The Combined Use of Phenothiazines and Statins Strongly Affects Doxorubicin-Resistance, Apoptosis, and Cox-2 Activity in Colon Cancer Cells

 , , ,
, , ,
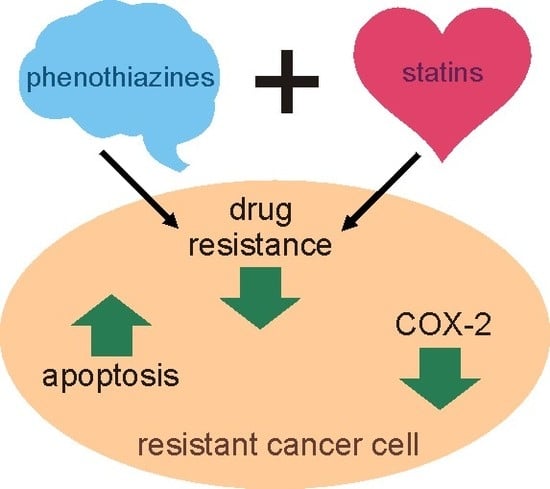
Abstract
:
1. Introduction
2. Results
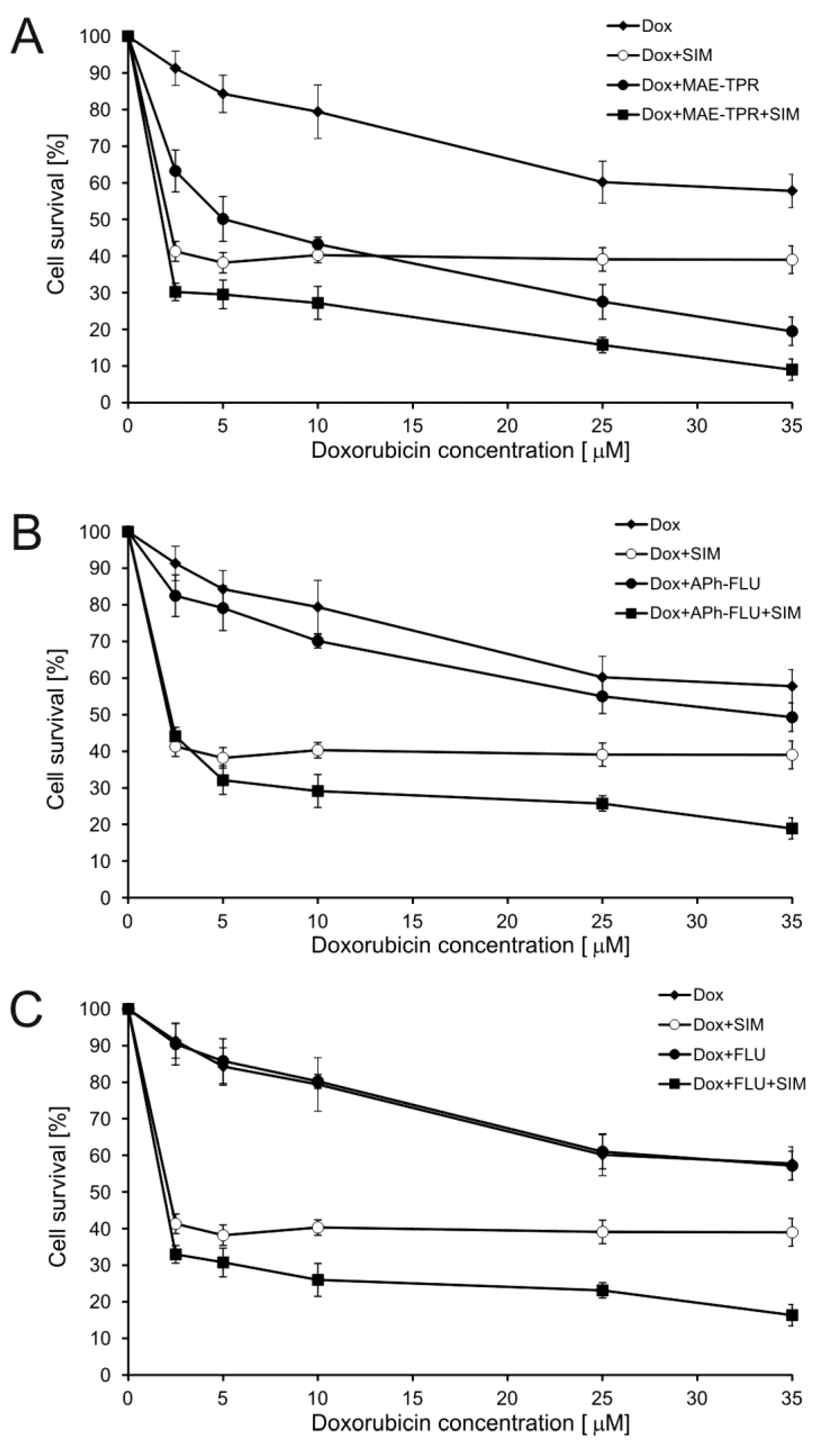
2.1. Doxorubicin Cytotoxicity
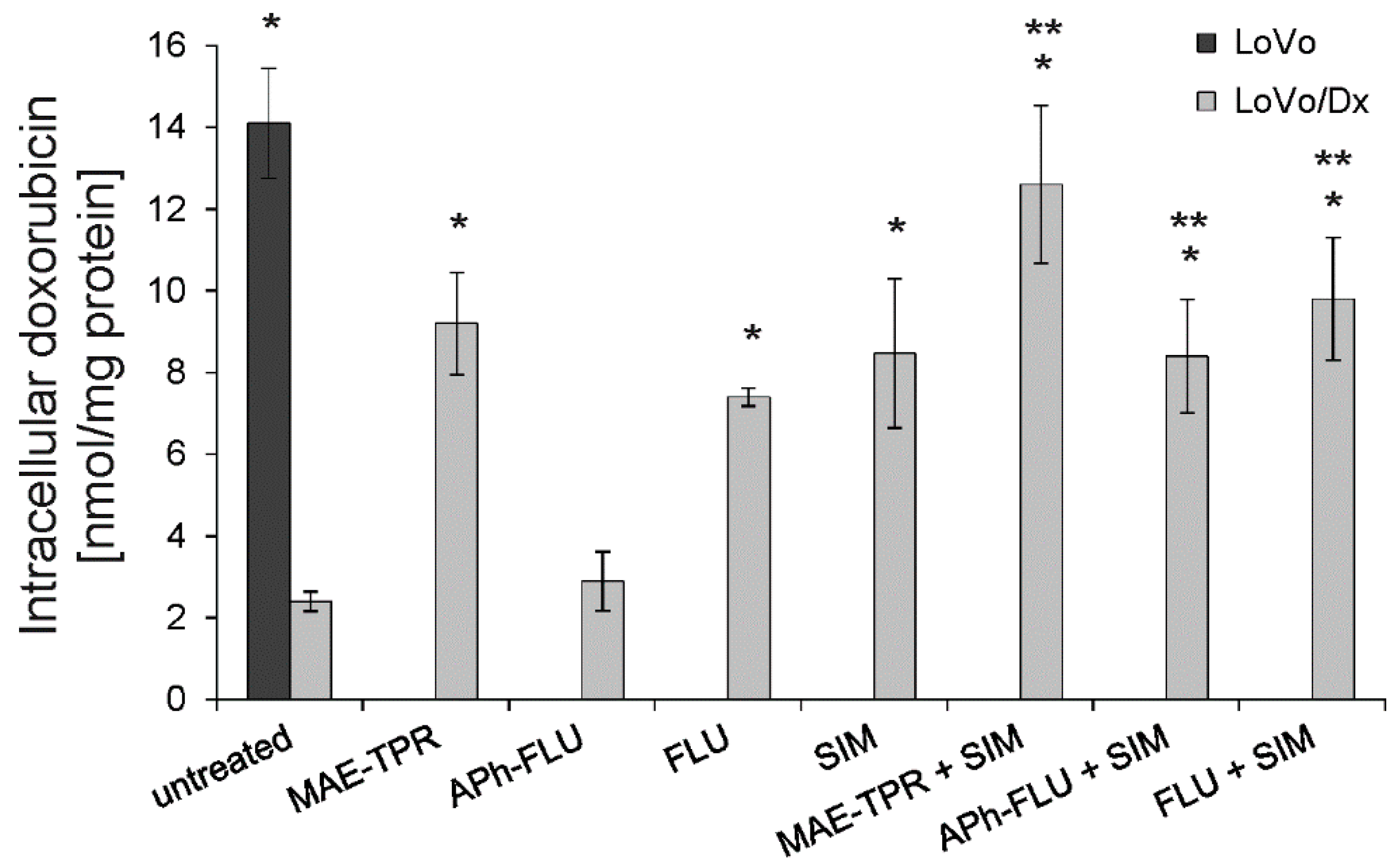
2.2. Doxorubicin Intracellular Accumulation
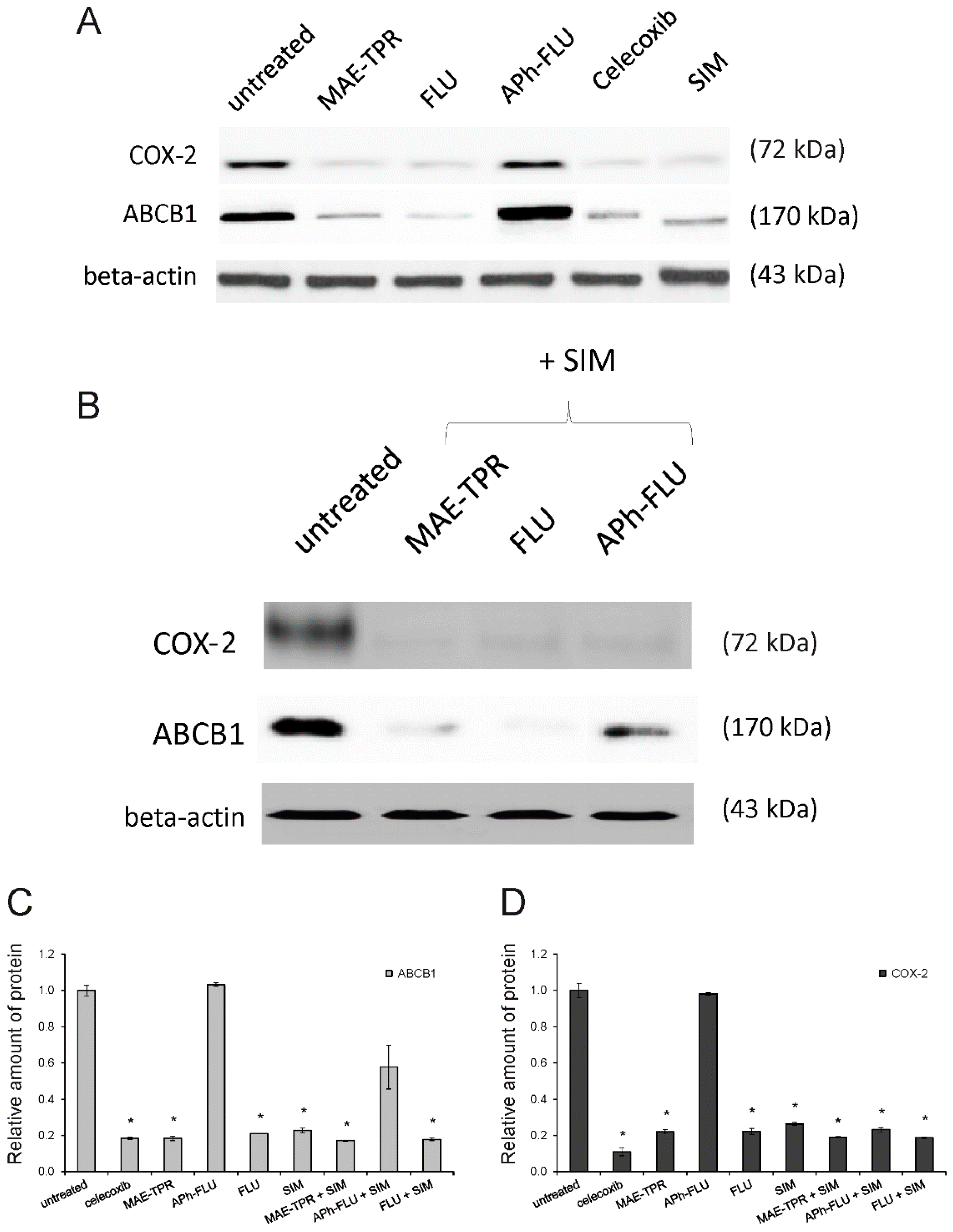
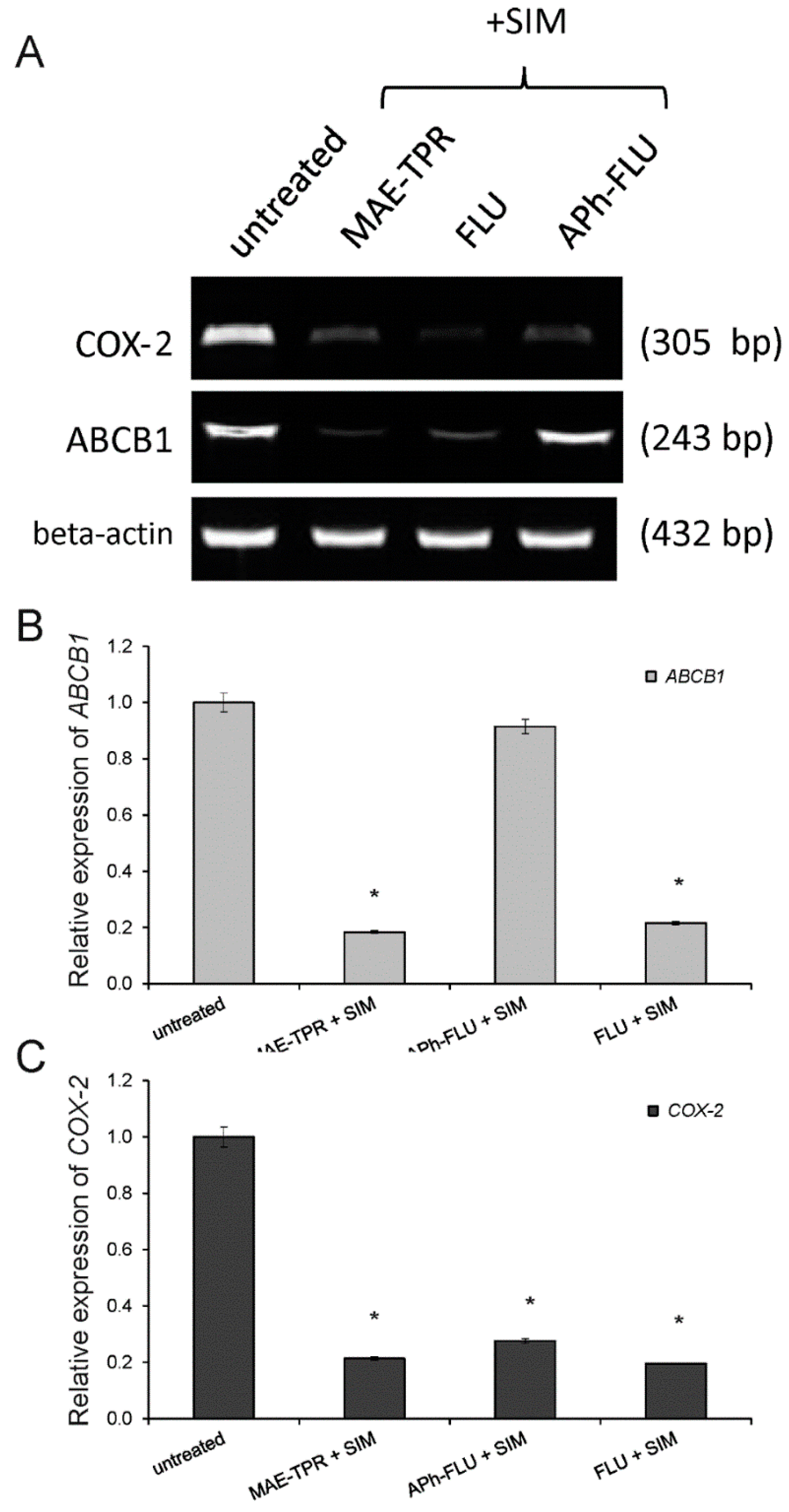
2.3. ABCB1 Expression
2.4. COX-2 Expression
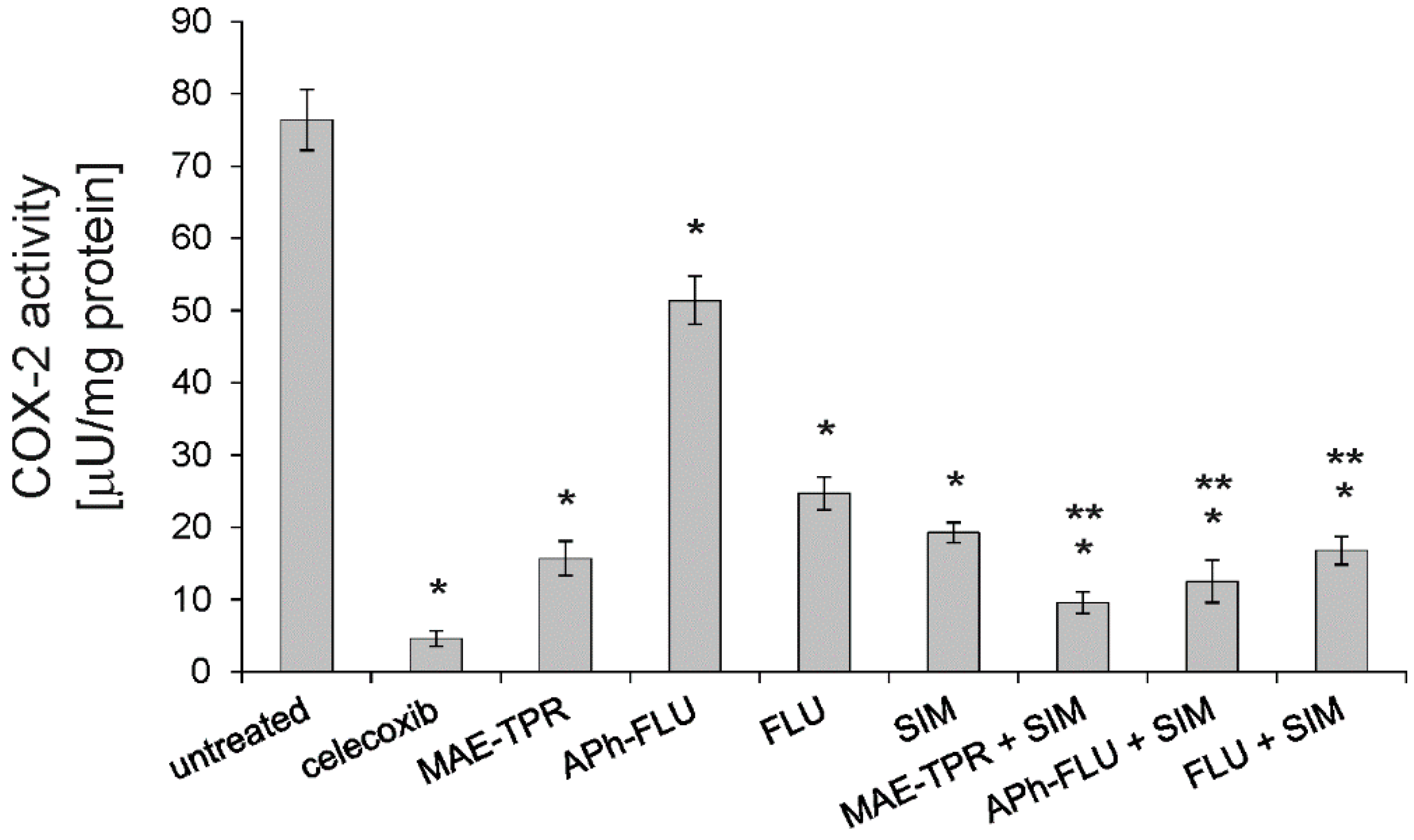
2.5. COX-2 Activity
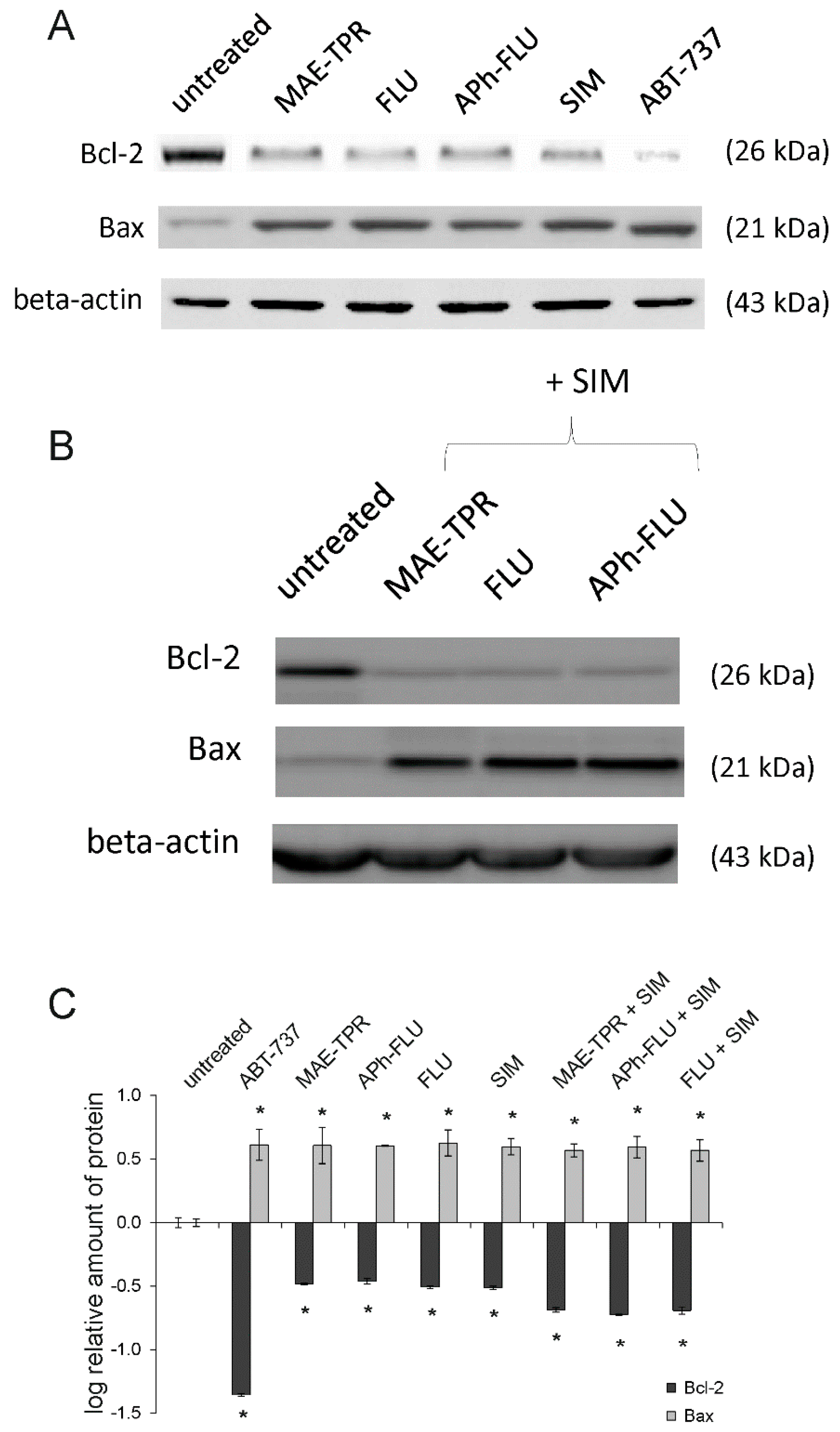
2.6. Bcl-2 and Bax Expression
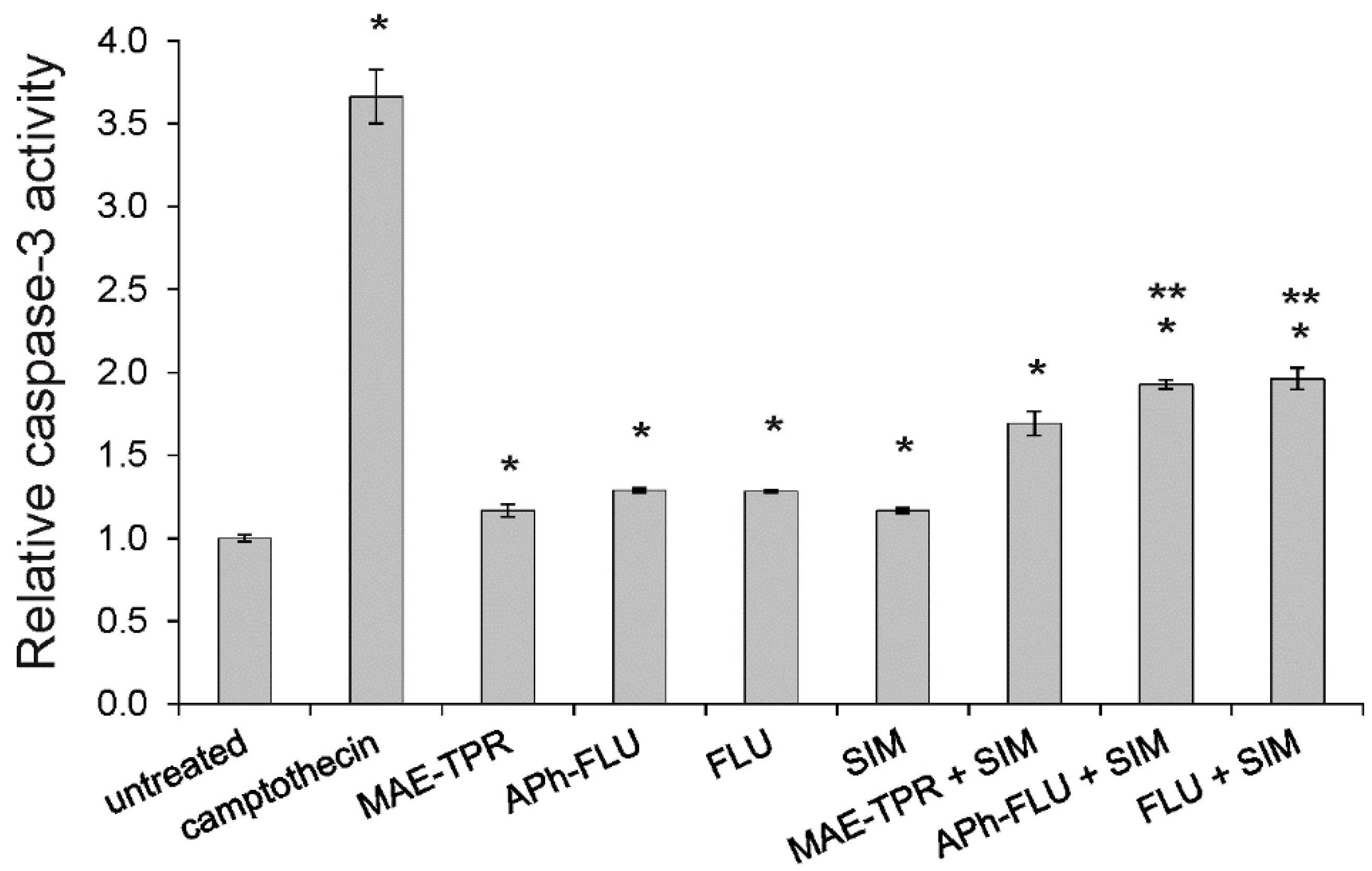
2.7. Caspase-3 Activity
3. Discussion
4. Materials and methods
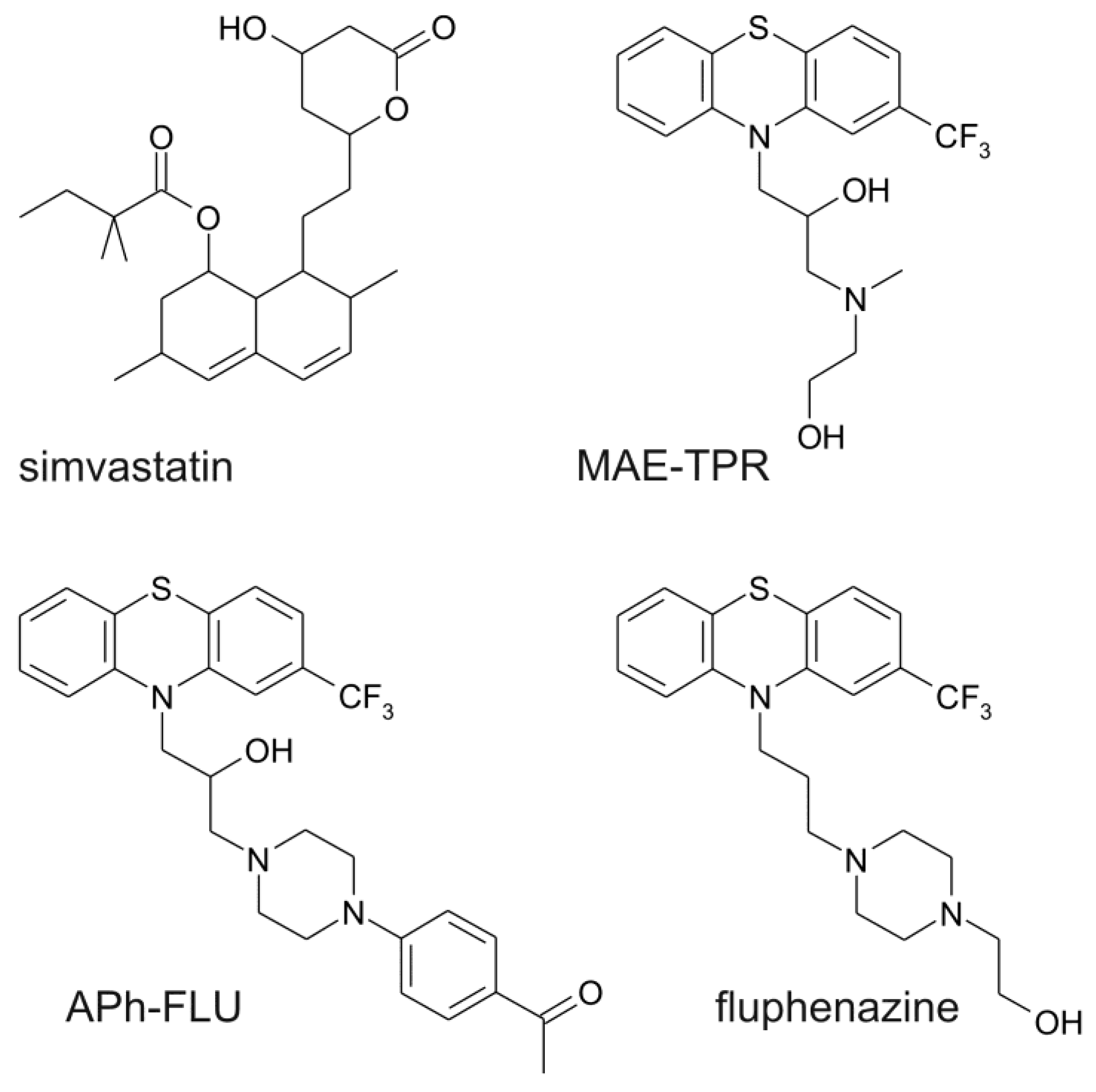
4.1. Chemicals
4.2. Cell Lines
4.3. Cell Viability Assay
4.4. Isobolographic Analysis
4.5. Intracellular Accumulation of Doxorubicin
4.6. Caspase 3 Activation Assay
4.7. Western Blot Analysis
4.8. Polymerase Chain Reaction
4.9. Cyclooxygenase (COX-2) Assay
- B—amount of resorufin from standard curve;
- ΔT—reaction time;
- M—protein amount added into the reaction well.
4.10. Data Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Kane, S.E. Multidrug resistance of cancer cells. Adv. Drug Res. 1996, 28, 181–252. [Google Scholar] [CrossRef]
- Lage, H. An overview of cancer multidrug resistance: A still unsolved problem. Cell. Mol. Life Sci. 2008, 65, 3145–3167. [Google Scholar] [CrossRef] [PubMed]
- Sano, H.; Kawahito, Y.; Wilder, R.L.; Hashiramoto, A.; Mukai, S.; Asai, K.; Kimura, S.; Kato, H.; Kondo, M.; Hla, T. Expression of cyclooxygenase-1 and -2 in human colorectal cancer. Cancer Res. 1995, 55, 3785–3789. [Google Scholar] [PubMed]
- Half, E.; Tang, X.M.; Gwyn, K.; Sahin, A.; Wathen, K.; Sinicrope, F.A. Cyclooxygenase-2 expression in human breast cancers and adjacent ductal carcinoma in situ. Cancer Res. 2002, 62, 1676–1681. [Google Scholar] [PubMed]
- Surowiak, P.; Pawelczyk, K.; Maciejczyk, A.; Pudelko, M.; Kolodziej, J.; Zabel, M.; Murawa, D.; Drag, M.; Gansukh, T.; Dietel, M.; et al. Positive correlation between cyclooxygenase 2 and the expression of ABC transporters in non-small cell lung cancer. Anticancer Res. 2008, 28, 2967–2974. [Google Scholar] [PubMed]
- Qiu, Z.Q.; Qiu, Z.R. Sensitivity of gastric cancer cells to chemotherapy drugs in elderly patients and its correlation with cyclooxygenase-2 expression. Asian Pac. J. Cancer Prev. 2015, 16, 3447–3450. [Google Scholar] [CrossRef] [PubMed]
