Effects of Cold-inducible RNA-binding Protein (CIRP) on Liver Glycolysis during Acute Cold Exposure in C57BL/6 Mice

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Acute Cold Exposure Induced Changes in Core Body Temperature
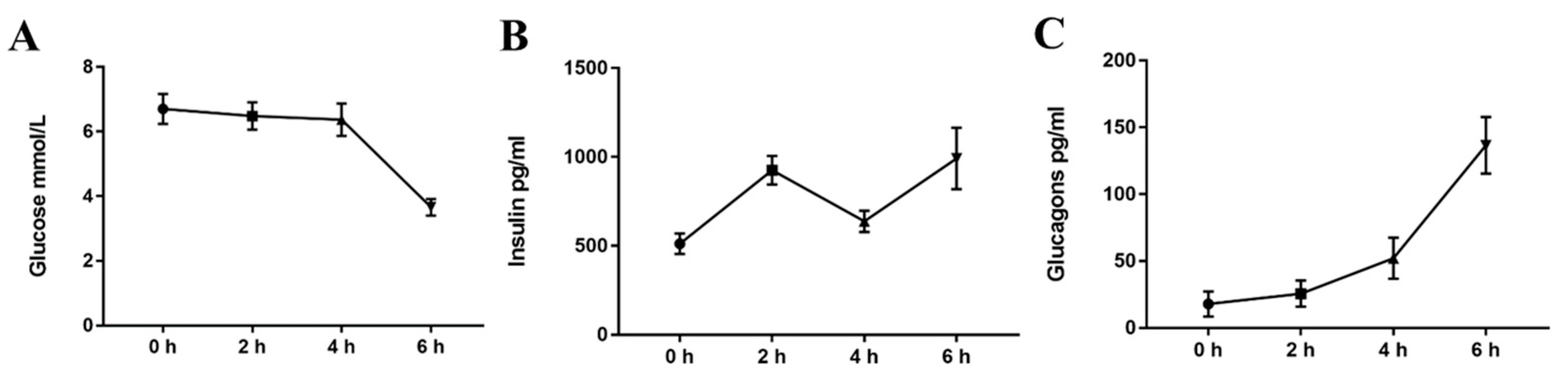
2.2. Acute Cold Exposure Induces A Plasma Glucose, Insulin, and Glucagon Time-Course
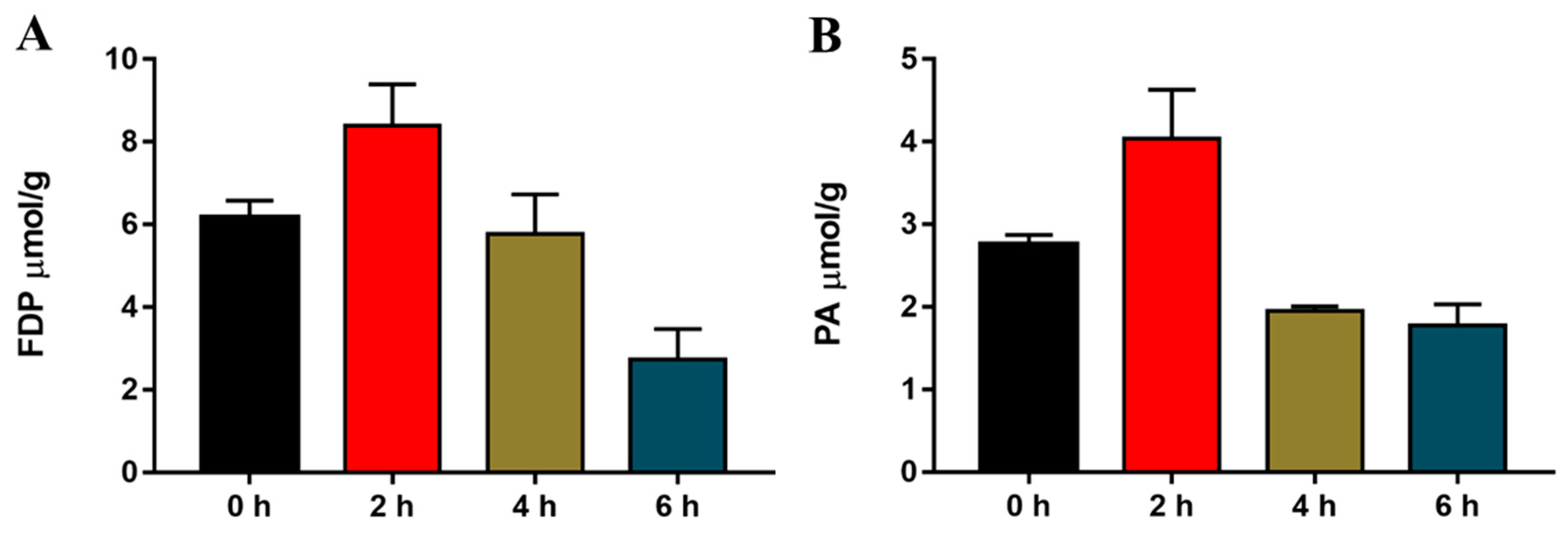
2.3. Acute Cold Exposure-Induced Changes in Hepatic Fructose-1,6-diphosphate (FDP) and Pyruvic Acid (PA) in Mice
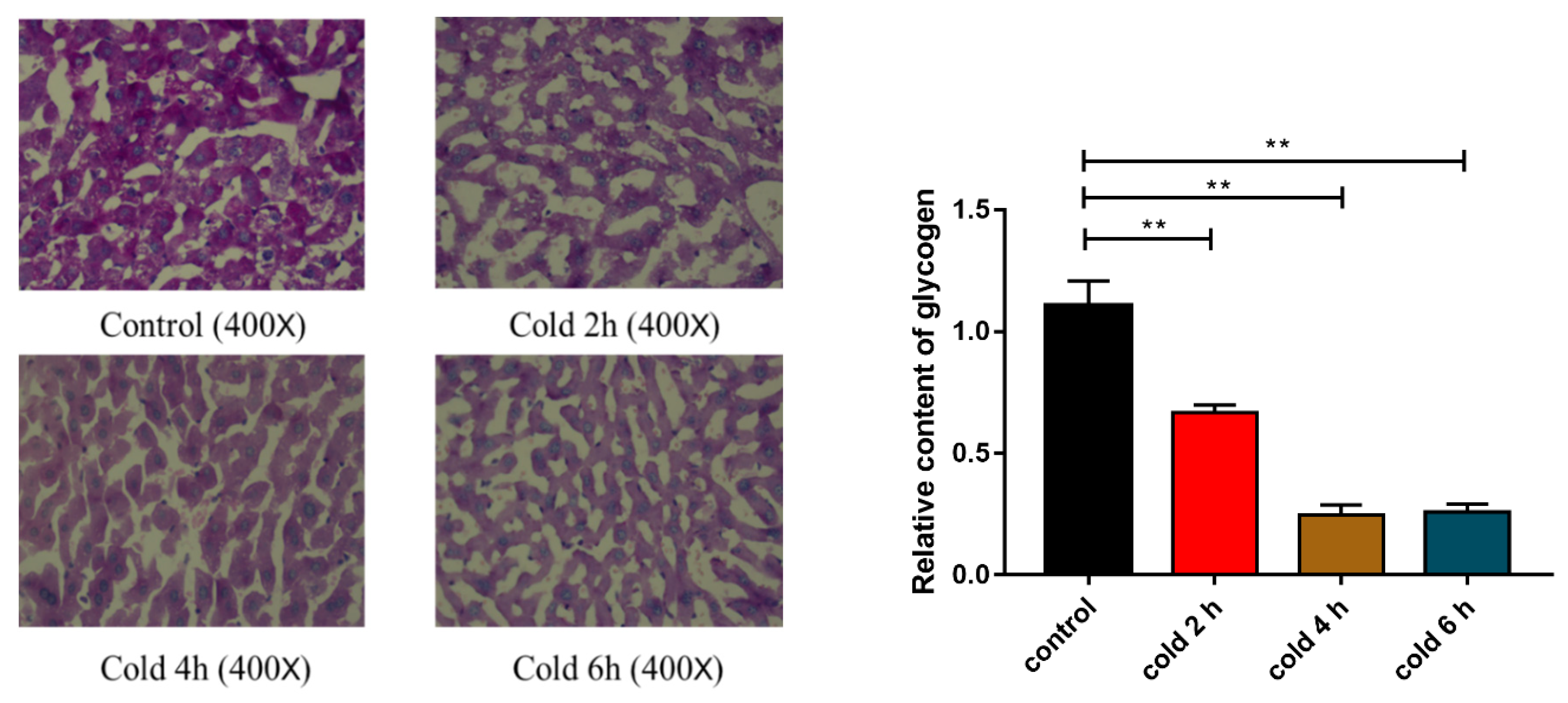
2.4. Acute Cold Exposure Induces Glycogen Depletion
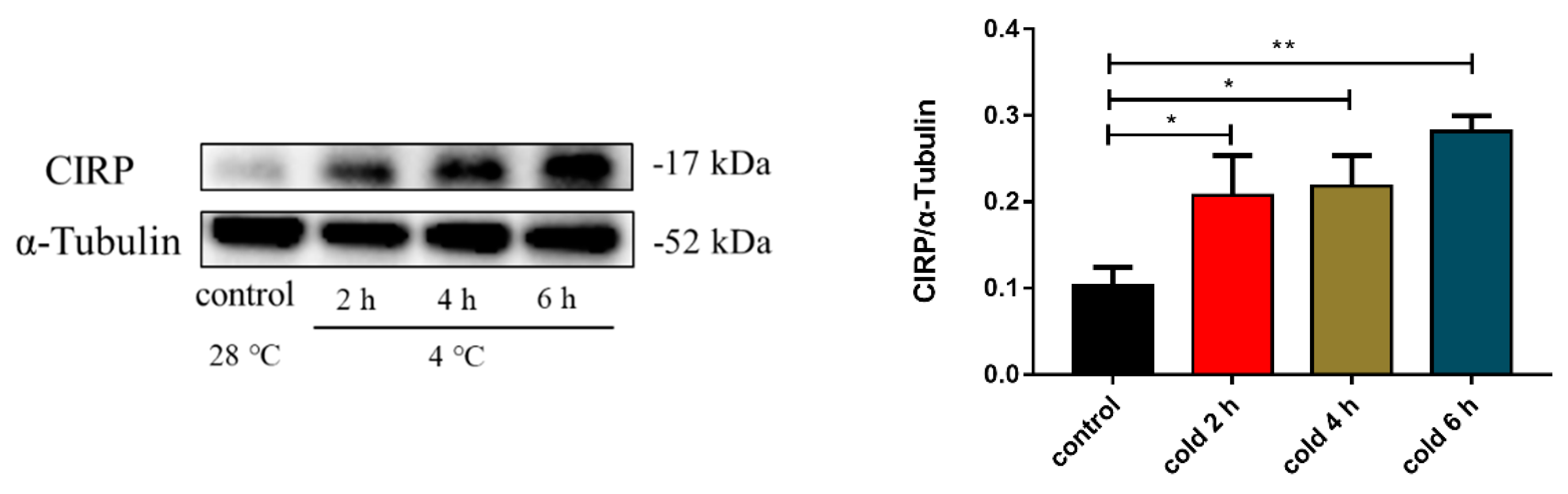
2.5. Acute Cold Exposure Upregulates CIRP Expression
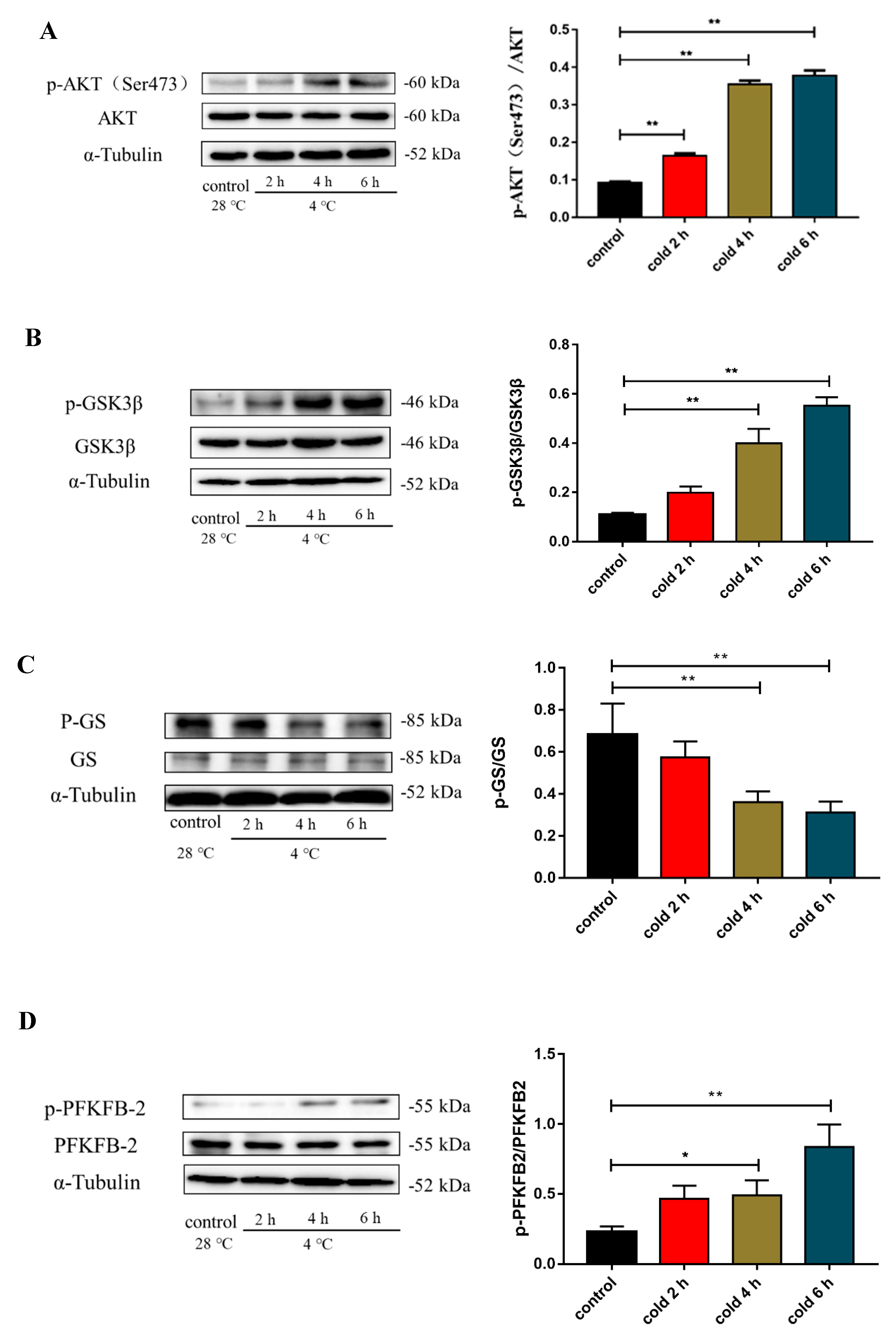
2.6. Acute Cold Exposure Activates the AKT Signaling Pathway
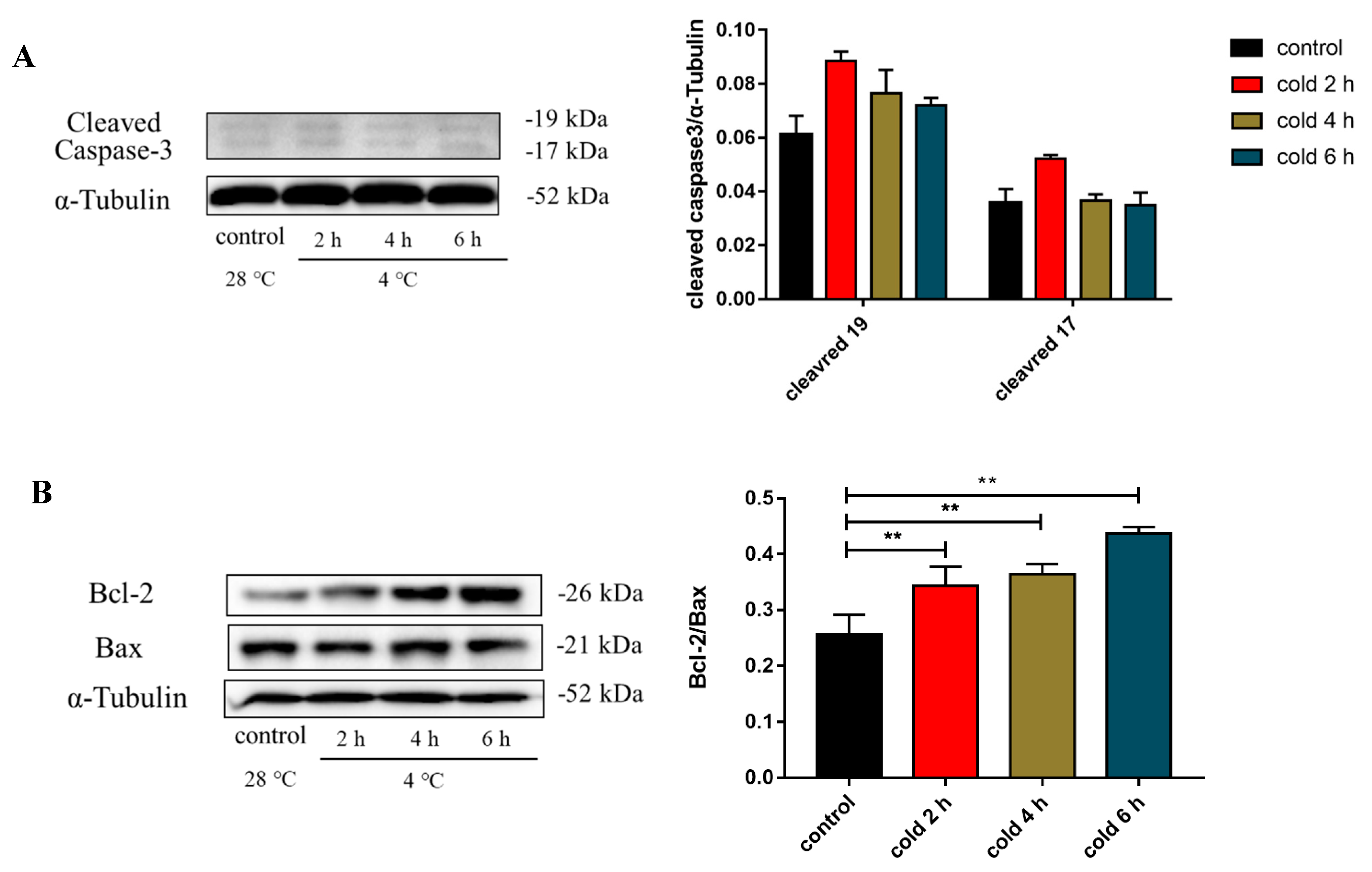
2.7. Acute Cold Exposure Induces Apoptosis
2.8. Effect of AKT on Glucose Metabolism in C57BL/6 Mouse Liver under Cold Exposure
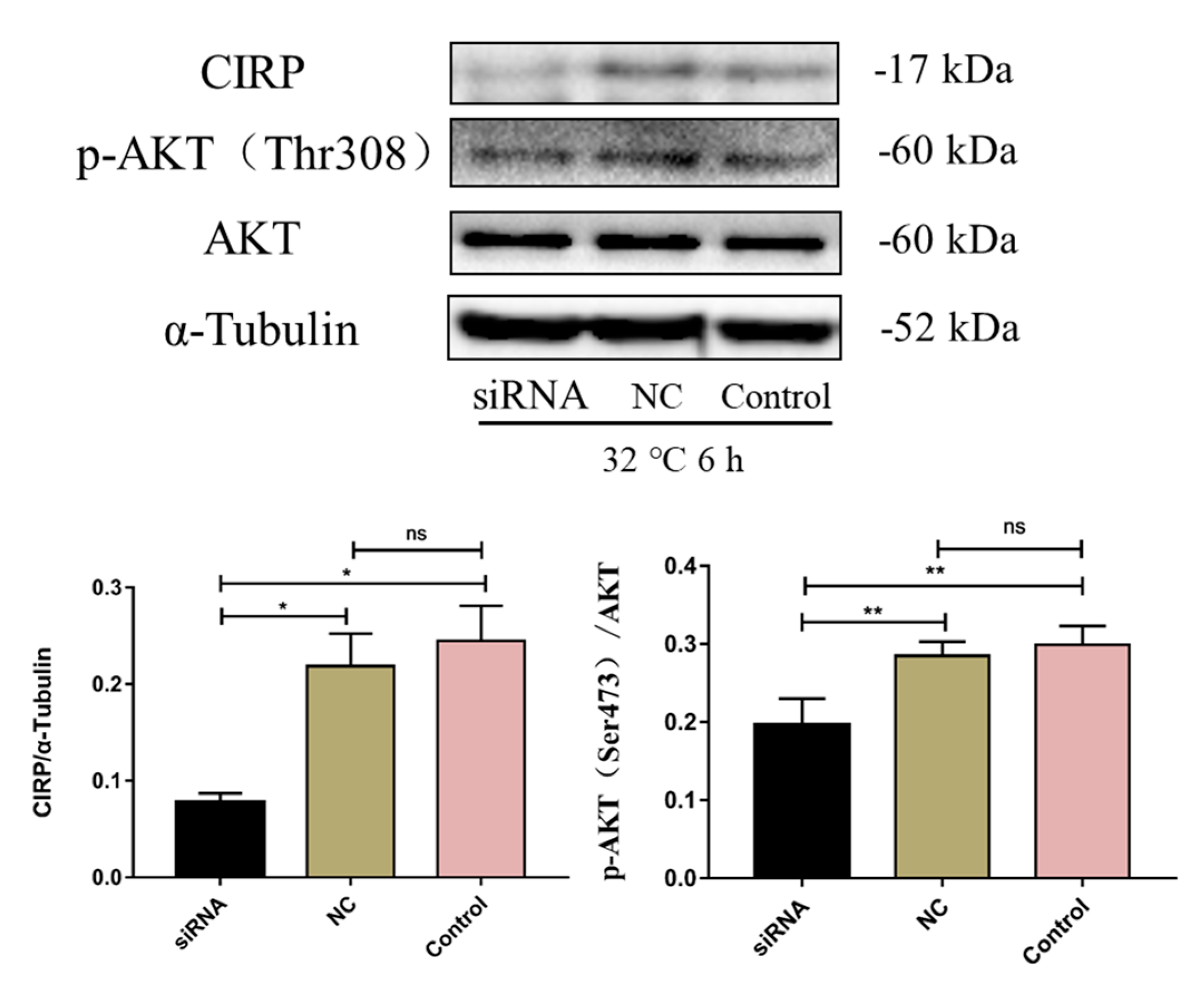
2.9. CIRP Silencing Inhibits AKT Activity
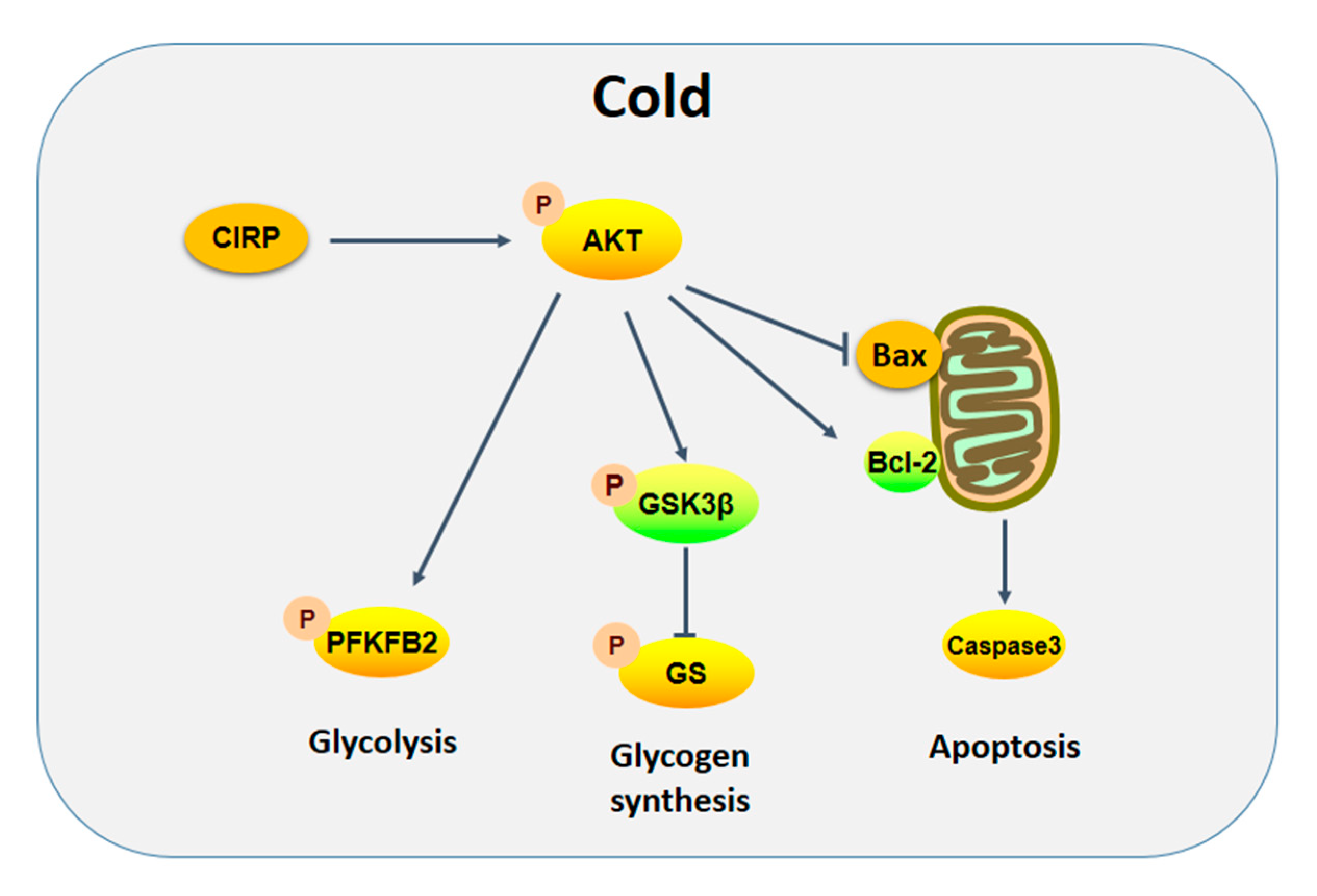
3. Discussion
4. Materials and Methods
4.1. Animals
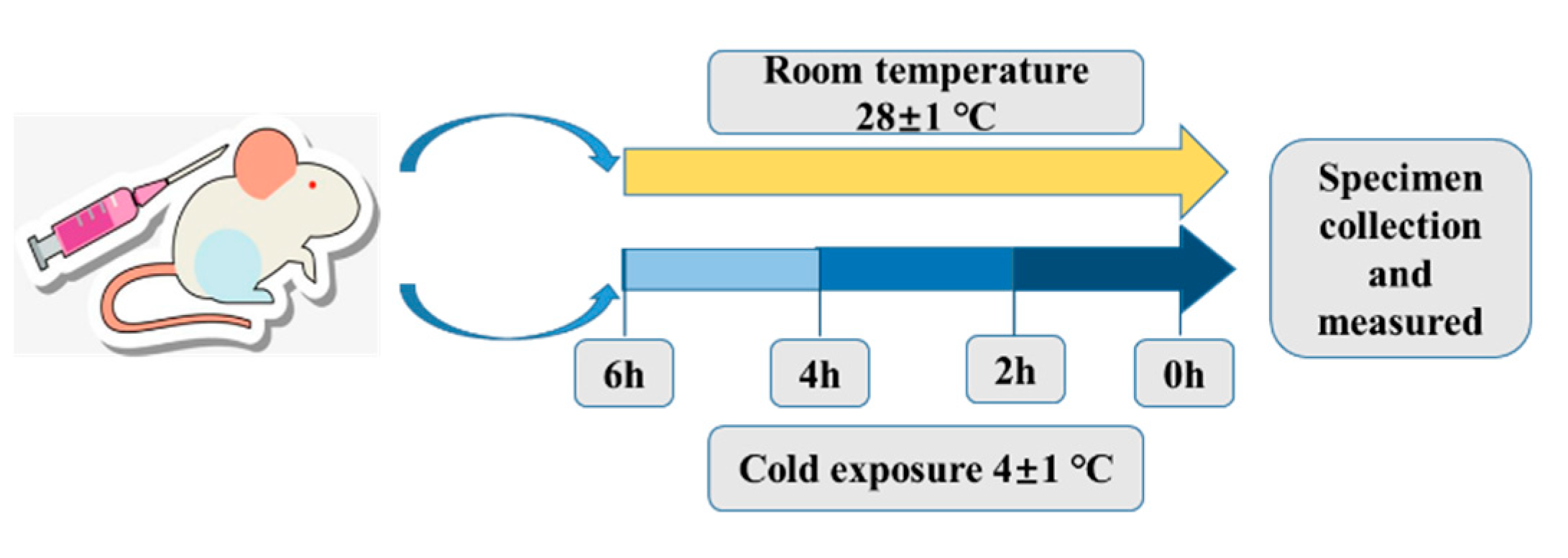
4.2. Cold Exposure Experiment, Core Body Temperature Measurement and Sample Collection
4.3. Cell Culture and Treatment
4.4. siRNA Transfections
4.5. Measurement of Biochemical Parameters
4.6. PAS Staining
4.7. Western Blot Analysis
4.8. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Bozkurt, A.; Ghandour, S.; Okboy, N.; Oner, S.; Arbak, S.; Coskun, T.; Yeğen, BC. Inflammatory response to cold injury in remote organs is reduced by corticotropin-releasing factor. Regul. Pept. 2001, 99, 131–139. [Google Scholar] [CrossRef]
