Testis-Specific SEPT12 Expression Affects SUN Protein Localization and is Involved in Mammalian Spermiogenesis


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
1.1. Mammalian Spermiogenesis and Male Infertility
1.2. Roles of Septins in Mammalian Spermatogenesis
1.3. Sperm Head Formation and SUN Proteins
1.4. SEPT12/SPAG4/LAMIN Complex
2. Results
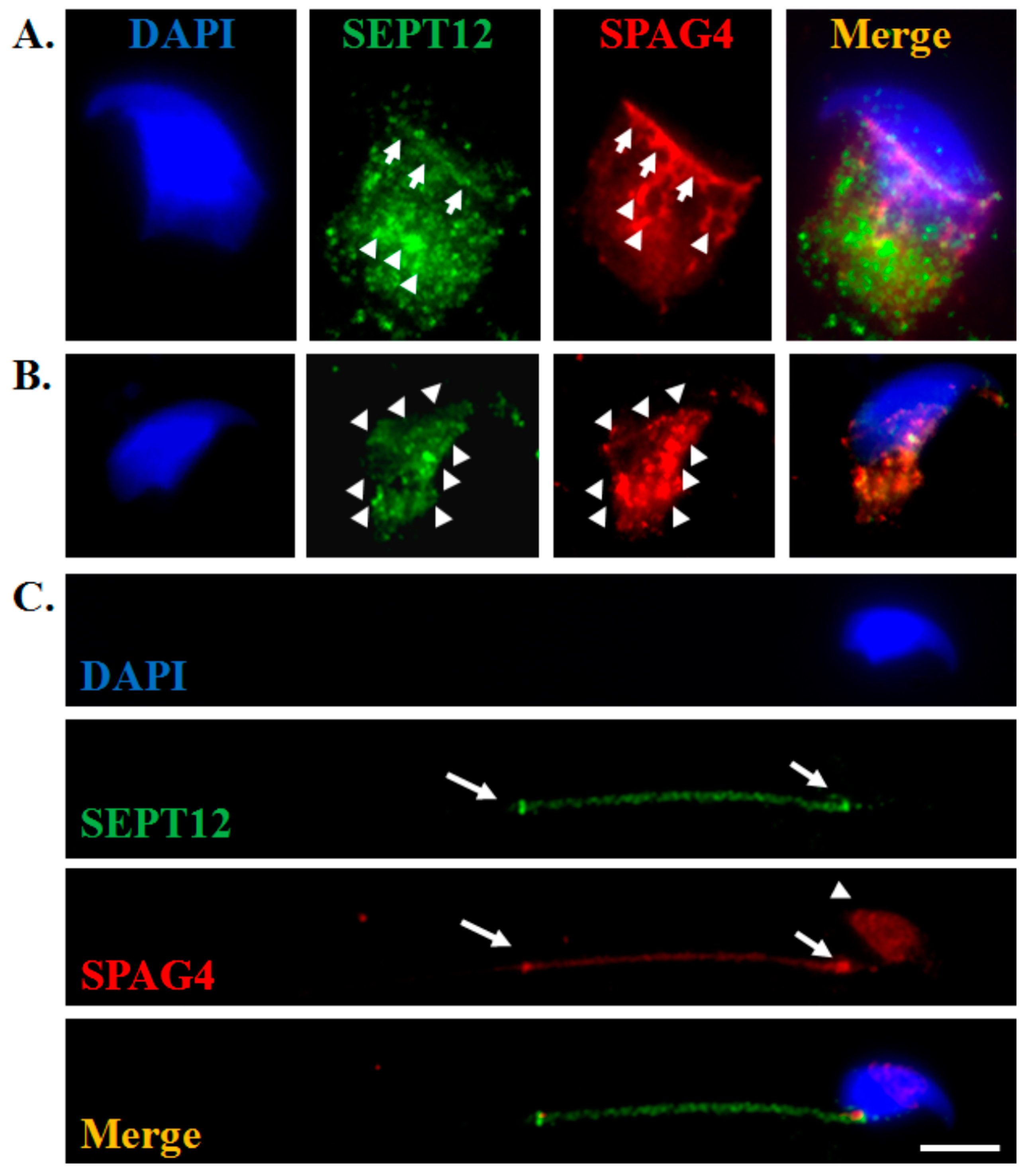
2.1. SEPT12 is Colocalized With SPAG4 During Murine Spermiogenesis
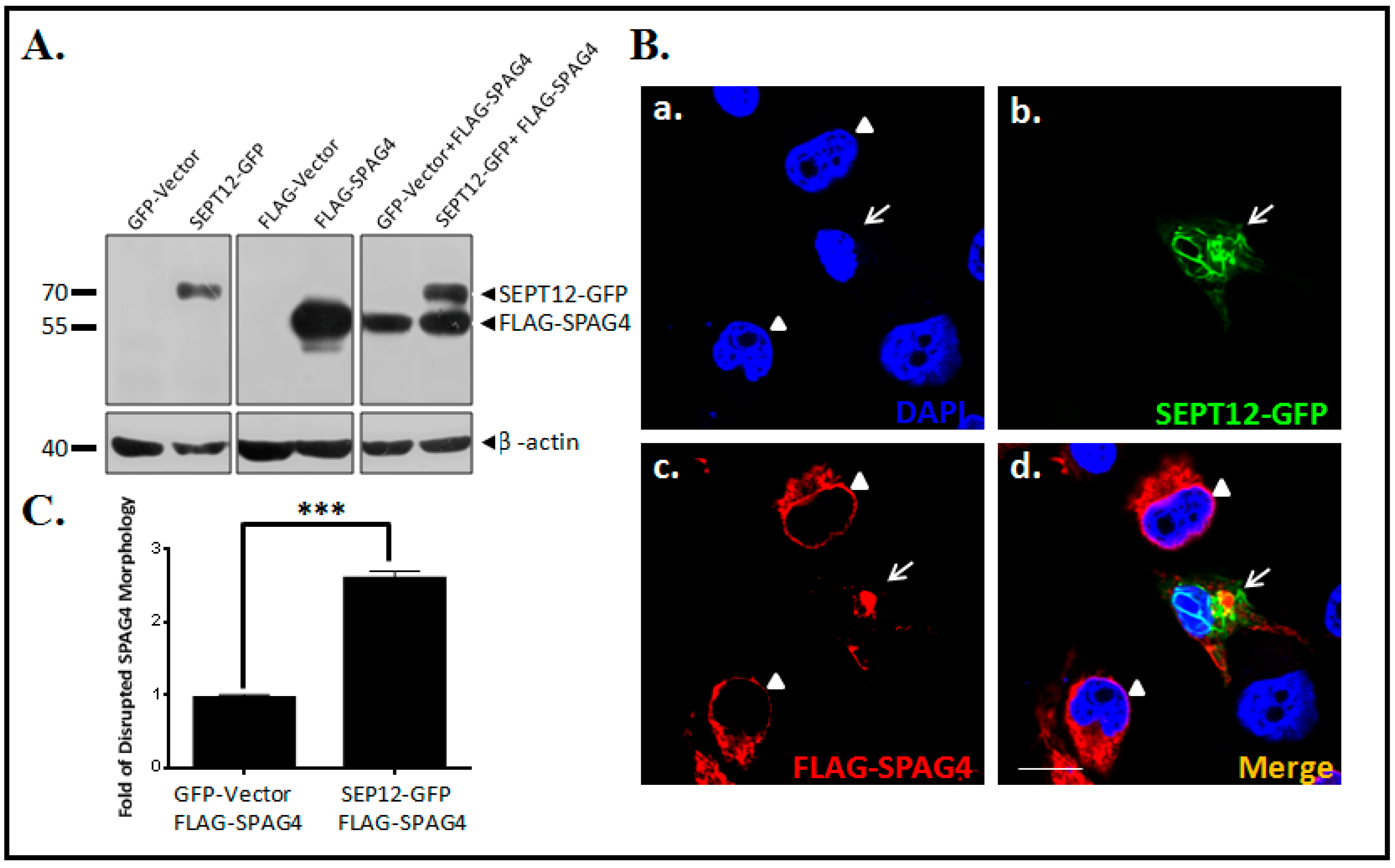
2.2. SEPT12 Alters SPAG4 Localization
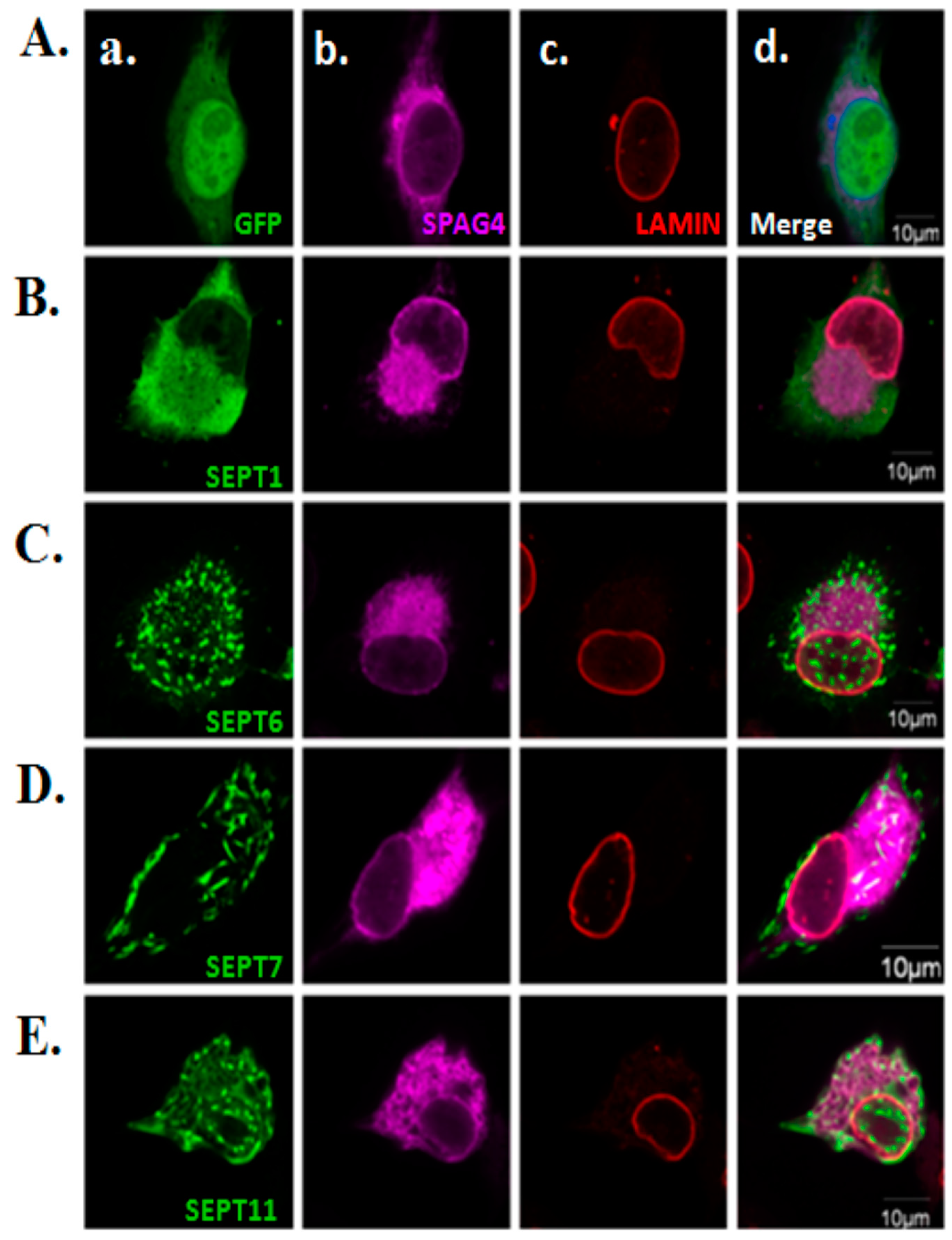
2.3. Morphological Effects of SPAG4 on SEPT1/6/7/11
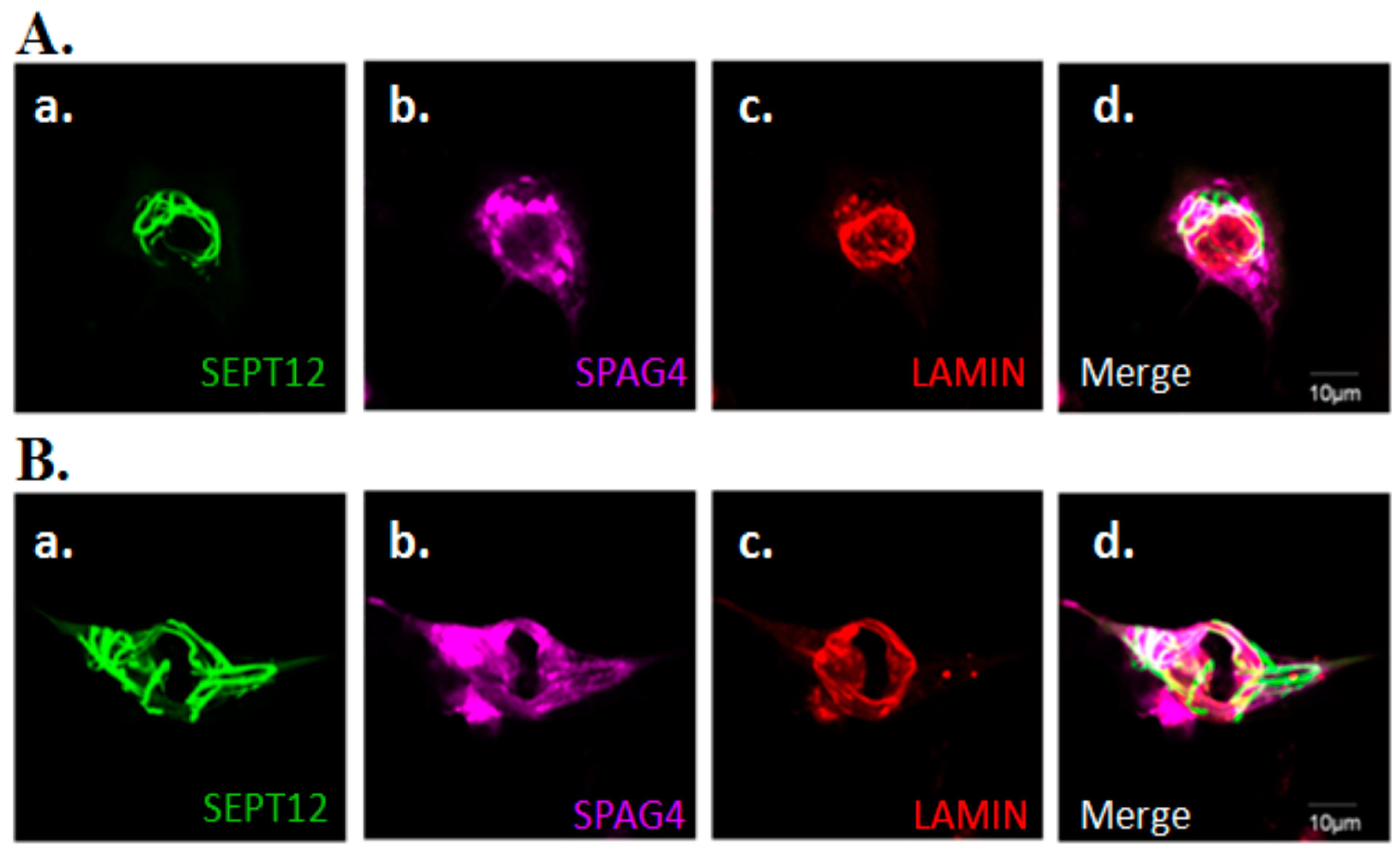
2.4. Effects of SEPT12 on SPAG4/LAMIN complexes
3. Discussion
3.1. Localization of SEPT12 is Similar to That of SPAG4 During Murine Spermiogenesis
3.2. Localization of SPAG4 in Different Cell Models
3.3. Possible Roles of SEPT12/SPAG4/LAMIN Complexes During Sperm Head Shaping
3.4. Teratozoospermia and LAMIN/SAPG4/SEPT12 Complexes
4. Experimental Section
4.1. Isolated Murine Testicular Germ Cells
4.2. Immunofluorescence Assay and Western Blotting
4.3. Cloning and Transfection
4.4. Cell Culture
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Kierszenbaum, A.L.; Rivkin, E.; Tres, L.L. Acroplaxome, an F-actin-keratin-containing plate, anchors the acrosome to the nucleus during shaping of the spermatid head. Mol. Biol. Cell 2003, 14, 4628–4640. [Google Scholar] [CrossRef] [PubMed]
- Cooke, H.J.; Saunders, P.T. Mouse models of male infertility. Nat. Rev. Genet. 2002, 3, 790–801. [Google Scholar] [CrossRef] [PubMed]
