Bioavailability and In Vivo Antioxidant Activity of a Standardized Polyphenol Mixture Extracted from Brown Propolis


Abstract
:1. Introduction
2. Results
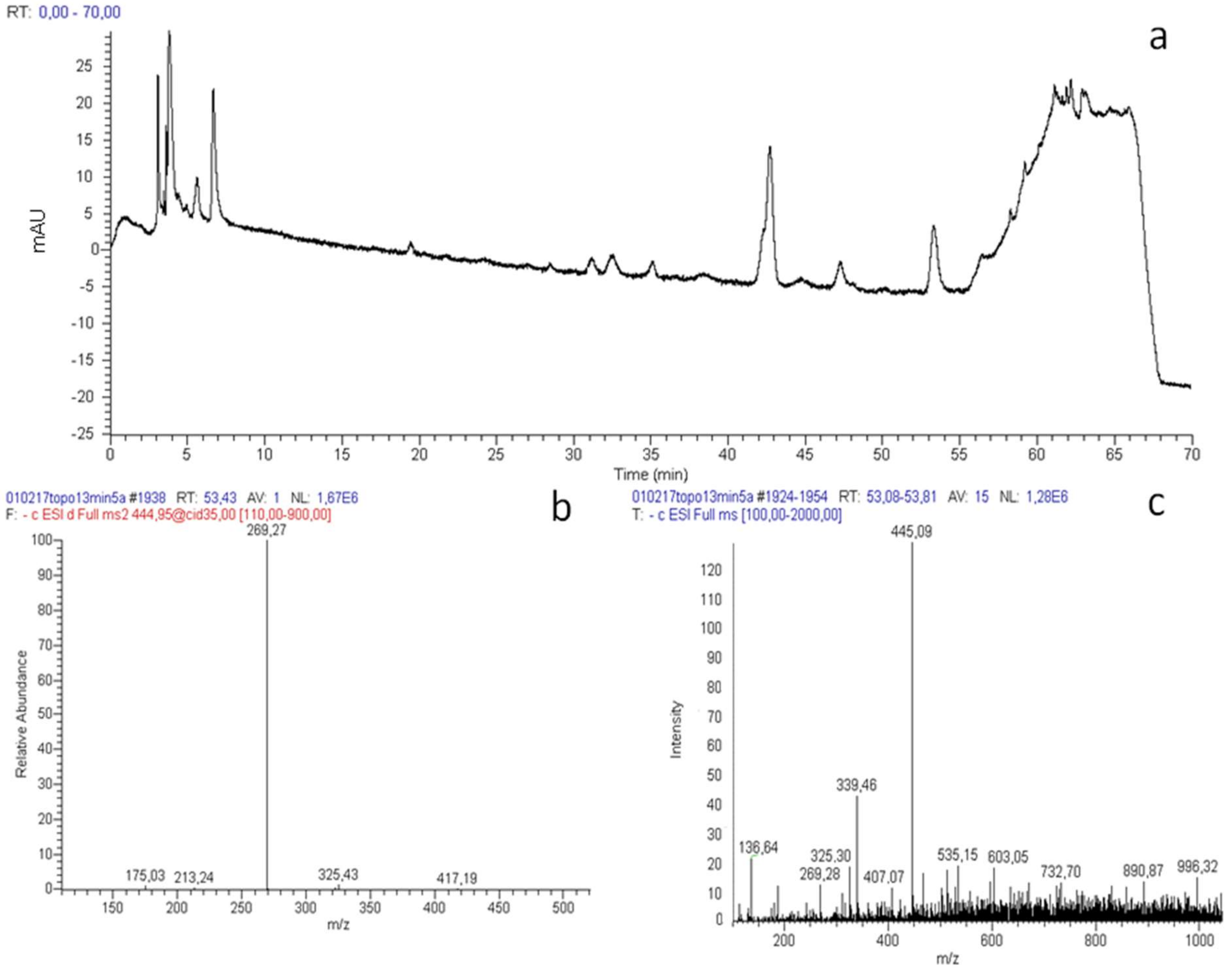
2.1. Analysis of Propolis Polyphenolic Content
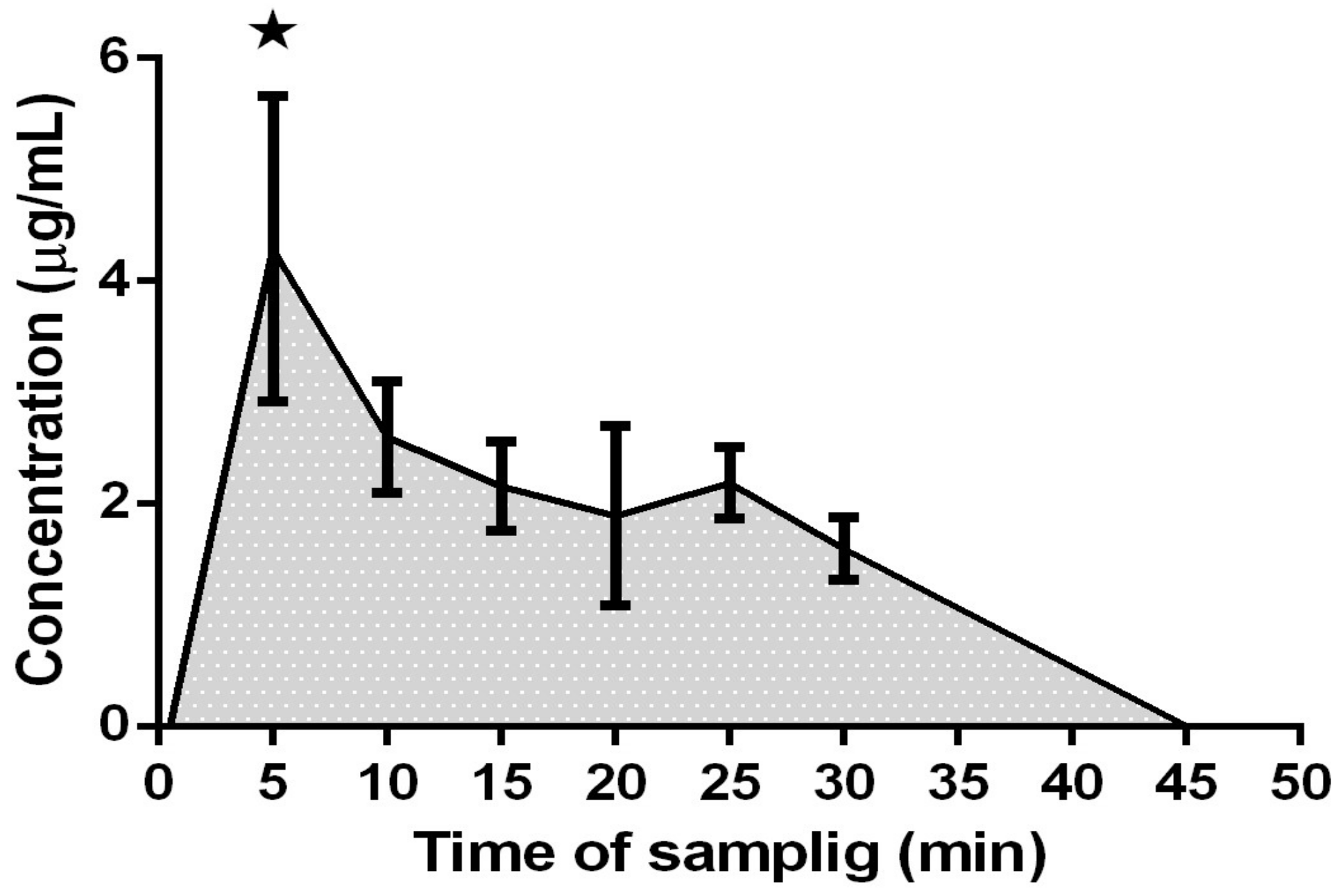
2.1.1. Acute Treatment
2.1.2. Prolonged Treatment
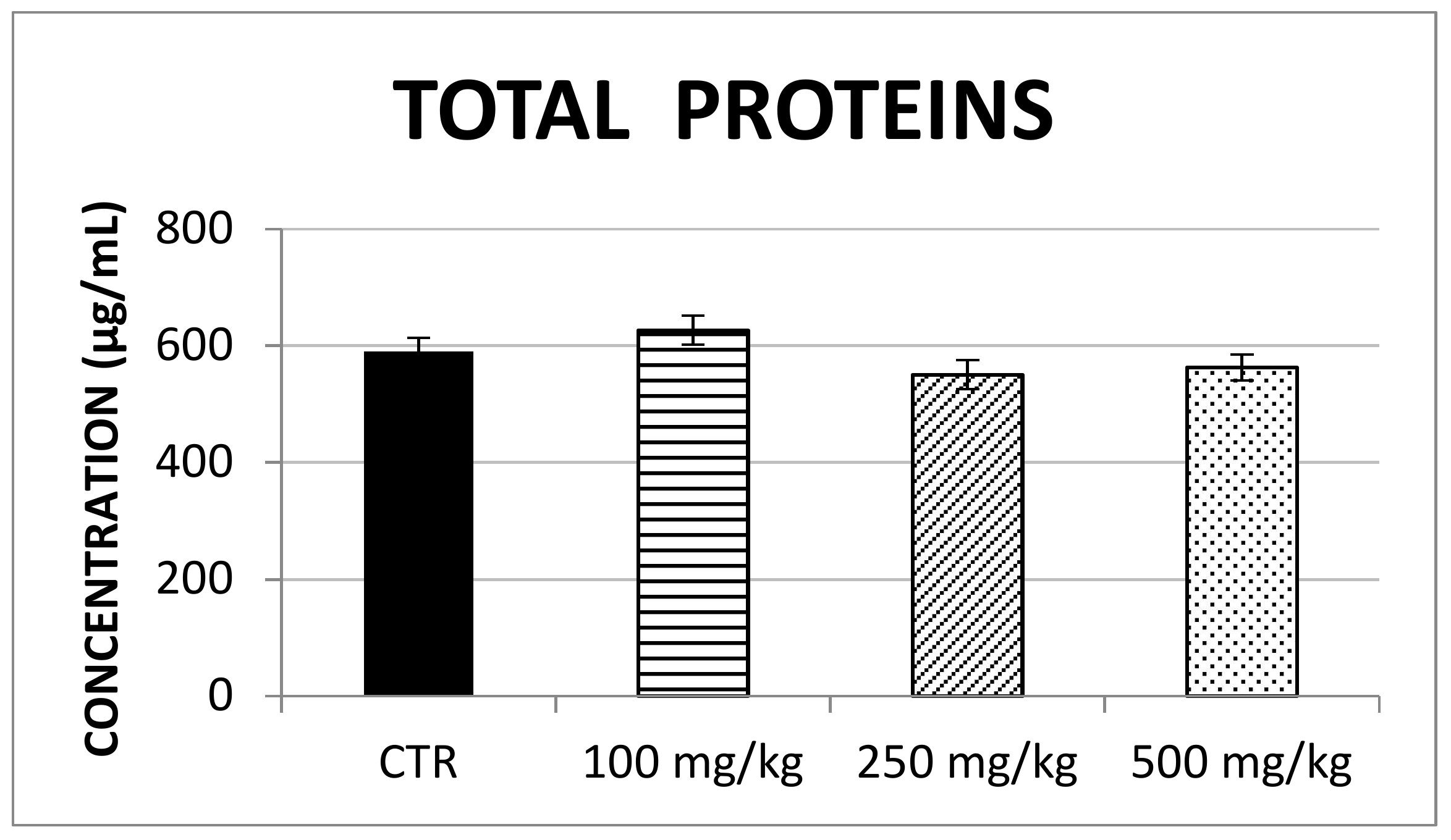
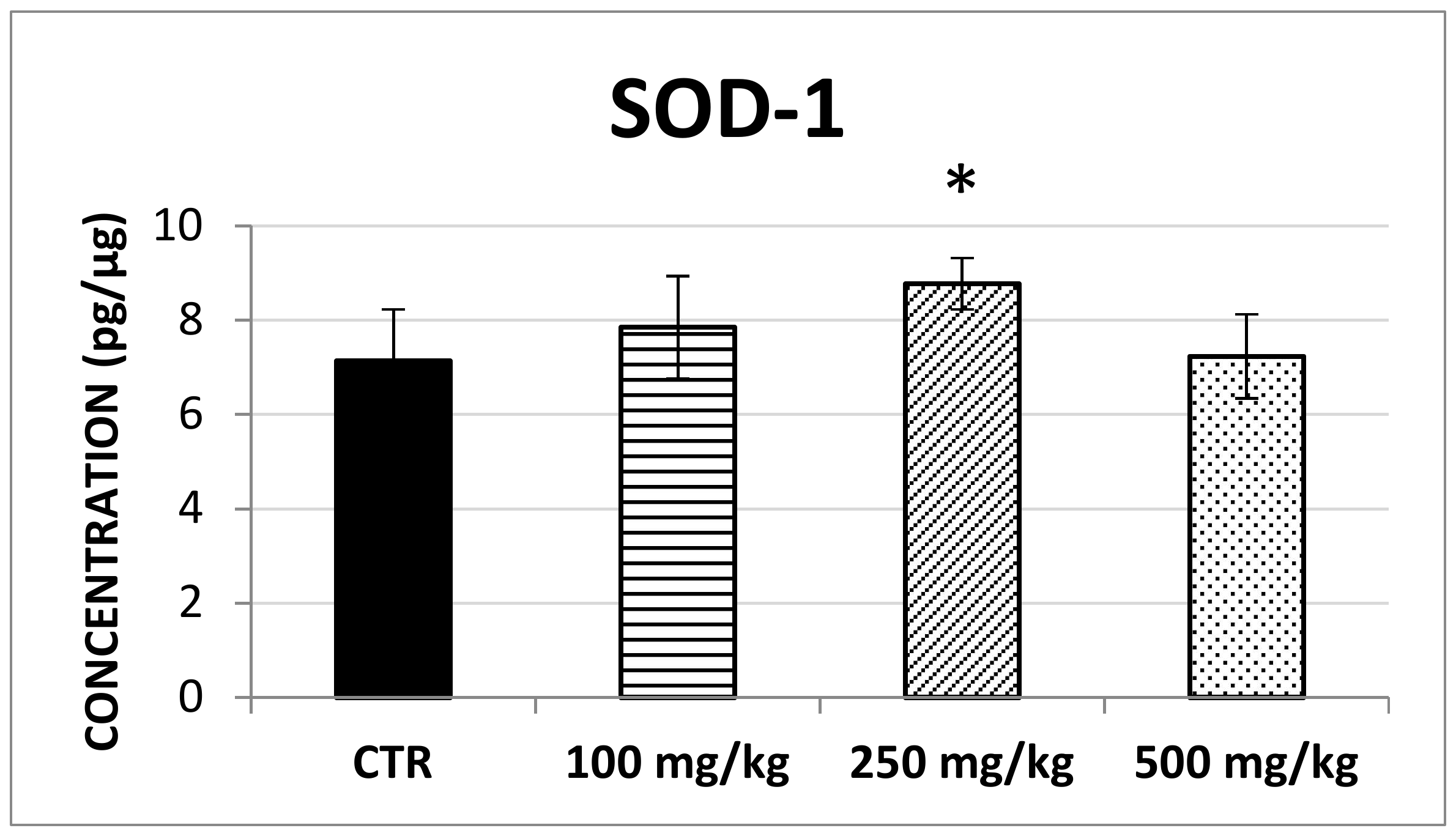
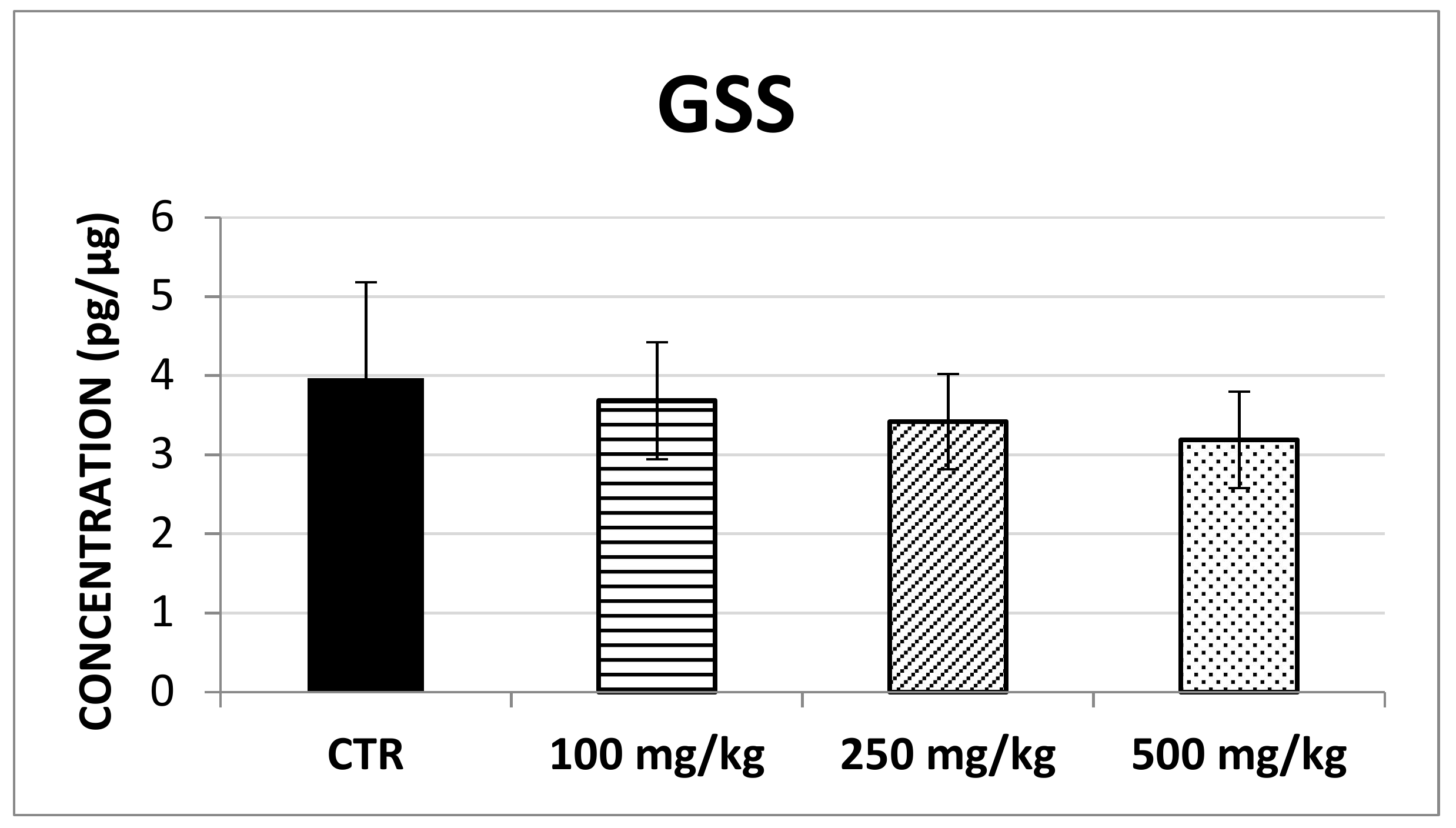
2.2. In Vivo Evaluation of Antioxidant Enzymes
3. Discussion
3.1. Bioavailability
3.2. In Vivo Antioxidant Effect
4. Materials and Methods
4.1. Materials
4.2. Determination of the Main Polyphenols in Brown Propolis M.E.D. Extract
4.3. Animal Model and Diet
4.4. Analysis of Propolis Bioavailability
4.4.1. Plasma Sample Preparation
4.4.2. Extraction of Galangin and Its Metabolites from Liver
4.4.3. RP-HPLC-PDA-ESI-MSn
4.5. Antioxidant Enzyme Quantification
4.5.1. Liver Sample Preparation
4.5.2. Bicinchoninic Acid Assay
4.5.3. ELISA Test
4.6. Statistical Analysis
5. Limitations and Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Park, Y.K.; Paredes-Guzman, J.F.; Aguiar, C.L.; Alencar, S.M.; Fujiwara, F.Y. Chemical constituents in Baccharis dracunculifolia as the main botanical origin of southeastern Brazilian propolis. J. Agric. Food Chem. 2004, 52, 1100–1103. [Google Scholar] [CrossRef] [PubMed]
