FTIR and Raman Spectroscopy-Based Biochemical Profiling Reflects Genomic Diversity of Clinical Candida Isolates That May Be Useful for Diagnosis and Targeted Therapy of Candidiasis

Abstract
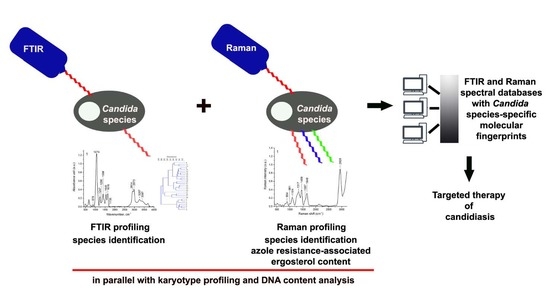
:
1. Introduction
2. Results and Discussion
2.1. Karyotype Profiling and DNA Content Analysis
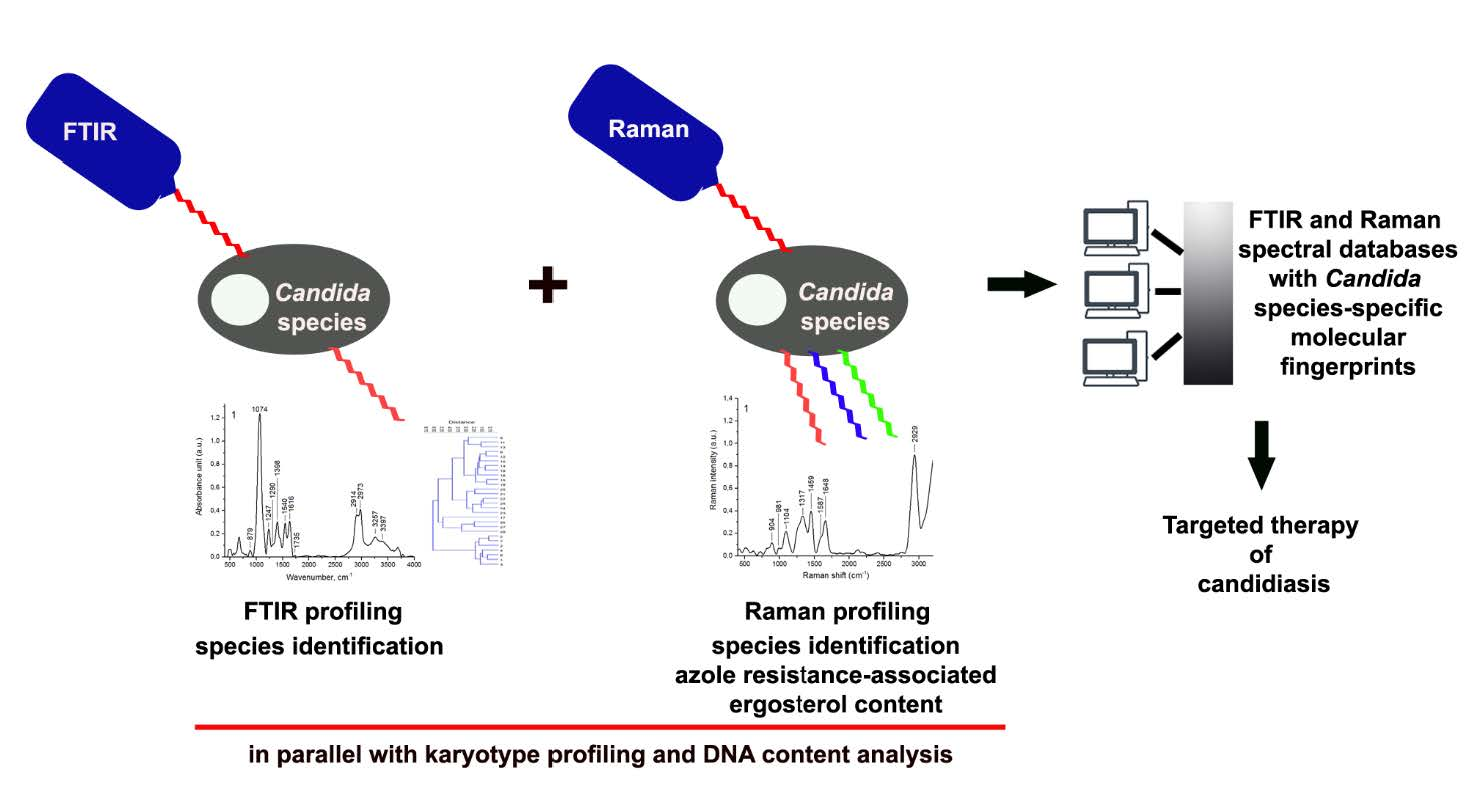
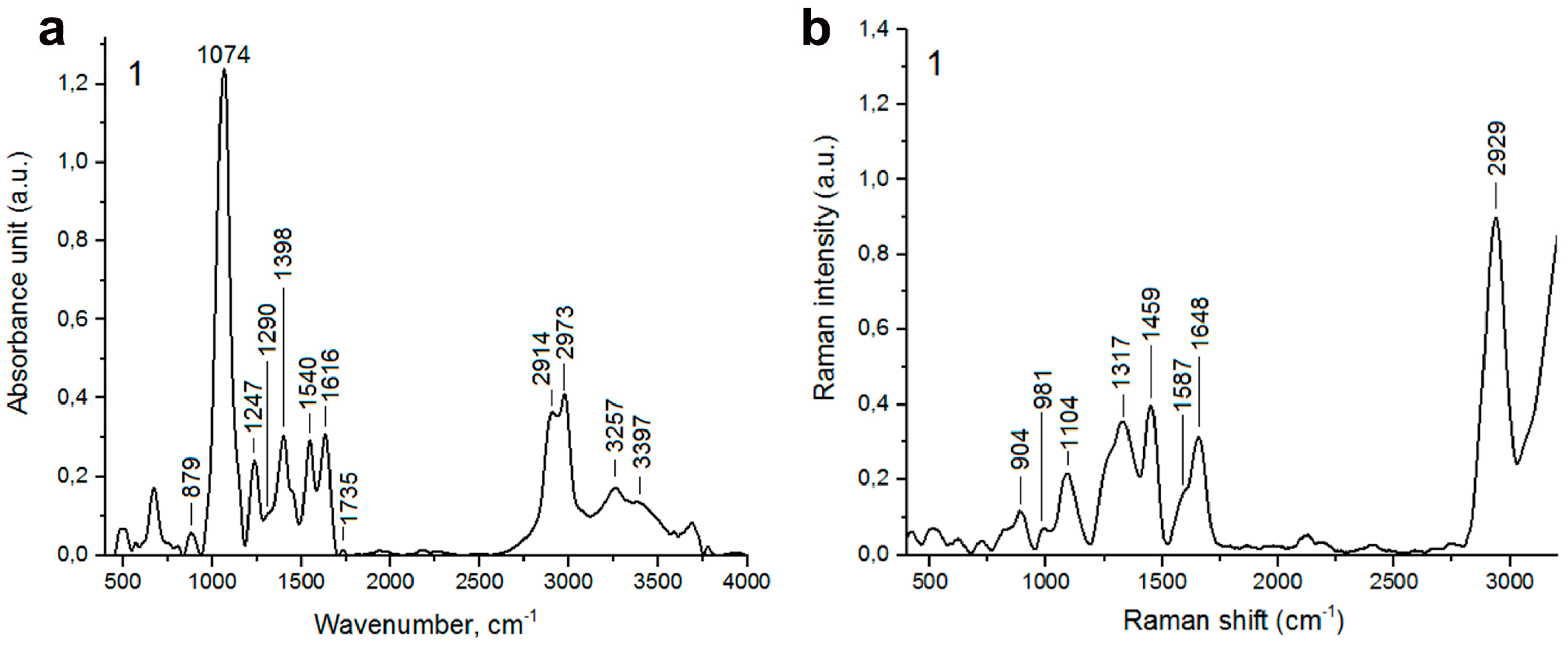
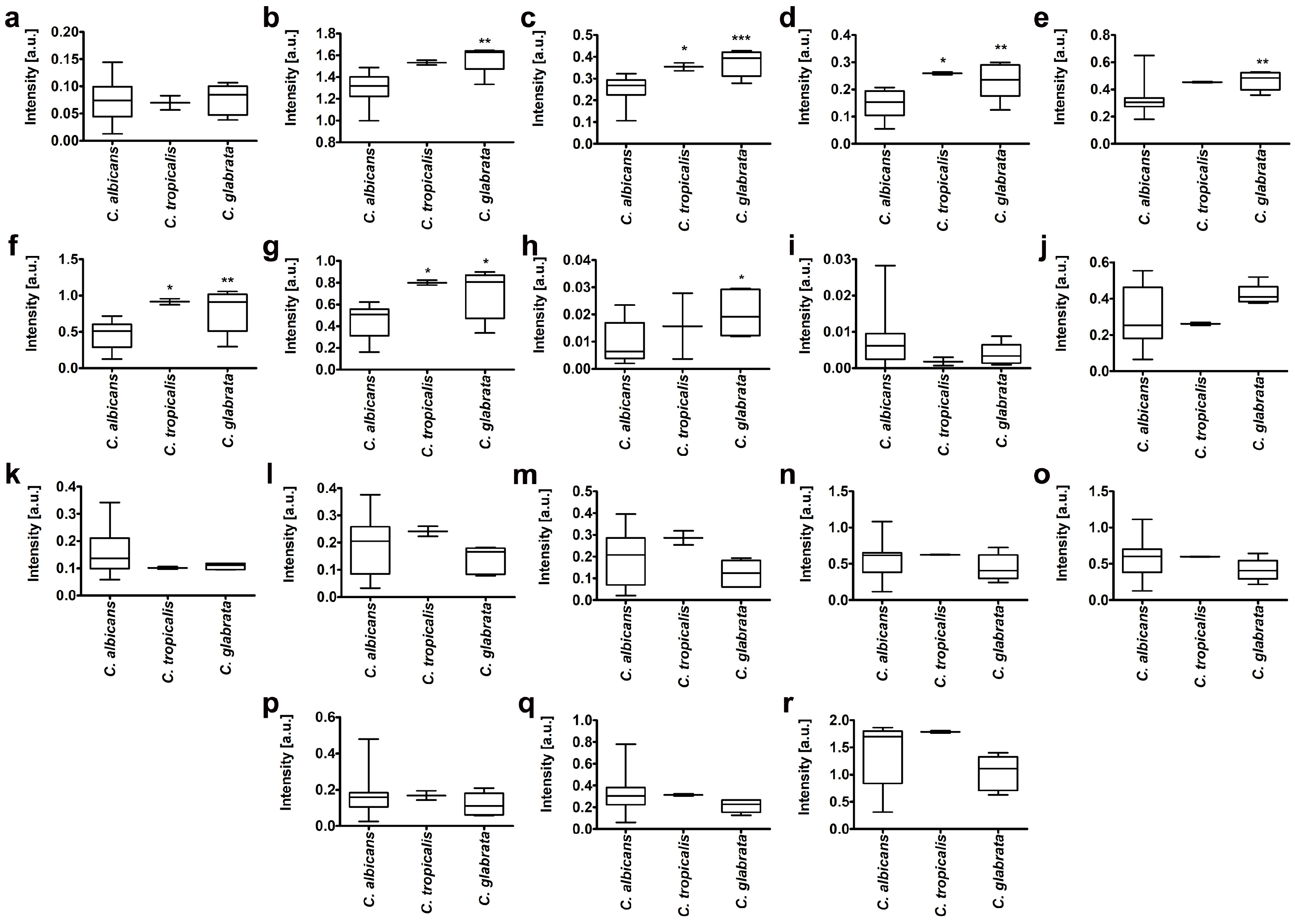
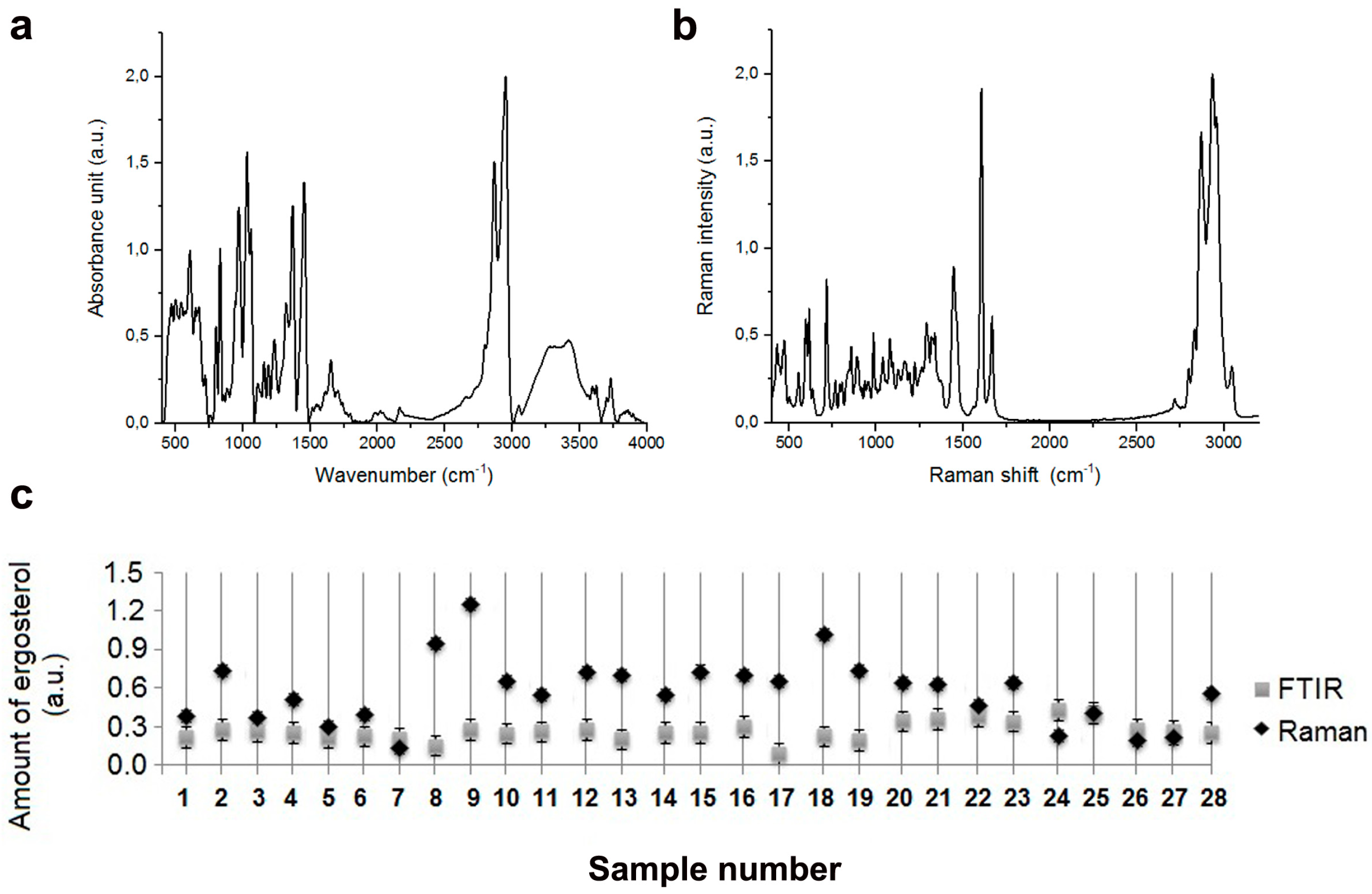
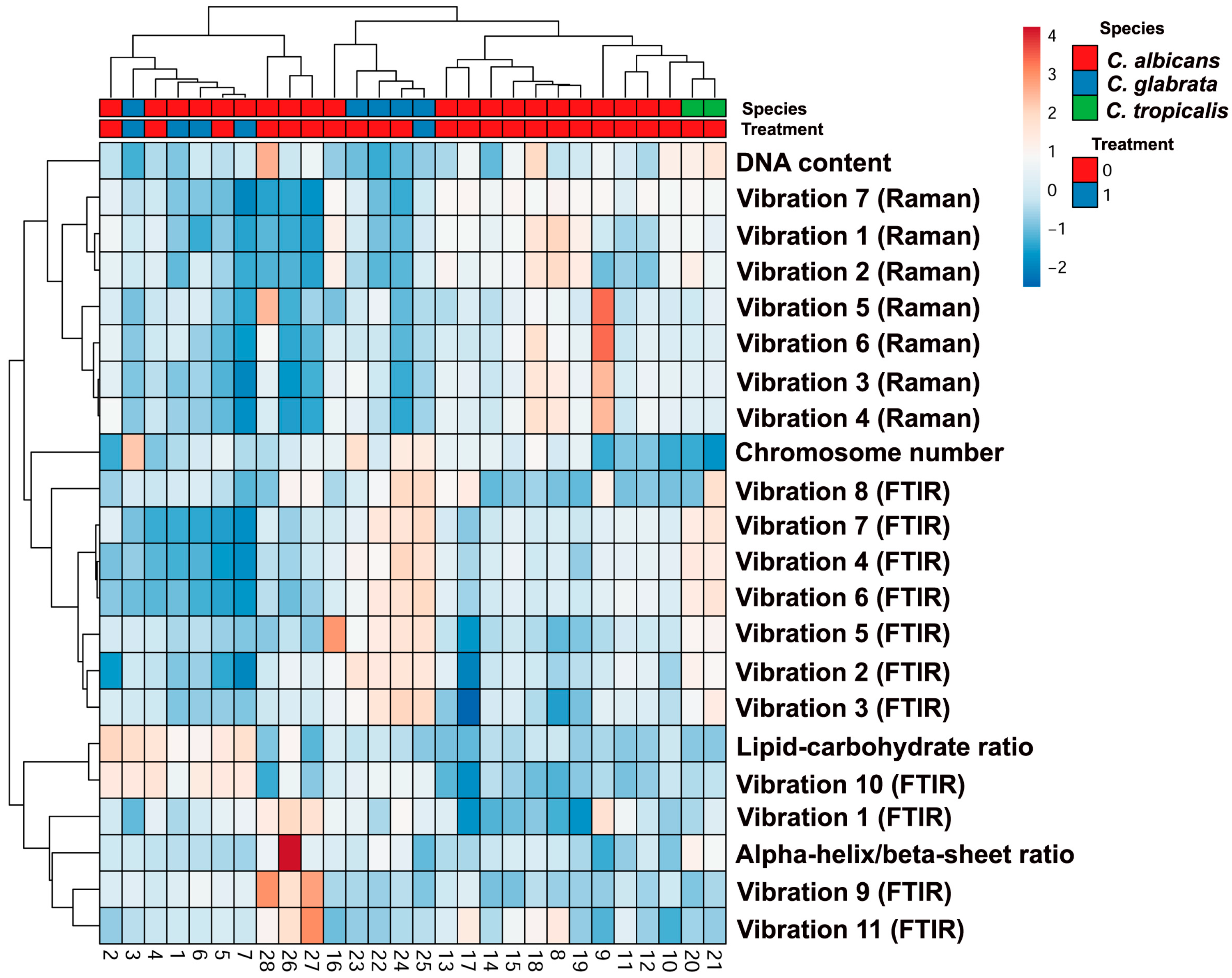
2.2. Biochemical Features Reflect Genomic Diversity and Plasticity of Candida Cells
3. Materials and Methods
3.1. Ethics Statement
3.2. Clinical Specimens and Reference Strains
3.3. Culture Conditions and Species Identification
3.4. Preparation of Agarose-Embedded Yeast DNA
3.5. Pulsed-Field Gel Electrophoresis (PFGE)
3.6. DNA Content Analysis
3.7. Glycogen Storage Assay
3.8. FTIR Spectroscopy
3.9. Deconvolution of Amide I region (1600–1700 cm−1)
3.10. Raman Spectroscopy
3.11. Lipid-Carbohydrate Ratio
3.12. Ergosterol Content
3.13. Multivariate Data Analysis
3.14. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Calderone, R.A. Introduction and historical perspectives. In Candida and Candidiasis; Calderone, R.A., Ed.; ASM Press: Washington, DC, USA, 2002; pp. 15–25. [Google Scholar]
- Antinori, S.; Milazzo, L.; Sollima, S.; Galli, M.; Corbellino, M. Candidemia and invasive candidiasis in adults: A narrative review. Eur. J. Intern. Med. 2016, 34, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Wilson, L.S.; Reyes, C.M.; Stolpman, M.; Speckman, J.; Allen, K.; Beney, J. The direct cost and incidence of systemic fungal infections. Value Health 2002, 5, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Pappas, P.G.; Lionakis, M.S.; Arendrup, M.C.; Ostrosky-Zeichner, L.; Kullberg, B.J. Invasive candidiasis. Nat. Rev. Dis. Primers 2018, 4, 18026. [Google Scholar] [CrossRef] [PubMed]
- Diekema, D.; Arbefeville, S.; Boyken, L.; Kroeger, J.; Pfaller, M. The changing epidemiology of healthcare-associated candidemia over three decades. Diagn Microbiol. Infect. Dis. 2012, 73, 45–48. [Google Scholar] [CrossRef] [PubMed]
- Kauffman, C.A.; Vazquez, J.A.; Sobel, J.D.; Gallis, H.A.; McKinsey, D.S.; Karchmer, A.W.; Sugar, A.M.; Sharkey, P.K.; Wise, G.J.; Mangi, R.; et al. Prospective multicenter surveillance study of funguria in hospitalized patients. The National Institute for Allergy and Infectious Diseases (NIAID) Mycoses Study Group. Clin. Infect. Dis. 2000, 30, 14–18. [Google Scholar] [CrossRef] [PubMed]
- Ruan, S.Y.; Hsueh, P.R. Invasive candidiasis: An overview from Taiwan. J. Formos. Med. Assoc. 2009, 108, 443–451. [Google Scholar] [CrossRef]