- Venditti, P.; Pamplona, R.; Ayala, V.; De Rosa, R.; Caldarone, G.; Di Meo, S. Differential effects of experimental and cold-induced hyperthyroidism on factors inducing rat liver oxidative damage. J. Exp. Biol. 2006, 209, 817–825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lleonart, M.E. A new generation of proto-oncogenes: Cold-inducible RNA binding proteins. Biochim. Et Biophys. Acta 2010, 1805, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Sepa-Kishi, D.M.; Sotoudeh-Nia, Y.; Iqbal, A.; Bikopoulos, G.; Ceddia, R.B. Cold acclimation causes fiber type-specific responses in glucose and fat metabolism in rat skeletal muscles. Sci. Rep. 2017, 7, 15430. [Google Scholar] [CrossRef]
- Wang, J.; Chen, Y.; Zhang, W.; Zheng, G.; Meng, S.; Che, H.; Ke, T.; Yang, J.; Chen, J.; Luo, W. Akt activation protects liver cells from apoptosis in rats during acute cold exposure. Int. J. Biol. Sci. 2013, 9, 509–517. [Google Scholar] [CrossRef] [PubMed]
- Meneghini, A.; Ferreira, C.; Abreu, L.C.; Ferreira, M.; Ferreira Filho, C.; Valenti, V.E.; Murad, N. Cold stress effects on cardiomyocytes nuclear size in rats: Light microscopic evaluation. Braz. J. Cardiovasc. Surg. 2008, 23, 530–533. [Google Scholar] [CrossRef]
- Sheikh, M.S.; Carrier, F.; Papathanasiou, M.A.; Hollander, M.C.; Zhan, Q.; Yu, K.; Fornace, A.J., Jr. Identification of several human homologs of hamster DNA damage-inducible transcripts. Cloning and characterization of a novel UV-inducible cDNA that codes for a putative RNA-binding protein. J. Biol. Chem. 1997, 272, 26720–26726. [Google Scholar] [CrossRef] [PubMed]
- Nishiyama, H.; Itoh, K.; Kaneko, Y.; Kishishita, M.; Yoshida, O.; Fujita, J. A glycine-rich RNA-binding protein mediating cold-inducible suppression of mammalian cell growth. J. Cell Biol. 1997, 137, 899–908. [Google Scholar] [CrossRef]
- Saito, T.; Sugimoto, K.; Adachi, Y.; Wu, Q.; Mori, K.J. Cloning and characterization of amphibian cold inducible RNA-binding protein. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2000, 125, 237–245. [Google Scholar] [CrossRef]
