Cell Signaling Pathways in Mammary Carcinoma Induced in Rats with Low versus High Inherent Aerobic Capacity


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Characteristics of Induced Tumors
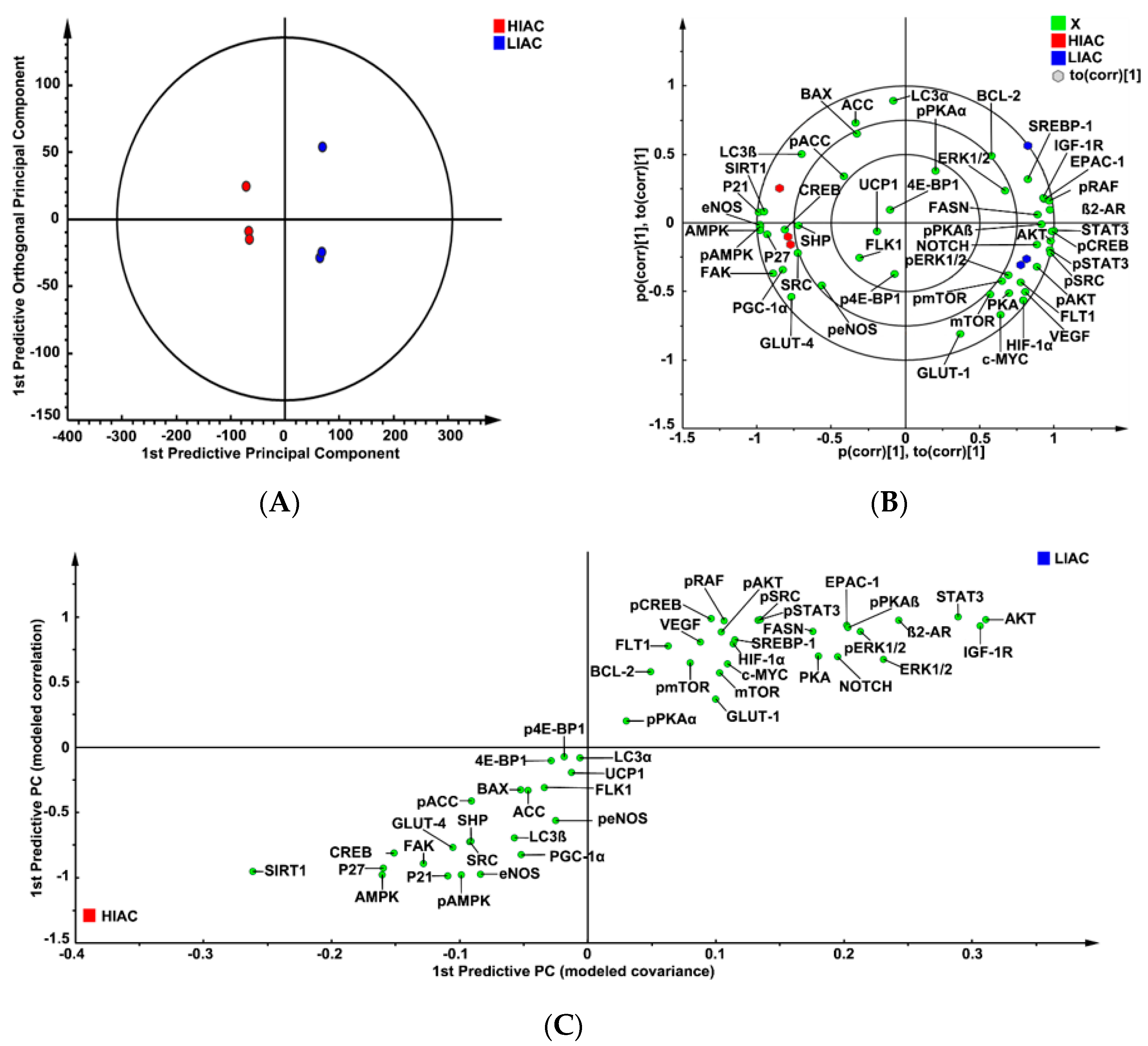
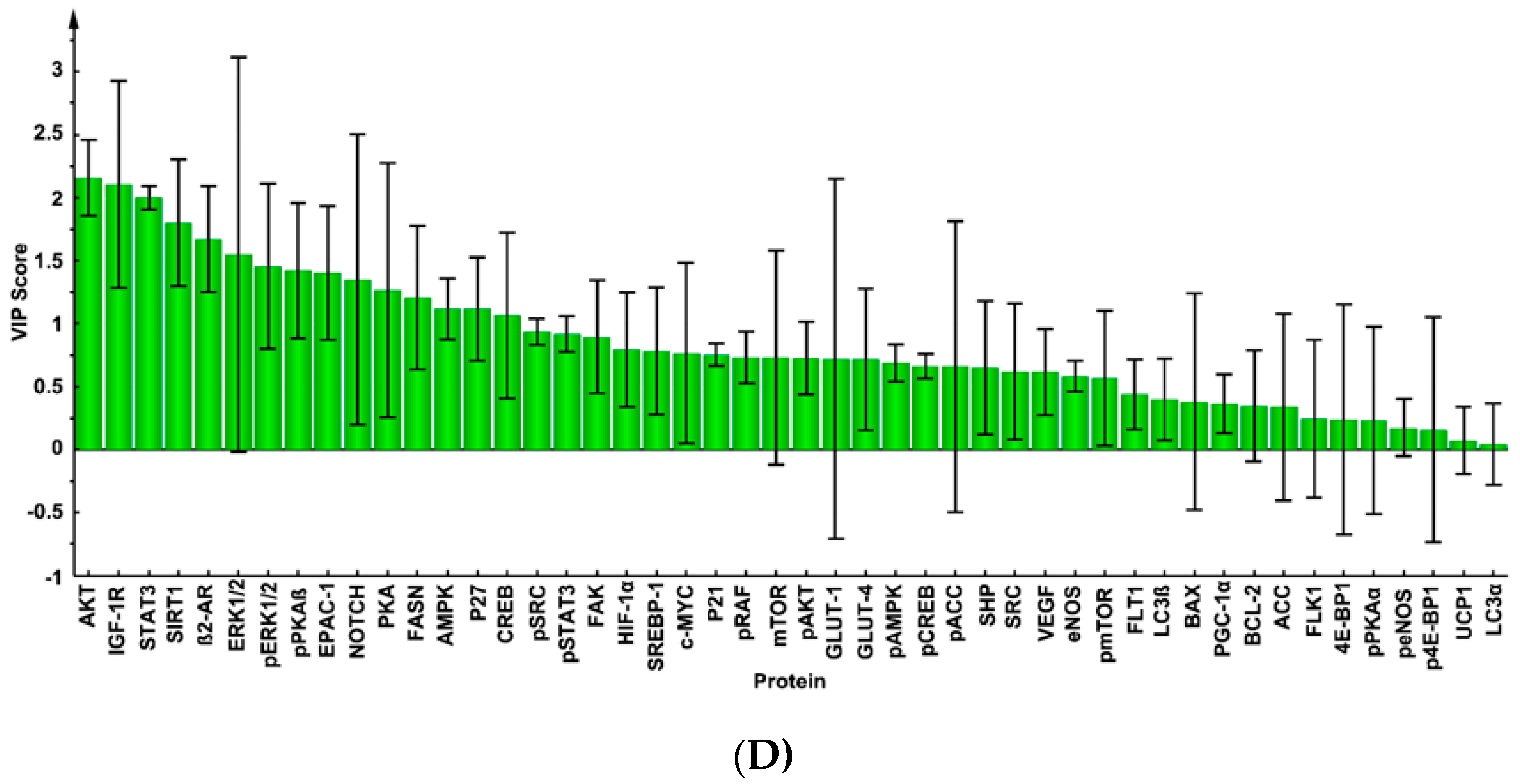
2.2. Mammary Tumor Protein Expression
2.3. Mammary Gland Protein Expression
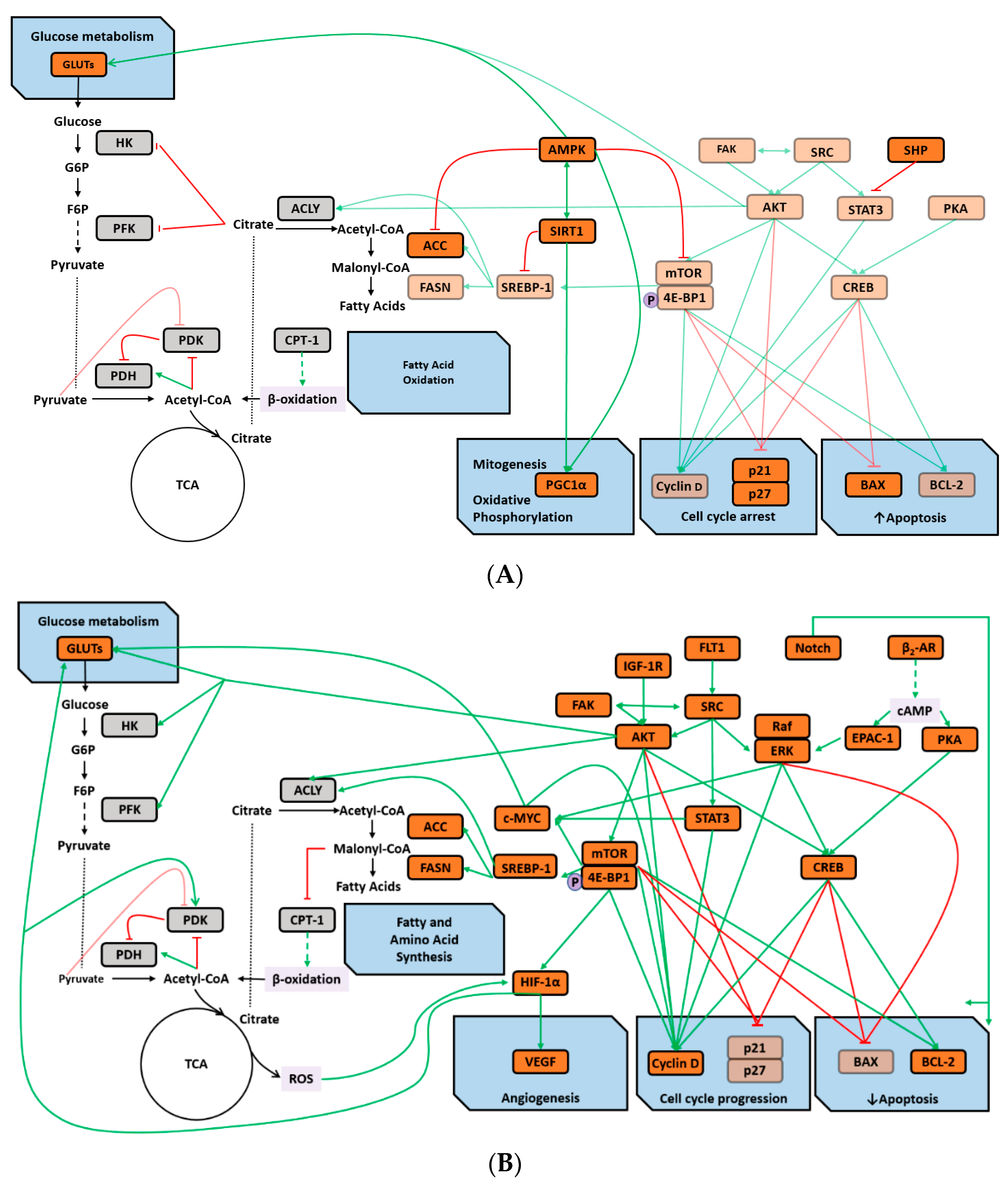
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Immunohistochemistry
4.3. Protein Expression
4.4. Data Evaluation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| 4E-BP1 | 4E Binding Protein 1 |
| ACC | Acetyl-CoA-carboxylase |
| ACLY | ATP-citrate Lyase |
| AKT | Protein Kinase B |
| AMPK | AMP-activated kinase |
| BAX | BCL-2-associated X protein. |
| β2-AR | β2-adrenergic Receptor |
| BCL-2 | B cell lymphoma/leukaemia-2 protein |
| cAMP | Cyclic Adenosine Monophosphate |
| CKI | Cyclin-dependent Kinase Inhibitor |
| c-MYC | Oncogenic transcription factor |
| CPT-I | Carnitine Palmitoyltransferase I |
| CREB | Cyclic-AMP-responsive element binding protein |
| DPC | Days Post Carcinogen |
| EPAC | Guanine-nucleotide exchange proteins activated by cAMP 1 |
| ER | Estrogen Receptor |
| ERK1/2 | Mitogen-activated protein kinase, extracellular signal regulated kinases 1 and 2 |
| FAK | Focal Adhesion Kinase |
| FASN | Fatty Acid Synthase |
| FLT1 | Vascular endothelial growth factor receptor 1 |
| GLUTs | Glucose transporters |
| HIAC | High Inherent Aerobic Capacity |
| HIF-1α | Hypoxia Inducible Factor-1α |
| HK | Hexokinase |
| HRS | Hormone Receptor Signaling |
| IAC | Inherent Aerobic Capacity |
| IGF-1R | Insulin Growth Factor 1 Receptor |
| JKCI | Jack-knifed Confidence Intervals |
| LIAC | Low Inherent Aerobic Capacity |
| MNU | 1-methyl-1-nitrosurea |
| mTOR | Mammalian Target of Rapamycin |
| OPLS-DA | Orthogonal Projections to Latent Structures for Discriminant Analysis |
| p21 | Cyclin-dependent kinase inhibitor 1 |
| p27 | Cyclin dependent kinase inhibitor 1B |
| p4E-BP1 | Phospho 4E Binding Protein |
| pAKT | Phospho protein Kinase B |
| PCA | Principle Component Analysis |
| pCREB | Phospho cyclic-AMP-responsive Element Binding protein |
