Discovery of Galangin as a Potential DPP-4 Inhibitor That Improves Insulin-Stimulated Skeletal Muscle Glucose Uptake: A Combinational Therapy for Diabetes

 , , and
, , and 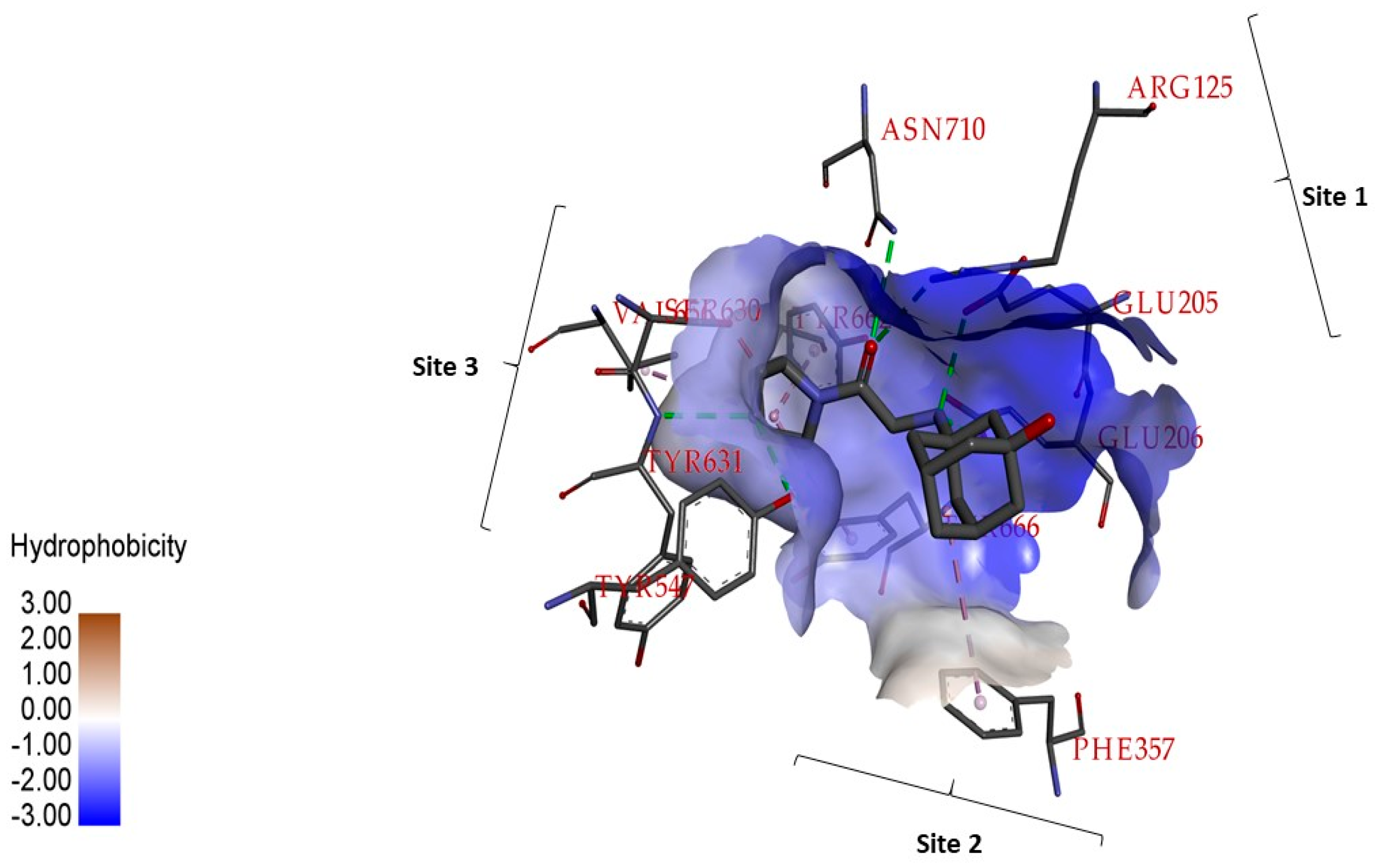{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
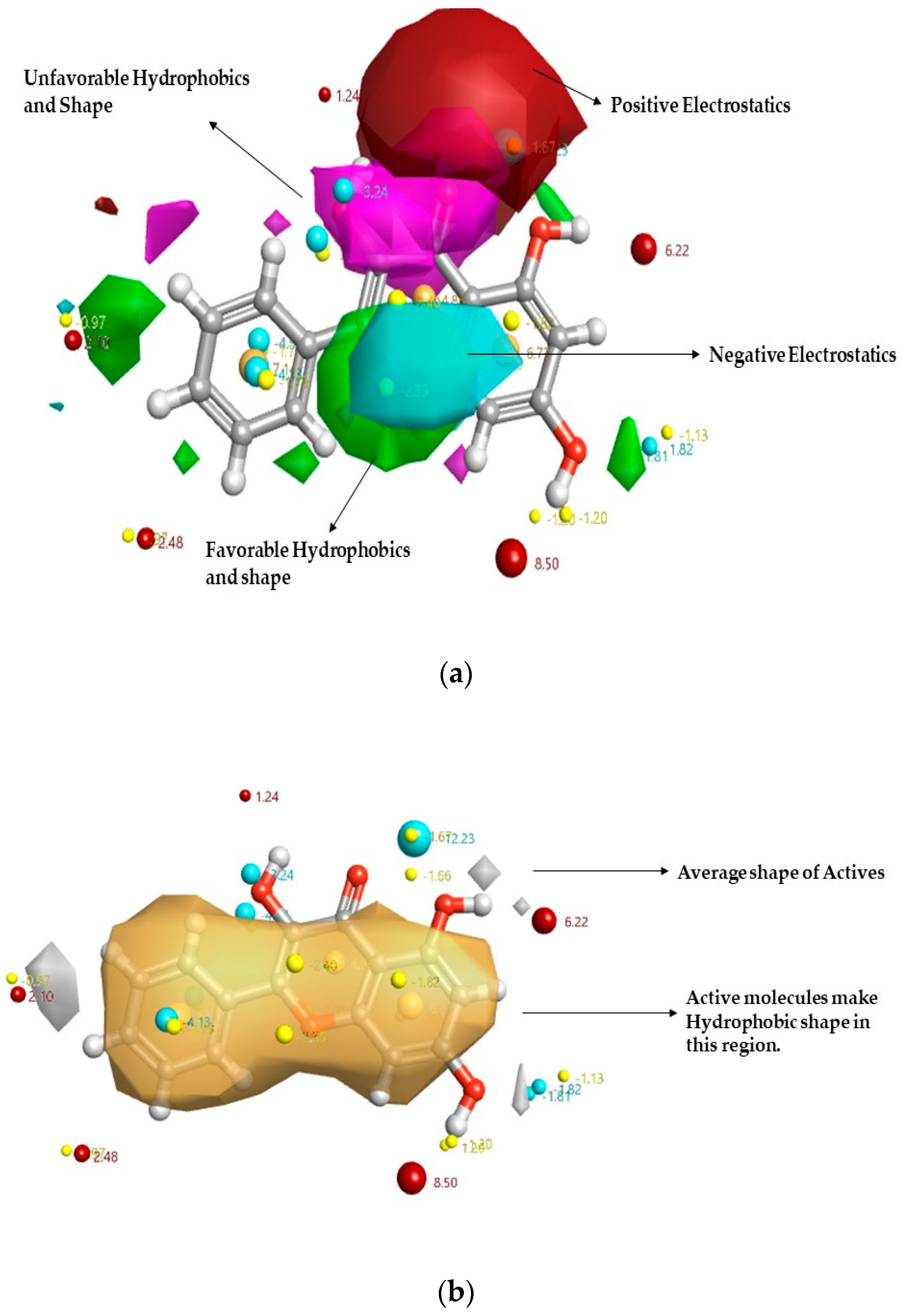
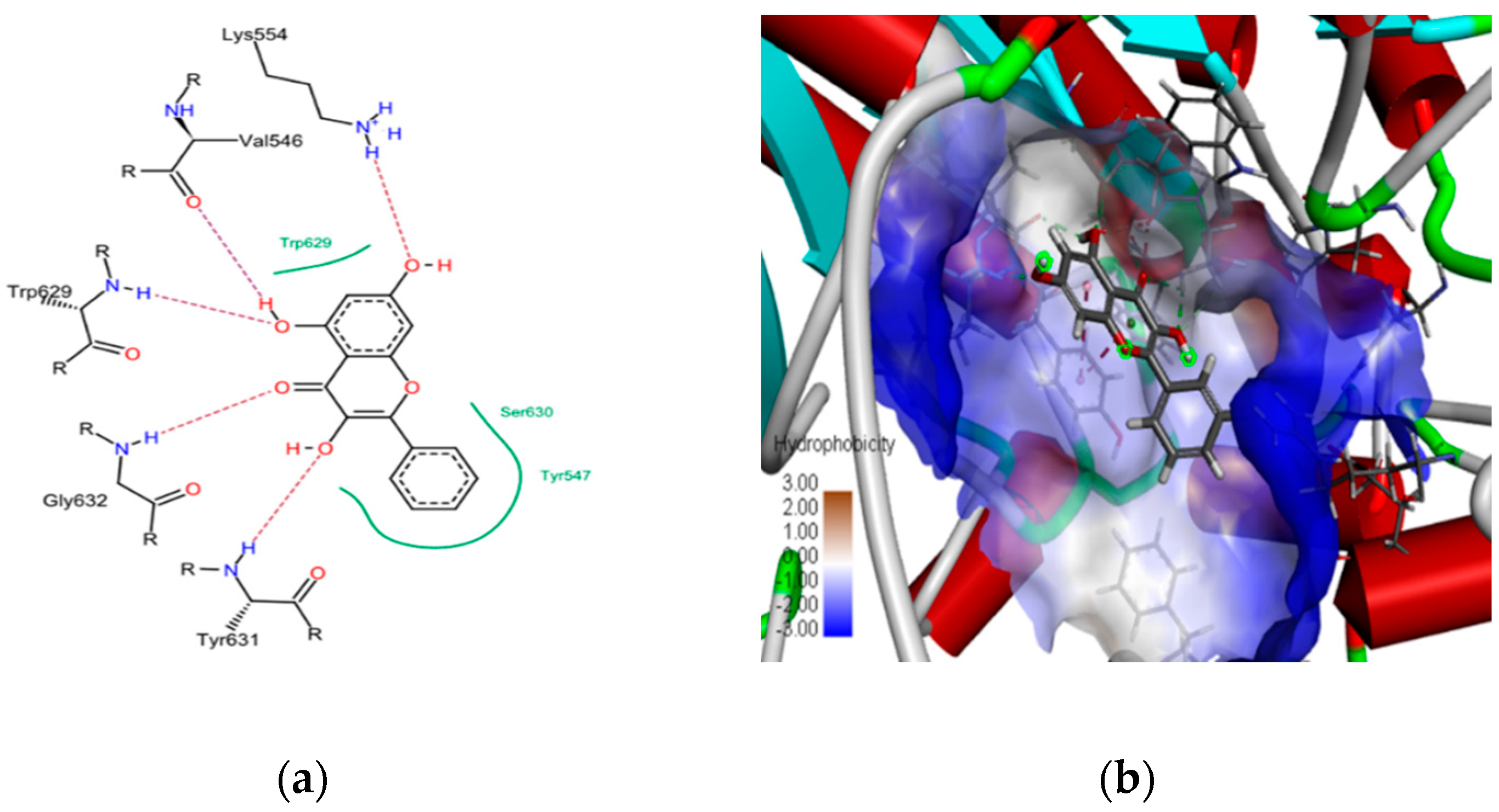
2.1. In Silico Studies on DPP-4 Enzyme Interactions with Flavanol Galangin
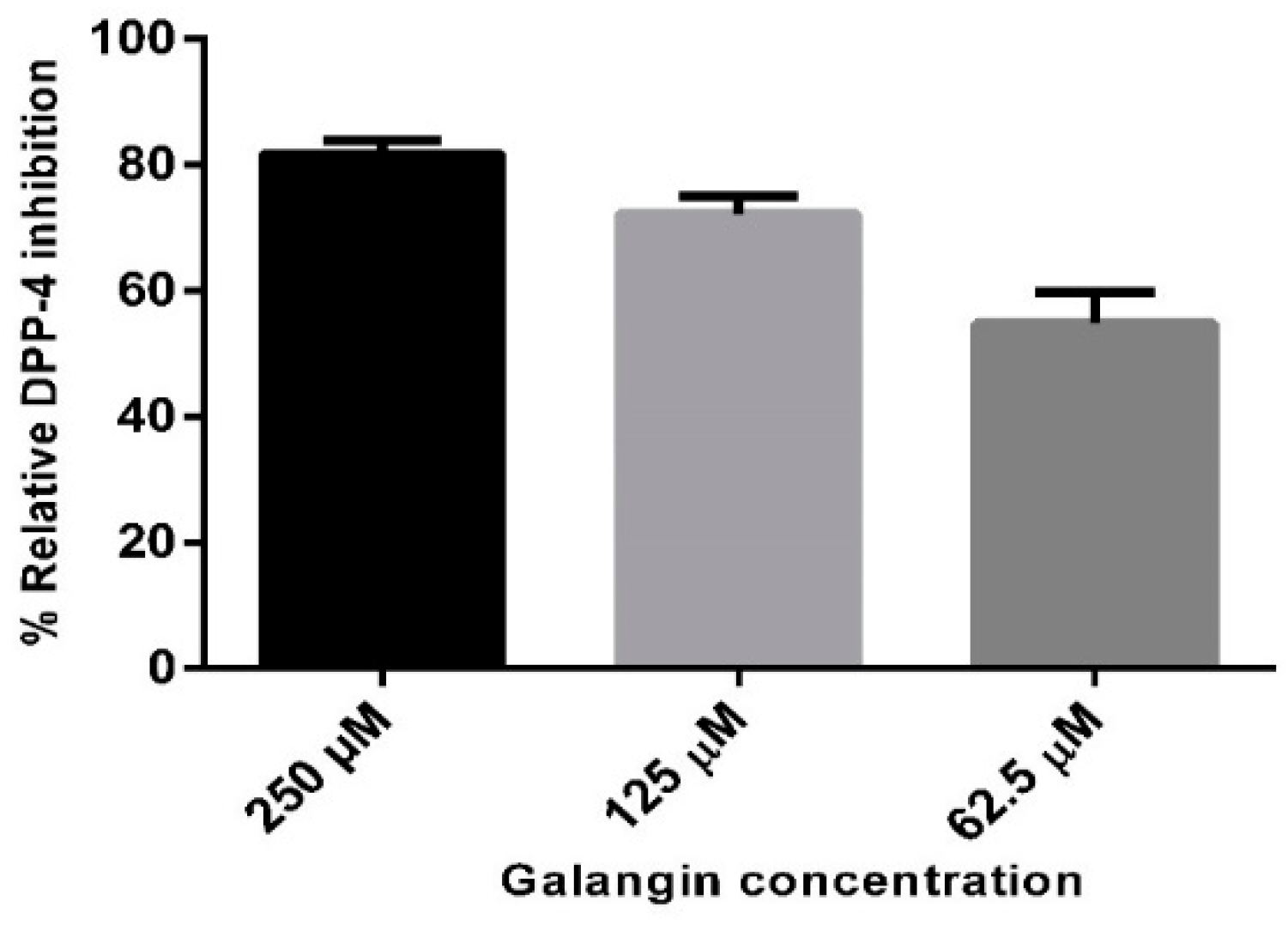
2.2. In Vitro Studies on Validation of as Galangin Inhibiting DPP-4 Enzyme
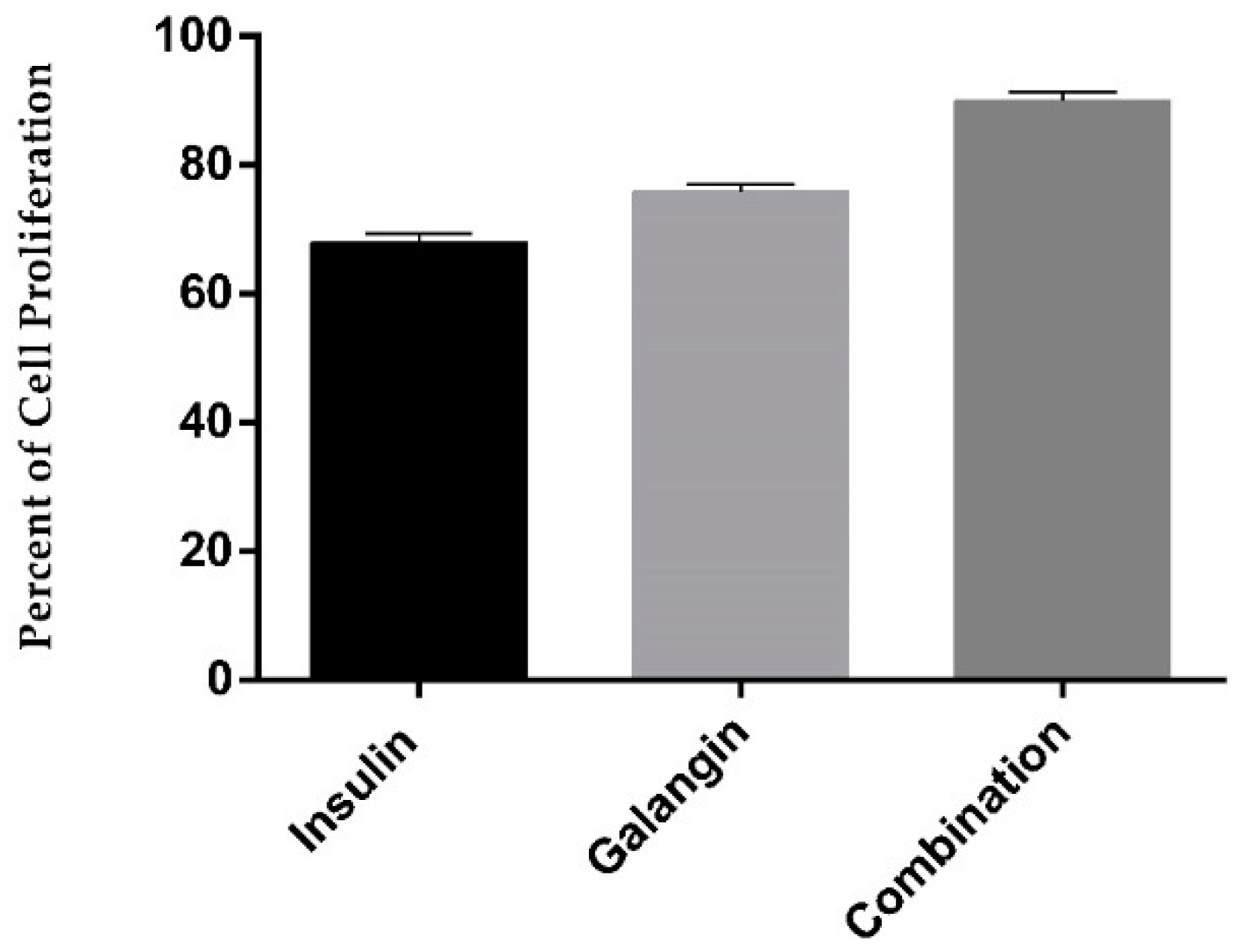
2.3. Galangin in Combination with Insulin Promotes Differentiated Skeletal Muscle Cell Proliferation
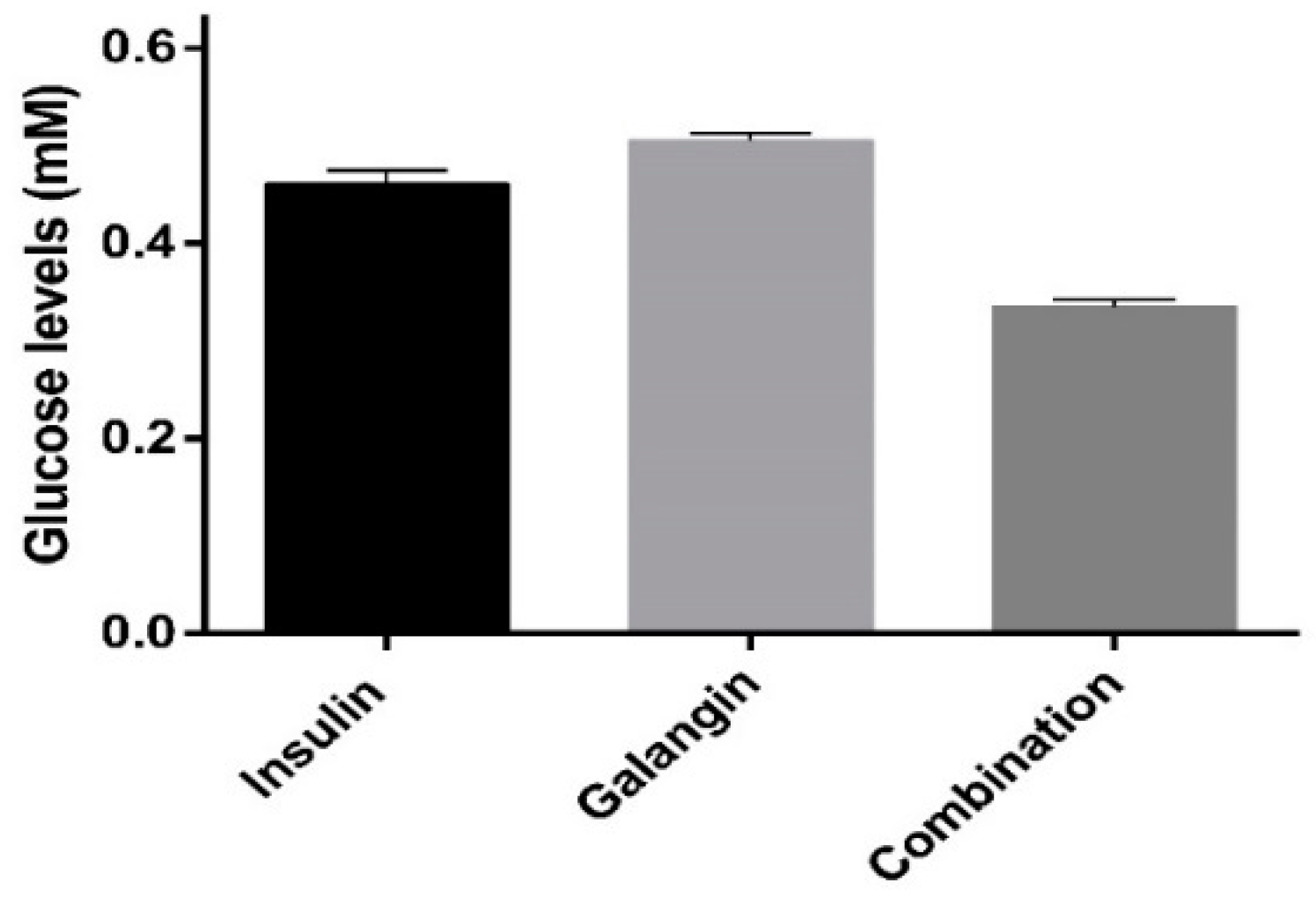
2.4. Glucose Metabolism
3. Materials and Methods
3.1. Chemicals and Reagents
3.2. Molecular Docking Simulations
3.3. In Vitro DPP-4 Inhibition Assay
3.4. Cell Culture
3.5. Cell Proliferation Assay
3.6. Glucose Quantification in Cell Supernatant Using 1H-NMR Spectroscopy
3.7. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ogurtsova, K.; da Rocha Fernandes, J.D.; Huang, Y.; Linnenkamp, U.; Guariguata, L.; Cho, N.H.; Cavan, D.; Shaw, J.E.; Makaroff, L.E. IDF Diabetes Atlas: Global estimates for the prevalence of diabetes for 2015 and 2040. Diabetes Res. Clin. Pract. 2017, 128, 40–50. [Google Scholar] [CrossRef]
- American Diabetes Association. 2. Classification and Diagnosis of Diabetes: Standards of Medical Care in Diabetes—2018. Diabetes Care 2018, 41, S13–S27. [Google Scholar] [CrossRef] [PubMed]