- Sugimoto, K.; Jiang, H. Cold stress and light signals induce the expression of cold-inducible RNA binding protein (cirp) in the brain and eye of the Japanese treefrog (Hyla japonica). Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2008, 151, 628–636. [Google Scholar] [CrossRef]
- Xia, Z.; Zheng, X.; Zheng, H.; Liu, X.; Yang, Z.; Wang, X. Cold-inducible RNA-binding protein (CIRP) regulates target mRNA stabilization in the mouse testis. FEBS Lett. 2012, 586, 3299–3308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, Y.; Yang, P.H.; Tanner, J.A.; Huang, J.D.; Li, M.; Lee, H.F.; Xu, R.H.; Kung, H.F.; Lin, M.C. Cold-inducible RNA binding protein is required for the expression of adhesion molecules and embryonic cell movement in Xenopus laevis. Biochem. Biophys. Res. Commun. 2006, 344, 416–424. [Google Scholar] [CrossRef] [PubMed]
- van Venrooy, S.; Fichtner, D.; Kunz, M.; Wedlich, D.; Gradl, D. Cold-inducible RNA binding protein (CIRP), a novel XTcf-3 specific target gene regulates neural development in Xenopus. BMC Dev. Biol. 2008, 8, 77. [Google Scholar] [CrossRef] [PubMed]
- Xia, Z.; Jiang, K.; Liu, T.; Zheng, H.; Liu, X.; Zheng, X. The protective effect of Cold-inducible RNA-binding protein (CIRP) on testicular torsion/detorsion: An experimental study in mice. J. Pediatric Surg. 2013, 48, 2140–2147. [Google Scholar] [CrossRef]
- Xia, Z.P.; Zheng, X.M.; Zheng, H.; Liu, X.J.; Liu, G.Y.; Wang, X.H. Downregulation of cold-inducible RNA-binding protein activates mitogen-activated protein kinases and impairs spermatogenic function in mouse testes. Asian J. Androl. 2012, 14, 884–889. [Google Scholar] [CrossRef] [Green Version]
- Jo, J.W.; Lee, J.R.; Jee, B.C.; Suh, C.S.; Kim, S.H. Exposing mouse oocytes to necrostatin 1 during in vitro maturation improves maturation, survival after vitrification, mitochondrial preservation, and developmental competence. Reprod. Sci. 2015, 22, 615–625. [Google Scholar] [CrossRef] [PubMed]