- Liguori, G.; Di Onofrio, V.; Lucariello, A.; Galle, F.; Signoriello, G.; Colella, G.; D’Amora, M.; Rossano, F. Oral candidiasis: A comparison between conventional methods and multiplex polymerase chain reaction for species identification. Oral Microbiol. Immunol. 2009, 24, 76–78. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, G.M.; Elizondo, M.; Ayala, J. Trends in species distribution and susceptibility of bloodstream isolates of Candida collected in Monterrey, Mexico, to seven antifungal agents: Results of a 3-year (2004 to 2007) surveillance study. J. Clin. Microbiol. 2008, 46, 2902–2905. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Gaitan, A.; Moret, A.M.; Tasias-Pitarch, M.; Aleixandre-Lopez, A.I.; Martinez-Morel, H.; Calabuig, E.; Salavert-Lleti, M.; Ramirez, P.; Lopez-Hontangas, J.L.; Hagen, F.; et al. An outbreak due to Candida auris with prolonged colonisation and candidaemia in a tertiary care European hospital. Mycoses 2018, 61, 498–505. [Google Scholar] [CrossRef] [PubMed]
- Silva, S.; Negri, M.; Henriques, M.; Oliveira, R.; Williams, D.W.; Azeredo, J. Candida glabrata, Candida parapsilosis and Candida tropicalis: Biology, epidemiology, pathogenicity and antifungal resistance. FEMS Microbiol. Rev. 2012, 36, 288–305. [Google Scholar] [CrossRef] [PubMed]
- Alexander, B.D.; Ashley, E.D.; Reller, L.B.; Reed, S.D. Cost savings with implementation of PNA FISH testing for identification of Candida albicans in blood cultures. Diagn Microbiol. Infect. Dis. 2006, 54, 277–282. [Google Scholar] [CrossRef] [PubMed]
- Ellepola, A.N.; Morrison, C.J. Laboratory diagnosis of invasive candidiasis. J. Microbiol. 2005, 43, 65–84. [Google Scholar] [PubMed]
- Dijck, P.V.; Sjollema, J.; Cammue, B.P.A.; Lagrou, K.; Berman, J.; d’Enfert, C.; Andes, D.R.; Arendrup, M.C.; Brakhage, A.A.; Calderone, R.; et al. Methodologies for in vitro and in vivo evaluation of efficacy of antifungal and antibiofilm agents and surface coatings against fungal biofilms. Microb. Cell 2018, 5, 300–326. [Google Scholar] [CrossRef] [PubMed]
- Selmecki, A.; Forche, A.; Berman, J. Genomic plasticity of the human fungal pathogen Candida albicans. Eukaryot. Cell 2010, 9, 991–1008. [Google Scholar] [CrossRef] [PubMed]
- Berman, J. Ploidy plasticity: A rapid and reversible strategy for adaptation to stress. FEMS Yeast Res. 2016, 16. [Google Scholar] [CrossRef] [PubMed]
- Todd, R.T.; Forche, A.; Selmecki, A. Ploidy Variation in Fungi: Polyploidy, Aneuploidy, and Genome Evolution. Microbiol. Spectr. 2017, 5. [Google Scholar] [CrossRef]
- Magee, B.B.; Magee, P.T. Electrophoretic karyotypes and chromosome numbers in Candida species. J. Gen. Microbiol. 1987, 133, 425–430. [Google Scholar] [PubMed]
- Rustchenko-Bulgac, E.P. Variations of Candida albicans electrophoretic karyotypes. J. Bacteriol. 1991, 173, 6586–6596. [Google Scholar] [CrossRef] [PubMed]