- La Greca, A.M.; MacKey, E.R. Type 1 diabetes mellitus. In Behavioral Approaches to Chronic Disease in Adolescence: A Guide to Integrative Care; Springer: New York, NY, USA, 2009; ISBN 9780387876863. [Google Scholar]
- Chaudhury, A.; Duvoor, C.; Reddy Dendi, V.S.; Kraleti, S.; Chada, A.; Ravilla, R.; Marco, A.; Shekhawat, N.S.; Montales, M.T.; Kuriakose, K.; et al. Clinical Review of Antidiabetic Drugs: Implications for Type 2 Diabetes Mellitus Management. Front. Endocrinol. 2017, 8, 6. [Google Scholar] [CrossRef]
- Alhadramy, M.S. Diabetes and oral therapies a review of oral therapies for diabetes mellitus. J. Taibah Univ. Med. Sci. 2016, 11, 317–329. [Google Scholar] [CrossRef]
- Milder, T.; Stocker, S.; Abdel Shaheed, C.; McGrath-Cadell, L.; Samocha-Bonet, D.; Greenfield, J.; Day, R. Combination Therapy with an SGLT2 Inhibitor as Initial Treatment for Type 2 Diabetes: A Systematic Review and Meta-Analysis. J. Clin. Med. 2019, 8, 45. [Google Scholar] [CrossRef]
- Moon, M.K.; Hur, K.Y.; Ko, S.H.; Park, S.O.; Lee, B.W.; Kim, J.H.; Rhee, S.Y.; Kim, H.J.; Choi, K.M.; Kim, N.H. Combination therapy of oral hypoglycemic agents in patients with type 2 diabetes mellitus. Korean J. Intern. Med. 2017, 32, 974–983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Massi-Benedetti, M.; Orsini-Federici, M. Treatment of type 2 diabetes with combined therapy: What are the pros and cons? Diabetes Care 2008, 31 (Suppl. 2), S131–S135. [Google Scholar] [CrossRef]