- Gerber, A.; Saini, C.; Curie, T.; Emmenegger, Y.; Rando, G.; Gosselin, P.; Gotic, I.; Gos, P.; Franken, P.; Schibler, U. The systemic control of circadian gene expression. Diabetesobesity Metab. 2015, 17 (Suppl. 1), 23–32. [Google Scholar] [CrossRef] [Green Version]
- Bellesi, M.; de Vivo, L.; Tononi, G.; Cirelli, C. Effects of sleep and wake on astrocytes: Clues from molecular and ultrastructural studies. BMC Biol. 2015, 13, 66. [Google Scholar] [CrossRef]
- Jian, F.; Chen, Y.; Ning, G.; Fu, W.; Tang, H.; Chen, X.; Zhao, Y.; Zheng, L.; Pan, S.; Wang, W.; et al. Cold inducible RNA binding protein upregulation in pituitary corticotroph adenoma induces corticotroph cell proliferation via Erk signaling pathway. Oncotarget 2016, 7, 9175–9187. [Google Scholar] [CrossRef] [Green Version]
- Sakurai, T.; Kashida, H.; Watanabe, T.; Hagiwara, S.; Mizushima, T.; Iijima, H.; Nishida, N.; Higashitsuji, H.; Fujita, J.; Kudo, M. Stress response protein cirp links inflammation and tumorigenesis in colitis-associated cancer. Cancer Res. 2014, 74, 6119–6128. [Google Scholar] [CrossRef]
- Sakurai, T.; Yada, N.; Watanabe, T.; Arizumi, T.; Hagiwara, S.; Ueshima, K.; Nishida, N.; Fujita, J.; Kudo, M. Cold-inducible RNA-binding protein promotes the development of liver cancer. Cancer Sci. 2015, 106, 352–358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolognese, A.C.; Sharma, A.; Yang, W.L.; Nicastro, J.; Coppa, G.F.; Wang, P. Cold-inducible RNA-binding protein activates splenic T cells during sepsis in a TLR4-dependent manner. Cell. Mol. Immunol. 2018, 15, 38–47. [Google Scholar] [CrossRef]
- Qiang, X.; Yang, W.L.; Wu, R.; Zhou, M.; Jacob, A.; Dong, W.; Kuncewitch, M.; Ji, Y.; Yang, H.; Wang, H.; et al. Cold-inducible RNA-binding protein (CIRP) triggers inflammatory responses in hemorrhagic shock and sepsis. Nat. Med. 2013, 19, 1489–1495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ode, Y.; Aziz, M.; Wang, P. CIRP increases ICAM-1(+) phenotype of neutrophils exhibiting elevated iNOS and NETs in sepsis. J. Leukoc. Biol. 2018, 103, 693–707. [Google Scholar] [CrossRef]