- Daugsch, A.; Moraes, C.S.; Fort, P.; Park, Y.K. Brazilian red propolis—Chemical composition and botanical origin. Evid. Based Complement. Altern. Med. 2008, 5, 435–441. [Google Scholar] [CrossRef] [PubMed]
- Castaldo, S.; Capasso, F. Propolis, an old remedy used in modern medicine. Fitoterapia 2002, 73, 1–6. [Google Scholar] [CrossRef]
- Alencar, S.M.; Oldoni, T.L.C.; Castro, M.L.; Cabral, I.S.R.; Costa-Neto, C.M.; Cury, J.A.; Rosalen, P.L.; Ikegaki, M. Chemical composition and biological activity of a new type of Brazilian propolis: Red propolis. J. Ethnopharmacol. 2007, 113, 278–283. [Google Scholar] [CrossRef] [PubMed]
- Marcucci, M.C. Propolis: Chemical composition, biological properties and therapeutic activity. Apidologie 1995, 26, 83–99. [Google Scholar] [CrossRef]
- De Castro, S.L. Propolis: Biological and pharmacological activities. Therapeutic uses of this bee-product. Annu. Rev. Biomed. Sci. 2001, 3, 49–83. [Google Scholar] [CrossRef]
- Juliano, C.; Pala, C.L.; Cossu, M. Preparation and characterisation of polymeric films containing propolis. J. Drug Deliv. Sci. Technol. 2007, 17, 177–181. [Google Scholar] [CrossRef]
- Wang, T.; Chen, L.; Wu, W.; Long, Y.; Wang, R. Potential cytoprotection: Antioxidant defence by caffeic acid phenethyl ester against free radical-induced damage of lipids, DNA, and proteins. Can. J. Physiol. Pharmacol. 2008, 86, 279–287. [Google Scholar] [CrossRef]