- Rivas, D.A.; Fielding, R.A. Skeletal Muscle. In Encyclopedia of Human Nutrition; Academic Press: Cambridge, MA, USA, 2012; ISBN 9780123848857. [Google Scholar]
- Lee, M.J.; Kim, E.H.; Bae, S.J.; Choe, J.; Jung, C.H.; Lee, W.J.; Kim, H.K. Protective role of skeletal muscle mass against progression from metabolically healthy to unhealthy phenotype. Clin. Endocrinol. 2018, 90, 102–113. [Google Scholar] [CrossRef]
- Malin, S.K.; Huang, H.; Mulya, A.; Kashyap, S.R.; Kirwan, J.P. Lower dipeptidyl peptidase-4 following exercise training plus weight loss is related to increased insulin sensitivity in adults with metabolic syndrome. Peptides 2013, 47, 142–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Röhrborn, D.; Wronkowitz, N.; Eckel, J. DPP4 in diabetes. Front. Immunol. 2015, 6, 386. [Google Scholar] [PubMed]
- Monami, M.; Iacomelli, I.; Marchionni, N.; Mannucci, E. Dipeptydil peptidase-4 inhibitors in type 2 diabetes: A meta-analysis of randomized clinical trials. Nutr. Metab. Cardiovasc. Dis. 2010, 20, 224–235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Todd, J.F.; Bloom, S.R. Incretins and other peptides in the treatment of diabetes. Diabet. Med. 2007, 24, 223–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Hu, Y.; Liu, T. Recent Advances in Non-Peptidomimetic Dipeptidyl Peptidase 4 Inhibitors: Medicinal Chemistry and Preclinical Aspects. Curr. Med. Chem. 2012, 19, 3982–3999. [Google Scholar] [CrossRef]
