Cryopreservation of Canine Primary Dorsal Root Ganglion Neurons and Its Impact upon Susceptibility to Paramyxovirus Infection

Abstract
:1. Introduction
2. Results
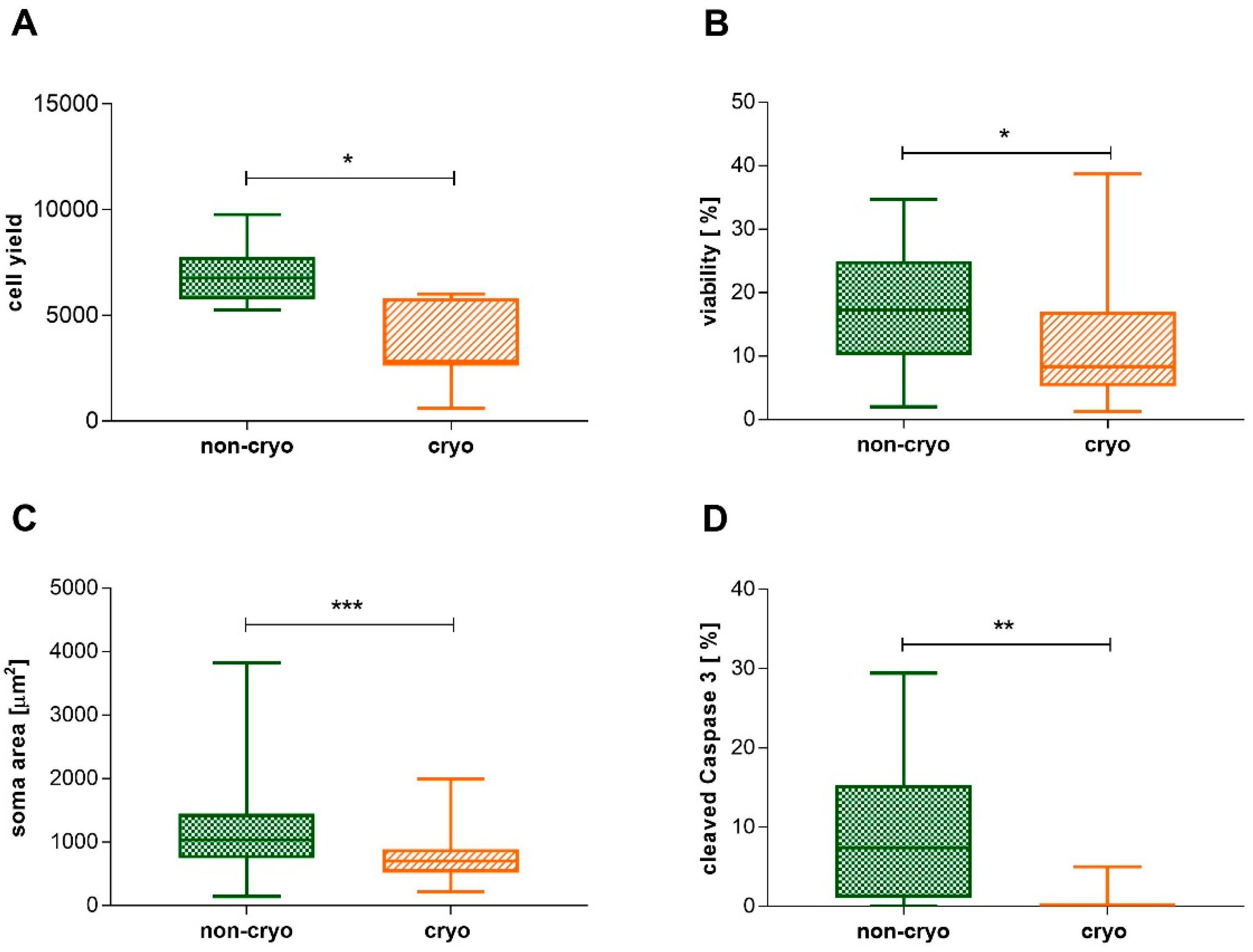
2.1. Non-Infected Cultures, Non-Cryopreserved and Cryopreserved: Neuronal Morphology, Cell Yield, Viability, and Cleaved Caspase 3-Expression
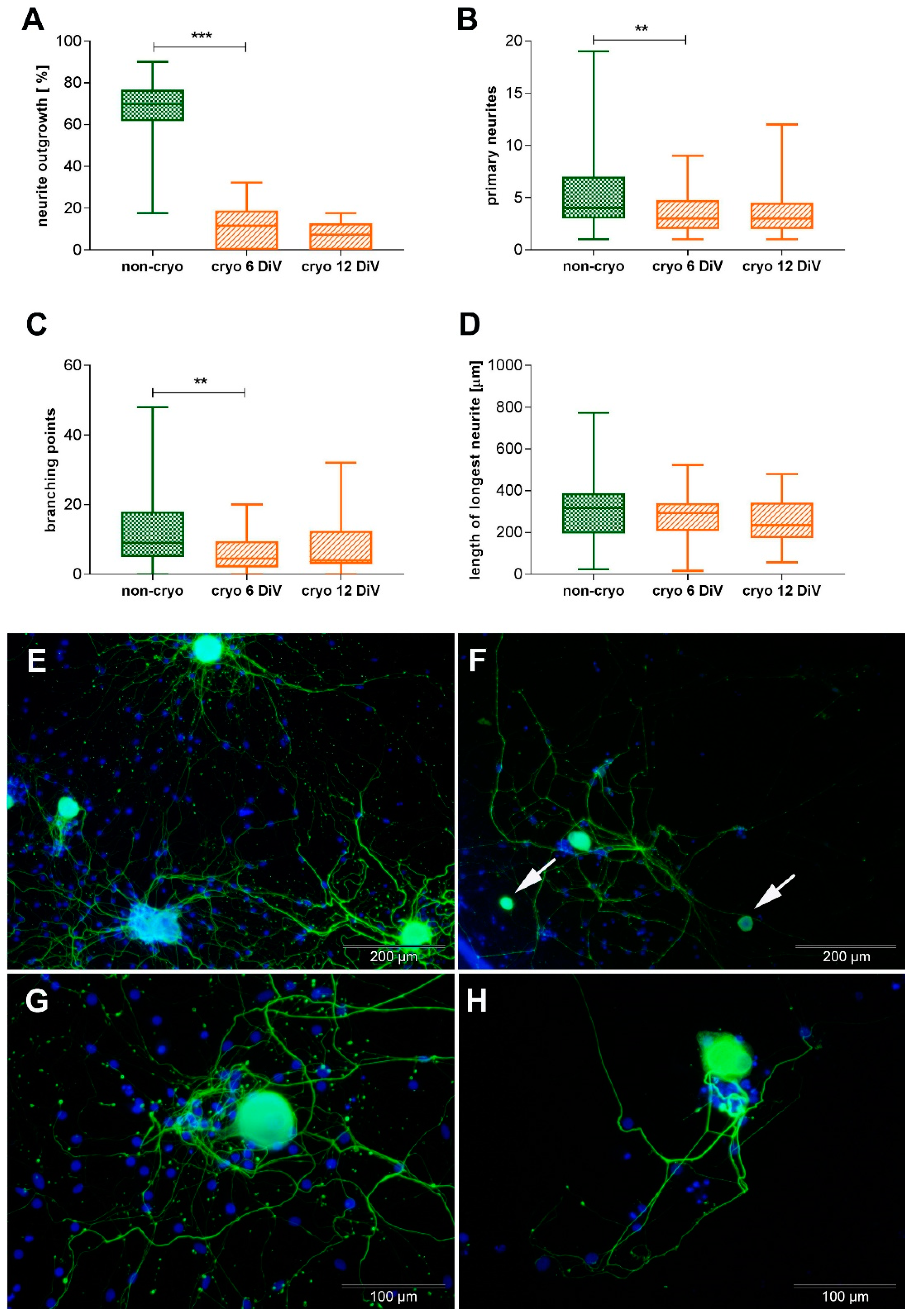
2.2. Non-Infected Cultures, Non-Cryopreserved and Cryopreserved: Neurite-Outgrowth
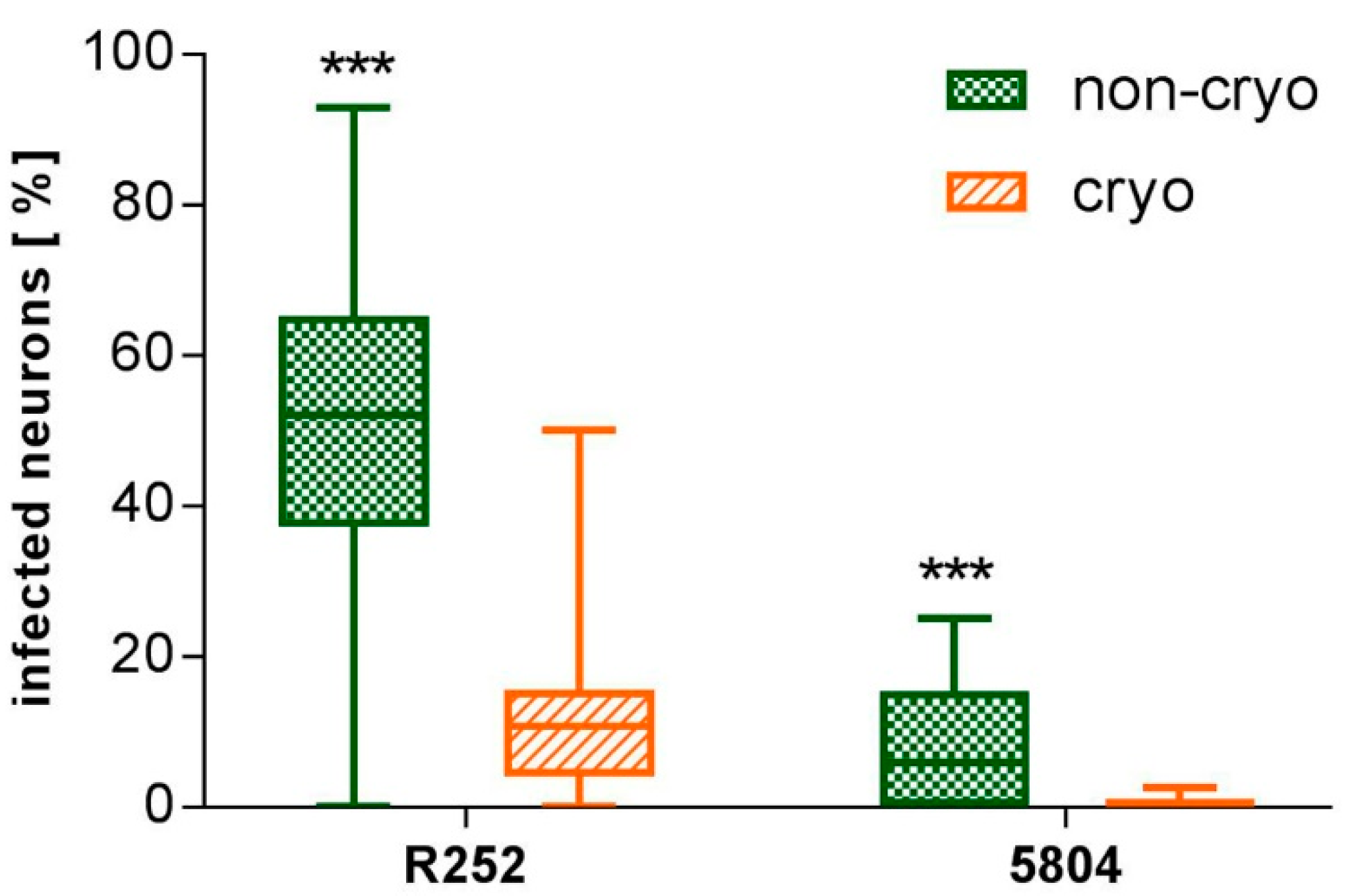
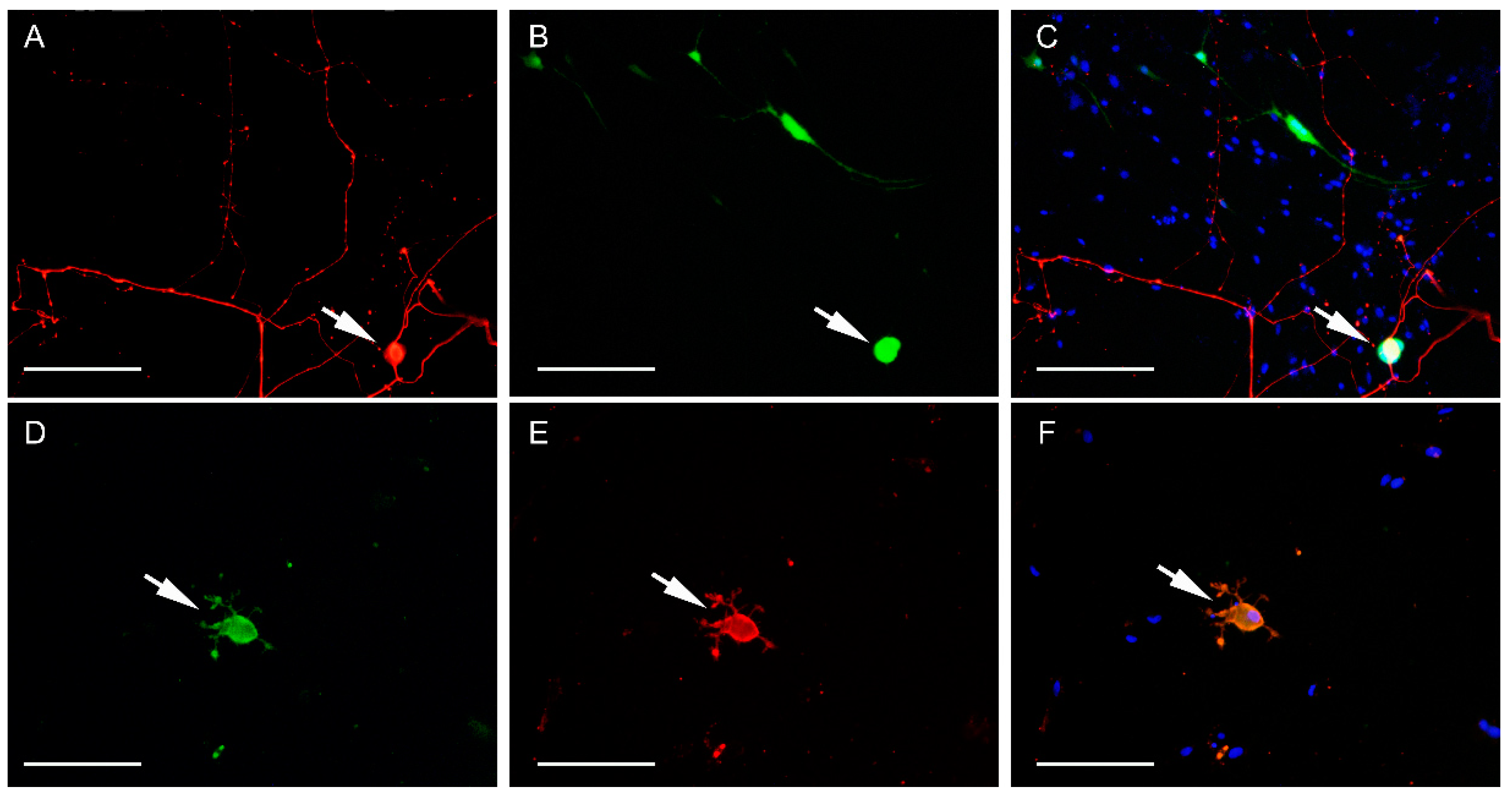
2.3. Non-Cryopreserved and Cryopreserved Cultures: Susceptibility to Canine Distemper Virus (CDV) Infection
3. Discussion
4. Materials and Methods
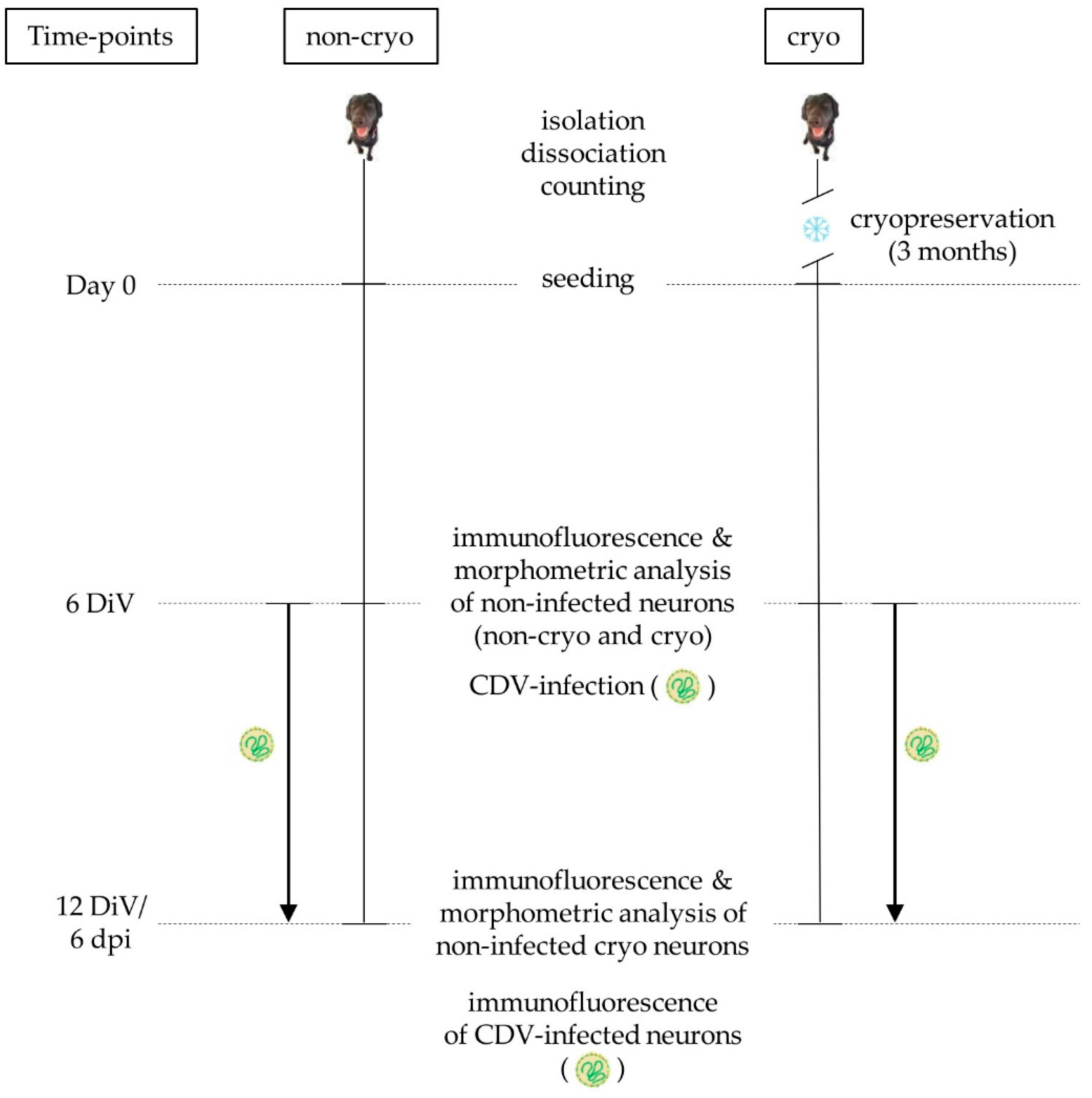
4.1. Isolation and Cultivation of Primary Canine Dorsal Root Ganglion Neurons
4.2. Cryopreservation and Cell Yield
4.3. Virus Strains and Virus Infection
4.4. Immunofluorescence
4.5. Comparative Analysis of Cell Morphology, Viability, Cleaved Caspase 3-Expression and Neurite Outgrowth of Non-Infected, Non-Cryopreserved and Cryopreserved Neurons
4.6. Evaluation of Neuronal CDV-Infection
4.7. Illustrations and Graphs
4.8. Statistics
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Buch, A.; Muller, O.; Ivanova, L.; Dohner, K.; Bialy, D.; Bosse, J.B.; Pohlmann, A.; Binz, A.; Hegemann, M.; Nagel, C.H.; et al. Inner tegument proteins of Herpes Simplex Virus are sufficient for intracellular capsid motility in neurons but not for axonal targeting. PLoS Pathog. 2017, 13, e1006813. [Google Scholar] [CrossRef] [PubMed]
- Gaburro, J.; Bhatti, A.; Sundaramoorthy, V.; Dearnley, M.; Green, D.; Nahavandi, S.; Paradkar, P.N.; Duchemin, J.B. Zika virus-induced hyper excitation precedes death of mouse primary neuron. Virol. J. 2018, 15, 79. [Google Scholar] [CrossRef] [PubMed]
- Omar, M.; Bock, P.; Kreutzer, R.; Ziege, S.; Imbschweiler, I.; Hansmann, F.; Peck, C.T.; Baumgärtner, W.; Wewetzer, K. Defining the morphological phenotype: 2’,3’-cyclic nucleotide 3’-phosphodiesterase (CNPase) is a novel marker for in situ detection of canine but not rat olfactory ensheathing cells. Cell Tissue Res. 2011, 344, 391–405. [Google Scholar] [CrossRef] [PubMed]