- Casaroto, A.R.; Lara, V.S. Phytomedicines for Candida-associated denture stomatitis. Fitoterapia 2010, 81, 323–328. [Google Scholar] [CrossRef]
- Kurek-Górecka, A.; Rzepecka-Stojko, A.; Górecki, M.; Stojko, J.; Sosada, M.; Świerczek-Zięba, G. Structure and antioxidant activity of polyphenols derived from propolis. Molecules 2013, 19, 78–101. [Google Scholar] [CrossRef]
- Chee, H.Y. In vitro evaluation of the antifungal activity of propolis extract on Cryptococcus neoformans and Candida albicans. Mycobiology 2002, 30, 93. [Google Scholar] [CrossRef]
- Banskota, A.H.; Tezuka, Y.; Adnyana, I.K.; Midorikawa, K.; Matsushige, K.; Message, D.; Huertas, A.A.; Kadota, S. Cytotoxic, hepatoprotective and free radical scavenging effects of propolis from Brazil, Peru, The Netherlands and China. J. Ethnopharmacol. 2000, 72, 239–246. [Google Scholar] [CrossRef]
- Zaccaria, V.; Curti, V.; Di Lorenzo, A.; Baldi, A.; Maccario, C.; Sommatis, S.; Mocchi, R.; Daglia, M. Effect of green and brown propolis extracts on the expression levels of microRNAs, mRNAs and proteins, related to oxidative stress and inflammation. Nutrients 2017, 10, 1090. [Google Scholar] [CrossRef] [PubMed]
- Porrini, M.; Riso, P. Factors influencing the bioavailability of antioxidants in foods: A critical appraisal. Nutr. Metab. Cardiovasc. Dis. 2008, 18, 647–650. [Google Scholar] [CrossRef] [PubMed]
- D’Archivio, M.; Filesi, C.; Varì, R.; Scazzocchio, B.; Masella, R. Bioavailability of the polyphenols: Status and controversies. Int. J. Mol. Sci. 2010, 4, 1321–1342. [Google Scholar] [CrossRef]