- Zettl, H.; Schubert-Zsilavecz, M.; Steinhilber, D. Medicinal chemistry of incretin mimetics and DPP-4 inhibitors. ChemMedChem 2010, 5, 179–185. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, B.; Hennig, M.; Mattei, P. Molecular Recognition of Ligands in Dipeptidyl Peptidase IV. Curr. Med. Chem. 2007, 7, 609–620. [Google Scholar] [CrossRef]
- Pissurlenkar, R.R.S.; Shaikh, M.S.; Coutinho, E.C. 3D-QSAR studies of Dipeptidyl peptidase IV inhibitors using a docking based alignment. J. Mol. Model. 2007, 13, 1047–1071. [Google Scholar] [CrossRef] [PubMed]
- Meduru, H.; Wang, Y.T.; Tsai, J.J.P.; Chen, Y.C. Finding a potential dipeptidyl peptidase-4 (DPP-4) inhibitor for type-2 diabetes treatment based on molecular docking, pharmacophore generation, and molecular dynamics simulation. Int. J. Mol. Sci. 2016, 17, 920. [Google Scholar] [CrossRef] [PubMed]
- Lovshin, J.A.; Drucker, D.J. Incretin-based therapies for type 2 diabetes mellitus. Nat. Rev. Endocrinol. 2009, 5, 262–269. [Google Scholar] [CrossRef] [PubMed]
- Thomsen, R.W.; Pedersen, L.; Møller, N.; Kahlert, J.; Beck-Nielsen, H.; Sørensen, H.T. Incretin-based therapy and risk of acute pancreatitis: A nationwide population-based case-control study. Diabetes Care 2015, 38, 1089–1098. [Google Scholar] [CrossRef] [PubMed]