| PDH | Pyruvate Dehydrogenase |
| PDK | Pyruvate Dehydrogenase Kinase |
| pERK1/2 | Phosopho mitogen-activated protein kinase, extracellular signal regulated kinases 1 and 2 |
| PFK | Phosphofructokinase |
| PGC | Peroxisome proliferator-activated receptor gamma coactivator |
| PGC-1α | Peroxisome proliferator-activated receptor gamma coactivator 1-alpha |
| PKA | Protein kinsae A |
| pmTOR | Phospho mammalian target of rapamycin |
| pPKAα/β | Protein kinsae A alpha / beta |
| PR | Progesterone Receptor |
| pSRC | Phospho non-receptor tyrosine kinase SRC |
| pSTAT3 | Phospho signal transducer and activator of transcription 3 |
| RAF | Rapidly Accelerated Fibrosarcoma kinase |
| ROS | Reactive Oxygen Species |
| SIRT-1 | Silent mating type information regulation 2 homolog 1 |
| SRC | Non-receptor tyrosine kinase SRC |
| SREBP1 | Sterol Regulatory Element-binding Protein 1 |
| STAT3 | Signal and transducer of transcription 3 |
| TCA | Tricarboxylic Acid |
| VEGF | Vascular Endothelial Growth Factor; |
| VIP | Variable Importance Projection |
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2018. CA Cancer J. Clin. 2018, 68, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Brenner, D.R.; Brockton, N.T.; Kotsopoulos, J.; Cotterchio, M.; Boucher, B.A.; Courneya, K.S.; Knight, J.A.; Olivotto, I.A.; Quan, M.L.; Friedenreich, C.M. Breast cancer survival among young women: A review of the role of modifiable lifestyle factors. Cancer Causes Control 2016, 27, 459–472. [Google Scholar] [CrossRef]
- Heitz, A.E.; Baumgartner, R.N.; Baumgartner, K.B.; Boone, S.D. Healthy lifestyle impact on breast cancer-specific and all-cause mortality. Breast Cancer Res. Treat. 2018, 167, 171–181. [Google Scholar] [CrossRef] [PubMed]
- De Boer, M.C.; Worner, E.A.; Verlaan, D.; van Leeuwen, P.A.M. The Mechanisms and Effects of Physical Activity on Breast Cancer. Clin. Breast Cancer 2017, 17, 272–278. [Google Scholar] [CrossRef] [PubMed]
- Friedenreich, C.M. Physical activity and breast cancer: Review of the epidemiologic evidence and biologic mechanisms. Recent Results Cancer Res. 2011, 188, 125–139. [Google Scholar] [PubMed]