- Iwaguchi, S.; Homma, M.; Tanaka, K. Variation in the electrophoretic karyotype analysed by the assignment of DNA probes in Candida albicans. J. Gen. Microbiol. 1990, 136, 2433–2442. [Google Scholar] [CrossRef] [PubMed]
- Pfaller, M.A.; Rhine-Chalberg, J.; Redding, S.W.; Smith, J.; Farinacci, G.; Fothergill, A.W.; Rinaldi, M.G. Variations in fluconazole susceptibility and electrophoretic karyotype among oral isolates of Candida albicans from patients with AIDS and oral candidiasis. J. Clin. Microbiol. 1994, 32, 59–64. [Google Scholar] [PubMed]
- Perepnikhatka, V.; Fischer, F.J.; Niimi, M.; Baker, R.A.; Cannon, R.D.; Wang, Y.K.; Sherman, F.; Rustchenko, E. Specific chromosome alterations in fluconazole-resistant mutants of Candida albicans. J. Bacteriol. 1999, 181, 4041–4049. [Google Scholar] [PubMed]
- Hickman, M.A.; Zeng, G.; Forche, A.; Hirakawa, M.P.; Abbey, D.; Harrison, B.D.; Wang, Y.M.; Su, C.H.; Bennett, R.J.; Wang, Y.; et al. The ‘obligate diploid’ Candida albicans forms mating-competent haploids. Nature 2013, 494, 55–59. [Google Scholar] [CrossRef] [PubMed]
- Selmecki, A.; Bergmann, S.; Berman, J. Comparative genome hybridization reveals widespread aneuploidy in Candida albicans laboratory strains. Mol. Microbiol. 2005, 55, 1553–1565. [Google Scholar] [CrossRef] [PubMed]
- Harrison, B.D.; Hashemi, J.; Bibi, M.; Pulver, R.; Bavli, D.; Nahmias, Y.; Wellington, M.; Sapiro, G.; Berman, J. A tetraploid intermediate precedes aneuploid formation in yeasts exposed to fluconazole. PLoS Biol. 2014, 12, e1001815. [Google Scholar] [CrossRef] [PubMed]
- Bader, O.; Weig, M.; Taverne-Ghadwal, L.; Lugert, R.; Gross, U.; Kuhns, M. Improved clinical laboratory identification of human pathogenic yeasts by matrix-assisted laser desorption ionization time-of-flight mass spectrometry. Clin. Microbiol. Infect. 2011, 17, 1359–1365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, K.E.; Ellis, B.C.; Lee, R.; Stamper, P.D.; Zhang, S.X.; Carroll, K.C. Prospective evaluation of a matrix-assisted laser desorption ionization-time of flight mass spectrometry system in a hospital clinical microbiology laboratory for identification of bacteria and yeasts: A bench-by-bench study for assessing the impact on time to identification and cost-effectiveness. J. Clin. Microbiol. 2012, 50, 3301–3308. [Google Scholar] [PubMed]
- Sockalingum, G.D.; Sandt, C.; Toubas, D.; Gomez, J.; Pina, P.; Beguinot, I.; Witthuhn, F.; Aubert, D.; Allouch, P.; Pinon, J.M.; et al. FTIR characterization of Candida species: A study on some reference strains and pathogenic C. albicans isolates from HIV+ patients. Vib. Spectrosc. 2002, 28, 137–146. [Google Scholar] [CrossRef]
- Essendoubi, M.; Toubas, D.; Bouzaggou, M.; Pinon, J.M.; Manfait, M.; Sockalingum, G.D. Rapid identification of Candida species by FT-IR microspectroscopy. Biochim. Biophys. Acta 2005, 1724, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Colabella, C.; Corte, L.; Roscini, L.; Shapaval, V.; Kohler, A.; Tafintseva, V.; Tascini, C.; Cardinali, G. Merging FT-IR and NGS for simultaneous phenotypic and genotypic identification of pathogenic Candida species. PLoS ONE 2017, 12, e0188104. [Google Scholar] [CrossRef] [PubMed]