- Carlsen, E.; Giwercman, A.; Keiding, N.; Skakkebaek, N.E. Evidence for decreasing quality of semen during past 50 years. BMJ 1992, 305, 609–613. [Google Scholar] [CrossRef] [PubMed]
- Huang, P.-H.; Wang, H.-Y.; Huang, C.-C.; Lee, Y.-T.; Yue, C.-H.; Chen, M.-C.; Lin, H. Suppression of Breast Cancer Cell Growth by Her2-Reduced AR Serine 81 Phosphorylation. Chin. J. Physiol. 2016, 59, 232–239. [Google Scholar] [CrossRef] [PubMed]
- Takizawa, P.A.; DeRisi, J.L.; Wilhelm, J.E.; Vale, R.D. Plasma membrane compartmentalization in yeast by messenger RNA transport and a septin diffusion barrier. Science 2000, 290, 341–344. [Google Scholar] [CrossRef] [PubMed]
- Mostowy, S.; Cossart, P. Septins: The fourth component of the cytoskeleton. Nat. Rev. Mol. Cell Biol. 2012, 13, 183–194. [Google Scholar] [CrossRef] [PubMed]
- Sirajuddin, M.; Farkasovsky, M.; Hauer, F.; Kuhlmann, D.; Macara, I.G.; Weyand, M.; Stark, H.; Wittinghofer, A. Structural insight into filament formation by mammalian septins. Nature 2007, 449, 311–315. [Google Scholar] [CrossRef] [PubMed]
- Ihara, M.; Kinoshita, A.; Yamada, S.; Tanaka, H.; Tanigaki, A.; Kitano, A.; Goto, M.; Okubo, K.; Nishiyama, H.; Ogawa, O.; et al. Cortical organization by the septin cytoskeleton is essential for structural and mechanical integrity of mammalian spermatozoa. Dev. Cell 2005, 8, 343–352. [Google Scholar] [CrossRef] [PubMed]
- Hall, P.A.; Jung, K.; Hillan, K.J.; Russell, S.E. Expression profiling the human septin gene family. J. Pathol. 2005, 206, 269–278. [Google Scholar] [CrossRef] [PubMed]