- Cragg, G.M.; Newman, D.J. Natural products: A continuing source of novel drug leads. Biochim. Biophys. Acta-Gen. Subj. 2013, 1830, 3670–3695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cragg, G.M.; Newman, D.J.; Snader, K.M. Natural products in drug discovery and development. J. Nat. Prod. 1997, 60, 52–60. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Demain, A.L. Natural Products: Drug Discovery and Therapeutic Medicine; Humana Press: New York, NY, USA, 2005; ISBN 9781588293831. [Google Scholar]
- Rienks, J.; Barbaresko, J.; Oluwagbemigun, K.; Schmid, M.; Nöthlings, U. Polyphenol exposure and risk of type 2 diabetes: Dose-response meta-analyses and systematic review of prospective cohort studies. Am. J. Clin. Nutr. 2018, 108, 49–61. [Google Scholar] [CrossRef] [PubMed]
- Unnikrishnan, M.K.; Veerapur, V.; Nayak, Y.; Mudgal, P.P.; Mathew, G. Antidiabetic, Antihyperlipidemic and Antioxidant Effects of the Flavonoids. In Polyphenols in Human Health and Disease; Academic Press: Cambridge, MA, USA, 2013; ISBN 9780123984562. [Google Scholar]
- Fantini, M.; Benvenuto, M.; Masuelli, L.; Frajese, G.V.; Tresoldi, I.; Modesti, A.; Bei, R. In vitro and in vivo antitumoral effects of combinations of polyphenols, or polyphenols and anticancer drugs: Perspectives on cancer treatment. Int. J. Mol. Sci. 2015, 16, 9236–9282. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Narayanan, S.; Chang, K.O. Inhibition of influenza virus replication by plant-derived isoquercetin. Antivir. Res. 2010, 88, 227–235. [Google Scholar] [CrossRef] [PubMed]
- Daglia, M. Polyphenols as antimicrobial agents. Curr. Opin. Biotechnol. 2012, 23, 174–181. [Google Scholar] [CrossRef] [PubMed]