- Ibelings, M.S.; Maquelin, K.; Endtz, H.P.; Bruining, H.A.; Puppels, G.J. Rapid identification of Candida spp. in peritonitis patients by Raman spectroscopy. Clin. Microbiol. Infect. 2005, 11, 353–358. [Google Scholar] [CrossRef] [PubMed]
- Maquelin, K.; Choo-Smith, L.P.; Endtz, H.P.; Bruining, H.A.; Puppels, G.J. Rapid identification of Candida species by confocal Raman microspectroscopy. J. Clin. Microbiol. 2002, 40, 594–600. [Google Scholar] [CrossRef] [PubMed]
- Silge, A.; Heinke, R.; Bocklitz, T.; Wiegand, C.; Hipler, U.C.; Rosch, P.; Popp, J. The application of UV resonance Raman spectroscopy for the differentiation of clinically relevant Candida species. Anal. Bioanal. Chem. 2018, 410, 5839–5847. [Google Scholar] [CrossRef] [PubMed]
- Sudbery, P.E. Growth of Candida albicans hyphae. Nat. Rev. Microbiol. 2011, 9, 737–748. [Google Scholar] [CrossRef] [PubMed]
- Larone, D.H. Medically Important Fungi: A Guide to Identification; ASM Press: Washington, DC, USA, 2011. [Google Scholar]
- Chibana, H.; Beckerman, J.L.; Magee, P.T. Fine-resolution physical mapping of genomic diversity in Candida albicans. Genome Res. 2000, 10, 1865–1877. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, K.M.; Kokosar, J.; Guo, X.; Gu, Z.; Ishchuk, O.P.; Piskur, J. Genome structure and dynamics of the yeast pathogen Candida glabrata. FEMS Yeast Res. 2014, 14, 529–535. [Google Scholar] [CrossRef] [PubMed]
- Dujon, B.; Sherman, D.; Fischer, G.; Durrens, P.; Casaregola, S.; Lafontaine, I.; De Montigny, J.; Marck, C.; Neuveglise, C.; Talla, E.; et al. Genome evolution in yeasts. Nature 2004, 430, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Healey, K.R.; Jimenez Ortigosa, C.; Shor, E.; Perlin, D.S. Genetic Drivers of Multidrug Resistance in Candida glabrata. Front. Microbiol. 2016, 7, 1995. [Google Scholar] [CrossRef] [PubMed]
- Klempp-Selb, B.; Rimek, D.; Kappe, R. Karyotyping of Candida albicans and Candida glabrata from patients with Candida sepsis. Mycoses 2000, 43, 159–163. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.H.; Chae, M.J.; Song, J.W.; Jung, S.I.; Cho, D.; Kee, S.J.; Kim, S.H.; Shin, M.G.; Suh, S.P.; Ryang, D.W. Changes in karyotype and azole susceptibility of sequential bloodstream isolates from patients with Candida glabrata candidemia. J. Clin. Microbiol. 2007, 45, 2385–2391. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.Y.; Chen, Y.C.; Lo, H.J.; Chen, K.W.; Li, S.Y. Assessment of Candida glabrata strain relatedness by pulsed-field gel electrophoresis and multilocus sequence typing. J. Clin. Microbiol. 2007, 45, 2452–2459. [Google Scholar] [CrossRef] [PubMed]
