A Cotton (Gossypium hirsutum) Myo-Inositol-1-Phosphate Synthase (GhMIPS1D) Gene Promotes Root Cell Elongation in Arabidopsis


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
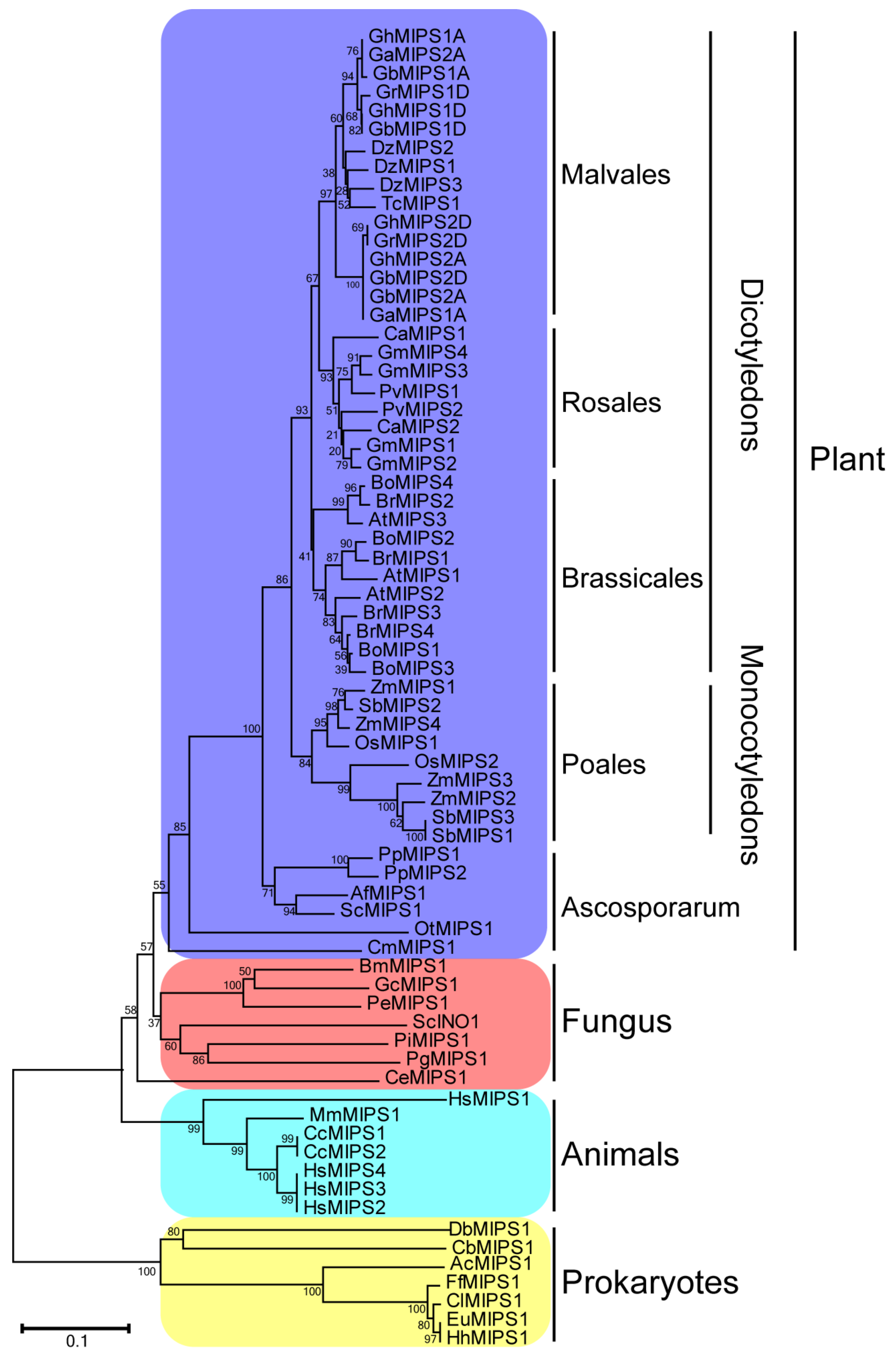
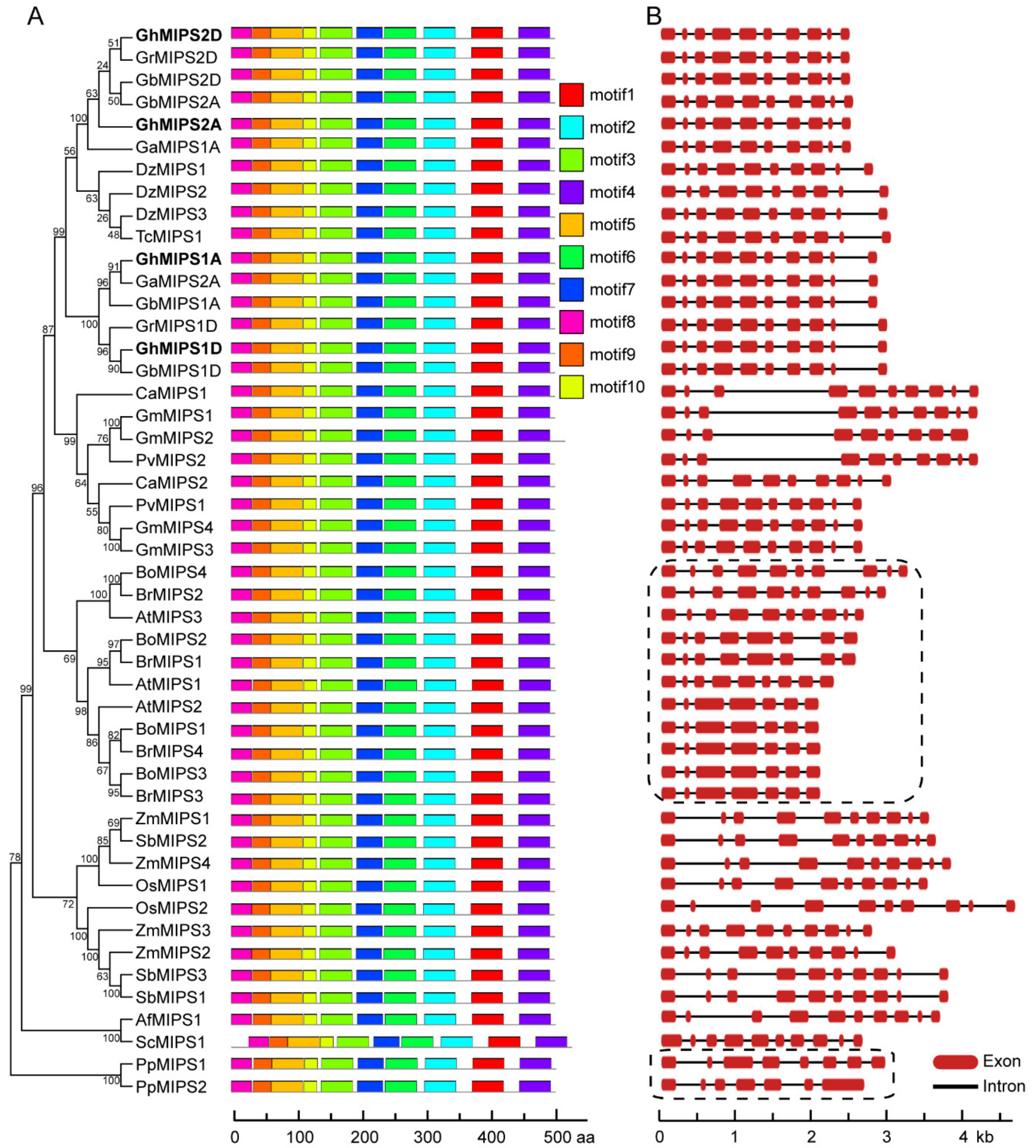
2.1. Characterization, Phylogenetic Relationship and Motif Distribution Analyses of GhMIPS Gene Family Members in G. hirsutum
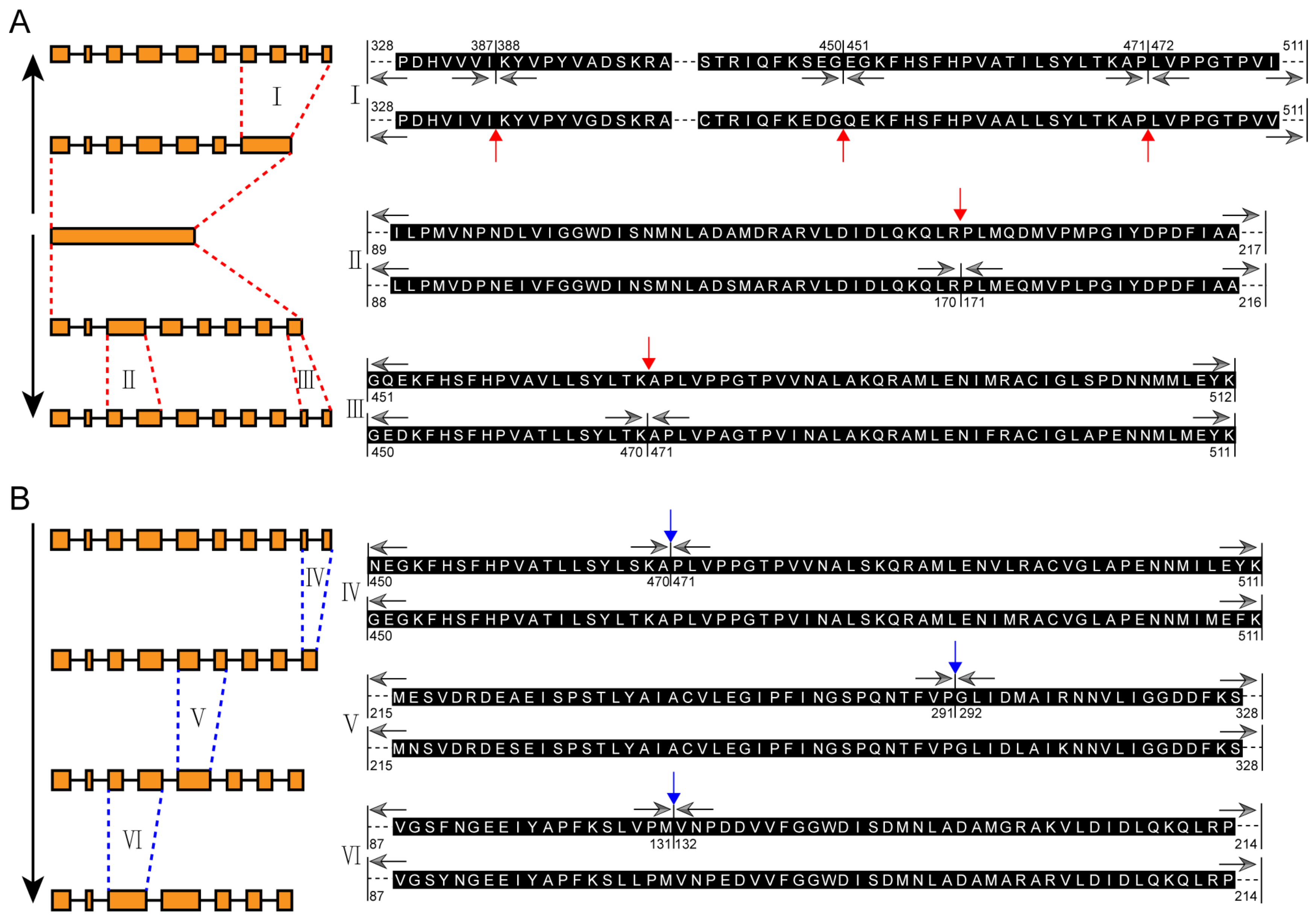
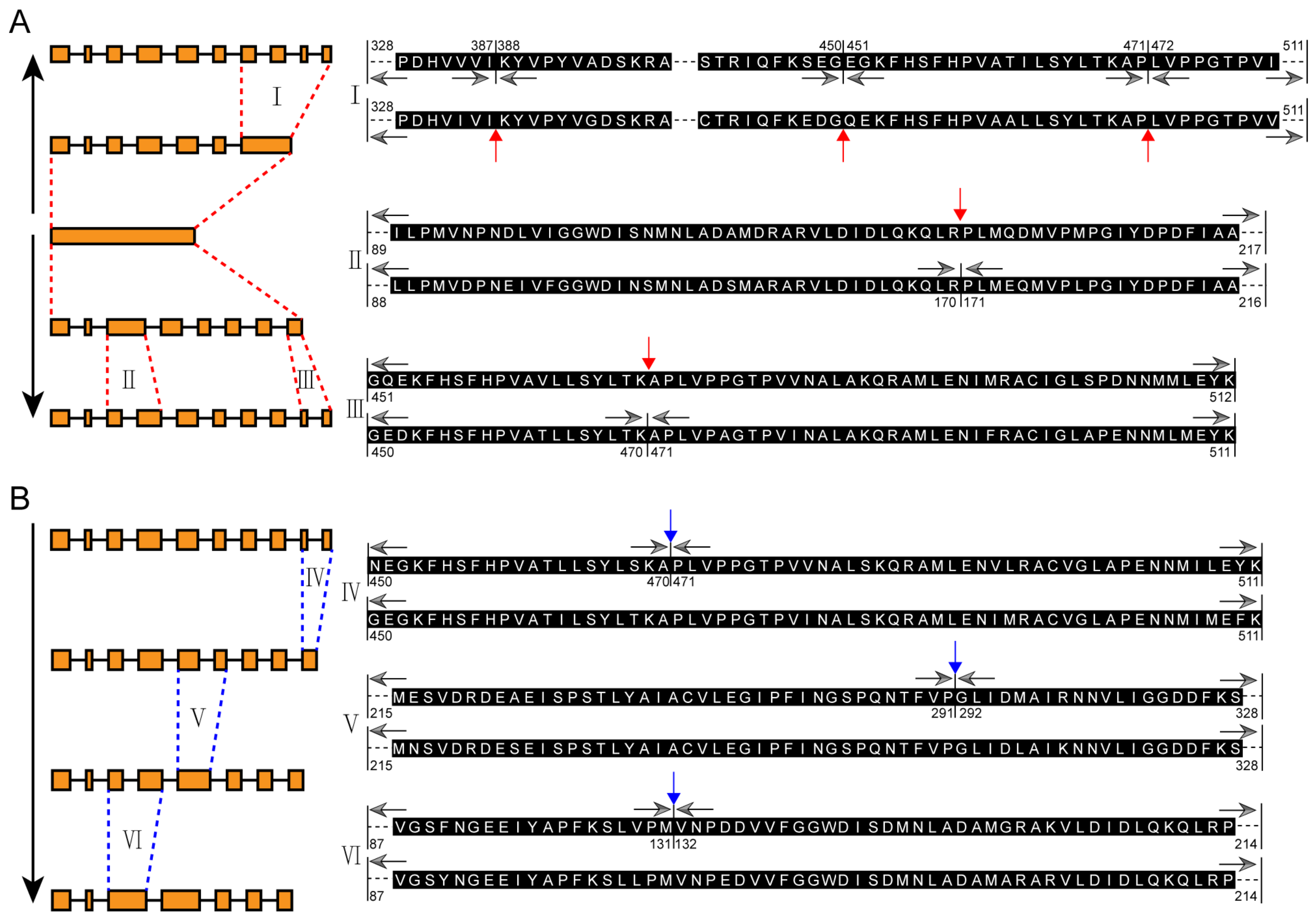
2.2. Exon-Intron Structure Evolution Analysis of MIPS Gene Family
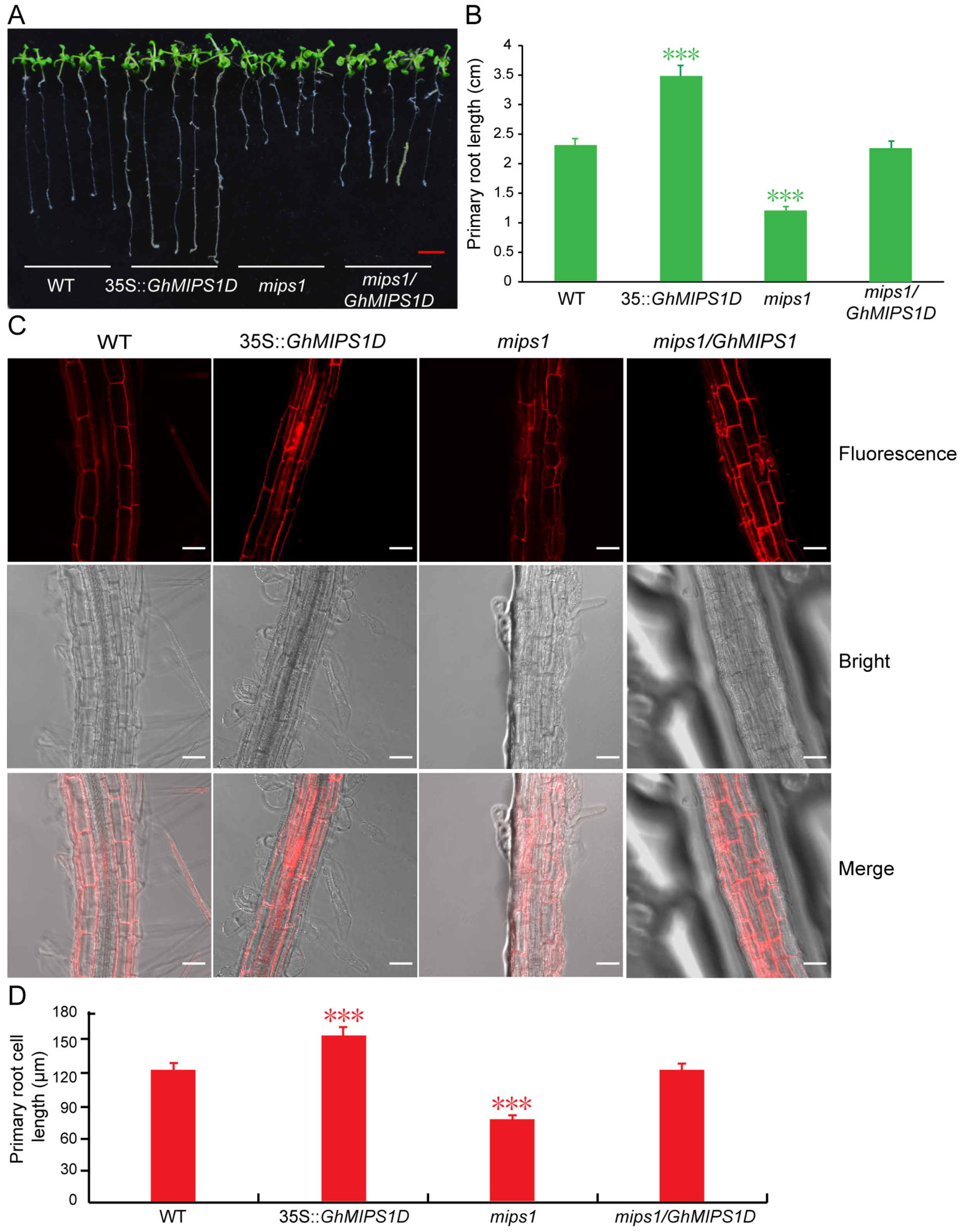
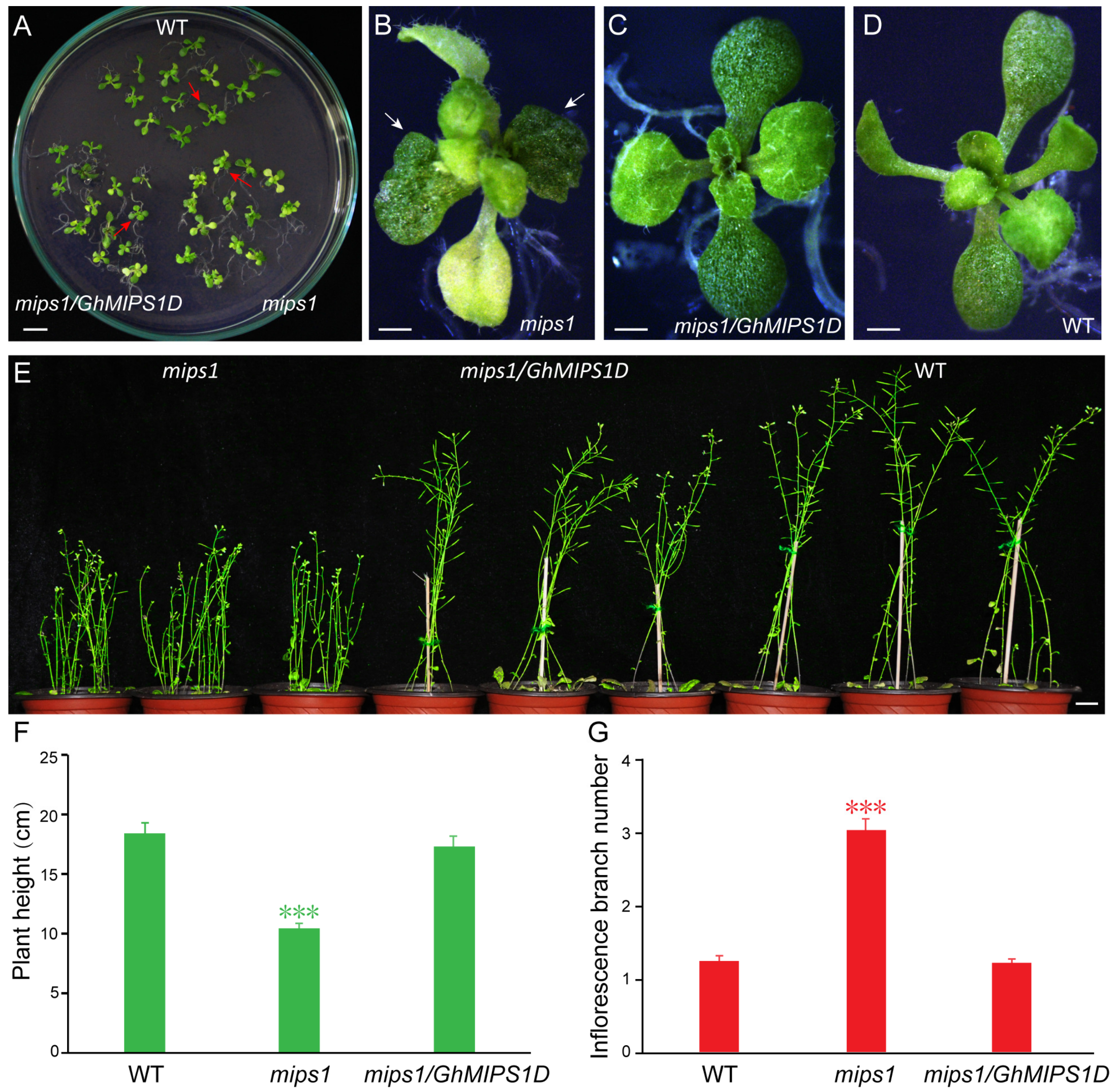
2.3. Functional Complementary Analysis of GhMIPS1D in the Loss-of-Function Arabidopsis mips1 Mutant
2.4. Ectopic Expression of GhMIPS1D Promotes Root Cell Elongation in Arabidopsis
3. Discussion
4. Materials and Methods
4.1. Sequence Acquirement and Chromosomal Distribution of GhMIPS Genes
4.2. Phylogenic and Evolutionary Analyses
4.3. Plant Materials
4.4. Construction of Vectors, Subcellular Localization Analysis and Ectopic Expression of GhMIPS1D in Arabidopsis
4.5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| APX | Ascorbate peroxidase |
| AsA | Ascorbic acid |
| BLAST | Basic local alignment search tool |
| DPA | Day post anthesis |
| G6P | d-glucose 6-phosphate |
| GlcA | d-glucuronic acid |
| IMP | Myo-inositol monophosphatase |
| Ins | Myo-inositol |
| IP | Myo-inositol polyphosphate |
| MEGA | Molecular evolutionary genetics analysis |
| MIPS | Myo-inositol-1-phosphate synthase |
| MS | Murashige and Skoog |
| PtdIns | phospholipid phosphatidylinositol |
| PtdInsP | phosphoinositide phosphate |
| RNAi | RNA interference |
| UDP-GlcA | UDP-d-glucuronic acid |
| WT | Wild-type |
References
- Majumder, A.L.; Chatterjee, A.; Dastidar, K.G.; Majee, M. Diversification and evolution of L-myo-inositol 1-phosphate synthase. FEBS Lett. 2003, 553, 3–10. [Google Scholar] [CrossRef]