- Hausen, B.M.; Wollenweber, E.; Senff, H.; Post, B. Propolis allergy: (I). Origin, properties, usage and literature review. Contact Dermat. 1987, 17, 163–177. [Google Scholar] [CrossRef]
- Gardana, C.; Simonetti, P.; Berti, C.; Pietta, P. Evaluation of propolis polyphenols absorption in humans by liquid chromatography/tandem mass spectrometry. Rapid. Commun. Mass Spectrom. 2007, 21, 3849–3854. [Google Scholar] [CrossRef] [PubMed]
- Nabavi, S.F.; Braidy, N.; Habtemariam, S.; Orhan, I.E.; Daglia, M.; Manayi, A.; Gortzi, O.; Nabavi, S.M. Neuroprotective effects of chrysin: From chemistry to medicine. Neurochem. Int. 2015, 90, 224–231. [Google Scholar] [CrossRef] [PubMed]
- Konishi, Y.; Hitomi, Y.; Yoshida, M.; Yoshioka, E. Absorption and bioavailability of artepillin c in rats after oral administration. J. Agric. Food Chem. 2015, 53, 9928–9933. [Google Scholar] [CrossRef]
- Konishi, Y.; Hitomi, Y.; Yoshida, M.; Yoshioka, E. Pharmacokinetic study of caffeic and rosmarinic acids in rats after oral administration. J. Agric. Food Chem. 2015, 53, 4740–4746. [Google Scholar] [CrossRef]
- Konishi, Y.; Kobayashi, S. Microbial metabolites of ingested caffeic acid are absorbed by the monocarboxylic acid transporter (MCT) in intestinal Caco-2 cell monolayers. J. Agric. Food Chem. 2004, 52, 6418–6424. [Google Scholar] [CrossRef] [PubMed]