- Techangamsuwan, S.; Imbschweiler, I.; Kreutzer, R.; Kreutzer, M.; Baumgärtner, W.; Wewetzer, K. Similar behaviour and primate-like properties of adult canine Schwann cells and olfactory ensheathing cells in long-term culture. Brain Res. 2008, 1240, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Jeffery, N.D.; Smith, P.M.; Lakatos, A.; Ibanez, C.; Ito, D.; Franklin, R.J. Clinical canine spinal cord injury provides an opportunity to examine the issues in translating laboratory techniques into practical therapy. Spinal Cord. 2006, 44, 584–593. [Google Scholar] [CrossRef] [PubMed]
- Spitzbarth, I.; Bock, P.; Haist, V.; Stein, V.M.; Tipold, A.; Wewetzer, K.; Baumgärtner, W.; Beineke, A. Prominent microglial activation in the early proinflammatory immune response in naturally occurring canine spinal cord injury. J. Neuropathol. Exp. Neurol. 2011, 70, 703–714. [Google Scholar] [CrossRef] [PubMed]
- Wewetzer, K.; Radtke, C.; Kocsis, J.; Baumgärtner, W. Species-specific control of cellular proliferation and the impact of large animal models for the use of olfactory ensheathing cells and Schwann cells in spinal cord repair. Exp. Neurol. 2011, 229, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Beineke, A.; Puff, C.; Seehusen, F.; Baumgärtner, W. Pathogenesis and immunopathology of systemic and nervous canine distemper. Vet. Immunol. Immunopathol. 2009, 127, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Griot, C.; Vandevelde, M.; Schobesberger, M.; Zurbriggen, A. Canine distemper, a re-emerging morbillivirus with complex neuropathogenic mechanisms. Anim. Health Res. Rev. 2003, 4, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Nesseler, A.; Baumgärtner, W.; Gaedke, K.; Zurbriggen, A. Abundant expression of viral nucleoprotein mRNA and restricted translation of the corresponding viral protein in inclusion body polioencephalitis of canine distemper. J. Comp. Pathol. 1997, 116, 291–301. [Google Scholar] [CrossRef]
- Nesseler, A.; Baumgärtner, W.; Zurbriggen, A.; Orvell, C. Restricted virus protein translation in canine distemper virus inclusion body polioencephalitis. Vet. Microbiol. 1999, 69, 23–28. [Google Scholar] [CrossRef]
- Liebert, U.G.; Baczko, K.; Budka, H.; ter Meulen, V. Restricted expression of measles virus proteins in brains from cases of subacute sclerosing panencephalitis. J. Gen. Virol. 1986, 67, 2435–2444. [Google Scholar] [CrossRef] [PubMed]
