Placenta Transcriptome Profiling in Intrauterine Growth Restriction (IUGR)

 , , ,
, , ,  , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. Statistics of the Placenta Transcriptome
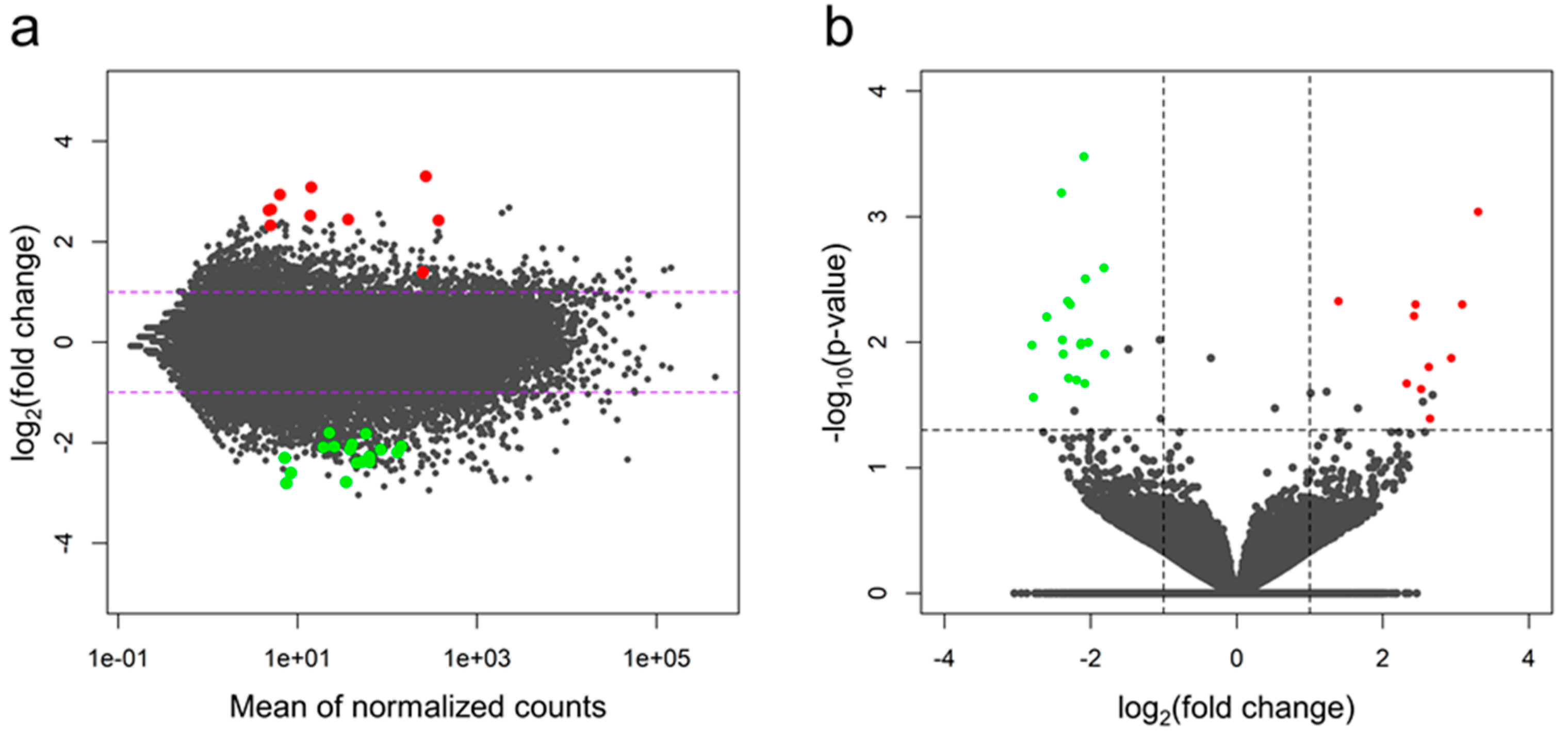
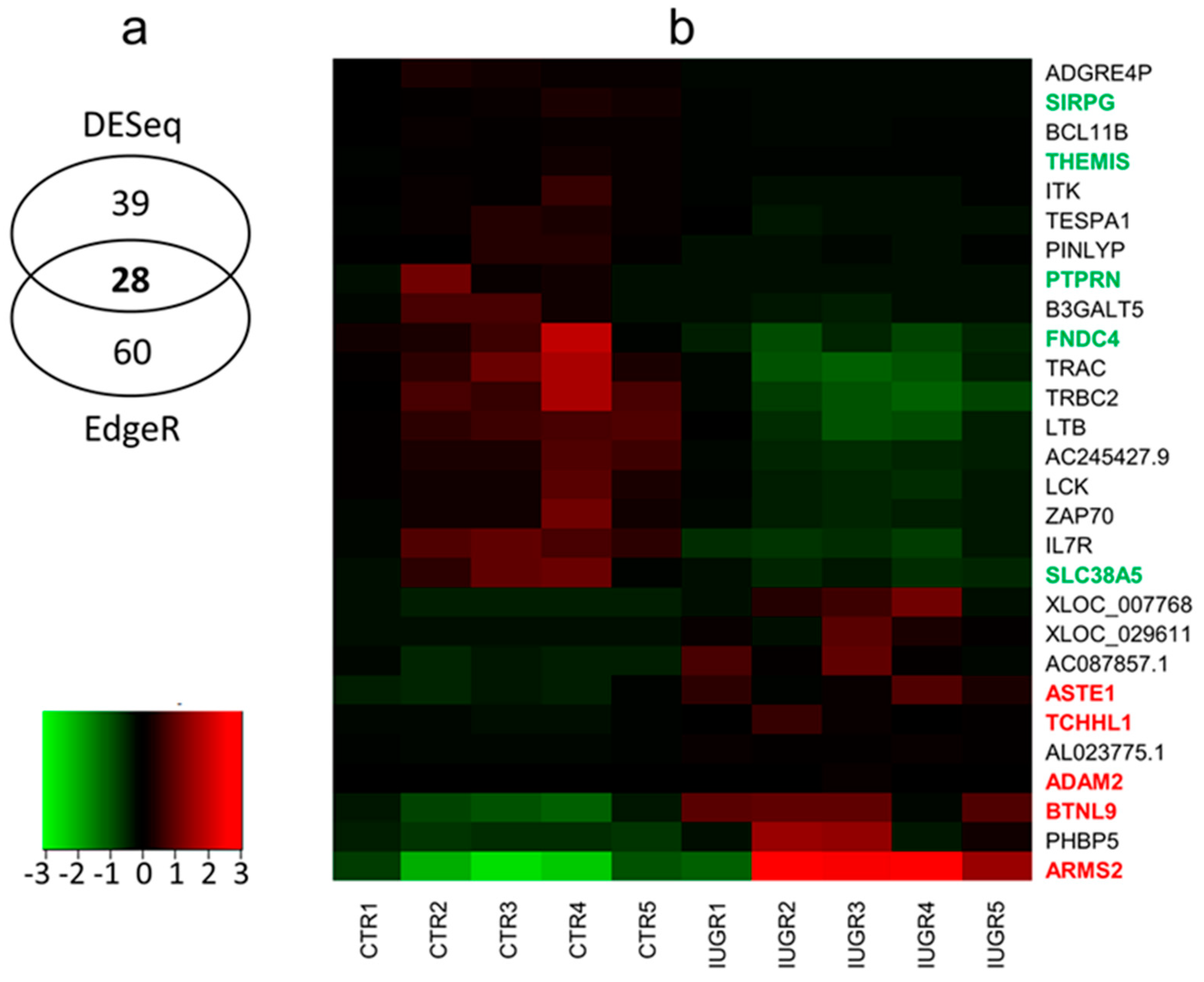
2.2. Differentially-expressed Genes (DEGs) and Functional Annotations
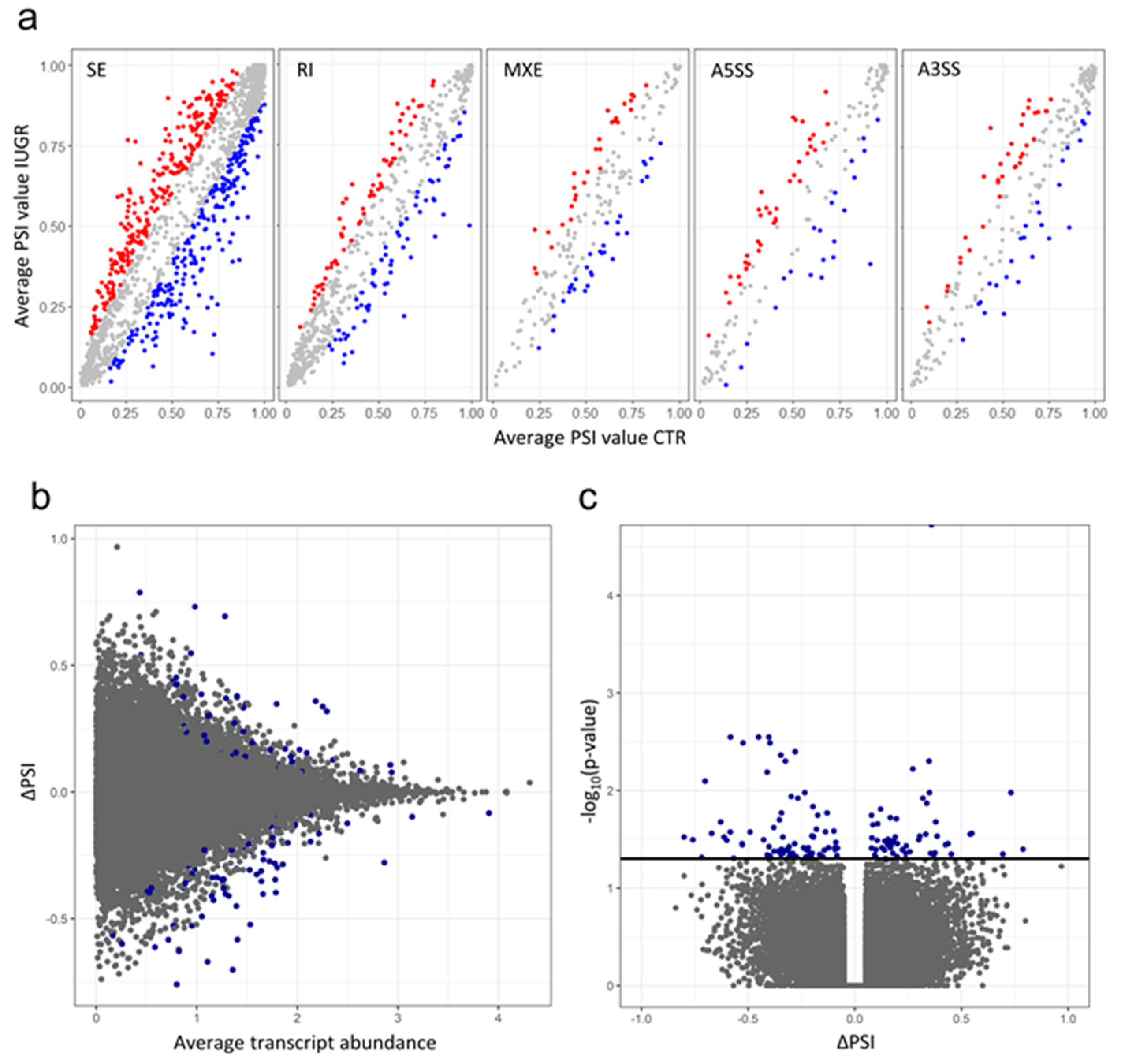
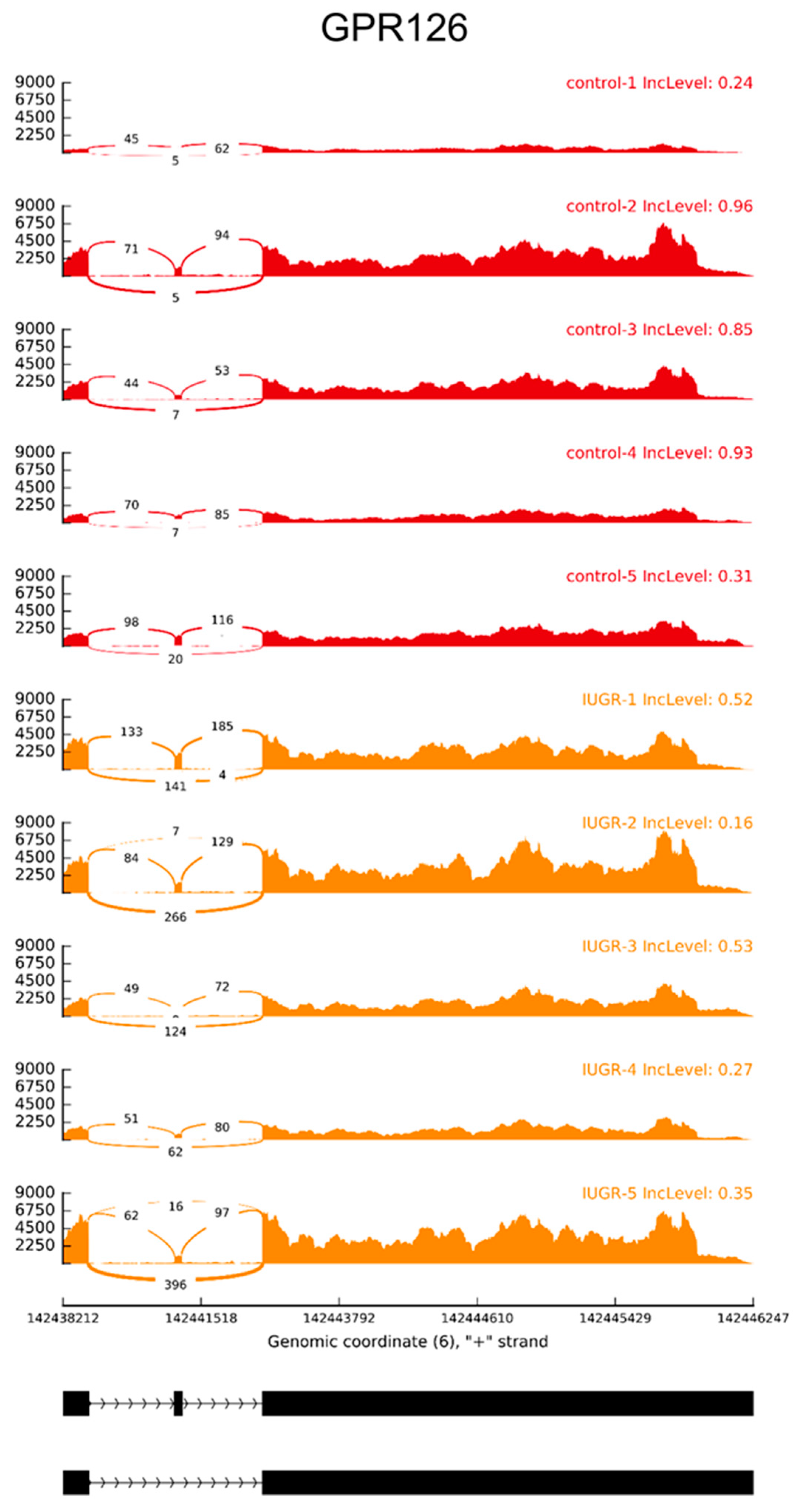
2.3. Alternative Splicing Patterns
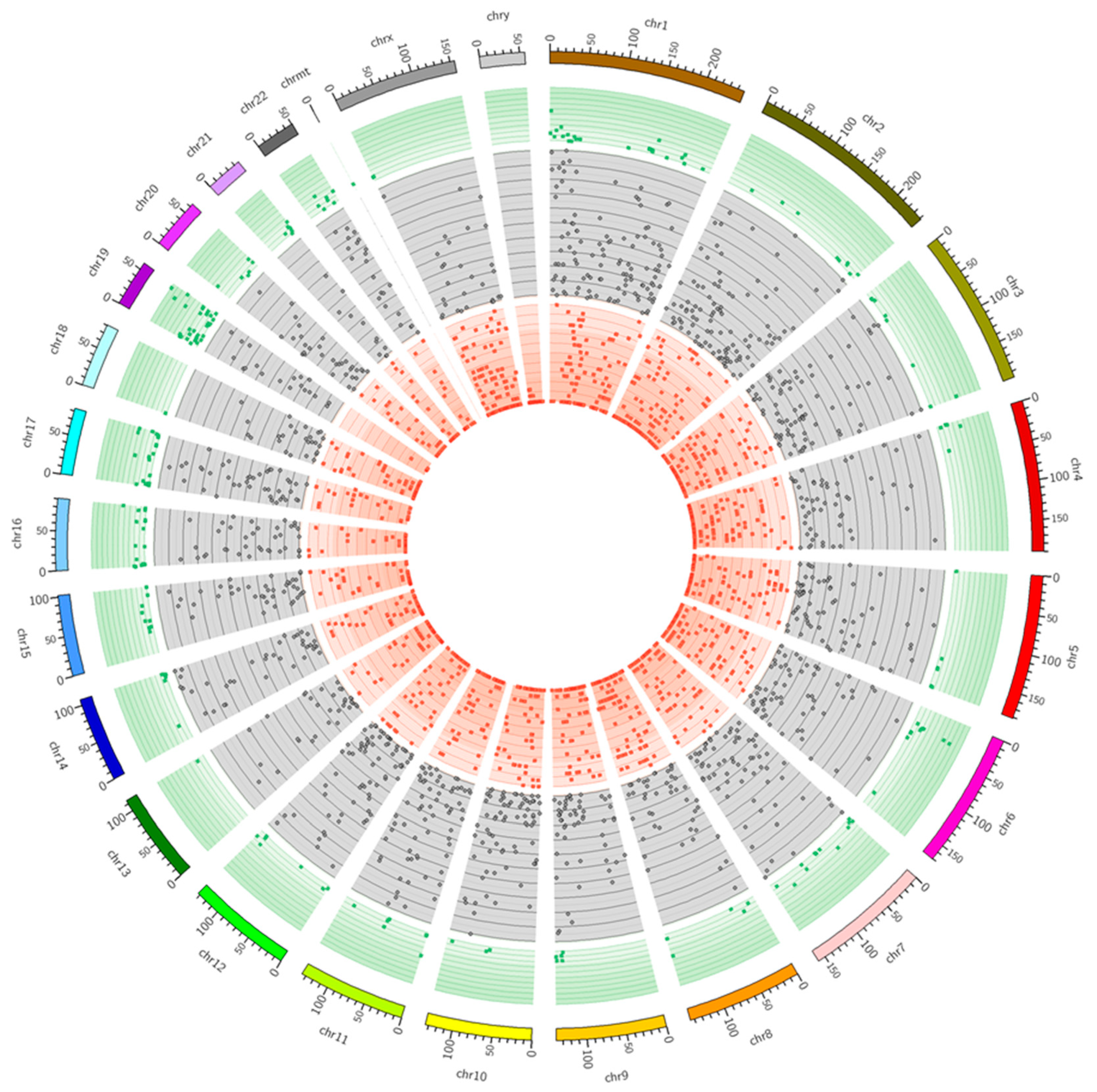
2.4. SNV Calling and RNA Editing Prediction
3. Discussion
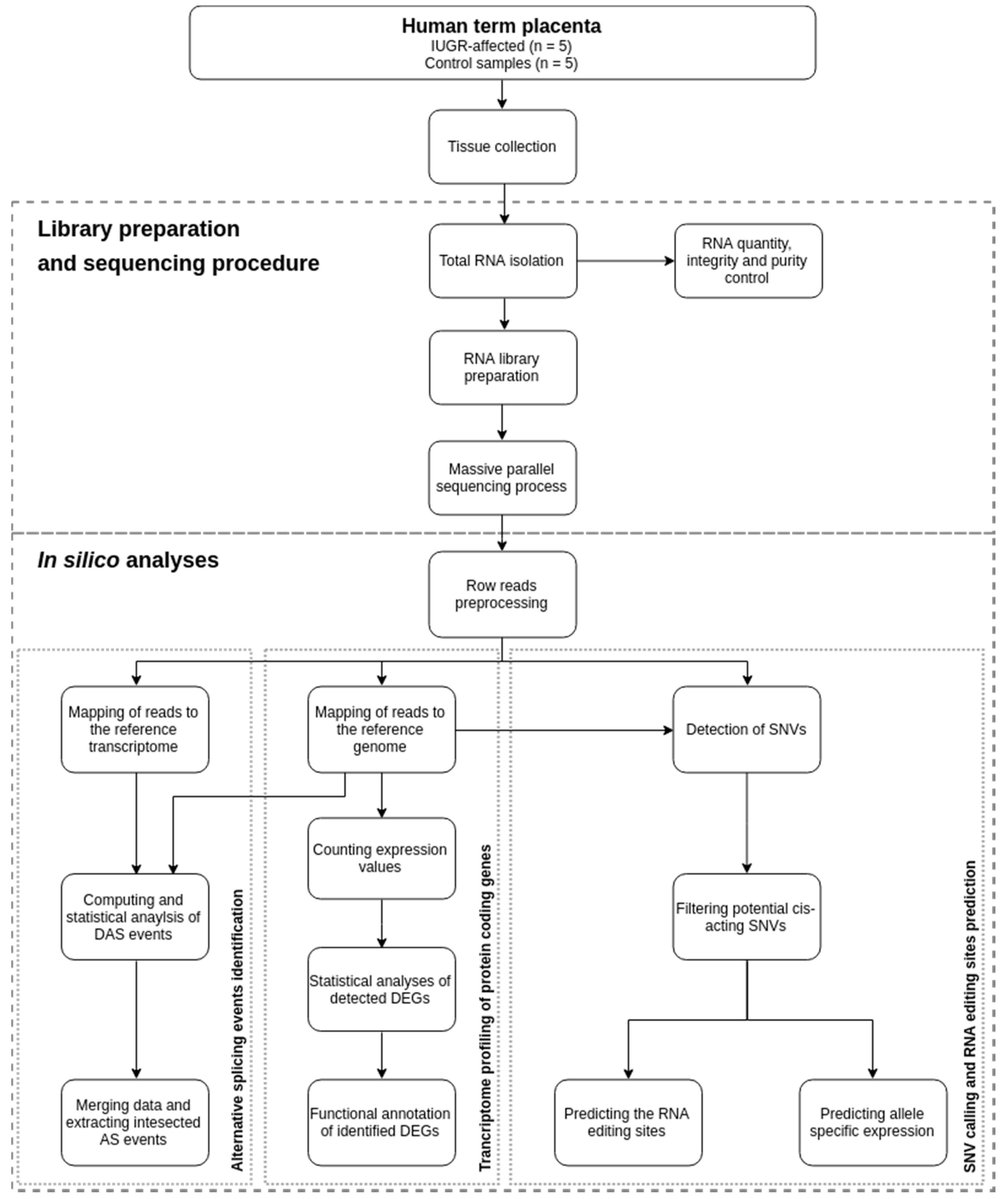
4. Materials and Methods
4.1. Ethics Statement
4.2. Clinical Characteristics of Placental Samples
4.3. Library Preparation and Sequencing Procedure
4.4. Quality Control and Mapping Processes
4.5. Transcriptome Profiling of Protein-coding Genes
4.6. Alternative Splicing Events
4.7. SNV calling and RNA Editing Sites Prediction
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Availability of Supporting Data
References
- Albu, A.R.; Anca, A.F.; Horhoianu, V.V.; Horhoianu, I.A. Predictive factors for intrauterine growth restriction. J. Med. Life 2014, 7, 165–171. [Google Scholar] [PubMed]
- Vayssière, C.; Sentilhes, L.; Ego, A.; Bernard, C.; Cambourieu, D.; Flamant, C.; Gascoin, G.; Gaudineau, A.; Grangé, G.; Houfflin-Debarge, V.; et al. Fetal growth restriction and intra-uterine growth restriction: Guidelines for clinical practice from the French College of Gynaecologists and Obstetricians. Eur. J. Obstet. Gynecol. Reprod. Biol. 2015, 193, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Unterscheider, J.; Daly, S.; Geary, M.P.; Kennelly, M.M.; McAuliffe, F.M.; O’Donoghue, K.; Hunter, A.; Morrison, J.J.; Burke, G.; Dicker, P.; et al. Optimizing the Definition of Intrauterine Growth Restriction. Obstet. Gynecol. Surv. 2013, 68, 549–551. [Google Scholar] [CrossRef]
- Sharma, D.; Sharma, P.; Shastri, S. Genetic, metabolic and endocrine aspect of intrauterine growth restriction: An update. J. Matern. Neonatal Med. 2017, 30, 2263–2275. [Google Scholar] [CrossRef] [PubMed]
- Carducci, B.; Bhutta, Z.A. Care of the growth-restricted newborn. Best Pract. Res. Clin. Obstet. Gynaecol. 2018, 49, 103–116. [Google Scholar] [CrossRef]
- Sharma, D.; Shastri, S.; Farahbakhsh, N.; Sharma, P. Intrauterine growth restriction—Part 1. J. Matern. Neonatal Med. 2016, 29, 3977–3987. [Google Scholar] [CrossRef]
- Figueras, F.; Gratacos, E. An integrated approach to fetal growth restriction. Best Pract. Res. Clin. Obstet. Gynaecol. 2017, 38, 48–58. [Google Scholar] [CrossRef]
- Hadlock, F.P.; Harrist, R.B.; Sharman, R.S.; Deter, R.L.; Park, S.K. Estimation of fetal weight with the use of head, body, and femur measurements—A prospective study. Am. J. Obstet. Gynecol. 1985, 151, 333–337. [Google Scholar] [CrossRef]
- Sahoo, R.K.; Nayak, S.; Panda, S.K.; Pati, P.B.; Sahoo, S.; Sahu, M.C. Doppler assessment of the fetus with intrauterine growth restriction. Int. J. Pharm. Sci. Rev. Res. 2015, 32, 162–170. [Google Scholar] [CrossRef]
- Villalaín, C.; Herraiz, I.; Quezada, M.S.; Gómez-Arriaga, P.I.; Gómez-Montes, E.; Galindo, A. Fetal Biometry and Doppler Study for the Assessment of Perinatal Outcome in Stage I Late-Onset Fetal Growth Restriction. Fetal Diagn. Ther. 2018, 44, 1–7. [Google Scholar] [CrossRef]
- Sharma, D.; Farahbakhsh, N.; Shastri, S.; Sharma, P. Intrauterine growth restriction—Part 2. J. Matern. Neonatal Med. 2016, 29, 4037–4048. [Google Scholar] [CrossRef] [PubMed]
- Gordijn, S.J.; Beune, I.M.; Thilaganathan, B.; Papageorghiou, A.; Baschat, A.A.; Baker, P.N.; Silver, R.M.; Wynia, K.; Ganzevoort, W. Consensus definition of fetal growth restriction: A Delphi procedure. Ultrasound Obstet. Gynecol. 2016, 48, 333–339. [Google Scholar] [CrossRef] [PubMed]
- Capros, H.; Scoricova, I.; Mihalcean, L. Intrauterine growth restriction: Contemporary issues in diagnosis and management. Mold. Med. J. 2017, 60, 26–30. [Google Scholar] [CrossRef]