- Saito, K.; Fukuda, N.; Matsumoto, T.; Iribe, Y.; Tsunemi, A.; Kazama, T.; Yoshida-Noro, C.; Hayashi, N. Moderate low temperature preserves the stemness of neural stem cells and suppresses apoptosis of the cells via activation of the cold-inducible RNA binding protein. Brain Res. 2010, 1358, 20–29. [Google Scholar] [CrossRef] [PubMed]
- Li, J.H.; Zhang, X.; Meng, Y.; Li, C.S.; Ji, H.; Yang, H.M.; Li, S.Z. Cold inducible RNA-binding protein inhibits hippocampal neuronal apoptosis under hypothermia by regulating redox system. Acta Physiol. Sin. 2015, 67, 386–392. [Google Scholar]
- Alkabie, S.; Boileau, A.J. The Role of Therapeutic Hypothermia After Traumatic Spinal Cord Injury—A Systematic Review. World Neurosurg. 2016, 86, 432–449. [Google Scholar] [CrossRef] [PubMed]
- Acosta, F.M.; Martinez-Tellez, B.; Sanchez-Delgado, G.; Alcantara, J.M.A.; Acosta-Manzano, P.; Morales-Artacho, A.J.; Ruiz, J.R. Physiological responses to acute cold exposure in young lean men. PLoS ONE. 2018, 13, e0196543. [Google Scholar]
- Sharabi, K.; Tavares, C.D.; Rines, A.K.; Puigserver, P. Molecular pathophysiology of hepatic glucose production. Mol. Asp. Med. 2015, 46, 21–33. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wahl, R. Responses of the insulin signaling pathways in the brown adipose tissue of rats following cold exposure. PLoS ONE 2014, 9, e99772. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Ishida, C.T.; Shu, C.; Kleiner, G.; Sanchez-Quintero, M.J.; Bianchetti, E.; Quinzii, C.M.; Westhoff, M.A.; Karpel-Massler, G.; Siegelin, M.D. Inhibition of Bcl-2/Bcl-xL and c-MET causes synthetic lethality in model systems of glioblastoma. Sci. Rep. 2018, 8, 7373. [Google Scholar] [CrossRef] [PubMed]
- Betz, M.J.; Enerback, S. Targeting thermogenesis in brown fat and muscle to treat obesity and metabolic disease. Nat. Rev. Endocrinol. 2018, 14, 77–87. [Google Scholar] [CrossRef]