- Isaacson, S.H.; Asher, D.M.; Godec, M.S.; Gibbs, C.J., Jr.; Gajdusek, D.C. Widespread, restricted low-level measles virus infection of brain in a case of subacute sclerosing panencephalitis. Acta Neuropathol. 1996, 91, 135–139. [Google Scholar] [CrossRef] [PubMed]
- Noyce, R.S.; Delpeut, S.; Richardson, C.D. Dog nectin-4 is an epithelial cell receptor for canine distemper virus that facilitates virus entry and syncytia formation. Virology 2013, 436, 210–220. [Google Scholar] [CrossRef] [PubMed]
- Tatsuo, H.; Ono, N.; Yanagi, Y. Morbilliviruses use signaling lymphocyte activation molecules (CD150) as cellular receptors. J. Virol. 2001, 75, 5842–5850. [Google Scholar] [CrossRef] [PubMed]
- Orlando, E.A.; Imbschweiler, I.; Gerhauser, I.; Baumgärtner, W.; Wewetzer, K. In vitro characterization and preferential infection by canine distemper virus of glial precursors with Schwann cell characteristics from adult canine brain. Neuropathol. Appl. Neurobiol. 2008, 34, 621–637. [Google Scholar] [CrossRef] [PubMed]
- Techangamsuwan, S.; Haas, L.; Rohn, K.; Baumgärtner, W.; Wewetzer, K. Distinct cell tropism of canine distemper virus strains to adult olfactory ensheathing cells and Schwann cells in vitro. Virus Res. 2009, 144, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Confer, A.W.; Kahn, D.E.; Koestner, A.; Krakowka, S. Biological properties of a canine distemper virus isolate associated with demyelinating encephalomyelitis. Infect. Immun. 1975, 11, 835–844. [Google Scholar] [PubMed]
- Summers, B.A.; Greisen, H.A.; Appel, M.J.G. Canine distemper encephalomyelitis: Variation with virus strain. J. Comp. Pathol. 1984, 94, 65–75. [Google Scholar] [CrossRef]
- Von Messling, V.; Milosevic, D.; Cattaneo, R. Tropism illuminated: Lymphocyte-based pathways blazed by lethal morbillivirus through the host immune system. Proc. Natl. Acad. Sci. USA 2004, 101, 14216–14221. [Google Scholar] [CrossRef] [PubMed]
- Luhmann, H.J.; Hanganu, I.; Kilb, W. Cellular physiology of the neonatal rat cerebral cortex. Brain Res. Bull. 2003, 60, 345–353. [Google Scholar] [CrossRef]
- Lu, J.; Karadsheh, M.; Delpire, E. Developmental regulation of the neuronal-specific isoform of K-Cl cotransporter KCC2 in postnatal rat brains. J. Neurobiol. 1999, 39, 558–568. [Google Scholar] [CrossRef]