- Ford, J.M.; Hait, W.N. Pharmacology of drugs that alter multidrug resistance in cancer. Pharmacol. Rev. 1990, 42, 155–199. [Google Scholar]
- Lehne, G. P-glycoprotein as a drug target in the treatment of multidrug resistant cancer. Curr. Drug Targets 2000, 1, 85–99. [Google Scholar] [CrossRef]
- Shukla, S.; Wu, C.P.; Ambudkar, S.V. Development of inhibitors of ATP-binding cassette drug transporters: Present status and challenges. Expert Opin. Drug Metab. Toxicol. 2008, 4, 205–223. [Google Scholar] [CrossRef]
- Ban, T.A. Fifty years chlorpromazine: A historical perspective. Neuropsychiatr. Dis. Treat. 2007, 3, 495–500. [Google Scholar]
- Tsuruo, T.; Iida, H.; Tsukagoshi, S.; Sakurai, Y. Increased accumulation of vincristine and adriamycin in drug-resistant P388 tumor cells following incubation with calcium antagonists and calmodulin inhibitors. Cancer Res. 1982, 42, 4730–4733. [Google Scholar] [PubMed]
- Konya, A.; Andor, A.; Satorhelyi, P.; Nemeth, K.; Kurucz, I. Inhibition of the MDR1 transporter by new phenothiazine derivatives. Biochem. Biophys. Res. Commun. 2006, 346, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Wesolowska, O.; Molnar, J.; Ocsovszki, I.; Michalak, K. Differential effect of phenothiazines on MRP1 and P-glycoprotein activity. In Vivo 2009, 23, 943–947. [Google Scholar] [PubMed]
- Ford, J.A.; Prozialeck, W.C.; Hait, W.N. Structural features determining activity of phenothiazines and related drugs for inhibition of cell growth and reversal of multidrug resistance. Mol. Pharmacol. 1989, 35, 105–115. [Google Scholar]
- Csonka, A.; Spengler, G.; Martins, A.; Ocsovszki, I.; Christensen, J.B.; Hendricks, O.; Kristiansen, J.E.; Amaral, L.; Molnar, J. Effect of thioridazine stereoisomers on the drug accumulation of mouse lymphoma and human prostate cancer cell lines in vitro. In Vivo 2013, 27, 815–820. [Google Scholar] [CrossRef] [PubMed]
- Miller, R.L.; Bukowski, R.M.; Budd, G.T.; Purvis, J.; Weick, J.K.; Shepard, K.; Midha, K.K.; Ganapathi, R. Clinical modulation of doxorubicin resistance by the calmodulin inhibitor trifluoperazine: A Phase I/II trial. J. Clin. Oncol. 1988, 5, 880–888. [Google Scholar] [CrossRef] [PubMed]