- Majumder, A.L.; Johnson, M.D.; Henry, S.A. 1-L-myo-inositol-1-phosphate synthase. Biochim. Biophys. Acta 1997, 1348, 245–256. [Google Scholar] [CrossRef]
- Johnson, M.D.; Wang, X. Differentially expressed forms of 1-L-myo-inositolphosphate synthase (EC5.5.1.4) in Phaseolus vulgaris. J. Biol. Chem. 1996, 271, 17215–17218. [Google Scholar] [CrossRef] [PubMed]
- Hait, N.C.; Chaudhuri, A.R.; Das, A.; Bhattacharyya, S.; Majumder, A.L. Processing and activation of chloroplast L-myo-inositol 1-phosphate synthase from Oryza sativa requires signals from both light and salt. Plant Sci. 2002, 162, 559–568. [Google Scholar] [CrossRef]
- Abu-abied, M.; Holland, D. The gene c-ino1 from Citrus paradise is highly homologous to tur1 and ino1 from the yeast and Spirodela encoding for myo-inositol phosphate synthase. Plant Physiol. 1994, 106, 1689. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.D.; Sussex, I.M. 1-L-myo-inositol 1-phosphate synthase from Arabidopsis thaliana. Plant Physiol. 1995, 107, 613–619. [Google Scholar] [CrossRef] [PubMed]
- Keller, R.; Brearley, C.A.; Trethewey, R.N.; Muller-Rober, B. Reduced inositol content and altered morphology in transgenic potato plants inhibited for 1-D-myo-inositol 3-phosphate synthase. Plant J. 1998, 16, 403–410. [Google Scholar] [CrossRef]
- Larson, S.R.; Raboy, V. Linkage mapping of maize and barley myo-inositol 1-phosphate synthase DNA sequences: Correspondence with low phytic acid mutation. Theor. Appl. Genet. 1999, 99, 27–36. [Google Scholar] [CrossRef]
- Iqbal, M.; Afzal, A.; Yaegashi, S.; Ruben, E.; Triwitayakorn, K.; Njiti, V.; Ahsan, R.; Wood, A.; Lightfoot, D. A pyramid of loci for partial resistance to Fusarium solani f. sp. glycines maintains myo-inositol-1-phosphate synthase expression in soybean roots. Theor. Appl. Genet. 2002, 105, 1115–1123. [Google Scholar] [CrossRef] [PubMed]
