic Virus Type 1 mRNA Enables Cap-Independent Translation Initiation. J. Virol. 2014, 88, 5936–5955. [Google Scholar] [CrossRef]
- Ji, B.; Harris, B.R.E.; Liu, Y.H.; Deng, Y.B.; Gradilone, S.A.; Cleary, M.P.; Liu, J.H.; Yang, D.Q. Targeting IRES-Mediated p53 Synthesis for Cancer Diagnosis and Therapeutics. Int. J. Mol. Sci. 2017, 18, E93. [Google Scholar] [CrossRef] [PubMed]
- Johnson, A.G.; Grosely, R.; Petrov, A.N.; Puglisi, J.D. Dynamics of IRES-mediated translation. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2017, 372, 20160177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boehringer, D.; Thermann, R.; Ostareck-Lederer, A.; Lewis, J.D.; Stark, H. Structure of the hepatitis C virus IRES bound to the human 80S ribosome: Remodeling of the HCV IRES. Structure 2005, 13, 1695–1706. [Google Scholar] [CrossRef]
- Spahn, C.M.; Kieft, J.S.; Grassucci, R.A.; Penczek, P.A.; Zhou, K.; Doudna, J.A.; Frank, J. Hepatitis C virus IRES RNA-induced changes in the conformation of the 40s ribosomal subunit. Science 2001, 291, 1959–1962. [Google Scholar] [CrossRef]
- Fernandez, I.S.; Bai, X.C.; Murshudov, G.; Scheres, S.H.; Ramakrishnan, V. Initiation of translation by cricket paralysis virus IRES requires its translocation in the ribosome. Cell 2014, 157, 823–831. [Google Scholar] [CrossRef]
- Hashem, Y.; des Georges, A.; Dhote, V.; Langlois, R.; Liao, H.Y.; Grassucci, R.A.; Pestova, T.V.; Hellen, C.U.; Frank, J. Hepatitis-C-virus-like internal ribosome entry sites displace eIF3 to gain access to the 40S subunit. Nature 2013, 503, 539–543. [Google Scholar] [CrossRef] [Green Version]
- Quade, N.; Boehringer, D.; Leibundgut, M.; van den Heuvel, J.; Ban, N. Cryo-EM structure of Hepatitis C virus IRES bound to the human ribosome at 3.9-A resolution. Nat. Commun. 2015, 6, 7646. [Google Scholar] [CrossRef]
- Yamamoto, H.; Collier, M.; Loerke, J.; Ismer, J.; Schmidt, A.; Hilal, T.; Sprink, T.; Yamamoto, K.; Mielke, T.; Burger, J.; et al. Molecular architecture of the ribosome-bound Hepatitis C Virus internal ribosomal entry site RNA. Embo J. 2015, 34, 3042–3058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Damiano, F.; Rochira, A.; Tocci, R.; Alemanno, S.; Gnoni, A.; Siculella, L. hnRNP A1 mediates the activation of the IRES-dependent SREBP-1a mRNA translation in response to endoplasmic reticulum stress. Biochem. J. 2013, 449, 543–553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, J.C.; Hsu, M.; Tarn, W.Y. Cell stress modulates the function of splicing regulatory protein RBM4 in translation control. Proc. Natl. Acad. Sci. USA 2007, 104, 2235–2240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, G.; Dhar, S.; Bedford, M.T. PRMT5 regulates IRES-dependent translation via methylation of hnRNP A1. Nucleic Acids Res. 2017, 45, 4359–4369. [Google Scholar] [CrossRef] [PubMed]
- Dorrello, N.V.; Peschiaroli, A.; Guardavaccaro, D.; Colburn, N.H.; Sherman, N.E.; Pagano, M. S6K1- and betaTRCP-mediated degradation of PDCD4 promotes protein translation and cell growth. Science 2006, 314, 467–471. [Google Scholar] [CrossRef]