- Hojman, P.; Gehl, J.; Christensen, J.F.; Pedersen, B.K. Molecular Mechanisms Linking Exercise to Cancer Prevention and Treatment. Cell Metab. 2018, 27, 10–21. [Google Scholar] [CrossRef]
- Sternfeld, B.; Goldman-Rosas, L. A systematic approach to selecting an appropriate measure of self-reported physical activity or sedentary behavior. J. Phys. Act. Health 2012, 9 (Suppl. 1), S19–S28. [Google Scholar] [CrossRef]
- Eriksen, L.; Curtis, T.; Gronbaek, M.; Helge, J.W.; Tolstrup, J.S. The association between physical activity, cardiorespiratory fitness and self-rated health. Prev. Med. 2013, 57, 900–902. [Google Scholar] [CrossRef]
- Wiggins, M.S.; Simonavice, E.M. Cancer prevention, aerobic capacity, and physical functioning in survivors related to physical activity: A recent review. Cancer Manag. Res. 2010, 2, 157–164. [Google Scholar] [CrossRef] [PubMed]
- Bouchard, C.; Blair, S.N.; Katzmarzyk, P.T. Less Sitting, More Physical Activity, or Higher Fitness? Mayo Clin. Proc. 2015, 90, 1533–1540. [Google Scholar] [CrossRef]
- Sarzynski, M.A.; Ghosh, S.; Bouchard, C. Genomic and transcriptomic predictors of response levels to endurance exercise training. J. Physiol. 2017, 595, 2931–2939. [Google Scholar] [CrossRef] [PubMed]
- Koch, L.G.; Britton, S.L. Artificial selection for intrinsic aerobic endurance running capacity in rats. Physiol. Genom. 2001, 5, 45–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koch, L.G.; Britton, S.L. Theoretical and Biological Evaluation of the Link between Low Exercise Capacity and Disease Risk. Cold Spring Harb. Perspect. Med. 2018, 8, a029868. [Google Scholar] [CrossRef]
- Thyfault, J.P.; Morris, E.M. Intrinsic (Genetic) Aerobic Fitness Impacts Susceptibility for Metabolic Disease. Exerc. Sport Sci. Rev. 2017, 45, 7–15. [Google Scholar] [CrossRef] [Green Version]
- Ritchie, R.H.; Leo, C.H.; Qin, C.; Stephenson, E.J.; Bowden, M.A.; Buxton, K.D.; Lessard, S.J.; Rivas, D.A.; Koch, L.G.; Britton, S.L.; et al. Low intrinsic exercise capacity in rats predisposes to age-dependent cardiac remodeling independent of macrovascular function. Am. J. Physiol. Heart Circ. Physiol. 2013, 304, H729–H739. [Google Scholar] [CrossRef]