- Chun, J.A.; Jin, U.H.; Lee, J.W.; Yi, Y.B.; Hyung, N.I.; Kang, M.H.; Pyee, J.H.; Suh, M.C.; Kang, C.W.; Seo, H.Y.; et al. Isolation and characterization of a myo-inositol 1-phosphate synthase cDNA from developing sesame (Sesamum indicum L.) seeds: Functional and differential expression, and salt-induced transcription during germination. Planta 2003, 216, 874–880. [Google Scholar] [PubMed]
- Bachhawat, N.; Mande, S.C. Complex evolution of the inositol-1-phosphate synthase gene among archaea and eubacteria. Trends Genet. 2000, 16, 111–113. [Google Scholar] [CrossRef]
- Hegeman, C.E.; Good, L.L.; Grabau, E.A. Expression of D-myo-inositol-3-phosphate synthase in soybean implication for phytic acid biosynthesis. Plant Physiol. 2001, 125, 1941–1948. [Google Scholar] [CrossRef] [PubMed]
- Gillaspy, G.E. The cellular language of myo-inositol signaling. New Phytol. 2011, 192, 823–839. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.J.; Zhou, C.; Yang, H.Y.; Roberts, M.F. Inositol 1-phosphate synthase from Archaeoglobus fulgidus is a class II aldolase. Biochemistry 2000, 39, 12415–12423. [Google Scholar] [CrossRef] [PubMed]
- Murphy, A.M.; Otto, B.; Brearley, C.A.; Carr, J.P.; Hanke, D.E. A role for inositol hexakisphosphate in the maintenance of basal resistance to plant pathogens. Plant J. 2008, 56, 638–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, P.H.; Raynaud, C.; Tcherkez, G.; Blanchet, S.; Massoud, K.; Domenichini, S.; Herry, Y.; Soubigou-Taconnat, L.; Lelarge-Trouverie, C.; Saindrenan, P.; et al. Crosstalks between myo-inositol metabolism, programmed cell death and basal immunity in Arabidopsis. PLoS ONE 2009, 4, e7364. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Qin, G.J.; Zhang, J.; Liang, Y.; Song, Y.Q.; Zhao, M.P.; Tsuge, T.; Aoyama, T.; Liu, J.J.; Gu, H.Y.; et al. D-myo-inositol-3-phosphate affects phosphatidylinositol-mediated endomembrane function in Arabidopsis and is essential for auxin-regulated embryogenesis. Plant Cell 2011, 23, 1352–1372. [Google Scholar] [CrossRef] [PubMed]