- Wallis, K.; Sjogren, M.; van Hogerlinden, M.; Silberberg, G.; Fisahn, A.; Nordstrom, K.; Larsson, L.; Westerblad, H.; de Escobar, G.M.; Shupliakov, O.; et al. Locomotor deficiencies and aberrant development of subtype-specific GABAergic interneurons caused by an unliganded thyroid hormone receptor alpha1. J. Neurosci. 2008, 28, 1904–1915. [Google Scholar] [CrossRef] [PubMed]
- Schock, S.C.; Jolin-Dahel, K.S.; Schock, P.C.; Theiss, S.; Arbuthnott, G.W.; Garcia-Munoz, M.; Staines, W.A. Development of dissociated cryopreserved rat cortical neurons in vitro. J. Neurosci. Methods 2012, 205, 324–333. [Google Scholar] [CrossRef] [PubMed]
- Russell, W.M.S.B.R. The Principles of Humane Experimental Technique; Methuen: London, UK, 1959. [Google Scholar]
- Amir, R.; Michaelis, M.; Devor, M. Membrane potential oscillations in dorsal root ganglion neurons: Role in normal electrogenesis and neuropathic pain. J. Neurosci. 1999, 19, 8589–8596. [Google Scholar] [CrossRef] [PubMed]
- An, L.; Li, G.; Si, J.; Zhang, C.; Han, X.; Wang, S.; Jiang, L.; Xie, K. Acrylamide Retards the Slow Axonal Transport of Neurofilaments in Rat Cultured Dorsal Root Ganglia Neurons and the Corresponding Mechanisms. Neurochem. Res. 2016, 41, 1000–1009. [Google Scholar] [CrossRef] [PubMed]
- Gold, M.S.; Dastmalchi, S.; Levine, J.D. Co-expression of nociceptor properties in dorsal root ganglion neurons from the adult rat in vitro. Neuroscience 1996, 71, 265–275. [Google Scholar] [CrossRef]
- Murali, S.S.; Napier, I.A.; Mohammadi, S.A.; Alewood, P.F.; Lewis, R.J.; Christie, M.J. High-voltage-activated calcium current subtypes in mouse DRG neurons adapt in a subpopulation-specific manner after nerve injury. J. Neurophysiol. 2015, 113, 1511–1519. [Google Scholar] [CrossRef] [PubMed]
- Scroggs, R.S.; Fox, A.P. Calcium current variation between acutely isolated adult rat dorsal root ganglion neurons of different size. J. Physiol. 1992, 445, 639–658. [Google Scholar] [CrossRef] [PubMed]
- Gerhauser, I.; Hahn, K.; Baumgärtner, W.; Wewetzer, K. Culturing adult canine sensory neurons to optimise neural repair. Vet. Rec. 2012, 170, 102. [Google Scholar] [CrossRef] [PubMed]