- Khurana, S.; Venkataraman, K.; Hollingsworth, A.; Piche, M.; Tai, T.C. Polyphenols: Benefits to the cardiovascular system in health and in aging. Nutrients 2013, 5, 3779–3827. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Durillo, F.; Segarra, A.; Villarejo, A.; Ramírez-Sánchez, M.; Prieto, I. Influence of Diet and Gender on Plasma DPP4 Activity and GLP-1 in Patients with Metabolic Syndrome: An Experimental Pilot Study. Molecules 2018, 23, 1564. [Google Scholar] [CrossRef] [PubMed]
- Kalhotra, P.; Chittepu, V.; Osorio-Revilla, G.; Gallardo-Velázquez, T. Structure-Activity Relationship and Molecular Docking of Natural Product Library Reveal Chrysin as a Novel Dipeptidyl Peptidase-4 (DPP-4) Inhibitor: An Integrated In Silico and In Vitro Study. Molecules 2018, 23, 1368. [Google Scholar] [CrossRef] [PubMed]
- Sato, H.; Kubota, N.; Kubota, T.; Takamoto, I.; Iwayama, K.; Tokuyama, K.; Moroi, M.; Sugi, K.; Nakaya, K.; Goto, M.; et al. Anagliptin increases insulin-induced skeletal muscle glucose uptake via an NO-dependent mechanism in mice. Diabetologia 2016, 59, 2426–2434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kutoh, E. Sitagliptin is effective and safe as add-on to insulin in patients with absolute insulin deficiency: A case series. J. Med. Case Rep. 2011, 5, 117. [Google Scholar] [CrossRef] [PubMed]
- Shankar, R.R.; Bao, Y.; Han, P.; Hu, J.; Ma, J.; Peng, Y.; Wu, F.; Xu, L.; Engel, S.S.; Jia, W. Sitagliptin added to stable insulin therapy with or without metformin in Chinese patients with type 2 diabetes. J. Diabetes Investig. 2017, 8, 321–329. [Google Scholar] [CrossRef] [PubMed]