- Kissel, H.; Georgescu, M.M.; Larisch, S.; Manova, K.; Hunnicutt, G.R.; Steller, H. The Sept4 septin locus is required for sperm terminal differentiation in mice. Dev. Cell 2005, 8, 353–364. [Google Scholar] [CrossRef] [PubMed]
- Kuo, Y.C.; Shen, Y.R.; Chen, H.I.; Lin, Y.H.; Wang, Y.Y.; Chen, Y.R.; Wang, C.Y.; Kuo, P.L. SEP12 orchestractes the formation of mammalian sperm annulus by orgaonizing core octameric complexes with other SEPT proteins. J. Cell Sci. 2015, 128, 923–935. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.H.; Lin, Y.M.; Teng, Y.N.; Hsieh, T.Y.; Lin, Y.S.; Kuo, P.L. Identification of ten novel genes involved in human spermatogenesis by microarray analysis of testicular tissue. Fertil. Steril. 2006, 86, 1650–1658. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.H.; Lin, Y.M.; Wang, Y.Y.; Yu, I.S.; Lin, Y.W.; Wang, Y.H.; Wu, C.M.; Pan, H.A.; Chao, S.C.; Yen, P.H.; et al. The Expression level of septin12 is critical for spermiogenesis. Am. J. Pathol 2009, 174, 1857–1868. [Google Scholar] [CrossRef] [PubMed]
- Kuo, Y.C.; Lin, Y.H.; Chen, H.I.; Wang, Y.Y.; Chiou, Y.W.; Lin, H.H.; Pan, H.A.; Wu, C.M.; Su, S.M.; Hsu, C.C.; et al. SEPT12 mutations cause male infertility with defective sperm annulus. Hum. Mutat 2012, 33, 710–719. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.H.; Wang, Y.Y.; Chen, H.I.; Kuo, Y.C.; Chiou, Y.W.; Lin, H.H.; Wu, C.M.; Hsu, C.C.; Chiang, H.S.; Kuo, P.L. SEPTIN12 Genetic Variants Confer Susceptibility to Teratozoospermia. PLoS ONE 2012, 7, e34011. [Google Scholar] [CrossRef] [PubMed]