- Tongtako, W.; Lehmbecker, A.; Wang, Y.; Hahn, K.; Baumgärtner, W.; Gerhauser, I. Canine dorsal root ganglia satellite glial cells represent an exceptional cell population with astrocytic and oligodendrocytic properties. Sci. Rep. 2017, 7, 13915. [Google Scholar] [CrossRef] [PubMed]
- Negishi, T.; Ishii, Y.; Kawamura, S.; Kuroda, Y.; Yoshikawa, Y. Cryopreservation of brain tissue for primary culture. Exp. Anim. 2002, 51, 383–390. [Google Scholar] [CrossRef] [PubMed]
- Pischedda, F.; Montani, C.; Obergasteiger, J.; Frapporti, G.; Corti, C.; Rosato Siri, M.; Volta, M.; Piccoli, G. Cryopreservation of Primary Mouse Neurons: The Benefit of Neurostore Cryoprotective Medium. Front. Cell Neurosci. 2018, 12, 81. [Google Scholar] [CrossRef] [PubMed]
- Borlongan, C.V.; Tajima, Y.; Trojanowski, J.Q.; Lee, V.M.; Sanberg, P.R. Transplantation of cryopreserved human embryonal carcinoma-derived neurons (NT2N cells) promotes functional recovery in ischemic rats. Exp. Neurol. 1998, 149, 310–321. [Google Scholar] [CrossRef] [PubMed]
- Uemura, M.; Ishiguro, H. Freezing behavior of adherent neuron-like cells and morphological change and viability of post-thaw cells. Cryobiology 2015, 70, 122–135. [Google Scholar] [CrossRef] [PubMed]
- Edamura, K.; Nakano, R.; Fujimoto, K.; Teshima, K.; Asano, K.; Tanaka, S. Effects of cryopreservation on the cell viability, proliferative capacity and neuronal differentiation potential of canine bone marrow stromal cells. J. Vet. Med. Sci. 2014, 76, 573–577. [Google Scholar] [CrossRef] [PubMed]
- Hancock, C.R.; Wetherington, J.P.; Lambert, N.A.; Condie, B.G. Neuronal differentiation of cryopreserved neural progenitor cells derived from mouse embryonic stem cells. Biochem. Biophys. Res. Commun. 2000, 271, 418–421. [Google Scholar] [CrossRef] [PubMed]
- Ladewig, J.; Koch, P.; Endl, E.; Meiners, B.; Opitz, T.; Couillard-Despres, S.; Aigner, L.; Brustle, O. Lineage selection of functional and cryopreservable human embryonic stem cell-derived neurons. Stem. Cells 2008, 26, 1705–1712. [Google Scholar] [CrossRef] [PubMed]
- Taupin, P. Cryopreservation of early postmitotic neuronal cells in culture. Expert Opin. Ther. Pat. 2009, 19, 265–268. [Google Scholar] [CrossRef] [PubMed]
- Ichikawa, J.; Yamada, R.X.; Muramatsu, R.; Ikegaya, Y.; Matsuki, N.; Koyama, R. Cryopreservation of granule cells from the postnatal rat hippocampus. J. Pharmacol. Sci. 2007, 104, 387–391. [Google Scholar] [CrossRef] [PubMed]