- Zeitlin, J.; El Ayoubi, M.; Jarreau, P.H.; Draper, E.S.; Blondel, B.; Künzel, W.; Cuttini, M.; Kaminski, M.; Gortner, L.; Van Reempts, P.; et al. Impact of fetal growth restriction on mortality and morbidity in a very preterm birth cohort. J. Pediatr. 2010, 157, 733.e1–739.e1. [Google Scholar] [CrossRef]
- Serena, C.; Marchetti, G.; Rambaldi, M.P.; Ottanelli, S.; Di Tommaso, M.; Avagliano, L.; Pieralli, A.; Mello, G.; Mecacci, F. Stillbirth and fetal growth restriction. J. Matern. Neonatal Med. 2013, 26, 16–20. [Google Scholar] [CrossRef]
- Sharma, D.; Shastri, S.; Sharma, P. Intrauterine Growth Restriction: Antenatal and Postnatal Aspects. Clin. Med. Insights Pediatr. 2016, 10, 67–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Damodaram, M.; Story, L.; Kulinskaya, E.; Rutherford, M.; Kumar, S. Early adverse perinatal complications in preterm growth-restricted fetuses. Aust. N. Z. J. Obstet. Gynaecol. 2011, 51, 204–209. [Google Scholar] [CrossRef]
- Murray, E.; Fernandes, M.; Fazel, M.; Kennedy, S.; Villar, J.; Stein, A. Differential effect of intrauterine growth restriction on childhood neurodevelopment: A systematic review. BJOG An Int. J. Obstet. Gynaecol. 2015, 122, 1062–1072. [Google Scholar] [CrossRef]
- Wixey, J.A.; Chand, K.K.; Colditz, P.B.; Bjorkman, S.T. Review: Neuroinflammation in intrauterine growth restriction. Placenta 2017, 54, 117–124. [Google Scholar] [CrossRef] [Green Version]
- Gardosi, J.; Madurasinghe, V.; Williams, M.; Malik, A.; Francis, A. Maternal and fetal risk factors for stillbirth: Population based study. BMJ 2013, 346, f108. [Google Scholar] [CrossRef]
- Mayer, C.; Joseph, K.S. Fetal growth: A review of terms, concepts and issues relevant to obstetrics. Ultrasound Obstet. Gynecol. 2013, 41, 136–145. [Google Scholar] [CrossRef] [PubMed]
- Sarvari, S.I.; Rodriguez-Lopez, M.; Nuñez-Garcia, M.; Sitges, M.; Sepulveda-Martinez, A.; Camara, O.; Butakoff, C.; Gratacos, E.; Bijnens, B.; Crispi, F. Persistence of Cardiac Remodeling in Preadolescents With Fetal Growth Restriction. Circ. Cardiovasc. Imaging 2017, 10, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Yzydorczyk, C.; Armengaud, J.B.; Peyter, A.C.; Chehade, H.; Cachat, F.; Juvet, C.; Siddeek, B.; Simoncini, S.; Sabatier, F.; Dignat-George, F.; et al. Endothelial dysfunction in individuals born after fetal growth restriction: Cardiovascular and renal consequences and preventive approaches. J. Dev. Orig. Health Dis. 2017, 8, 448–464. [Google Scholar] [CrossRef] [PubMed]
- Haroun, H.S.W. Intrauterine Growth Restriction. Anat. Physiol. Biochem. Int. J. 2017, 1, 555572. [Google Scholar] [CrossRef]
- Eydoux, P.; Choiset, A.; le Porrier, N.; Thépot, F.; Szpiro-Tapia, S.; Alliet, J.; Ramond, S.; Viel, J.F.; Gautier, E.; Morichon, N.; et al. Chromosomal prenatal diagnosis: Study of 936 cases of intrauterine abnormalities after ultrasound assessment. Prenat. Diagn. 1989, 9, 255–269. [Google Scholar] [CrossRef] [PubMed]
- Veerbeek, J.H.W.; Nikkels, P.G.J.; Torrance, H.L.; Gravesteijn, J.; Post Uiterweer, E.D.; Derks, J.B.; Koenen, S.V.; Visser, G.H.A.; Van Rijn, B.B.; Franx, A. Placental pathology in early intrauterine growth restriction associated with maternal hypertension. Placenta 2014, 35, 696–701. [Google Scholar] [CrossRef] [PubMed]
- Groom, K.M.; David, A.L. The role of aspirin, heparin, and other interventions in the prevention and treatment of fetal growth restriction. Am. J. Obstet. Gynecol. 2018, 218, S829–S840. [Google Scholar] [CrossRef]
- Salomon, L.J.; Malan, V. Bilan étiologique du retard de croissance intra-utérin (RCIU). La Revue Sage-Femme 2014, 13, 99–110. [Google Scholar] [CrossRef]
- Deshpande, S.S.; Balasinor, N.H. Placental Defects: An Epigenetic Perspective. Reprod. Sci. 2018, 25, 1143–1160. [Google Scholar] [CrossRef]
- John, R.M. Imprinted genes and the regulation of placental endocrine function: Pregnancy and beyond. Placenta 2017, 56, 86–90. [Google Scholar] [CrossRef] [Green Version]