- Pasch, E.; Link, J.; Beck, C.; Scheuerle, S.; Alsheimer, M. The LINC complex component Sun4 plays a crucial role in sperm head formation and fertility. Biol. Open 2015, 4, 1792–1802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kierszenbaum, A.L.; Tres, L.L. The acrosome-acroplaxome-manchette complex and the shaping of the spermatid head. Arch. Histol. Cytol. 2004, 67, 271–284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Donnell, L.; O’Bryan, M.K. Microtubules and spermatogenesis. Semin. Cell Dev. Biol. 2014, 30, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Gob, E.; Schmitt, J.; Benavente, R.; Alsheimer, M. Mammalian sperm head formation involves different polarization of two novel LINC complexes. PLoS ONE 2010, 5, e12072. [Google Scholar] [CrossRef] [PubMed]
- Haque, F.; Lloyd, D.J.; Smallwood, D.T.; Dent, C.L.; Shanahan, C.M.; Fry, A.M.; Trembath, R.C.; Shackleton, S. SUN1 interacts with nuclear lamin A and cytoplasmic nesprins to provide a physical connection between the nuclear lamina and the cytoskeleton. Mol. Cell Biol. 2006, 26, 3738–3751. [Google Scholar] [CrossRef] [PubMed]
- Tzur, Y.B.; Wilson, K.L.; Gruenbaum, Y. SUN-domain proteins: ‘Velcro’ that links the nucleoskeleton to the cytoskeleton. Nat. Rev. Mol. Cell Biol. 2006, 7, 782–788. [Google Scholar] [CrossRef] [PubMed]
- Shao, X.; Tarnasky, H.A.; Lee, J.P.; Oko, R.; van der Hoorn, F.A. Spag4, a novel sperm protein, binds outer dense-fiber protein Odf1 and localizes to microtubules of manchette and axoneme. Dev. Biol. 1999, 211, 109–123. [Google Scholar] [CrossRef] [PubMed]