- Mattson, M.P.; Kater, S.B. Development and selective neurodegeneration in cell cultures from different hippocampal regions. Brain Res. 1989, 490, 110–125. [Google Scholar] [CrossRef]
- Robert, M.C.; Juan de Paz, L.; Graf, D.A.; Gazzin, S.; Tiribelli, C.; Bottai, H.; Rodriguez, J.V. Cryopreservation by slow cooling of rat neuronal cells. Cryobiology 2016, 72, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Seggio, A.M.; Ellison, K.S.; Hynd, M.R.; Shain, W.; Thompson, D.M. Cryopreservation of transfected primary dorsal root ganglia neurons. J. Neurosci. Methods 2008, 173, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Negishi, T.; Ishii, Y.; Kawamura, S.; Kuroda, Y.; Yoshikawa, Y. Cryopreservation and primary culture of cerebral neurons from cynomolgus monkeys (Macaca fascicularis). Neurosci. Lett. 2002, 328, 21–24. [Google Scholar] [CrossRef]
- Kay, G.W.; Oswald, M.J.; Palmer, D.N. The development and characterisation of complex ovine neuron cultures from fresh and frozen foetal neurons. J. Neurosci. Methods 2006, 155, 98–108. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, A.; Onodera, T.; Ikeda, H.; Kitani, H. Isolation and characterisation of fetal bovine brain cells in primary culture. Res. Vet. Sci. 2000, 69, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Cai, R.S.; Xue, D.L.; Jiang, X.H. Cryopreservation and culture of the human fetal brain tissues. J. Tongji Med. Univ. 1993, 13, 138–142. [Google Scholar] [PubMed]
- Mattson, M.P.; Rychlik, B. Cell culture of cryopreserved human fetal cerebral cortical and hippocampal neurons: Neuronal development and responses to trophic factors. Brain Res. 1990, 522, 204–214. [Google Scholar] [CrossRef]
- Silani, V.; Pizzuti, A.; Strada, O.; Falini, A.; Buscaglia, M.; Scarlato, G. Human neuronal cell viability demonstrated in culture after cryopreservation. Brain Res. 1988, 473, 169–174. [Google Scholar] [CrossRef]
- Paynter, S.J. Principles and practical issues for cryopreservation of nerve cells. Brain Res. Bull 2008, 75, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Swett, J.W.; Paramore, C.G.; Turner, D.A. Quantitative estimation of cryopreservation viability in rat fetal hippocampal cells. Exp. Neurol. 1994, 129, 330–334. [Google Scholar] [CrossRef] [PubMed]