- Polakova, S.; Blume, C.; Zarate, J.A.; Mentel, M.; Jorck-Ramberg, D.; Stenderup, J.; Piskur, J. Formation of new chromosomes as a virulence mechanism in yeast Candida glabrata. Proc. Natl. Acad. Sci. USA 2009, 106, 2688–2693. [Google Scholar] [CrossRef] [PubMed]
- Wertheimer, N.B.; Stone, N.; Berman, J. Ploidy dynamics and evolvability in fungi. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2016, 371. [Google Scholar] [CrossRef] [PubMed]
- Hickman, M.A.; Paulson, C.; Dudley, A.; Berman, J. Parasexual Ploidy Reduction Drives Population Heterogeneity Through Random and Transient Aneuploidy in Candida albicans. Genetics 2015, 200, 781–794. [Google Scholar] [CrossRef] [PubMed]
- Kaur, R.; Domergue, R.; Zupancic, M.L.; Cormack, B.P. A yeast by any other name: Candida glabrata and its interaction with the host. Curr. Opin. Microbiol. 2005, 8, 378–384. [Google Scholar] [CrossRef] [PubMed]
- Morrow, C.A.; Fraser, J.A. Ploidy variation as an adaptive mechanism in human pathogenic fungi. Semin. Cell Dev. Biol. 2013, 24, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Selmecki, A.; Forche, A.; Berman, J. Aneuploidy and isochromosome formation in drug-resistant Candida albicans. Science 2006, 313, 367–370. [Google Scholar] [CrossRef] [PubMed]
- Selmecki, A.; Gerami-Nejad, M.; Paulson, C.; Forche, A.; Berman, J. An isochromosome confers drug resistance in vivo by amplification of two genes, ERG11 and TAC1. Mol. Microbiol. 2008, 68, 624–641. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Yang, F.; Li, D.; Zhou, M.; Wang, X.; Xu, Q.; Zhang, Y.; Yan, L.; Jiang, Y. Trisomy of chromosome R confers resistance to triazoles in Candida albicans. Med. Mycol. 2015, 53, 302–309. [Google Scholar] [CrossRef] [PubMed]
- Anderson, M.Z.; Saha, A.; Haseeb, A.; Bennett, R.J. A chromosome 4 trisomy contributes to increased fluconazole resistance in a clinical isolate of Candida albicans. Microbiology 2017, 163, 856–865. [Google Scholar] [CrossRef] [PubMed]
- Harari, Y.; Ram, Y.; Rappoport, N.; Hadany, L.; Kupiec, M. Spontaneous Changes in Ploidy Are Common in Yeast. Curr. Biol. 2018, 28, 825–835. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.O.; Sherlock, G.; Petrov, D.A. Whole Genome Analysis of 132 Clinical Saccharomyces cerevisiae Strains Reveals Extensive Ploidy Variation. G3 (Bethesda) 2016, 6, 2421–2434. [Google Scholar] [CrossRef] [PubMed]
- Naumann, D.; Helm, D.; Labischinski, H. Microbiological characterizations by FT-IR spectroscopy. Nature 1991, 351, 81–82. [Google Scholar] [CrossRef] [PubMed]
- Maquelin, K.; Kirschner, C.; Choo-Smith, L.P.; van den Braak, N.; Endtz, H.P.; Naumann, D.; Puppels, G.J. Identification of medically relevant microorganisms by vibrational spectroscopy. J. Microbiol. Methods 2002, 51, 255–271. [Google Scholar] [CrossRef]