- Kracklauer, M.P.; Wiora, H.M.; Deery, W.J.; Chen, X.; Bolival, B., Jr.; Romanowicz, D.; Simonette, R.A.; Fuller, M.T.; Fischer, J.A.; Beckingham, K.M. The Drosophila SUN protein Spag4 cooperates with the coiled-coil protein Yuri Gagarin to maintain association of the basal body and spermatid nucleus. J. Cell Sci. 2010, 123 Pt 16, 2763–2772. [Google Scholar] [CrossRef] [Green Version]
- Calvi, A.; Wong, A.S.; Wright, G.; Wong, E.S.; Loo, T.H.; Stewart, C.L.; Burke, B. SUN4 is essential for nuclear remodeling during mammalian spermiogenesis. Dev. Biol. 2015, 407, 321–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mouassite, M.; Camougrand, N.; Schwob, E.; Demaison, G.; Laclau, M.; Guerin, M. The ‘SUN’ family: Yeast SUN4/SCW3 is involved in cell septation. Yeast 2000, 16, 905–919. [Google Scholar] [CrossRef]
- Yeh, C.H.; Kuo, P.L.; Wang, Y.Y.; Wu, Y.Y.; Chen, M.F.; Lin, D.Y.; Lai, T.H.; Chiang, H.S.; Lin, Y.H. SEPT12/SPAG4/LAMINB1 complexes are required for maintaining the integrity of the nuclear envelope in postmeiotic male germ cells. PLoS ONE 2015, 10, e0120722. [Google Scholar] [CrossRef] [PubMed]
- Hasan, S.; Guttinger, S.; Muhlhausser, P.; Anderegg, F.; Burgler, S.; Kutay, U. Nuclear envelope localization of human UNC84A does not require nuclear lamins. FEBS Lett. 2006, 580, 1263–1268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, X.; Xu, R.; Yu, J.; Xu, T.; Zhuang, Y.; Han, M. SUN1 is required for telomere attachment to nuclear envelope and gametogenesis in mice. Dev. Cell 2007, 12, 863–872. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.H.; Chou, C.K.; Hung, Y.C.; Yu, I.S.; Pan, H.A.; Lin, S.W.; Kuo, P.L. SEPT12 deficiency causes sperm nucleus damage and developmental arrest of preimplantation embryos. Fertil. Steril. 2011, 95, 363–365. [Google Scholar] [CrossRef] [PubMed]
- Kuo, P.L.; Chiang, H.S.; Wang, Y.Y.; Kuo, Y.C.; Chen, M.F.; Yu, I.S.; Teng, Y.N.; Lin, S.W.; Lin, Y.H. SEPT12-microtubule complexes are required for sperm head and tail formation. Int. J. Mol. Sci. 2013, 14, 22102–22116. [Google Scholar] [CrossRef] [PubMed]
- Palermo, G.; Joris, H.; Devroey, P.; Van Steirteghem, A.C. Pregnancies after intracytoplasmic injection of single spermatozoon into an oocyte. Lancet 1992, 340, 17–18. [Google Scholar] [CrossRef]
- Silber, S.J. Evaluation and treatment of male infertility. Clin. Obstet. Gynecol. 2000, 43, 854–888. [Google Scholar] [CrossRef] [PubMed]
- Yeh, Y.C.; Yang, V.C.; Huang, S.C.; Lo, N.W. Stage-dependent expression of extra-embryonic tissue-spermatogenesis-homeobox gene 1 (ESX1) protein, a candidate marker for X chromosome-bearing sperm. Reprod. Fertil. Dev. 2005, 17, 447–455. [Google Scholar] [CrossRef] [PubMed]
- Ke, C.C.; Lin, Y.H.; Wang, Y.Y.; Wu, Y.Y.; Chen, M.F.; Ku, W.C.; Chiang, H.S.; Lai, T.H. TBC1D21 Potentially Interacts with and Regulates Rap1 during Murine Spermatogenesis. Int. J. Mol. Sci. 2018, 19, 3292. [Google Scholar] [CrossRef] [PubMed]