- Slater, E.E.; McDonald, J.S. Mechanism of action and biological profile of HMG CoA reductase inhibitors. A new therapeutic alternative. Drugs 1988, 36, 72–82. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Botas, J.; Ferruelo, A.J.; Suarez, Y.; Fernandez, C.; Gomez-Coronado, D.; Lasuncion, M.A. Dose-dependent effects of lovastatin on cell cycle progression. Distinct requirement of cholesterol and non-sterol mevalonate derivatives. Biochim. Biophys. Acta 2001, 1532, 185–194. [Google Scholar] [CrossRef]
- Cafforio, P.; Dammacco, F.; Gernone, A.; Silvestris, F. Statins activate the mitochondrial pathway of apoptosis in human lymphoblasts and myeloma cells. Carcinogenesis 2005, 26, 883–891. [Google Scholar] [CrossRef] [Green Version]
- Cho, S.J.; Kim, J.S.; Kim, J.M.; Lee, J.Y.; Jung, H.C.; Song, I.S. Simvastatin induces apoptosis in human colon cancer cells and in tumor xenografts, and attenuates colitis-associated colon cancer in mice. Int. J. Cancer 2008, 123, 951–957. [Google Scholar] [CrossRef] [Green Version]
- Hoque, A.; Chen, H.; Xu, X.C. Statin induces apoptosis and cell growth arrest in prostate cancer cells. Cancer Epidemiol. Biomark. Prev. 2008, 17, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Palko-Labuz, A.; Sroda-Pomianek, K.; Wesolowska, O.; Kostrzewa-Suslow, E.; Uryga, A.; Michalak, K. MDR reversal and pro-apoptotic effects of statins and statins combined with flavonoids in colon cancer cells. Biomed. Pharmacother. 2019, 109, 511–1522. [Google Scholar] [CrossRef] [PubMed]
- Sroda-Pomianek, K.; Michalak, K.; Palko-Labuz, A.; Dziegiel, P.; Pula, B.; Swiątek, P.; Wesolowska, O. Cytotoxic and multidrug resistance reversal activity of new phenothiazine derivative is strongly enhanced by theobromine, a phytochemical from cocoa. Eur. J. Pharmacol. 2019, 849, 124–134. [Google Scholar] [CrossRef] [PubMed]
- Sroda-Pomianek, K.; Michalak, K.; Swiątek, P.; Pola, A.; Palko-Labuz, A.; Wesolowska, O. Increased lipid peroxidation, apoptosis and selective cytotoxicity in colon cancer cell line LoVo and its doxorubicin-resistant subline LoVo/Dx in the presence of newly synthesized phenothiazine derivatives. Biomed. Pharmacother. 2018, 106, 624–636. [Google Scholar] [CrossRef] [PubMed]
- Wesolowska, O.; Wisniewski, J.; Sroda, K.; Krawczenko, A.; Bielawska-Pohl, A.; Paprocka, M.; Dus, D.; Michalak, K. 8-Prenylnaringenin is an inhibitor of multidrug resistance-associated transporters, P-glycoprotein and MRP1. Eur. J. Pharmacol. 2010, 644, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Sroda-Pomianek, K.; Michalak, K.; Palko-Labuz, A.; Uryga, A.; Szczesniak-Siega, B.; Wesolowska, O. Simvastatin Strongly Augments Proapoptotic, Anti-inflammatory and cytotoxic activity of oxicam derivatives in doxorubicin-resistant colon cancer cells. Anticancer Res. 2019, 39, 727–734. [Google Scholar] [CrossRef] [PubMed]