- Lipka, A.; Paukszto, L.; Majewska, M.; Jastrzebski, J.P.; Myszczynski, K.; Panasiewicz, G.; Szafranska, B. Identification of differentially-expressed placental transcripts during multiple gestations in the Eurasian beaver (Castor fiber L.). Reprod. Fertil. Dev. 2017, 29, 2073. [Google Scholar] [CrossRef]
- Lipka, A.; Panasiewicz, G.; Majewska, M.; Paukszto, L.; Bieniek-Kobuszewska, M.; Szafranska, B. Identification of Placental Aspartic Proteinase in the Eurasian Beaver (Castor fiber L.). Int. J. Mol. Sci. 2018, 19, 1229. [Google Scholar] [CrossRef] [PubMed]
- Majewska, M.; Lipka, A.; Paukszto, L.; Jastrzebski, J.P.; Myszczynski, K.; Gowkielewicz, M.; Jozwik, M.; Majewski, M.K. Transcriptome profile of the human placenta. Funct. Integr. Genomics 2017, 17, 551–563. [Google Scholar] [CrossRef] [Green Version]
- Majewska, M.; Lipka, A.; Paukszto, L.; Jastrzebski, J.P.; Gowkielewicz, M.; Jozwik, M.; Majewski, M.K. Preliminary RNA-Seq Analysis of Long Non-Coding RNAs Expressed in Human Term Placenta. Int. J. Mol. Sci. 2018, 19, 1894. [Google Scholar] [CrossRef] [PubMed]
- Juiz, N.A.; Torrejón, I.; Burgos, M.; Torres, A.M.F.; Duffy, T.; Cayo, N.M.; Tabasco, A.; Salvo, M.; Longhi, S.A.; Schijman, A.G. Alterations in Placental Gene Expression of Pregnant Women with Chronic Chagas Disease. Am. J. Pathol. 2018, 188, 1345–1353. [Google Scholar] [CrossRef] [PubMed]
- Sitras, V.; Paulssen, R.; Leirvik, J.; Vårtun, Å.; Acharya, G. Placental Gene Expression Profile in Intrauterine Growth Restriction Due to Placental Insufficiency. Reprod. Sci. 2009, 16, 701–711. [Google Scholar] [CrossRef] [PubMed]
- Leftwich, H.K.; Stetson, B.; Sabol, B.; Leung, K.; Hibbard, J.; Wilkins, I. Growth restriction: Identifying fetuses at risk. J. Matern. Neonatal Med. 2018, 31, 1962–1966. [Google Scholar] [CrossRef] [PubMed]
- Kingdom, J.; Huppertz, B.; Seaward, G.; Kaufmann, P. Development of the placental villous tree and its consequences for fetal growth. Eur. J. Obstet. Gynecol. Reprod. Biol. 2000, 92, 35–43. [Google Scholar] [CrossRef]
- Fitzgerald, B.; Levytska, K.; Kingdom, J.; Walker, M.; Baczyk, D.; Keating, S. Villous trophoblast abnormalities in extremely preterm deliveries with elevated second trimester maternal serum hCG or inhibin-A. Placenta 2011, 32, 339–345. [Google Scholar] [CrossRef]
- Dunk, C.E.; Roggensack, A.M.; Cox, B.; Perkins, J.E.; Senius, F.; Keating, S.; Weksberg, R.; Kingdom, J.C.P.; Adamson, S.L. A distinct microvascular endothelial gene expression profile in severe IUGR placentas. Placenta 2012, 33, 285–293. [Google Scholar] [CrossRef]
- Fu, B.; Tian, Z.; Wei, H. TH17 cells in human recurrent pregnancy loss and pre-eclampsia. Cell. Mol. Immunol. 2014, 11, 564–570. [Google Scholar] [CrossRef] [Green Version]
- Piccinni, M.-P.; Lombardelli, L.; Logiodice, F.; Kullolli, O.; Romagnani, S.; Le Bouteiller, P. T helper cell mediated-tolerance towards fetal allograft in successful pregnancy. Clin. Mol. Allergy 2015, 13, 9. [Google Scholar] [CrossRef] [PubMed]
- Granger, J.P.; Alexander, B.T.; Llinas, M.T.; Bennett, W.A.; Khalil, R.A. Pathophysiology of preeclampsia: Linking placental ischemia/hypoxia with microvascular dysfunction. Microcirculation 2002, 9, 147–160. [Google Scholar] [CrossRef]
- LaMarca, B. The role of immune activation in contributing to vascular dysfunction and the pathophysiology of hypertension during preeclampsia. Minerva Ginecol. 2010, 62, 105–120. [Google Scholar] [CrossRef] [PubMed]
- Sankaralingam, S.; Arenas, I.A.; Lalu, M.M.; Davidge, S.T. Preeclampsia: Current understanding of the molecular basis of vascular dysfunction. Expert Rev. Mol. Med. 2006, 8, 1–20. [Google Scholar] [CrossRef]
- Trogstad, L.; Magnus, P.; Stoltenberg, C. Pre-eclampsia: Risk factors and causal models. Best Pract. Res. Clin. Obstet. Gynaecol. 2011, 25, 329–342. [Google Scholar] [CrossRef] [PubMed]
- Ness, R.B.; Sibai, B.M. Shared and disparate components of the pathophysiologies of fetal growth restriction and preeclampsia. Am. J. Obstet. Gynecol. 2006, 195, 40–49. [Google Scholar] [CrossRef]