- Rivas, D.A.; Lessard, S.J.; Saito, M.; Friedhuber, A.M.; Koch, L.G.; Britton, S.L.; Yaspelkis, B.B., III; Hawley, J.A. Low intrinsic running capacity is associated with reduced skeletal muscle substrate oxidation and lower mitochondrial content in white skeletal muscle. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2011, 300, R835–R843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petriz, B.A.; Gomes, C.P.; Almeida, J.A.; de Oliveira, G.P.J.; Ribeiro, F.M.; Pereira, R.W.; Franco, O.L. The Effects of Acute and Chronic Exercise on Skeletal Muscle Proteome. J. Cell. Physiol. 2017, 232, 257–269. [Google Scholar] [CrossRef]
- Karvinen, S.; Silvennoinen, M.; Vainio, P.; Sistonen, L.; Koch, L.G.; Britton, S.L.; Kainulainen, H. Effects of intrinsic aerobic capacity, aging and voluntary running on skeletal muscle sirtuins and heat shock proteins. Exp. Gerontol. 2016, 79, 46–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gavini, C.K.; Mukherjee, S.; Shukla, C.; Britton, S.L.; Koch, L.G.; Shi, H.; Novak, C.M. Leanness and heightened nonresting energy expenditure: Role of skeletal muscle activity thermogenesis. Am. J. Physiol. Endocrinol. Metab. 2014, 306, E635–E647. [Google Scholar] [CrossRef] [PubMed]
- Gan, L.; Ma, D.; Li, M.; Yang, F.C.; Rogers, R.S.; Wheatley, J.L.; Koch, L.G.; Britton, S.L.; Thyfault, J.P.; Geiger, P.C.; et al. Region-specific differences in bioenergetic proteins and protein response to acute high fat diet in brains of low and high capacity runner rats. Neurosci. Lett. 2018, 674, 49–53. [Google Scholar] [CrossRef] [PubMed]
- Cooper, M.A.; Jack, M.M.; Ryals, J.M.; Hayley, P.; Escher, T.; Koch, L.G.; Britton, S.L.; Raupp, S.M.; Winter, M.K.; McCarson, K.E.; et al. Rats bred for low and high running capacity display alterations in peripheral tissues and nerves relevant to neuropathy and pain. Brain Behav. 