- Aloud, A.A.; Chinnadurai, V.; Govindasamy, C.; Alsaif, M.A.; Al-Numair, K.S. Galangin, a dietary flavonoid, ameliorates hyperglycaemia and lipid abnormalities in rats with streptozotocin-induced hyperglycaemia. Pharm. Biol. 2018, 56, 302–308. [Google Scholar] [CrossRef]
- Aloud, A.A.; Veeramani, C.; Govindasamy, C.; Alsaif, M.A.; El Newehy, A.S.; Al-Numair, K.S. Galangin, a dietary flavonoid, improves antioxidant status and reduces hyperglycemia-mediated oxidative stress in streptozotocin-induced diabetic rats. Redox Rep. 2017, 22, 290–300. [Google Scholar] [CrossRef] [PubMed]
- Berman, H.M.; Battistuz, T.; Bhat, T.N.; Bluhm, W.F.; Bourne, P.E.; Burkhardt, K.; Feng, Z.; Gilliland, G.L.; Iype, L.; Jain, S.; et al. The protein data bank. Acta Crystallogr. Sect. D Biol. Crystallogr. 2002, 58, 899–907. [Google Scholar] [CrossRef]
- Kramer, B.; Rarey, M.; Lengauer, T. Evaluation of the FlexX incremental construction algorithm for protein-ligand docking. Proteins Struct. Funct. Genet. 1999, 37, 228–241. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, N.; Sato, T.; Kawasaki, K.; Murosaki, S.; Yamamoto, Y. A nonradioisotope, enzymatic assay for 2-deoxyglucose uptake in L6 skeletal muscle cells cultured in a 96-well microplate. Anal. Biochem. 2006, 351, 139–145. [Google Scholar] [CrossRef]