- Konishi, Y.; Hitomi, Y.; Yoshioka, E. Intestinal absorption of p -coumaric and gallic acids in rats after oral administration. J. Agric. Food Chem. 2004, 52, 2527–2532. [Google Scholar] [CrossRef]
- Zhao, Z.; Egashira, Y.; Sanada, H. Ferulic acid is quickly absorbed from rat stomach as the free form and then conjugated mainly in liver. J. Nutr. 2004, 134, 3083–3088. [Google Scholar] [CrossRef] [PubMed]
- Lahouel, M.; Boulkour, S.; Segueni, N.; Fillastre, J.P. The flavonoids effect against vinblastine, cyclophosphamide and paracetamol toxicity by inhibition of lipid-peroxydation and increasing liver glutathione concentration. Pathol. Biol. (Paris) 2004, 52, 314–322. [Google Scholar] [CrossRef] [PubMed]
- De Barros, M.P.; Lemos, M.; Maistro, E.L.; Leite, M.F.; Sousa, J.P.B.; Bastos, J.K.; de Andrade, S.F. Evaluation of antiulcer activity of the main phenolic acids found in Brazilian green propolis. J. Ethnopharmacol. 2008, 120, 372–377. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.F.; Lin, C.C.; Lin, M.H.; Lin, Y.S.; Lin, S.C. Cytoprotection by propolis ethanol extract of acute absolute ethanol-induced gastric mucosal lesions. Am. J. Chin. Med. 2002, 30, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Jasprica, I.; Mornar, A.; Debeljak, Ž.; Smolčić-Bubalo, A.; Medić-Šarić, M.; Mayer, L.; Romić, Ž.; Bućan, K.; Balog, T.; Sobočanec, S.; Šverko, V. In vivo study of propolis supplementation effects on antioxidative status and red blood cells. J. Ethnopharmacol. 2007, 110, 548–554. [Google Scholar] [CrossRef]