- Ford, J.M.; Bruggemann, E.P.; Pastan, I.; Gottesman, M.M.; Hait, W.N. Cellular and biochemical characterization of thioxanthenes for reversal of multidrug resistance in human and murine cell lines. Cancer Res. 1990, 50, 1748–1756. [Google Scholar]
- Wesolowska, O.; Wisniewski, J.; Sroda-Pomianek, K.; Bielawska-Pohl, A.; Paprocka, M.; Dus, D.; Duarte, N.; Ferreira, M.J.; Michalak, K. Multidrug resistance reversal and apoptosis induction in human colon cancer cells by some flavonoids present in citrus plants. J. Nat. Prod. 2012, 75, 1896–1902. [Google Scholar] [CrossRef]
- Takacs, D.; Csonka, A.; Horvath, A.; Windt, T.; Gajdacs, M.; Riedl, Z.; Hajos, G.; Amaral, L.; Molnar, J.; Spengler, G. Reversal of ABCB1-related multidrug resistance of colonic adenocarcinoma cells by phenothiazines. Anticancer Res. 2015, 35, 3245–3251. [Google Scholar]
- Goard, C.A.; Mather, R.G.; Vinepal, B.; Clendening, J.W.; Martirosyan, A.; Boutros, P.C.; Sharom, F.J.; Penn, L.Z. Differential interactions between statins and P-glycoprotein: Implications for exploiting statins as anticancer agents. Int. J. Cancer 2010, 127, 2936–2948. [Google Scholar] [CrossRef]
- Sieczkowski, E.; Lehner, C.; Ambros, P.F.; Hohenegger, M. Double impact on p-glycoprotein by statins enhances doxorubicin cytotoxicity in human neuroblastoma cells. Int. J. Cancer 2010, 126, 2025–2035. [Google Scholar] [CrossRef] [PubMed]
- Glodkowska-Mrowka, E.; Mrowka, P.; Basak, G.W.; Niesiobedzka-Krezel, J.; Seferynska, I.; Wlodarski, P.K.; Jakobisiak, M.; Stoklosa, T. Statins inhibit ABCB1 and ABCG2 drug transporter activity in chronic myeloid leukemia cells and potentiate antileukemic effects of imatinib. Exp. Hematol. 2014, 42, 439–447. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, A.C.; Neri, E.A.; Veríssimo-Filho, S.; Rebouças, N.A.; Hirata, R.D.; Yu, A.M. Atorvastatin attenuation of ABCB1 expression is mediated by microRNA miR-491-3p in Caco-2 cells. Eur. J. Pharm. Sci. 2016, 93, 431–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lapczuk-Romanska, J.; Wajda, A.; Pius-Sadowska, E.; Kurzawski, M.; Niedzielski, A.; Machalinski, B.; Drozdzik, M. Effects of simvastatin on nuclear receptors, drug metabolizing enzymes and transporters expression in Human Umbilical Vein Endothelial Cells. Pharmacol. Rep. 2018, 70, 875–880. [Google Scholar] [CrossRef]