2017, 7, e00780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cox-York, K.A.; Sheflin, A.M.; Foster, M.T.; Gentile, C.L.; Kahl, A.; Koch, L.G.; Britton, S.L.; Weir, T.L. Ovariectomy results in differential shifts in gut microbiota in low versus high aerobic capacity rats. Physiol. Rep. 2015, 3, e12488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, H.J.; Jones, L.W.; Koch, L.G.; Britton, S.L.; Neil, E.S.; McGinley, J.N. Inherent aerobic capacity-dependent differences in breast carcinogenesis. Carcinogenesis 2017, 38, 920–928. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Jiang, W.; McGinley, J.N.; Thompson, H.J. Defining the role of histone deacetylases in the inhibition of mammary carcinogenesis by dietary energy restriction (DER): Effects of suberoylanilide hydroxamic acid (SAHA) and DER in a rat model. Cancer Prev. Res. 2013, 6, 290–298. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.; Zhu, Z.; Thompson, H.J. Effects of limiting energy availability via diet and physical activity on mammalian target of rapamycin-related signaling in rat mammary carcinomas. Carcinogenesis 2013, 34, 378–387. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Jiang, W.; Zacher, J.H.; Neil, E.S.; McGinley, J.N.; Thompson, H.J. Effects of energy restriction and wheel running on mammary carcinogenesis and host systemic factors in a rat model. Cancer Prev. Res. 2012, 5, 414–422. [Google Scholar] [CrossRef] [PubMed]
- Thompson, H.J.; Wolfe, P.; McTiernan, A.; Jiang, W.; Zhu, Z. Wheel running-induced changes in plasma biomarkers and carcinogenic response in the 1-methyl-1-nitrosourea-induced rat model for breast cancer. Cancer Prev. Res. 2010, 3, 1484–1492. [Google Scholar] [CrossRef] [PubMed]
- Thompson, H.J.; Jiang, W.; Zhu, Z. Candidate mechanisms accounting for effects of physical activity on breast carcinogenesis. IUBMB Life 2009, 61, 895–901. [Google Scholar] [CrossRef]
- Zhu, Z.; Jiang, W.; McGinley, J.N.; Thompson, H.J. Energetics and mammary carcinogenesis: Effects of moderate-intensity running and energy intake on cellular processes and molecular mechanisms in rats. J. Appl. Physiol. 2009, 106, 911–918. [Google Scholar] [CrossRef]
- DeMarco, V.G.; Johnson, M.S.; Ma, L.; Pulakat, L.; Mugerfeld, I.; Hayden, M.R.; Garro, M.; Knight, W.; Britton, S.L.; Koch, L.G.; et al. Overweight female rats selectively breed for low aerobic capacity exhibit increased myocardial fibrosis and diastolic dysfunction. Am. J. Physiol. Heart Circ. Physiol. 2012, 302, H1667–H1682. [Google Scholar] [CrossRef] [Green Version]