- Suk, F.M.; Lin, S.Y.; Lin, R.J.; Hsine, Y.H.; Liao, Y.J.; Fang, S.U.; Liang, Y.C. Bortezomib inhibits Burkitt’s lymphoma cell proliferation by downregulating sumoylated hnRNP K and c-Myc expression. Oncotarget 2015, 6, 25988–26001. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Gao, F.H. Role and molecular mechanism of heterogeneous nuclear ribonucleoprotein K in tumor development and progression. Biomed. Rep. 2016, 4, 657–663. [Google Scholar] [CrossRef] [Green Version]
- Pickering, B.M.; Mitchell, S.A.; Spriggs, K.A.; Stoneley, M.; Willis, A.E. Bag-1 internal ribosome entry segment activity is promoted by structural changes mediated by poly(rC) binding protein 1 and recruitment of polypyrimidine tract binding protein 1. Mol. Cell Biol. 2004, 24, 5595–5605. [Google Scholar] [CrossRef]
- Braunschweig, U.; Gueroussov, S.; Plocik, A.M.; Graveley, B.R.; Blencowe, B.J. Dynamic integration of splicing within gene regulatory pathways. Cell 2013, 152, 1252–1269. [Google Scholar] [CrossRef]
- Pestova, T.V.; Shatsky, I.N.; Hellen, C.U. Functional dissection of eukaryotic initiation factor 4F: The 4A subunit and the central domain of the 4G subunit are sufficient to mediate internal entry of 43S preinitiation complexes. Mol. Cell Biol. 1996, 16, 6870–6878. [Google Scholar] [CrossRef] [PubMed]
- Pestova, T.V.; Hellen, C.U.; Shatsky, I.N. Canonical eukaryotic initiation factors determine initiation of translation by internal ribosomal entry. Mol. Cell Biol. 1996, 16, 6859–6869. [Google Scholar] [CrossRef] [PubMed]
- Sizova, D.V.; Kolupaeva, V.G.; Pestova, T.V.; Shatsky, I.N.; Hellen, C.U. Specific interaction of eukaryotic translation initiation factor 3 with the 5′ nontranslated regions of hepatitis C virus and classical swine fever virus RNAs. J. Virol. 1998, 72, 4775–4782. [Google Scholar] [PubMed]
- Pestova, T.V.; de Breyne, S.; Pisarev, A.V.; Abaeva, I.S.; Hellen, C.U. eIF2-dependent and eIF2-independent modes of initiation on the CSFV IRES: A common role of domain II. Embo J. 2008, 27, 1060–1072. [Google Scholar] [CrossRef] [PubMed]
- Balagopal, V.; Parker, R. Polysomes, P bodies and stress granules: States and fates of eukaryotic mRNAs. Curr. Opin. Cell Biol. 2009, 21, 403–408. [Google Scholar] [CrossRef]
- Borghese, F.; Michiels, T. The leader protein of cardioviruses inhibits stress granule assembly. J. Virol. 2011, 85, 9614–9622. [Google Scholar] [CrossRef] [PubMed]
- Guil, S.; Long, J.C.; Caceres, J.F. hnRNP A1 relocalization to the stress granules reflects a role in the stress response. Mol. Cell Biol. 2006, 26, 5744–5758. [Google Scholar] [CrossRef]
- Galan, A.; Lozano, G.; Pineiro, D.; Martinez-Salas, E. G3BP1 interacts directly with the FMDV IRES and negatively regulates translation. Febs J. 2017, 284, 3202–3217. [Google Scholar] [CrossRef]
- Fujimura, K.; Katahira, J.; Kano, F.; Yoneda, Y.; Murata, M. Selective localization of PCBP2 to cytoplasmic processing bodies. Biochim. Biophys. Acta 2009, 1793, 878–887. [Google Scholar] [CrossRef] [Green Version]
- Fierro-Monti, I.; Mohammed, S.; Matthiesen, R.; Santoro, R.; Burns, J.S.; Williams, D.J.; Proud, C.G.; Kassem, M.; Jensen, O.N.; Roepstorff, P. Quantitative proteomics identifies Gemin5, a scaffolding protein involved in ribonucleoprotein assembly, as a novel partner for eukaryotic initiation factor 4E. J. Proteome Res. 2006, 5, 1367–1378. [Google Scholar] [CrossRef]
- Pacheco, A.; Lopez de Quinto, S.; Ramajo, J.; Fernandez, N.; Martinez-Salas, E. A novel role for Gemin5 in mRNA translation. Nucleic Acids Res. 2009, 37, 582–590. [Google Scholar] [CrossRef] [PubMed]