- Myatt, L. Placental adaptive responses and fetal programming. J. Physiol. 2006, 572, 25–30. [Google Scholar] [CrossRef] [Green Version]
- Faraci, M.; Renda, E.; Monte, S.; Di Prima, F.A.F.; Valenti, O.; De Domenico, R.; Giorgio, E.; Hyseni, E. Fetal growth restriction: Current perspectives. J. Prenat. Med. 2011, 5, 31–33. [Google Scholar] [PubMed]
- Pagani, F.; Baralle, F.E. Genomic variants in exons and introns: Identifying the splicing spoilers. Nat. Rev. Genet. 2004, 5, 389–396. [Google Scholar] [CrossRef] [PubMed]
- Park, E.; Pan, Z.; Zhang, Z.; Lin, L.; Xing, Y. The Expanding Landscape of Alternative Splicing Variation in Human Populations. Am. J. Hum. Genet. 2018, 102, 11–26. [Google Scholar] [CrossRef]
- Mazzucchelli, R.; Durum, S.K. Interleukin-7 receptor expression: Intelligent design. Nat. Rev. Immunol. 2007, 7, 144–154. [Google Scholar] [CrossRef]
- Palmer, M.J.; Mahajan, V.S.; Trajman, L.C.; Irvine, D.J.; Lauffenburger, D.A.; Chen, J. Interleukin-7 Receptor Signaling Network: An Integrated Systems Perspective. Cell. Mol. Immunol. 2008, 5, 79–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, L.; Li, J.; Xu, H.L.; Xu, B.; Tong, X.H.; Kwak-Kim, J.; Liu, Y.S. IL-7/IL-7R signaling pathway might play a role in recurrent pregnancy losses by increasing inflammatory Th17 cells and decreasing Treg cells. Am. J. Reprod. Immunol. 2016, 76, 454–464. [Google Scholar] [CrossRef] [PubMed]
- Inchley, C.S.; Østerholt, H.C.D.; Sonerud, T.; Fjærli, H.O.; Nakstad, B. Downregulation of IL7R, CCR7, and TLR4 in the cord blood of children with respiratory syncytial virus disease. J. Infect. Dis. 2013, 208, 1431–1435. [Google Scholar] [CrossRef] [PubMed]
- Liao, C.Y.; Yu, H.W.; Cheng, C.N.; Chen, J.S.; Lin, C.W.; Chen, P.C.; Shieh, C.C. A novel pathogenic mutation on Interleukin-7 receptor leading to severe combined immunodeficiency identified with newborn screening and whole exome sequencing. J. Microbiol. Immunol. Infect. 2018, S1684-1182(18)30049-5. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.; Yun, S.; Choi, G.H.; Jung, T.W. Fibronectin Type III Domain Containing 4 attenuates hyperlipidemia-induced insulin resistance via suppression of inflammation and ER stress through HO-1 expression in adipocytes. Biochem. Biophys. Res. Commun. 2018, 502, 129–136. [Google Scholar] [CrossRef]
- Goyer, B.; Thériault, M.; Gendron, S.P.; Brunette, I.; Rochette, P.J.; Proulx, S. Extracellular Matrix and Integrin Expression Profiles in Fuchs Endothelial Corneal Dystrophy Cells and Tissue Model. Tissue Eng. Part A 2018, 24, 607–615. [Google Scholar] [CrossRef] [PubMed]
- Bosma, M.; Gerling, M.; Pasto, J.; Georgiadi, A.; Graham, E.; Shilkova, O.; Iwata, Y.; Almer, S.; Söderman, J.; Toftgård, R.; et al. FNDC4 acts as an anti-inflammatory factor on macrophages and improves colitis in mice. Nat. Commun. 2016, 7, 11314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arnett, H.A.; Viney, J.L. Immune modulation by butyrophilins. Nat. Rev. Immunol. 2014, 14, 559–569. [Google Scholar] [CrossRef]
- Zhong, Y.; Johnson, A.J.; Byrd, J.C.; Dubovsky, J.A. Targeting Interleukin-2-Inducible T-cell Kinase (ITK) in T-Cell Related Diseases. Postdoc J. 2014, 2, 1–11. [Google Scholar] [CrossRef]
- Alme, C.; Satwani, P.; Alobeid, B.; Bhagat, G.; Kelly, K.M. Atypical clinical course in pediatric Hodgkin lymphoma: Association with germline mutations in interleukin-2-inducible T-cell kinase. J. Pediatr. Hematol. Oncol. 2015, 37, 507–508. [Google Scholar] [CrossRef] [PubMed]
- Vélez, J.I.; Lopera, F.; Sepulveda-Falla, D.; Patel, H.R.; Johar, A.S.; Chuah, A.; Tobón, C.; Rivera, D.; Villegas, A.; Cai, Y.; et al. APOE*E2 allele delays age of onset in PSEN1 E280A Alzheimer’s disease. Mol. Psychiatry 2016, 21, 916–924. [Google Scholar] [CrossRef] [PubMed]