- Butler, H.J.; Ashton, L.; Bird, B.; Cinque, G.; Curtis, K.; Dorney, J.; Esmonde-White, K.; Fullwood, N.J.; Gardner, B.; Martin-Hirsch, P.L.; et al. Using Raman spectroscopy to characterize biological materials. Nat. Protoc. 2016, 11, 664–687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maquelin, K.; Kirschner, C.; Choo-Smith, L.P.; Ngo-Thi, N.A.; van Vreeswijk, T.; Stammler, M.; Endtz, H.P.; Bruining, H.A.; Naumann, D.; Puppels, G.J. Prospective study of the performance of vibrational spectroscopies for rapid identification of bacterial and fungal pathogens recovered from blood cultures. J. Clin. Microbiol. 2003, 41, 324–329. [Google Scholar] [CrossRef] [PubMed]
- Galichet, A.; Sockalingum, G.D.; Belarbi, A.; Manfait, M. FTIR spectroscopic analysis of Saccharomyces cerevisiae cell walls: Study of an anomalous strain exhibiting a pink-colored cell phenotype. FEMS Microbiol. Lett. 2001, 197, 179–186. [Google Scholar] [CrossRef] [PubMed]
- Burattini, E.; Cavagna, M.; Dell’Anna, R.; Campeggi, F.M.; Monti, F.; Rossi, F.; Torriani, S. A FTIR microspectroscopy study of autolysis in cells of the wine yeast Saccharomyces cerevisiae. Vib. Spectrosc. 2008, 47, 139–147. [Google Scholar] [CrossRef]
- Mihoubi, W.; Sahli, E.; Gargouri, A.; Amiel, C. FTIR spectroscopy of whole cells for the monitoring of yeast apoptosis mediated by p53 over-expression and its suppression by Nigella sativa extracts. PLoS ONE 2017, 12, e0180680. [Google Scholar] [CrossRef] [PubMed]
- Wrobel, T.P.; Mateuszuk, L.; Chlopicki, S.; Malek, K.; Baranska, M. Imaging of lipids in atherosclerotic lesion in aorta from ApoE/LDLR-/- mice by FT-IR spectroscopy and Hierarchical Cluster Analysis. Analyst 2011, 136, 5247–5255. [Google Scholar] [CrossRef] [PubMed]
- Barth, A.; Zscherp, C. What vibrations tell us about proteins. Q. Rev. Biophys. 2002, 35, 369–430. [Google Scholar] [CrossRef] [PubMed]
- Mikoliunaite, L.; Rodriguez, R.D.; Sheremet, E.; Kolchuzhin, V.; Mehner, J.; Ramanavicius, A.; Zahn, D.R. The substrate matters in the Raman spectroscopy analysis of cells. Sci. Rep. 2015, 5, 13150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harz, M.; Rosch, P.; Popp, J. Vibrational spectroscopy-a powerful tool for the rapid identification of microbial cells at the single-cell level. Cytometry A 2009, 75, 104–113. [Google Scholar] [CrossRef] [PubMed]
- Arthington-Skaggs, B.A.; Jradi, H.; Desai, T.; Morrison, C.J. Quantitation of ergosterol content: Novel method for determination of fluconazole susceptibility of Candida albicans. J. Clin. Microbiol. 1999, 37, 3332–3337. [Google Scholar] [PubMed]
- Li, Q.Q.; Tsai, H.F.; Mandal, A.; Walker, B.A.; Noble, J.A.; Fukuda, Y.; Bennett, J.E. Sterol uptake and sterol biosynthesis act coordinately to mediate antifungal resistance in Candida glabrata under azole and hypoxic stress. Mol. Med. Rep. 2018, 17, 6585–6597. [Google Scholar] [PubMed]