- Dormoi, J.; Pascual, A.; Briolant, S.; Amalvict, R.; Charras, S.; Baret, E.; Huyghues des Etages, E.; Feraud, M.; Pradines, B. Proveblue (methylene blue) as an antimalarial agent: In vitro synergy with dihydroartemisinin and atorvastatin. Antimicrob. Agents Chemother. 2012, 56, 3467–3469. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Srivastava, V.K.; Kumar, A. Synthesis and anti-inflamatory activity of some heterocyclic derivatives of phenothiazine. Pharmazie 2005, 60, 18–20. [Google Scholar] [PubMed]
- Kubota, K.; Kurebayashi, H.; Miyachi, H.; Tobe, M.; Onishi, M.; Isobe, Y. Synthesis and structure-activity relationships of phenothiazine carboxylic acids having pyrimidine-dione as novel histamine H(1) antagonists. Bioorg. Med. Chem. Lett. 2009, 19, 2766–2771. [Google Scholar] [CrossRef]
- Stewart, K.D.; Loren, S.; Frey, L.; Otis, E.; Klinghofer, V.; Hulkower, K.I. Discovery of a new cyclooxygenase-2 lead compound through 3-D database searching and combinatorial chemistry. Bioorg. Med. Chem. Lett. 1998, 8, 529–534. [Google Scholar] [CrossRef]
- Matralis, A.N.; Kourounakis, A.P. Design of novel potent antihyperlipidemic agents with antioxidant/anti-inflammatory properties: Exploiting phenothiazine’s strong antioxidant activity. J. Med. Chem. 2014, 57, 2568–2581. [Google Scholar] [CrossRef]
- Habib, A.; Shamseddeen, I.; Nasrallah, M.S.; Antoun, T.A.; Nemer, G.; Bertoglio, J.; Badreddine, R.; Badr, K.F. Modulation of COX2 expression by statins in human monocytic cells. FASEB J. 2007, 21, 1665–1674. [Google Scholar] [CrossRef]
- Haviv, Y.; Avrahami, D.; Ovadia, H.; Ben-Hur, T.; Gabizon, R.; Sharon, R. Induced neuroprotection independently from PrPSc accumulation in a mouse model for prion disease treated with simvastatin. Arch. Neurol. 2008, 65, 762–775. [Google Scholar] [CrossRef] [PubMed]
- Soliman, N.A.; Keshk, W.A.; Rizk, F.H.; Ibrahim, M.A. The possible ameliorative effect of simvastatin versus sulfasalazine on acetic acid induced ulcerative colitis in adult rats. Chem. Biol. Interact. 2019, 298, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Sun, D.; Chen, J.; Guan, N.; Huo, X.; Xi, H. Simvastatin attenuates angiotensin II-induced inflammation and oxidative stress in human mesangial cells. Mol. Med. Rep. 2015, 11, 1246–1251. [Google Scholar] [CrossRef] [PubMed]