- Benz, C.C.; Yau, C. Ageing, oxidative stress and cancer: Paradigms in parallax. Nat. Rev. Cancer 2008, 8, 875–879. [Google Scholar] [CrossRef]
- Favier, A. Oxidative stress: Value of its demonstration in medical biology and problems posed by the choice of a marker. Ann. Biol. Clin. (Paris) 1997, 55, 9–16. [Google Scholar]
- Zamora-Ros, R.; Knaze, V.; Rothwell, J.A.; Hémon, B.; Moskal, A.; Overvad, K.; Tjønneland, A.; Kyrø, C.; Fagherazzi, G.; Boutron-Ruault, M.C.; et al. Dietary polyphenol intake in europe: The european prospective investigation into cancer and nutrition (EPIC) study. Eur. J. Nutr. 2016, 55, 1359–1375. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Lu, Y.; Chen, Y.; Cheng, J. The role of Nrf2 in oxidative stress-induced endothelial injuries. J. Endocrinol. 2015, 225, 83–99. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Pu, L.; Wei, J.; Li, J.; Wu, J.; Xin, Z.; Gao, W.; Guo, C. Brazilian green propolis improves antioxidant function in patients with type 2 diabetes mellitus. Int. J. Environ. Res. Public Health 2016, 13, 498. [Google Scholar] [CrossRef] [PubMed]
- Williams, R.J.; Spencer, J.P.; Rice-Evans, C. Flavonoids: Antioxidants or signalling molecules? Free Radic. Biol. Med. 2004, 36, 838–849. [Google Scholar] [CrossRef]