- Chiu, L.D.; Hullin-Matsuda, F.; Kobayashi, T.; Torii, H.; Hamaguchi, H.O. On the origin of the 1602 cm−1 Raman band of yeasts; contribution of ergosterol. J. Biophotonics 2012, 5, 724–728. [Google Scholar] [CrossRef] [PubMed]
- Deregowska, A.; Skoneczny, M.; Adamczyk, J.; Kwiatkowska, A.; Rawska, E.; Skoneczna, A.; Lewinska, A.; Wnuk, M. Genome-wide array-CGH analysis reveals YRF1 gene copy number variation that modulates genetic stability in distillery yeasts. Oncotarget 2015, 6, 30650–30663. [Google Scholar] [CrossRef] [PubMed]
- Chester, V.E. Heritable glycogen-storage deficiency in yeast and its induction by ultra-violet light. J. Gen. Microbiol. 1968, 51, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Chiu, L.D.; Ho, S.H.; Shimada, R.; Ren, N.Q.; Ozawa, T. Rapid in vivo lipid/carbohydrate quantification of single microalgal cell by Raman spectral imaging to reveal salinity-induced starch-to-lipid shift. Biotechnol. Biofuels 2017, 10, 9. [Google Scholar] [CrossRef] [PubMed]
- Metsalu, T.; Vilo, J. ClustVis: A web tool for visualizing clustering of multivariate data using Principal Component Analysis and heatmap. Nucleic Acids Res. 2015, 43, W566–W570. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate | Isolate Identification (API® Candida Biochemical Test) | Isolate Identification (DNA Sequencing) | GenBank Accession Number | Sequence Identity (%)/Query Cover (%) | Isolation Site | Sex | Treatment |
|---|---|---|---|---|---|---|---|
| 1. | Candida albicans 4316 | Candida albicans M366B | KP675609.1 | 99/98 | bronchoalveolar lavage | female | voriconazole |
| 2. | Candida albicans 154 | Candida albicans NG76 | MH019247.1 | 99/96 | vagina | female | |
| 3. | Candida albicans 4200 | Candida glabrata | AM492797.1 | 98/98 | sputum | male | fluconazole |
| 4. | Candida albicans 4248 | Candida albicans M215B | KP675383.1 | 99/98 | urine | female | |
| 5. | Candida albicans 4310 | Candida albicans CGP41 | MF276783.1 | 99/100 | sputum | female | |
| 6. | Candida albicans M/529 | Candida albicans M179A | KP675353.1 | 99/100 | bronchoalveolar lavage | male | fluconazole |
| 7. | Candida albicans 532 | Candida albicans M221B | KP675393.1 | 99/98 | bronchoalveolar lavage | male | |
| 8. | Candida albicans 4331 | Candida albicans M363B | KP675603.1 | 99/98 | inoculation from urine | male | fluconazole |
| 9. | Candida albicans 4324 | Candida albicans M366B | KP675609.1 | 99/97 | sputum | female | |
| 10. | Candida albicans 534 | Candida sp. | KY996547.1 | 99/97 | bronchoalveolar lavage | male | |
| 11. | Candida albicans 152 | Candida albicans M357B | KP675591.1 | 99/99 | vagina | female | |
| 12. | Candida albicans 153 | Candida albicans H291B | KP675000.1 | 89/97 | vagina | female | |