- Qin, J.; Mai, Y.; Li, Y.; Jiang, Z.; Gao, Y. Effect of mild hypothermia preconditioning against low temperature (4 degrees C) induced rat liver cell injury in vitro. PLoS ONE 2017, 12, e0176652. [Google Scholar] [CrossRef] [PubMed]
- Richter, C.; Schweizer, M.; Cossarizza, A.; Franceschi, C. Control of apoptosis by the cellular ATP level. FEBS Lett. 1996, 378, 107–110. [Google Scholar] [CrossRef] [Green Version]
- Manning, B.D.; Cantley, L.C. AKT/PKB signaling: Navigating downstream. Cell 2007, 129, 1261–1274. [Google Scholar] [CrossRef]
- Han, C.Y.; Lim, S.W.; Koo, J.H.; Kim, W.; Kim, S.G. PHLDA3 overexpression in hepatocytes by endoplasmic reticulum stress via IRE1-Xbp1s pathway expedites liver injury. Gut 2016, 65, 1377–1388. [Google Scholar] [CrossRef] [PubMed]
- He, N.; Jia, J.J.; Xie, H.Y.; Li, J.H.; He, Y.; Yin, S.Y.; Liang, R.P.; Jiang, L.; Liu, J.F.; Xu, K.D.; et al. Partial Inhibition of HO-1 Attenuates HMP-Induced Hepatic Regeneration against Liver Injury in Rats. Oxidative Med. Cell. Longev. 2018, 2018, 9108483. [Google Scholar] [CrossRef] [PubMed]
- Leng, J.; Wang, Z.; Fu, C.L.; Zhang, J.; Ren, S.; Hu, J.N.; Jiang, S.; Wang, Y.P.; Chen, C.; Li, W. NF-kappaB and AMPK/PI3K/Akt signaling pathways are involved in the protective effects of Platycodon grandiflorum saponins against acetaminophen-induced acute hepatotoxicity in mice. Phytother. Res. 2018, 32, 2235–2246. [Google Scholar] [CrossRef]
- Baillet, A.; Hograindleur, M.A.; El Benna, J.; Grichine, A.; Berthier, S.; Morel, F.; Paclet, M.H. Unexpected function of the phagocyte NADPH oxidase in supporting hyperglycolysis in stimulated neutrophils: Key role of 6-phosphofructo-2-kinase. FASEB J. 2017, 31, 663–673. [Google Scholar] [CrossRef] [PubMed]
- Rui, L. Energy metabolism in the liver. Compr. Physiol. 2014, 4, 177–197. [Google Scholar] [PubMed]
- Bickler, P.E.; Kelleher, J.A. Fructose-1,6-bisphosphate stabilizes brain intracellular calcium during hypoxia in rats. Stroke 1992, 23, 1617–1622. [Google Scholar] [CrossRef] [PubMed]