- Donahue, J.L.; Alford, S.R.; Torabinejad, J.; Kerwin, R.E.; Nourbakhsh, A.; Ray, W.K.; Hernick, M.; Huang, X.; Lyons, B.M.; Hein, P.P.; et al. The Arabidopsis myo-inositol 1-phosphate synthase1 gene is required for myo-inositol synthesis and suppression of cell death. Plant Cell 2010, 22, 888–903. [Google Scholar] [CrossRef] [PubMed]
- Raboy, V. Seeds for a better future: ‘Low phytate’ grains help to overcome malnutrition and reduce pollution. Trends Plant Sci. 2001, 6, 458–462. [Google Scholar] [CrossRef]
- Baig, M.M.; Kelly, S.; Loewus, F. L-ascorbic acid biosynthesis in higher plants from L-gulono-1, 4-lactone and L-galactono-1, 4-lactone. Plant Physiol. 1970, 46, 277–280. [Google Scholar] [CrossRef] [PubMed]
- Allison, J.H.; Stewart, M.A. Myo-inositol and ascorbic acid in developing rat brain. J. Neurochem. 1973, 20, 1785–1788. [Google Scholar] [CrossRef] [PubMed]
- Banhegyi, G.; Braun, L.; Csala, M.; Puskás, F.; Mandl, J. Ascorbate metabolism and its regulation in animals. Free Radic. Biol. Med. 1997, 23, 793–803. [Google Scholar] [CrossRef]
- Loewus, F. Inositol metabolism and cell wall formation in plants. Fed. Proc. 1965, 24, 855–862. [Google Scholar] [PubMed]
- Loewus, F.A. Inositol and plant cell wall polysaccharide biogenesis. Subcell. Biochem. 2006, 39, 21–45. [Google Scholar] [PubMed]
- Pang, C.Y.; Wang, H.; Pang, Y.; Xu, C.; Jiao, Y.; Qin, Y.M.; Western, T.L.; Yu, S.X.; Zhu, Y.X. Comparative proteomics indicates that biosynthesis of pectic precursors is important for cotton fiber and Arabidopsis root hair elongation. Mol. Cell Proteom. 2010, 9, 2019–2033. [Google Scholar] [CrossRef] [PubMed]