- Majzoub, K.; Hafirassou, M.L.; Meignin, C.; Goto, A.; Marzi, S.; Fedorova, A.; Verdier, Y.; Vinh, J.; Hoffmann, J.A.; Martin, F.; et al. RACK1 controls IRES-mediated translation of viruses. Cell 2014, 159, 1086–1095. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Ji, H.; Doudna, J.A.; Leary, J.A. Mass spectrometric analysis of the human 40S ribosomal subunit: Native and HCV IRES-bound complexes. Protein Sci. 2005, 14, 1438–1446. [Google Scholar] [CrossRef] [PubMed]
- Colon-Ramos, D.A.; Shenvi, C.L.; Weitzel, D.H.; Gan, E.C.; Matts, R.; Cate, J.; Kornbluth, S. Direct ribosomal binding by a cellular inhibitor of translation. Nat. Struct. Mol. Biol. 2006, 13, 103–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoo, S.J.; Huh, J.R.; Muro, I.; Yu, H.; Wang, L.; Wang, S.L.; Feldman, R.M.; Clem, R.J.; Muller, H.A.; Hay, B.A. Hid, Rpr and Grim negatively regulate DIAP1 levels through distinct mechanisms. Nat. Cell Biol. 2002, 4, 416–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernandez, G.; Vazquez-Pianzola, P.; Sierra, J.M.; Rivera-Pomar, R. Internal ribosome entry site drives cap-independent translation of reaper and heat shock protein 70 mRNAs in Drosophila embryos. RNA 2004, 10, 1783–1797. [Google Scholar] [CrossRef] [Green Version]
- Vazquez-Pianzola, P.; Hernandez, G.; Suter, B.; Rivera-Pomar, R. Different modes of translation for hid, grim and sickle mRNAs in Drosophila. Cell Death Differ. 2007, 14, 286–295. [Google Scholar] [CrossRef]
- Fuchs, G.; Diges, C.; Kohlstaedt, L.A.; Wehner, K.A.; Sarnow, P. Proteomic analysis of ribosomes: Translational control of mRNA populations by glycogen synthase GYS1. J. Mol. Biol. 2011, 410, 118–130. [Google Scholar] [CrossRef]
- Francisco-Velilla, R.; Fernandez-Chamorro, J.; Ramajo, J.; Martinez-Salas, E. The RNA-binding protein Gemin5 binds directly to the ribosome and regulates global translation. Nucleic Acids Res. 2016, 44, 8335–8351. [Google Scholar] [CrossRef]
- Pineiro, D.; Fernandez, N.; Ramajo, J.; Martinez-Salas, E. Gemin5 promotes IRES interaction and translation control through its C-terminal region. Nucleic Acids Res. 2013, 41, 1017–1028. [Google Scholar] [CrossRef]
- Xue, S.; Barna, M. Specialized ribosomes: A new frontier in gene regulation and organismal biology. Nat. Rev. Mol. Cell Biol. 2012, 13, 355–369. [Google Scholar] [CrossRef] [PubMed]
- Landry, D.M.; Hertz, M.I.; Thompson, S.R. RPS25 is essential for translation initiation by the Dicistroviridae and hepatitis C viral IRESs. Genes Dev. 2009, 23, 2753–2764. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
| ITAF | Also Known As | Regulated IRESs | Regulation | Stimulus | Roles in Cell Physiology and Diseases | References |
|---|---|---|---|---|---|---|
| Class I: ITAFs with nucleocytoplasmic translocation | ||||||
| Annexin A2 | p53 | activator | ER stress | cancer | [76] | |
| CUGBP1 | CELF1 | SHMT-1, p27kip | inhibitor/activator | UV irradiation | DNA repair, cell proliferation | [77,78] |
| DAP5 | P97, NAT1, eIF4GII | Bcl-2, Bcl-XL, BAX, APAF-1, DAP5, Δ40p53, CDK1, HIAP2, c-myc, XIAP | activator | viral infection, apoptosis, ER stress, serum starvation, g-irradiation | cell survival or programmed cell death | [79,80,81,82,83,84,85,86,87,88] |