- Zelko, I.N.; Mariani, T.J.; Folz, R.J. Superoxide dismutase multigene family: A comparison of the CuZn-SOD (SOD1), Mn-SOD (SOD2), and EC-SOD (SOD3) gene structures, evolution, and expression. Free Radic. Biol. Med. 2002, 33, 337–349. [Google Scholar] [CrossRef]
- Cui-ping, Z.; Shuai, H.; Wen-ting, W.; Shun, P.; Xiao-ge, S.; Ya-jing, L.; Fu-liang, H. Development of high-performance liquid chromatographic for quality and authenticity control of Chinese propolis. J. Food Sci. 2014, 79, 1315–1322. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Fish, B.L.; Moulder, J.E.; Medhora, M.; Baker, J.E.; Mader, M.; Cohen, E.P. Safety and blood sample volume and quality of a refined retro-orbital bleeding technique in rats using a lateral approach. Lab Anim. 2014, 43, 63–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Polyphenols | % Area |
|---|---|
| Quercetin | 1.3 |
| Apigenin | 1.8 |
| Pinobanksin | 1.6 |
| Chrysin | 7.5 |
| Pinocembrin | 5.0 |
| Galangin | 7.8 |
| Mouse | Time 1 | Time 2 | Mouse | Time 1 | Time 2 |
|---|---|---|---|---|---|
| 1 | 5′ | 20′ | |||
| 2 | 10′ | 25′ | 16 | 10′ | 25′ |
| 3 | 15′ | 30′ | 17 | 15′ | 30′ |
| 4 | 5′ | 20′ | 18 | 30″ | 45′ |
| 5 | 10′ | 25′ | 19 | 60′ | 120′ |
| 6 | 15′ | 30′ | 20 | 30″ | 45′ |
| 7 | 5′ | 20′ | 21 | 60′ | 120′ |
| 8 | 10′ | 25′ | 22 | 30″ | 45′ |
| 9 | 15′ | 30′ | 23 | 60′ | 120′ |
| 10 | 5′ | 20′ | 24 | 30″ | 45′ |
| 11 | 10′ | 25′ | 25 | 60′ | 120′ |
| 12 | 15′ | 30′ | 26 | 30″ | 45′ |