- Lessard, S.J.; Rivas, D.A.; Chen, Z.P.; van Denderen, B.J.; Watt, M.J.; Koch, L.G.; Britton, S.L.; Kemp, B.E.; Hawley, J.A. Impaired skeletal muscle beta-adrenergic activation and lipolysis are associated with whole-body insulin resistance in rats bred for low intrinsic exercise capacity. Endocrinology 2009, 150, 4883–4891. [Google Scholar] [CrossRef] [PubMed]
- Pajvani, U.B.; Accili, D. The new biology of diabetes. Diabetologia 2015, 58, 2459–2468. [Google Scholar] [CrossRef] [Green Version]
- Landor, S.K.; Mutvei, A.P.; Mamaeva, V.; Jin, S.; Busk, M.; Borra, R.; Gronroos, T.J.; Kronqvist, P.; Lendahl, U.; Sahlgren, C.M. Hypo- and hyperactivated Notch signaling induce a glycolytic switch through distinct mechanisms. Proc. Natl. Acad. Sci. USA 2011, 108, 18814–18819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, R.L.; Soeters, M.R.; Wust, R.C.I.; Houtkooper, R.H. Metabolic flexibility as an adaptation to energy resources and requirements in health and disease. Endocr. Rev. 2018, 39, 489–517. [Google Scholar] [CrossRef]
- Thompson, H.J.; Neuhouser, M.L.; Lampe, J.W.; McGinley, J.N.; Neil, E.S.; Schwartz, Y.; McTiernan, A. Effect of low or high glycemic load diets on experimentally induced mammary carcinogenesis in rats. Mol. Nutr. Food Res. 2016, 60, 1416–1426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, H.J.; McGinley, J.N.; Rothhammer, K.; Singh, M. Rapid induction of mammary intraductal proliferations, ductal carcinoma in situ and carcinomas by the injection of sexually immature female rats with 1-methyl-1-nitrosourea. Carcinogenesis 1995, 16, 2407–2411. [Google Scholar] [CrossRef] [PubMed]
- Thompson, H.J.; Singh, M.; McGinley, J. Classification of premalignant and malignant lesions developing in the rat mammary gland after injection of sexually immature rats with 1-methyl-1-nitrosourea. J. Mammary Gland Boil. Neoplasia 2000, 5, 201–210. [Google Scholar] [CrossRef]