| 13. | Candida albicans 521 | Candida albicans 125A | KP765018.1 | 96/97 | pharynx | male | |
| 14. | Candida albicans 4369 | Candida albicans H296B | KP675010.1 | 100/99 | wound | female | |
| 15. | Candida albicans 556 | Candida albicans n96b | KP675680.1 | 100/96 | bronchoalveolar lavage | female | |
| 16. | Candida albicans 563 | Candida albicans M349A | KP675580.1 | 100/97 | bronchoalveolar lavage | male | |
| 17. | Candida albicans 4335 | Candida albicans H257B | KP674940.1 | 88/97 | bronchoalveolar lavage | female | |
| 18. | Candida albicans 539 | Candida albicans H294A | KP675005.1 | 100/98 | bronchoalveolar lavage | male | |
| 19. | Candida albicans 7363 | Candida albicans M215B | KP675383.1 | 100/98 | bronchoalveolar lavage | female | |
| 20. | Candida tropicalis 4403 | Candida tropicalis H260C | KP674945.1 | 99/98 | sputum | male | |
| 21. | Candida tropicalis 4114 | Candida tropicalis CTR817 | KX664669.1 | 99/98 | bronchoalveolar lavage | male | |
| 22. | Candida glabrata 520 | Candida glabrata IFM 64525 | LC317501.1 | 99/97 | bronchoalveolar lavage | male | |
| 23. | Candida glabrata 4570 | Candida glabrata M9 | LC389275.1 | 99/97 | sputum | male | |
| 24. | Candida glabrata 144 | Candida glabrata H160 | MF187244.1 | 99/97 | vagina | female | |
| 25. | Candida glabrata 4246 | Candida glabrata H160 | LC389261.1 | 99/97 | urine | female | fluconazole |
| 26. | Candida albicans 302 | Candida albicans H194B | KP674872.1 | 100/98 | reference strain—n | ||
| 27. | Candida albicans SC5314 | Candida albicans SC5314 | CP017630.1 | 99/98 | reference strain—2n | ||
| 28. | Candida albicans T15 | Candida albicans B280A | KP674535.1 | 99/97 | reference strain—4n |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Potocki, L.; Depciuch, J.; Kuna, E.; Worek, M.; Lewinska, A.; Wnuk, M. FTIR and Raman Spectroscopy-Based Biochemical Profiling Reflects Genomic Diversity of Clinical Candida Isolates That May Be Useful for Diagnosis and Targeted Therapy of Candidiasis. Int. J. Mol. Sci. 2019, 20, 988. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20040988
Potocki L, Depciuch J, Kuna E, Worek M, Lewinska A, Wnuk M. FTIR and Raman Spectroscopy-Based Biochemical Profiling Reflects Genomic Diversity of Clinical Candida Isolates That May Be Useful for Diagnosis and Targeted Therapy of Candidiasis. International Journal of Molecular Sciences. 2019; 20(4):988. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20040988
Chicago/Turabian StylePotocki, Leszek, Joanna Depciuch, Ewelina Kuna, Mariusz Worek, Anna Lewinska, and Maciej Wnuk. 2019. "FTIR and Raman Spectroscopy-Based Biochemical Profiling Reflects Genomic Diversity of Clinical Candida Isolates That May Be Useful for Diagnosis and Targeted Therapy of Candidiasis" International Journal of Molecular Sciences 20, no. 4: 988. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20040988