- Varga, B.; Csonka, A.; Csonka, A.; Molnar, J.; Amaral, L.; Spengler, G. Possible biological and clinical applications of phenothiazines. Anticancer Res. 2017, 37, 5983–5993. [Google Scholar] [CrossRef] [PubMed]
- Gil-Ad, I.; Shtaif, B.; Levkovitz, Y.; Dayag, M.; Zeldich, E.; Weizman, A. Characterization of phenothiazine-induced apoptosis in neuroblastoma and glioma cell lines: Clinical relevance and possible application for brain-derived tumors. J. Mol. Neurosci. 2004, 22, 189–198. [Google Scholar] [CrossRef]
- Gil-Ad, I.; Shtaif, B.; Levkovitz, Y.; Nordenberg, J.; Taler, M.; Korov, I.; Weizman, A. Phenothiazines induce apoptosis in a B16 mouse melanoma cell line and attenuate in vivo melanoma tumor growth. Oncol. Rep. 2006, 15, 107–112. [Google Scholar] [CrossRef]
- Zong, D.; Haag, P.; Yakymovych, I.; Lewensohn, R.; Viktorsson, K. Chemosensitization by phenothiazines in human lung cancer cells: impaired resolution of γH2AX and increased oxidative stress elicit apoptosis associated with lysosomal expansion and intense vacuolation. Cell Death Dis. 2011, 2, e181. [Google Scholar] [CrossRef]
- Seervi, M.; Rani, A.; Sharma, A.K.; Santhosh Kumar, T.R. ROS mediated ER stress induces Bax-Bak dependent and independent apoptosis in response to thioridazine. Biomed. Pharmacother. 2018, 106, 200–209. [Google Scholar] [CrossRef]
- Wang, B.; Luo, Y.; Zhou, X.; Li, R. Trifluoperazine induces apoptosis through the upregulation of Bax/Bcl-2 and downregulated phosphorylation of AKT in mesangial cells and improves renal function in lupus nephritis mice. Int. J. Mol. Med. 2018, 41, 3278–3286. [Google Scholar] [CrossRef]
- Qi, X.F.; Zheng, L.; Lee, K.J.; Kim, D.H.; Kim, C.S.; Cai, D.Q.; Wu, Z.; Qin, J.W.; Yu, Y.H.; Kim, S.K. HMG-CoA reductase inhibitors induce apoptosis of lymphoma cells by promoting ROS generation and regulating Akt, Erk and p38 signals via suppression of mevalonate pathway. Cell Death Dis. 2013, 4, e518. [Google Scholar] [CrossRef]
- Shen, Y.Y.; Yuan, Y.; Du, Y.Y.; Pan, Y.Y. Molecular mechanism underlying the anticancer effect of simvastatin on MDA-MB-231 human breast cancer cells. Mol. Med. Rep. 2015, 12, 623–630. [Google Scholar] [CrossRef]