- Li, H.B.; Qin, Y.M.; Pang, Y.; Song, W.Q.; Mei, W.Q.; Zhu, Y.X. A cotton ascorbate peroxidase is involved in hydrogen peroxide homeostasis during fibre cell development. New Phytol. 2007, 175, 462–471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tao, C.C.; Jin, X.; Zhu, L.P.; Xie, Q.L.; Wang, X.C.; Li, H.B. Genome-wide investigation and expression profiling of APX gene family in Gossypium hirsutum provide new insights in redox homeostasis maintenance during different fiber development stages. Mol. Genet. Genom. 2018, 293, 685–697. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.J.; Xiao, G.H.; Liu, N.J.; Liu, D.; Chen, P.S.; Qin, Y.M.; Zhu, Y.X. Targeted Lipidomics Studies Reveal that Linolenic Acid Promotes Cotton Fiber Elongation by Activating Phosphatidylinositol and Phosphatidylinositol Monophosphate Biosynthesis. Mol. Plant 2015, 8, 911–921. [Google Scholar] [CrossRef] [PubMed]
- Kusano, H.; Testerink, C.; Vermeer, J.E.; Tsuge, T.; Shimada, H.; Oka, A.; Munnik, T.; Aoyama, T. The Arabidopsis Phosphatidylinositol Phosphate 5-Kinase PIP5K3 is a key regulator of root hair tip growth. Plant Cell 2008, 20, 367–380. [Google Scholar] [CrossRef] [PubMed]
- Stenzel, I.; Ischebeck, T.; König, S.; Hołubowska, A.; Sporysz, M.; Hause, B.; Heilmann, I. The type B phosphatidylinositol-4-phosphate 5-kinase 3 is essential for root hair formation in Arabidopsis thaliana. Plant Cell 2008, 20, 124–141. [Google Scholar] [CrossRef] [PubMed]
- Abid, G.; Silue, S.; Muhovski, Y.; Jacquemin, J.M.; Toussaint, A.; Baudoin, J.P. Role of myo-inositol phosphate synthase and sucrose synthase genes in plant seed development. Gene 2009, 439, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Cui, M.; Liang, D.; Wu, S.; Ma, F.W.; Lei, Y.S. Isolation and developmental expression analysis of L-myo-inositol-1-1phosphate synthase in four Actinidia species. Plant Physiol. Biochem. 