- Schierding, W.; Antony, J.; Karhunen, V.; Vääräsmäki, M.; Franks, S.; Elliott, P.; Kajantie, E.; Sebert, S.; Blakemore, A.; Horsfield, J.A.; et al. GWAS on prolonged gestation (post-term birth): Analysis of successive Finnish birth cohorts. J. Med. Genet. 2018, 55, 55–63. [Google Scholar] [CrossRef]
- Brown, A.S.; Hooton, J.; Schaefer, C.A.; Zhang, H.; Petkova, E.; Babulas, V.; Perrin, M.; Gorman, J.M.; Susser, E.S. Elevated Maternal Interleukin-8 Levels and Risk of Schizophrenia in Adult Offspring. Am. J. Psychiatry 2004, 161, 889–895. [Google Scholar] [CrossRef] [PubMed]
- Beversdorf, D.Q.; Manning, S.E.; Hillier, A.; Anderson, S.L.; Nordgren, R.E.; Walters, S.E.; Nagaraja, H.N.; Cooley, W.C.; Gaelic, S.E.; Bauman, M.L. Timing of Prenatal Stressors and Autism. J. Autism Dev. Disord. 2005, 35, 471–478. [Google Scholar] [CrossRef] [PubMed]
- Whitton, L.; Apostolova, G.; Rieder, D.; Dechant, G.; Rea, S.; Donohoe, G.; Morris, D.W. Genes regulated by SATB2 during neurodevelopment contribute to schizophrenia and educational attainment. PLoS Genet. 2018, 14, e1007515. [Google Scholar] [CrossRef] [PubMed]
- Lessel, D.; Gehbauer, C.; Bramswig, N.C.; Schluth-Bolard, C.; Venkataramanappa, S.; van Gassen, K.L.I.; Hempel, M.; Haack, T.B.; Baresic, A.; Genetti, C.A.; et al. BCL11B mutations in patients affected by a neurodevelopmental disorder with reduced type 2 innate lymphoid cells. Brain 2018, 141, 2299–2311. [Google Scholar] [CrossRef]
- Punwani, D.; Zhang, Y.; Yu, J.; Cowan, M.J.; Rana, S.; Kwan, A.; Adhikari, A.N.; Lizama, C.O.; Mendelsohn, B.A.; Fahl, S.P.; et al. Multisystem Anomalies in Severe Combined Immunodeficiency with Mutant BCL11B. N. Engl. J. Med. 2016, 375, 2165–2176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, X.; von Kampen, J.; Slaughter, C.A.; DeMartino, G.N. Relative Functions of the α and β Subunits of the Proteasome Activator, PA28. J. Biol. Chem. 1997, 272, 27994–28000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, K.; Mizushima, T.; Saeki, Y. The proteasome: Molecular machinery and pathophysiological roles. Biol. Chem. 2012, 393, 217–234. [Google Scholar] [CrossRef]
- Chen, J.-Y.; Xu, L.; Fang, W.-M.; Han, J.-Y.; Wang, K.; Zhu, K.-S. Identification of PA28β as a potential novel biomarker in human esophageal squamous cell carcinoma. Tumor Biol. 2017, 39, 101042831771978. [Google Scholar] [CrossRef] [PubMed]
- Kawasaki, M.; Sekigawa, I.; Nozawa, K.; Kaneko, H.; Takasaki, Y.; Takamori, K.; Ogawa, H. Changes in the gene expression of peripheral blood mononuclear cells during the menstrual cycle of females is associated with a gender bias in the incidence of systemic lupus erythematosus. Clin. Exp. Rheumatol. 2009, 27, 260–266. [Google Scholar] [PubMed]
- Bing, P.-F.; Xia, W.; Wang, L.; Zhang, Y.-H.; Lei, S.-F.; Deng, F.-Y. Common Marker Genes Identified from Various Sample Types for Systemic Lupus Erythematosus. PLoS ONE 2016, 11, e0156234. [Google Scholar] [CrossRef]
- Bai, Q.; Assou, S.; Haouzi, D.; Ramirez, J.-M.; Monzo, C.; Becker, F.; Gerbal-Chaloin, S.; Hamamah, S.; De Vos, J. Dissecting the First Transcriptional Divergence During Human Embryonic Development. Stem Cell Rev. Rep. 2012, 8, 150–162. [Google Scholar] [CrossRef] [PubMed]
- Abu Taha, A.; Schnittler, H.-J. Dynamics between actin and the VE–cadherin/catenin complex. Cell Adh. Migr. 2014, 8, 125–135. [Google Scholar] [CrossRef]
- Rutledge, N.S.; Weber, E.W.; Winger, R.; Tourtellotte, W.G.; Park, S.H.; Muller, W.A. CD99-like 2 (CD99L2)-deficient mice are defective in the acute inflammatory response. Exp. Mol. Pathol. 2015, 99, 455–459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zamoyska, R.; Basson, A.; Filby, A.; Legname, G.; Lovatt, M.; Seddon, B. The influence of the src-family kinases, Lck and Fyn, on T cell differentiation, survival and activation. Immunol. Rev. 2003, 191, 107–118. [Google Scholar] [CrossRef]
- Gladkikh, A.A.; Potashnikova, D.M.; Tatarskiy, V.; Yastrebova, M.; Khamidullina, A.; Barteneva, N.; Vorobjev, I. Comparison of the mRNA expression profile of B-cell receptor components in normal CD5-high B-lymphocytes and chronic lymphocytic leukemia: A key role of ZAP70. Cancer Med. 2017, 6, 2984–2997. [Google Scholar] [CrossRef]