- Knott, K.K.; McGinley, J.N.; Lubet, R.A.; Steele, V.E.; Thompson, H.J. Effect of the aromatase inhibitor vorozole on estrogen and progesterone receptor content of rat mammary carcinomas induced by 1-methyl-1-nitrosourea. Breast Cancer Res. Treat. 2001, 70, 171–183. [Google Scholar] [CrossRef]
- Sokal, R.R.; Rohlf, F.J. Biometry the Principles and Practice of Statistics in Biological Research; W.H. Freeman: New York, NY, USA, 1995. [Google Scholar]
- Gabrielsson, J.; Jonsson, H.; Airiau, C.; Schmidt, B.; Escott, R.; Trygg, J. OPLS methodology for analysis of pre-processing effects on spectroscopic data. Chemom. Intell. Lab. Syst. 2006, 84, 153–158. [Google Scholar] [CrossRef]
- Wiklund, S. Multivariate Analysis for Omics. Available online: http://metabolomics.se/Courses/MVA/MVA%20in%20Omics_Handouts_Exercises_Solutions_Thu-Fri.pdf (accessed on 10 August 2011).
- Trygg, J.; Wold, S. Orthogonal projections to latent structures (O-PLS). J. Chemom. 2002, 16, 119–128. [Google Scholar] [CrossRef]
- Wiklund, S.; Johansson, E.; Sjostrom, L.; Mellerowicz, E.J.; Edlund, U.; Shockcor, J.P.; Gottfries, J.; Moritz, T.; Trygg, J. Visualization of GC/TOF-MS-based metabolomics data for identification of biochemically interesting compounds using OPLS class models. Anal. Chem. 2008, 80, 115–122. [Google Scholar] [CrossRef] [PubMed]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lutsiv, T.; McGinley, J.N.; Neil, E.S.; Thompson, H.J. Cell Signaling Pathways in Mammary Carcinoma Induced in Rats with Low versus High Inherent Aerobic Capacity. Int. J. Mol. Sci. 2019, 20, 1506. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20061506
Lutsiv T, McGinley JN, Neil ES, Thompson HJ. Cell Signaling Pathways in Mammary Carcinoma Induced in Rats with Low versus High Inherent Aerobic Capacity. International Journal of Molecular Sciences. 2019; 20(6):1506. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20061506
Chicago/Turabian StyleLutsiv, Tymofiy, John N. McGinley, Elizabeth S. Neil, and Henry J. Thompson. 2019. "Cell Signaling Pathways in Mammary Carcinoma Induced in Rats with Low versus High Inherent Aerobic Capacity" International Journal of Molecular Sciences 20, no. 6: 1506. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20061506