- Lai, T.H.; Wu, Y.Y.; Wang, Y.Y.; Chen, M.F.; Wang, P.; Chen, T.M.; Wu, Y.N.; Chiang, H.S.; Kuo, P.L.; Lin, Y.H. SEPT12-NDC1 Complexes Are Required for Mammalian Spermiogenesis. Int. J. Mol. Sci. 2016, 17, 1911. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.Y.; Wang, Y.Y.; Chen, Y.L.; Chen, M.F.; Chiang, H.S.; Kuo, P.L.; Lin, Y.H. CDC42 Negatively Regulates Testis-Specific SEPT12 Polymerization. Int. J. Mol. Sci. 2018, 19, 2627. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.H.; Lin, Y.M.; Kuo, Y.C.; Wang, Y.Y.; Kuo, P.L. Identification and characterization of a novel Rab GTPase-activating protein in spermatids. Int. J. Androl. 2011, 34 Pt 2, e358–e367. [Google Scholar] [CrossRef]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yeh, C.-H.; Wang, Y.-Y.; Wee, S.-K.; Chen, M.-F.; Chiang, H.-S.; Kuo, P.-L.; Lin, Y.-H. Testis-Specific SEPT12 Expression Affects SUN Protein Localization and is Involved in Mammalian Spermiogenesis. Int. J. Mol. Sci. 2019, 20, 1163. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20051163
Yeh C-H, Wang Y-Y, Wee S-K, Chen M-F, Chiang H-S, Kuo P-L, Lin Y-H. Testis-Specific SEPT12 Expression Affects SUN Protein Localization and is Involved in Mammalian Spermiogenesis. International Journal of Molecular Sciences. 2019; 20(5):1163. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20051163
Chicago/Turabian StyleYeh, Chung-Hsin, Ya-Yun Wang, Shi-Kae Wee, Mei-Feng Chen, Han-Sun Chiang, Pao-Lin Kuo, and Ying-Hung Lin. 2019. "Testis-Specific SEPT12 Expression Affects SUN Protein Localization and is Involved in Mammalian Spermiogenesis" International Journal of Molecular Sciences 20, no. 5: 1163. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20051163