2013, 73, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Basak, P.; Maitra-Majee, S.; Das, J.K.; Mukherjee, A.; Ghosh Dastidar, S.; Pal Choudhury, P.; Lahiri Majumder, A. An evolutionary analysis identifies a conserved pentapeptide stretch containing the two essential lysine residues for rice L-myo-inositol 1-phosphate synthase catalytic activity. PLoS ONE 2017, 12, e0185351. [Google Scholar] [CrossRef] [PubMed]
- Fleet, C.M.; Yen, J.Y.; Hill, E.A.; Gillaspy, G.E. Co-suppression of AtMIPS demonstrates cooperation of MIPS1, MIPS2 and MIPS3 in maintaining myo-inositol synthesis. Plant Mol. Biol. 2018, 97, 253–263. [Google Scholar] [CrossRef] [PubMed]
- Nunes, A.C.; Vianna, G.R.; Cuneo, F.; Amaya-Farfán, J.; de Capdeville, G.; Rech, E.L.; Aragão, F.J. RNAi-mediated silencing of the myo-inositol-1-phosphate synthase gene (GmMIPS1) in transgenic soybean inhibited seed development and reduced phytate content. Planta 2006, 224, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Zhai, H.; Wang, F.B.; Si, Z.Z.; Huo, J.X.; Xing, L.; An, Y.Y.; He, S.Z.; Liu, Q.C. A myo-inositol-1-phosphate synthase gene, IbMIPS1, enhances salt and drought tolerance and stem nematode resistance in transgenic sweet potato. Plant Biotechnol. J. 2016, 14, 592–602. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.L.; Wang, C.Y.; Xiang, B.; Han, R.H.; Guo, Z.F. Hydrogen peroxide and nitric oxide mediated cold- and dehydration-induced myo-inositol phosphate synthase that confers multiple resistances to abiotic stresses. Plant Cell Environ. 2013, 36, 288–299. [Google Scholar] [CrossRef] [PubMed]
- Latrasse, D.; Jégu, T.; Meng, P.H.; Mazubert, C.; Hudik, E.; Delarue, M.; Charon, C.; Crespi, M.; Hirt, H.; Raynaud, C.; et al. Dual function of MIPS1 as a metabolic enzyme and transcriptional regulator. Nucleic Acids Res. 2013, 41, 2907–2917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valluru, R.; Van den Ende, W. Myo-inositol and beyond-emerging networks under stress. Plant Sci. 2011, 181, 387–400. [Google Scholar] [CrossRef] [PubMed]