- Ashkenazi, A.; Fairbrother, W.J.; Leverson, J.D.; Souers, A.J. From basic apoptosis discoveries to advanced selective BCL-2 family inhibitors. Nat. Rev. Drug Discov. 2017, 16, 273–284. [Google Scholar] [CrossRef]
- Luedde, T.; Kaplowitz, N.; Schwabe, R.F. Cell death and cell death responses in liver disease: Mechanisms and clinical relevance. Gastroenterology 2014, 147, 765–783.e4. [Google Scholar] [CrossRef]
- Kalkavan, H.; Green, D.R. MOMP, cell suicide as a BCL-2 family business. Cell Death Differ. 2018, 25, 46–55. [Google Scholar] [CrossRef]
- Sakurai, T.; Kudo, M.; Watanabe, T.; Itoh, K.; Higashitsuji, H.; Arizumi, T.; Inoue, T.; Hagiwara, S.; Ueshima, K.; Nishida, N.; et al. Hypothermia protects against fulminant hepatitis in mice by reducing reactive oxygen species production. Dig. Dis. 2013, 31, 440–446. [Google Scholar] [CrossRef] [PubMed]
- Lujan, D.A.; Ochoa, J.L.; Hartley, R.S. Cold-inducible RNA binding protein in cancer and inflammation. Wiley Interdiscip. Rev. Rna 2018, 9, e1462. [Google Scholar] [CrossRef] [PubMed]
- Zhong, P.; Huang, H. Recent progress in the research of cold-inducible RNA-binding protein. Future Sci. 2017, 3, Fso246. [Google Scholar] [CrossRef]
- Sakurai, T.; Itoh, K.; Higashitsuji, H.; Nonoguchi, K.; Liu, Y.; Watanabe, H.; Nakano, T.; Fukumoto, M.; Chiba, T.; Fujita, J. Cirp protects against tumor necrosis factor-alpha-induced apoptosis via activation of extracellular signal-regulated kinase. Biochim. Et Biophys. Acta 2006, 1763, 290–295. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Wang, Q.; Zhang, Y.; Yang, D.; Li, D.; Tang, B.; Li, X.; Yang, Y.; Ma, S. Intermittent cold stress enhances features of atherosclerotic plaque instability in apolipoprotein Edeficient mice. Mol. Med. Rep. 2014, 10, 1679–1684. [Google Scholar] [CrossRef]