- Daoud, G.; Rassart, É.; Masse, A.; Lafond, J. Src family kinases play multiple roles in differentiation of trophoblasts from human term placenta. J. Physiol. 2006, 571, 537–553. [Google Scholar] [CrossRef] [Green Version]
- Irtegun, S.; Akcora-Yıldız, D.; Pektanc, G.; Karabulut, C. Deregulation of c-Src tyrosine kinase and its downstream targets in pre-eclamptic placenta. J. Obstet. Gynaecol. Res. 2017, 43, 1278–1284. [Google Scholar] [CrossRef]
- Burton, G.J. Oxygen, the Janus gas; its effects on human placental development and function. J. Anat. 2009, 215, 27–35. [Google Scholar] [CrossRef]
- Fritsche, L.G.; Loenhardt, T.; Janssen, A.; Fisher, S.A.; Rivera, A.; Keilhauer, C.N.; Weber, B.H.F. Age-related macular degeneration is associated with an unstable ARMS2 (LOC387715) mRNA. Nat. Genet. 2008, 40, 892–896. [Google Scholar] [CrossRef]
- Wang, G.; Spencer, K.L.; Scott, W.K.; Whitehead, P.; Court, B.L.; Ayala-Haedo, J.; Mayo, P.; Schwartz, S.G.; Kovach, J.L.; Gallins, P.; et al. Analysis of the indel at the ARMS2 3UTR in age-related macular degeneration. Hum. Genet. 2010, 127, 595–602. [Google Scholar] [CrossRef]
- Sun, L.; Zhou, J.; Wang, K.; Wang, J.; Shang, L.; Zhang, J.; Wu, J.; Cram, D.S. Placental Up-Regulation of Leptin and ARMS2 is Associated with Growth Discordance in Monochorionic Diamniotic Twin Pregnancies. Twin Res. Hum. Genet. 2017, 20, 169–179. [Google Scholar] [CrossRef]
- Zeng, F.; Zhang, M.; Xu, Y.; Xu, H. ARMS2 interference leads to decrease of proinflammatory mediators. Graefe’s Arch. Clin. Exp. Ophthalmol. 2013, 251, 2539–2544. [Google Scholar] [CrossRef]
- Brett, K.E.; Ferraro, Z.M.; Yockell-Lelievre, J.; Gruslin, A.; Adamo, K.B. Maternal–Fetal nutrient transport in pregnancy pathologies: The role of the placenta. Int. J. Mol. Sci. 2014, 15, 16153–16185. [Google Scholar] [CrossRef]
- Huang, X.; Anderle, P.; Hostettler, L.; Baumann, M.U.; Surbek, D.V.; Ontsouka, E.C.; Albrecht, C. Identification of placental nutrient transporters associated with intrauterine growth restriction and pre-eclampsia. BMC Genomics 2018, 19, 173. [Google Scholar] [CrossRef]
- Day, P.E.L.; Cleal, J.K.; Lofthouse, E.M.; Goss, V.; Koster, G.; Postle, A.; Jackson, J.M.; Hanson, M.A.; Jackson, A.A.; Lewis, R.M. Partitioning of glutamine synthesised by the isolated perfused human placenta between the maternal and fetal circulations q. Placenta 2013, 34, 1223–1231. [Google Scholar] [CrossRef]
- Rodríguez, A.; Ortega, A.; Berumen, L.C.; García-Alcocer, M.G.; Giménez, C.; Zafra, F. Expression of the System N transporter (SNAT5/SN2) during development indicates its plausible role in glutamatergic neurotransmission. Neurochem. Int. 2014, 73, 166–171. [Google Scholar] [CrossRef]
- Bröer, S. The SLC38 family of sodium-amino acid co-transporters. Pflug. Arch. Eur. J. Physiol. 2014, 466, 155–172. [Google Scholar] [CrossRef]
- Mackenzie, B.; Erickson, J.D. Sodium-coupled neutral amino acid (System N/A) transporters of the SLC38 gene family. Pflug. Arch. Eur. J. Physiol. 2004, 447, 784–795. [Google Scholar] [CrossRef]
- Hunt, J.S.; Rasmussen, C.A. The tumor necrosis factor gene family in pregnancy. Placenta 1998, 19, 139–147. [Google Scholar] [CrossRef]
- Browning, J.L.; Ngam-ek, A.; Lawton, P.; DeMarinis, J.; Tizard, R.; Chow, E.P.; Hession, C.; O’Brine-Greco, B.; Foley, S.F.; Ware, C.F. Lymphotoxin β, a novel member of the TNF family that forms a heteromeric complex with lymphotoxin on the cell surface. Cell 1993, 72, 847–856. [Google Scholar] [CrossRef]
- Snyder, S.K.; Wessells, J.L.; Waterhouse, R.M.; Dveksler, G.S.; Wessner, D.H.; Wahl, L.M.; Zimmermann, W. Pregnancy-Specific Glycoproteins Function as Immunomodulators by Inducing Secretion of IL-10, IL-6 and TGF-beta1 by Human Monocytes. Am. J. Reprod. Immunol. 2001, 45, 205–216. [Google Scholar] [CrossRef]