| 13 | 5′ | 20′ | 27 | 60′ | 120′ |
| 14 | 10′ | 25′ | 28 | 30″ | 45′ |
| 15 | 15′ | 25′ | 29 | 60′ | 45′ |
| Time (min) | % Eluent A | % Eluent B |
|---|---|---|
| 0 | 70 | 30 |
| 10 | 60 | 40 |
| 30 | 45 | 55 |
| 50 | 30 | 70 |
| 55 | 0 | 100 |
| 60 | 0 | 100 |
| 65 | 70 | 30 |
| 70 | 70 | 30 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Curti, V.; Zaccaria, V.; Tsetegho Sokeng, A.J.; Dacrema, M.; Masiello, I.; Mascaro, A.; D’Antona, G.; Daglia, M. Bioavailability and In Vivo Antioxidant Activity of a Standardized Polyphenol Mixture Extracted from Brown Propolis. Int. J. Mol. Sci. 2019, 20, 1250. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20051250
Curti V, Zaccaria V, Tsetegho Sokeng AJ, Dacrema M, Masiello I, Mascaro A, D’Antona G, Daglia M. Bioavailability and In Vivo Antioxidant Activity of a Standardized Polyphenol Mixture Extracted from Brown Propolis. International Journal of Molecular Sciences. 2019; 20(5):1250. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20051250
Chicago/Turabian StyleCurti, Valeria, Vincenzo Zaccaria, Arold Jorel Tsetegho Sokeng, Marco Dacrema, Irene Masiello, Anna Mascaro, Giuseppe D’Antona, and Maria Daglia. 2019. "Bioavailability and In Vivo Antioxidant Activity of a Standardized Polyphenol Mixture Extracted from Brown Propolis" International Journal of Molecular Sciences 20, no. 5: 1250. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20051250