- Li, S.; Zhang, Z.; Xue, J.; Liu, A.; Zhang, H. Cold-inducible RNA binding protein inhibits H(2)O(2)-induced apoptosis in rat cortical neurons. Brain Res. 2012, 1441, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.N.; Ahn, S.M.; Jang, H.H. Cold-inducible RNA-binding protein, CIRP, inhibits DNA damage-induced apoptosis by regulating p53. Biochem. Biophys. Res. Commun. 2015, 464, 916–921. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Xue, J.; Zhang, H.; Li, S.; Liu, Y.; Xu, D.; Zou, M.; Zhang, Z.; Diao, J. Cloning, expression, and purification of cold inducible RNA-binding protein and its neuroprotective mechanism of action. Brain Res. 2015, 1597, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Xu, B.; Lian, S.; Li, S.Z.; Guo, J.R.; Wang, J.F.; Wang, D.; Zhang, L.P.; Yang, H.M. GABAB receptor mediate hippocampal neuroinflammation in adolescent male and female mice after cold expose. Brain Res. Bull. 2018, 142, 163–175. [Google Scholar] [CrossRef] [PubMed]











© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, P.; Yao, R.; Shi, H.; Liu, Y.; Lian, S.; Yang, Y.; Yang, H.; Li, S. Effects of Cold-inducible RNA-binding Protein (CIRP) on Liver Glycolysis during Acute Cold Exposure in C57BL/6 Mice. Int. J. Mol. Sci. 2019, 20, 1470. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20061470
Liu P, Yao R, Shi H, Liu Y, Lian S, Yang Y, Yang H, Li S. Effects of Cold-inducible RNA-binding Protein (CIRP) on Liver Glycolysis during Acute Cold Exposure in C57BL/6 Mice. International Journal of Molecular Sciences. 2019; 20(6):1470. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20061470
Chicago/Turabian StyleLiu, Peng, Ruizhi Yao, Hongzhao Shi, Yang Liu, Shuai Lian, Yuying Yang, Huanmin Yang, and Shize Li. 2019. "Effects of Cold-inducible RNA-binding Protein (CIRP) on Liver Glycolysis during Acute Cold Exposure in C57BL/6 Mice" International Journal of Molecular Sciences 20, no. 6: 1470. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20061470