- Stevenson-Paulik, J.; Bastidas, R.J.; Chiou, S.T.; Frye, R.A.; York, J.D. Generation of phytate-free seeds in Arabidopsis through disruption of inositol polyphosphate kinases. Proc. Natl. Acad. Sci. USA 2005, 102, 12612–12617. [Google Scholar] [CrossRef] [PubMed]
- Ischebeck, T.; Stenzel, I.; Heilmann, I. Type B phosphatidylinositol-4-phosphate 5-kinases mediate Arabidopsis and Nicotiana tabacum pollen tube growth by regulating apical pectin secretion. Plant Cell 2008, 20, 3312–3330. [Google Scholar] [CrossRef] [PubMed]
- Kusuda, H.; Koga, W.; Kusano, M.; Oikawa, A.; Saito, K.; Hirai, M.Y.; Yoshida, K.T. Ectopic expression of myo-inositol 3-phosphate synthase induces a wide range of metabolic changes and confers salt tolerance in rice. Plant Sci. 2015, 232, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Zheng, J.Y.; Long, H.; Xiao, Y.H.; Yan, X.Y.; Pei, Y. Auxin regulates cotton fiber initiation via GhPIN-mediated auxin transport. Plant Cell Physiol. 2017, 58, 385–397. [Google Scholar] [CrossRef] [PubMed]
- Samuel, Y.S.; Cheung, F.; Lee, J.J.; Ha, M.; Wei, N.E.; Sze, S.H.; Stelly, D.M.; Thaxton, P.; Triplett, B.; Town, C.D.; et al. Accumulation of genome-specific transcripts, transcription factors and phytohormonal regulators during early stages of fiber cell development in allotetraploid cotton. Plant J. 2006, 47, 761–765. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Xiong, L.M. myo-Inositol-1-phosphate synthase is required for polar auxin transport and organ development. J. Biol. Chem. 2010, 285, 24238–24247. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; Zhu, L.P.; Yao, Q.; Meng, X.R.; Ding, G.H.; Wang, D.; Xie, Q.L.; Tong, Z.; Tao, C.C.; Yu, L.; et al. Expression profiling of mitogen-activated protein kinase genes reveals their evolutionary and functional diversity in different rubber tree (Hevea brasiliensis) cultivars. Genes 2017, 8, 261. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.P.; Jin, X.; Xie, Q.L.; Yao, Q.; Wang, X.C.; Li, H.B. Calcium-dependant protein kinase family genes involved in ethylene-induced natural rubber production in different Hevea brasiliensis cultivars. Int. J. Mol. Sci. 2018, 19, 947. [Google Scholar] [CrossRef] [PubMed]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sánchez-Gracia, A. DnaSP 6: DNA sequence polymorphism analysis of large datasets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Xin, S.; Tao, C.C.; Jin, X.; Li, H.B. Cotton ascorbate oxidase promotes cell growth in cultured tobacco bright yellow-2 cells through generation of apoplast oxidation. Int. J. Mol. Sci. 2017, 18, 1346. [Google Scholar] [CrossRef] [PubMed]




© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, R.; Song, W.; Wang, F.; Cao, A.; Xie, S.; Chen, X.; Jin, X.; Li, H. A Cotton (Gossypium hirsutum) Myo-Inositol-1-Phosphate Synthase (GhMIPS1D) Gene Promotes Root Cell Elongation in Arabidopsis. Int. J. Mol. Sci. 2019, 20, 1224. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20051224
Ma R, Song W, Wang F, Cao A, Xie S, Chen X, Jin X, Li H. A Cotton (Gossypium hirsutum) Myo-Inositol-1-Phosphate Synthase (GhMIPS1D) Gene Promotes Root Cell Elongation in Arabidopsis. International Journal of Molecular Sciences. 2019; 20(5):1224. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20051224
Chicago/Turabian StyleMa, Rendi, Wangyang Song, Fei Wang, Aiping Cao, Shuangquan Xie, Xifeng Chen, Xiang Jin, and Hongbin Li. 2019. "A Cotton (Gossypium hirsutum) Myo-Inositol-1-Phosphate Synthase (GhMIPS1D) Gene Promotes Root Cell Elongation in Arabidopsis" International Journal of Molecular Sciences 20, no. 5: 1224. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20051224