- Gordon, Y.B.; Jeffrey, D.; Grudzinskas, J.G.; Chard, T.; Letchworth, A.T. Concentrations of pregnancy-specific β1-glycoprotein in maternal blood in normal pregnancy and in intrauterine growth retardation. Lancet 1977, 309, 331–333. [Google Scholar] [CrossRef]
- Zhao, L.; Triche, E.W.; Walsh, K.M.; Bracken, M.B.; Saftlas, A.F.; Hoh, J.; Dewan, A.T. Genome-wide association study identifies a maternal copy-number deletion in PSG11 enriched among preeclampsia patients. BMC Pregnancy Childbirth 2012, 12, 1–10. [Google Scholar] [CrossRef]
- Kadamb, R.; Mittal, S.; Bansal, N.; Batra, H.; Saluja, D. Sin3: Insight into its transcription regulatory functions. Eur. J. Cell Biol. 2013, 237–246. [Google Scholar] [CrossRef]
- Sukhikh, G.T.; Bourmenskaya, O.V.; Smolnikova, V.Y.; Krasnoschoka, O.E.; Trofimov, D.Y.; Donnikov, A.E.; Kalinina, E.A. Protamine and fertilin mRNA: Potential biomarkers of assisted reproductive technology outcomes. Bull. Exp. Biol. Med. 2012, 153, 513–515. [Google Scholar] [CrossRef]
- Depa-Martynów, M.; Kempisty, B.; Lianeri, L.; Pawel, P.; Jagodziñski, P.J. Association between fertilin β, protamines 1 and 2 and spermatid-specific linker histone H1-like protein mRNA levels, fertilization ability of human spermatozoa, and quality of preimplantation embryos. Folia Histochem. Cytobiol. 2007, 45, 79–85. [Google Scholar]
- Cascella, R.; Strafella, C.; Caputo, V.; Errichiello, V.; Zampatti, S.; Milano, F.; Potenza, S.; Mauriello, S.; Novelli, G.; Ricci, F.; et al. Towards the application of precision medicine in Age-Related Macular Degeneration. Prog. Retin. Eye Res. 2018, 63, 132–146. [Google Scholar] [CrossRef]
- Sharma, S.; Godbole, G.; Modi, D. Decidual Control of Trophoblast Invasion. Am. J. Reprod. Immunol. 2016, 75, 341–350. [Google Scholar] [CrossRef] [Green Version]
- Merviel, P.; Challier, J.-C.; Carbillon, L.; Foidart, J.-M.; Uzan, S. The Role of Integrins in Human Embryo Implantation. Fetal Diagn. Ther. 2001, 16, 364–371. [Google Scholar] [CrossRef]
- Su, E.J. Role of the fetoplacental endothelium in fetal growth restriction with abnormal umbilical artery Doppler velocimetry. Am. J. Obstet. Gynecol. 2015, 213, S123–S130. [Google Scholar] [CrossRef]
- Morley, L.C.; Shi, J.; Gaunt, H.J.; Hyman, A.J.; Webster, P.J.; Williams, C.; Forbes, K.; Walker, J.J.; Simpson, N.A.B.; Beech, D.J. Piezo1 channels are mechanosensors in human fetoplacental endothelial cells. MHR Basic Sci. Reprod. Med. 2018, 24, 510–520. [Google Scholar] [CrossRef]
- Liu, J.; Li, D.; Cao, B.; Li, Y.; Herva, R.; Piao, Y.; Wang, Y. Expression and Localization of SWAP-70 in Human Fetomaternal Interface and Placenta During Tubal Pregnancy and Normal Placentation. J. Histochem. Cytochem. 2007, 55, 701–708. [Google Scholar] [CrossRef] [Green Version]
- Mansilla, M.; Wang, Y.; Hyett, J.; da Silva Costa, F.; Nie, G. Serum podocalyxin for early detection of preeclampsia at 11–13 weeks of gestation. Placenta 2018, 71, 13–15. [Google Scholar] [CrossRef]
- Chen, Q.; Wang, Y.; Li, Y.; Zhao, M.; Nie, G. Serum podocalyxin is significantly increased in early-onset preeclampsia and may represent a novel marker of maternal endothelial cell dysfunction. J. Hypertens. 2017, 35, 2287–2294. [Google Scholar] [CrossRef]
- Di Bartolomeo, A.; Chauleur, C.; Gris, J.C.; Chapelle, C.; Noblot, E.; Laporte, S.; Raia-Barjat, T. Tissue factor pathway inhibitor for prediction of placenta-mediated adverse pregnancy outcomes in high-risk women: AngioPred study. PLoS ONE 2017, 12, e0173596. [Google Scholar] [CrossRef]
- Redline, R.W. Placental Pathology: A Systematic Approach with Clinical Correlations. Placenta 2008, 29, 86–91. [Google Scholar] [CrossRef]
- Landriscina, M.; Schinzari, G.; Di Leonardo, G.; Quirino, M.; Cassano, A.; D’Argento, E.; Lauriola, L.; Scerrati, M.; Prudovsky, I.; Barone, C. S100A13, a new marker of angiogenesis in human astrocytic gliomas. J. Neurooncol. 2006, 80, 251–259. [Google Scholar] [CrossRef]