- Andersen, N.D.; Srinivas, S.; Pinero, G.; Monje, P.V. A rapid and versatile method for the isolation, purification and cryogenic storage of Schwann cells from adult rodent nerves. Sci. Rep. 2016, 6, 31781. [Google Scholar] [CrossRef] [PubMed]
- Andres, K.H. Untersuchungen über den Feinbau von Spinalganglien. Zeitschrift für Zellforschung und Mikroskopische Anatomie 1961, 55, 1–48. [Google Scholar] [CrossRef] [PubMed]
- Roloff, F.; Ziege, S.; Baumgärtner, W.; Wewetzer, K.; Bicker, G. Schwann cell-free adult canine olfactory ensheathing cell preparations from olfactory bulb and mucosa display differential migratory and neurite growth-promoting properties in vitro. BMC Neurosci. 2013, 14, 141. [Google Scholar] [CrossRef] [PubMed]
- Usoskin, D.; Furlan, A.; Islam, S.; Abdo, H.; Lonnerberg, P.; Lou, D.; Hjerling-Leffler, J.; Haeggstrom, J.; Kharchenko, O.; Kharchenko, P.V.; et al. Unbiased classification of sensory neuron types by large-scale single-cell RNA sequencing. Nat. Neurosci. 2015, 18, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Bonfanti, L.; Bellardi, S.; Ghidella, S.; Gobetto, A.; Polak, J.M.; Merighi, A. Distribution of five peptides, three general neuroendocrine markers, and two synaptic-vesicle-associated proteins in the spinal cord and dorsal root ganglia of the adult and newborn dog: An immunocytochemical study. Am. J. Anat. 1991, 191, 154–166. [Google Scholar] [CrossRef] [PubMed]
- Molliver, D.C.; Wright, D.E.; Leitner, M.L.; Parsadanian, A.S.; Doster, K.; Wen, D.; Yan, Q.; Snider, W.D. IB4-binding DRG neurons switch from NGF to GDNF dependence in early postnatal life. Neuron 1997, 19, 849–861. [Google Scholar] [CrossRef]
- Heng, B.C.; Ye, C.P.; Liu, H.; Toh, W.S.; Rufaihah, A.J.; Yang, Z.; Bay, B.H.; Ge, Z.; Ouyang, H.W.; Lee, E.H.; et al. Loss of viability during freeze-thaw of intact and adherent human embryonic stem cells with conventional slow-cooling protocols is predominantly due to apoptosis rather than cellular necrosis. J. Biomed. Sci. 2006, 13, 433–445. [Google Scholar] [CrossRef] [PubMed]
- Mattson, M.P.; Dou, P.; Kater, S.B. Outgrowth-regulating actions of glutamate in isolated hippocampal pyramidal neurons. J. Neurosci. 1988, 8, 2087–2100. [Google Scholar] [CrossRef] [PubMed]