- Spampanato, C.; De Maria, S.; Sarnataro, M.; Giordano, E.; Zanfardino, M.; Baiano, S.; Cartenì, M.; Morelli, F. Simvastatin inhibits cancer cell growth by inducing apoptosis correlated to activation of Bax and down-regulation of BCL-2 gene expression. Int. J. Oncol. 2012, 40, 935–941. [Google Scholar] [CrossRef]
- Jakobisiak, M.; Golab, J. Statins can modulate effectiveness of antitumor therapeutic modalities. Med. Res. Rev. 2010, 30, 102–135. [Google Scholar] [CrossRef] [PubMed]
- Gazzerro, P.; Proto, M.C.; Gangemi, G.; Malfitano, A.M.; Ciaglia, E.; Pisanti, S.; Santoro, A.; Laezza, C.; Bifulco, M. Pharmacological actions of statins: A critical appraisal in the management of cancer. Pharmacol. Rev. 2012, 64, 102–146. [Google Scholar] [CrossRef] [PubMed]
- Feleszko, W.; Jalili, A.; Olszewska, D.; Mlynarczuk, I.; Grzela, T.; Giermasz, A.; Jakobisiak, M. Synergistic interaction between highly specific cyclooxygenase-2 inhibitor, MF-tricyclic and lovastatin in murine colorectal cancer cell lines. Oncol. Rep. 2002, 9, 879–885. [Google Scholar] [CrossRef] [PubMed]
- Xiao, H.; Zhang, Q.; Lin, Y.; Reddy, B.S.; Yang, C.S. Combination of atorvastatin and celecoxib synergistically induces cell cycle arrest and apoptosis in colon cancer cells. Int. J. Cancer 2008, 122, 2115–2124. [Google Scholar] [CrossRef] [Green Version]
- Reddy, B.S.; Wang, C.X.; Kong, A.N.; Khor, T.O.; Zheng, X.; Steele, V.E.; Kopelovich, L.; Rao, C.V. Prevention of azoxymethane-induced colon cancer by combination of low doses of atorvastatin, aspirin, and celecoxib in F 344 rats. Cancer Res. 2006, 66, 4542–4546. [Google Scholar] [CrossRef]
- Gasiorowski, K.; Malinka, W.; Swiatek, P.; Jaszczyszyn, A. Antimutagenic activity of new analogues of fluphenazine. Cell. Mol. Biol. Lett. 2003, 8, 927–942. [Google Scholar]
- Drewinko, B.; Romsdahl, M.M.; Yang, L.Y.; Ahearn, M.J.; Trujillo, J.M. Establishment of a Human Carcinoembryonic Antigen-producing Colon Adenocarcinoma Cell Line. Cancer Res. 1976, 36, 467–475. [Google Scholar]
- Grandi, M.; Geroni, C.; Giuliani, F.C. Isolation and characterization of a human colon adenocarcinoma cell line resistant to doxorubicin. Br. J. Cancer 1986, 54, 515–518. [Google Scholar] [CrossRef] [Green Version]
- Skehan, P.; Storeng, R.; Scudiero, D.; Monks, A.; McMahon, J.; Vistica, D.; Warren, J.T.; Bokesch, H.; Kenney, S.; Boyd, M.R. New colorimetric cytotoxicity assay for anticancer-drug screening. J. Natl. Cancer Inst. 1990, 82, 1107–1112. [Google Scholar] [CrossRef] [PubMed]