- Chittepu, V.; Kalhotra, P.; Gallardo-Velázquez, T.; Robles-de la Torre, R.; Osorio-Revilla, G.; Chittepu, V.C.S.R.; Kalhotra, P.; Gallardo-Velázquez, T.; Robles-de la Torre, R.R.; Osorio-Revilla, G. Designed Functional Dispersion for Insulin Protection from Pepsin Degradation and Skeletal Muscle Cell Proliferation: In Silico and In Vitro Study. Nanomaterials 2018, 8, 852. [Google Scholar] [CrossRef] [PubMed]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kalhotra, P.; Chittepu, V.C.S.R.; Osorio-Revilla, G.; Gallardo-Velázquez, T. Discovery of Galangin as a Potential DPP-4 Inhibitor That Improves Insulin-Stimulated Skeletal Muscle Glucose Uptake: A Combinational Therapy for Diabetes. Int. J. Mol. Sci. 2019, 20, 1228. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20051228
Kalhotra P, Chittepu VCSR, Osorio-Revilla G, Gallardo-Velázquez T. Discovery of Galangin as a Potential DPP-4 Inhibitor That Improves Insulin-Stimulated Skeletal Muscle Glucose Uptake: A Combinational Therapy for Diabetes. International Journal of Molecular Sciences. 2019; 20(5):1228. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20051228
Chicago/Turabian StyleKalhotra, Poonam, Veera C. S. R. Chittepu, Guillermo Osorio-Revilla, and Tzayhri Gallardo-Velázquez. 2019. "Discovery of Galangin as a Potential DPP-4 Inhibitor That Improves Insulin-Stimulated Skeletal Muscle Glucose Uptake: A Combinational Therapy for Diabetes" International Journal of Molecular Sciences 20, no. 5: 1228. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20051228