| FBP3 | FUBP3 | TXNIP | activator | Renal cell carcinoma | [89] | |
| FUS | LEF1 | activator | Cancer, amyotrophic lateral sclerosis | [90] | ||
| GRSF1 | c-myc, L-myc, N-myc | activator | cancer | [91] | ||
| H-ferritin | SHMT-1 | activator | UV irradiation | DNA repair | [77,92] | |
| HDMX | p53 | activator | DNA damage | tumour suppression | [93] | |
| hnRNPA1 | XIAP, FGF2, Nfil3, SREBP1-a, c-myc, BCL-XL, cyclin D1, APAF-1, sst2, ER-α HIF1-α | activator/inhibitor | FGF2, lipid accumulation, ER stress, osmotic shock, UV irradiation | multiple myeloma, circadian oscillation | [37,94,95,96,97,98,99,100,101,102] | |
| hnRNPC | hnRNP C1/C2 | p53, IGF1R, unr, c-myc, XIAP | activator | DNA damage, transcription inhibition, growth stimulus, cell cycle | inhibition of apoptosis, cancer | [103,104,105,106,107]] |
| hnRNPD | JKTBP1 | NRF | activator | UV irradiation | cell survival | [108,109] |
| hnRNPE | PCBP, alphaCP | c-myc, BAG1 | activator | Chemotoxic stress | cell survival, tumorigenesis | [110,111,112] |
| hnRNPH2 | SHMT1 | activator | UV irradiation | DNA repair | [77] | |
| hnRNPK | c-myc | activator | myoblast differentiation, proliferation, tumor progression | [111,113] | ||
| hnRNPL | Cat-1, p53, LINE-1 | activator | Amino-acid deprivation/ DNA damage | transposition inhibition | [114,115,116] | |
| hnRNPM | FGF1 | activator | myoblast differentiation | muscle regeneration | [117] | |
| hnRNPQ | NSAP1 | p53, rev-erb-a, Period1, AANAT, Bip, FMRP | activator | apoptosis/ heat shock | circadian oscillation/ cell survival/ axonal growth cone collapse/ Fragile X syndroma, autism | [118,119,120,121,122,123] |
| hnRNPR | AANAT | activator | circadian oscillation | [124] | ||
| HuR | ELAV1 | IGF1R, caspase-2, BcL-XL, XIAP, p27kip, Thrombomodulin | activator/inhibitor | amino-acid deprivation, IL-1b, | cytoprotection, inhibition of apoptosis, cell proliferation, breast cancer | [106,125,126,127,128,129,130] |
| La auto antigen | XIAP, Bip, RRBP1 | activator/inhibitor | serum starvation, paclitaxel, adriamycin | cell survival, malignancy maintenance, hepatocellular carcinoma | [106,125,126,127,128,129,130,131,132,133] | |
| Mdm2 | HDM2 | p53, XIAP | activator | DNA damage, ionizing radiation | resistance to radiation-induced apoptosis | [93,134] |
| NF45 | iIAP1, XIAP, NRF, ELG | activator | ER stress | polyploidy, senescence | [135] | |
| nPTB | IR | activator | cell density, insulin | cell proliferation | [136] | |
| nucleolin | p53, VEGFD, LINE-1 | activator/inhibitor | heat shock, DNA damage | transposition inhibition | [30,115,137,138] | |
| p54nrb | NONO | c-myc, L-myc, N-myc, APAF1, FGF1 | activator | myoblast differentiation, nucleolar stress, apoptosis | muscle regeneration | [91,117,139] |
| PDCD4 | P53, INR, IGF1R, BcL-XL, XIAP | activator/inhibitor | oxidative stress, absence of DNA damage, S6K2 inactivation, FGF2 pathway inhibition | apoptosis, tumour suppression | [140,141,142,143] | |
| PSF | SFPQ | p53, c-myc, L-myc, N-myc, BAG1, LEF1 | activator/inhibitor | nucleolar stress, apoptosis, ER stress | cancer | [76,90,91,139] |
| PTB | hnRNPI/ PTBP1 | p53, p27kip, PFK1, IR, Cat-1, APAF1, HIF1α, IRF2, rev-erb-a, unr, c-myc, N-myc, BAG1, Bip, ADAR1, TXNIP | activator/inhibitor | DNA damage, hypoxia, ER stress, amino-acid deprivation, cell density, insulin | circadian oscillation, cell cycle arrest, apoptosis | [89,91,107,112,114,136,144,145,146,147,148,149,150,151,152,153,154] |