- Palko-Labuz, A.; Sroda-Pomianek, K.; Uryga, A.; Kostrzewa-Suslow, E.; Michalak, K. Anticancer activity of baicalein and luteolin studied in colorectal adenocarcinoma LoVo cells and in drug-resistant LoVo/Dx cells. Biomed. Pharmacother. 2017, 88, 232–241. [Google Scholar] [CrossRef] [PubMed]
- Chou, T.C.; Martin, N. CompuSyn for Drug Combinations: PC Software and User’s Guide: A Computer Program for Quantitation of Synergism and Antagonism in Drug Combinations, and the Determination of IC50 and ED50 and LD50 Values; ComboSyn Inc.: Paramus, NJ, USA, 2005. [Google Scholar]
- Riganti, C.; Miraglia, E.; Viarisio, D.; Costamagna, C.; Pescarmona, G.; Ghigo, D.; Bosia, A. Nitric Oxide reverts the resistance to doxorubicin in human colon cancer cells by inhibiting the drug efflux. Cancer Res. 2005, 65, 516–525. [Google Scholar] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Concentration [M] | Ratio | Combination Index | ||
|---|---|---|---|---|
| Dox | MAE-TPR | SIM | (CI) | |
| 10 | 2.5 | 4:1 | 0.2261 | |
| 10 | 2.5 | 4:1 | 0.4092 | |
| 10 | 2.5 | 2.5 | 4:1:1 | 0.0748 |
| Dox | APh-Flu | SIM | ||
| 10 | 2.5 | 4:1 | 1.0101 | |
| 10 | 2.5 | 2.5 | 4:1:1 | 0.2116 |
| Dox | FLU | SIM | ||
| 10 | 2.5 | 4:1 | 0.8261 | |
| 10 | 2.5 | 2.5 | 4:1:1 | 0.348 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Środa-Pomianek, K.; Michalak, K.; Palko-Łabuz, A.; Uryga, A.; Świątek, P.; Majkowski, M.; Wesołowska, O. The Combined Use of Phenothiazines and Statins Strongly Affects Doxorubicin-Resistance, Apoptosis, and Cox-2 Activity in Colon Cancer Cells. Int. J. Mol. Sci. 2019, 20, 955. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20040955
Środa-Pomianek K, Michalak K, Palko-Łabuz A, Uryga A, Świątek P, Majkowski M, Wesołowska O. The Combined Use of Phenothiazines and Statins Strongly Affects Doxorubicin-Resistance, Apoptosis, and Cox-2 Activity in Colon Cancer Cells. International Journal of Molecular Sciences. 2019; 20(4):955. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20040955
Chicago/Turabian StyleŚroda-Pomianek, Kamila, Krystyna Michalak, Anna Palko-Łabuz, Anna Uryga, Piotr Świątek, Michał Majkowski, and Olga Wesołowska. 2019. "The Combined Use of Phenothiazines and Statins Strongly Affects Doxorubicin-Resistance, Apoptosis, and Cox-2 Activity in Colon Cancer Cells" International Journal of Molecular Sciences 20, no. 4: 955. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20040955