- Mattson, M.P.; Kater, S.B. Isolated hippocampal neurons in cryopreserved long-term cultures: Development of neuroarchitecture and sensitivity to NMDA. Int. J. Dev. Neurosci. 1988, 6, 439–452. [Google Scholar] [CrossRef]
- Techangamsuwan, S.A.; Enzo, D.; Cundi, O.; Imbschweiler, I.; Gerhauser, I.; Wewetzer, K.; Baumgärtner, W. Cell-cell Interaction Appears to Represent a Pivotal Factor for Susceptibility of Adult Canine Schwann Cell-like Brain Glia to Canine Distemper Virus Infection in vitro. Thai J. Vet. Med. 2011, 41, 213–225. [Google Scholar]
- Bottenstein, J.E.; Sato, G.H. Growth of a rat neuroblastoma cell line in serum-free supplemented medium. Proc. Natl. Acad. Sci. USA 1979, 76, 514–517. [Google Scholar] [CrossRef] [PubMed]
- Ziege, S.; Baumgärtner, W.; Wewetzer, K. Toward defining the regenerative potential of olfactory mucosa: Establishment of Schwann cell-free adult canine olfactory ensheathing cell preparations suitable for transplantation. Cell Transplant. 2013, 22, 355–367. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Condition | Non-Cryo | Cryo |
|---|---|---|
| glial cells [number/well] | 470 *** (+/− 164.21) | 225 *** (+/− 104.65) |
| glial cells [%/well] | 94.63 (+/− 2.56) | 93.20 (+/− 4.02) |
| neurons [number/well] | 26 * (+/− 13.66) | 18 * (+/−15.6) |
| neurons [%/well] | 5.37 (+/− 2.56) | 6.80 (+/− 5.66) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schwarz, S.; Spitzbarth, I.; Baumgärtner, W.; Lehmbecker, A. Cryopreservation of Canine Primary Dorsal Root Ganglion Neurons and Its Impact upon Susceptibility to Paramyxovirus Infection. Int. J. Mol. Sci. 2019, 20, 1058. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20051058
Schwarz S, Spitzbarth I, Baumgärtner W, Lehmbecker A. Cryopreservation of Canine Primary Dorsal Root Ganglion Neurons and Its Impact upon Susceptibility to Paramyxovirus Infection. International Journal of Molecular Sciences. 2019; 20(5):1058. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20051058
Chicago/Turabian StyleSchwarz, Sarah, Ingo Spitzbarth, Wolfgang Baumgärtner, and Annika Lehmbecker. 2019. "Cryopreservation of Canine Primary Dorsal Root Ganglion Neurons and Its Impact upon Susceptibility to Paramyxovirus Infection" International Journal of Molecular Sciences 20, no. 5: 1058. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20051058