| RHA | NDH II | p53 | activator | DNA damage | tumour suppression | [155] |
| SMAR 1 | p53 | activator/inhibitor | glucose deprivation | cancer (tumor suppressor) | [156] | |
| YB1 | YBX1 | c-myc, L-myc, N-myc, p16INK4 | activator | hypoxia | multiple myeloma, cancer | [91,145,157] |
| Class II: Cytoplasmic ITAFs related to translational machinery | ||||||
| 4E-BP1 | VEGFA, HIF1α, INR | activator | hypoxia, low nutrients, low insulin | cancer, Parkinson | [35,158] | |
| APP (AICD) | Δ40p53 | activator | Alzheimer disease | [159] | ||
| eeF1A2 | utrophin A | activator | muscle regeneration | [160] | ||
| eIF3 | c-myc, N-myc, XIAP | activator | apoptosis, hypoxia | cancer | [161,162] | |
| eIF4A | c-myc, N-myc | activator | apoptosis, hypoxia | cancer | [161] | |
| eIF4GI | APAF-1, DAP5, Bcl-2, Bip, c-myc, L-myc, N-myc, VEGFA, | activator | apoptosis, hypoxia | cancer | [35,86,161,163] | |
| eIF5B | XIAP | activator | apoptosis, hypoxia | cancer | [68] | |
| eL38 | Rpl38 | Hox | activator | development | [58] | |
| eS19 | Rps19 | BAG1, CSDE1, LamB1 | activator | erythroid differentiation, Diamond-Blackfan anemia | [164] | |
| eS25 | Rps25 | APAF-1, BAG1, c-myc, L-myc, Myb, p53, Set7 | activator | ER stress | multiple myeloma | [101,165,166] |
| Gemin5 | TXNIP | activator/inhibitor | Serum starvation | Spinal muscular atrophy | [89] | |
| Hepsin (also in plasmic membrane) | unr | inhibitor | Cell cycle regulation, Prostate cancer | [167] | ||
| PINK1 (also mitochondrial) | HIF1α | activator | hypoxia | Parkinson | [168] | |
| Rack1 | IGF1R | activator/inhibitor | Hepatocellular carcinoma | [169] | ||
| TCP80 | NF90, DRBP76 | p53 | activator | DNA damage | tumour suppression | [155] |
| uL1 | Rpl10A | IGF2, APP, Chmp2A, Bcl-2 | activator | Alzheimer, leukemia, mitochodrial dysfunction | [166,170] | |
| uL24 | Rpl26 | p53 | activator | DNA damage | tumour suppression | [137,138] |
| uL5 | Rpl11 | BAG1, CSDE1, LamB1 | activator | erythroid differentiation, Diamond-Blackfan anemia | [164] | |
| VASH1 (also secreted and nuclear) | Vasohibin 1 | FGF1 | activator | hypoxia | ischemic heart disease | [72] |
| Class III: ncRNA-constituted ITAFs | ||||||
| TRMP | p27kip | inhibitor | induced by p53 | inhibition of cell proliferation, tumor suppressor | [74] | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Godet, A.-C.; David, F.; Hantelys, F.; Tatin, F.; Lacazette, E.; Garmy-Susini, B.; Prats, A.-C. IRES Trans-Acting Factors, Key Actors of the Stress Response. Int. J. Mol. Sci. 2019, 20, 924. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20040924
Godet A-C, David F, Hantelys F, Tatin F, Lacazette E, Garmy-Susini B, Prats A-C. IRES Trans-Acting Factors, Key Actors of the Stress Response. International Journal of Molecular Sciences. 2019; 20(4):924. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20040924
Chicago/Turabian StyleGodet, Anne-Claire, Florian David, Fransky Hantelys, Florence Tatin, Eric Lacazette, Barbara Garmy-Susini, and Anne-Catherine Prats. 2019. "IRES Trans-Acting Factors, Key Actors of the Stress Response" International Journal of Molecular Sciences 20, no. 4: 924. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20040924