- Cui, H.; Wang, Y.; Huang, H.; Yu, W.; Bai, M.; Zhang, L.; Bryan, B.A.; Wang, Y.; Luo, J.; Li, D.; et al. GPR126 protein regulates developmental and pathological angiogenesis through modulation of VEGFR2 receptor signaling. J. Biol. Chem. 2014, 289, 34871–34885. [Google Scholar] [CrossRef]
- Lwaleed, B.A.; Bass, P.S. Tissue factor pathway inhibitor: Structure, biology and involvement in disease. J. Pathol. 2006, 208, 327–339. [Google Scholar] [CrossRef]
- Edstrom, C.S.; Calhoun, D.A.; Christensen, R.D. Expression of tissue factor pathway inhibitor in human fetal and placental tissues. Early Hum. Dev. 2000, 59, 77–84. [Google Scholar] [CrossRef]
- Aharon, A.; Brenner, B.; Katz, T.; Miyagi, Y.; Lanir, N. Tissue factor and tissue factor pathway inhibitor levels in trophoblast cells: Implications for placental hemostasis. Thromb. Haemost. 2004, 92, 776–786. [Google Scholar] [CrossRef]
- Ittel, A.; Bretelle, F.; Gris, J.C.; Chau, C.; Sébahoun, G.; Boubli, L.; Arnoux, D. Increased risk of gestational vascular complications in women with low free tissue factor pathway inhibitor plasma levels out of pregnancy. Thromb. Haemost. 2011, 105, 66–71. [Google Scholar] [CrossRef]
- Wong, G.W.; Krawczyk, S.A.; Kitidis-Mitrokostas, C.; Revett, T.; Gimeno, R.; Lodish, H.F. Molecular, biochemical and functional characterizations of C1q/TNF family members: Adipose-tissue-selective expression patterns, regulation by PPAR-γ agonist, cysteine-mediated oligomerizations, combinatorial associations and metabolic functions. Biochem. J. 2008, 416, 161–177. [Google Scholar] [CrossRef]
- Liu, Z.H.; Li, C.; Chen, J.W.; Shen, Y.; Gao, J.; Shen, W.F.; Zhang, R.Y.; Wang, X.Q.; Lu, L. C1q/TNF-related protein 1 promotes endothelial barrier dysfunction under disturbed flow. Biochem. Biophys. Res. Commun. 2017, 490, 580–586. [Google Scholar] [CrossRef]
- Kanemura, N.; Shibata, R.; Ohashi, K.; Ogawa, H.; Hiramatsu-Ito, M.; Enomoto, T.; Yuasa, D.; Ito, M.; Hayakawa, S.; Otaka, N.; et al. C1q/TNF-related protein 1 prevents neointimal formation after arterial injury. Atherosclerosis 2017, 257, 138–145. [Google Scholar] [CrossRef]
- Nicholson, A.M.; Finch, N.A.; Almeida, M.; Perkerson, R.B.; van Blitterswijk, M.; Wojtas, A.; Cenik, B.; Rotondo, S.; Inskeep, V.; Almasy, L.; et al. Prosaposin is a regulator of progranulin levels and oligomerization. Nat. Commun. 2016, 7, 11992. [Google Scholar] [CrossRef] [Green Version]
- Stubert, J.; Schattenberg, F.; Richter, D.-U.; Dieterich, M.; Briese, V. Trophoblastic progranulin expression is upregulated in cases of fetal growth restriction and preeclampsia. J. Perinat. Med. 2012, 40, 475–481. [Google Scholar] [CrossRef]
- Pyle, A.L.; Li, B.; Maupin, A.B.; Guzman, R.J.; Crimmins, D.L.; Olson, S.; Atkinson, J.B.; Young, P.P. Biomechanical stress induces novel arterial intima-enriched genes: Implications for vascular adaptation to stress. Cardiovasc. Pathol. 2010, 19, e13–e20. [Google Scholar] [CrossRef]
- Wakasugi, K.; Slike, B.M.; Hood, J.; Otani, A.; Ewalt, K.L.; Friedlander, M.; Cheresh, D.A.; Schimmel, P. A human aminoacyl-tRNA synthetase as a regulator of angiogenesis. Proc. Natl. Acad. Sci. USA 2002, 99, 173–177. [Google Scholar] [CrossRef] [Green Version]
- Liu, B.; Xu, Y.; Voss, C.; Qiu, F.H.; Zhao, M.Z.; Liu, Y.D.; Nie, J.; Wang, Z.L. Altered Protein Expression in Gestational Diabetes Mellitus Placentas Provides Insight into Insulin Resistance and Coagulation/Fibrinolysis Pathways. PLoS ONE 2012, 7, e44701. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [Green Version]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.-C.; Mendell, J.T.; Salzberg, S.L. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef] [Green Version]
- Pertea, M.; Kim, D.; Pertea, G.M.; Leek, J.T.; Salzberg, S.L. Transcript-level expression analysis of RNA-seq experiments with HISAT, StringTie and Ballgown. Nat. Protoc. 2016, 11, 1650–1667. [Google Scholar] [CrossRef]
- Anders, S.; Huber, W. Differential expression analysis for sequence count data. Genome Biol. 2010, 11, R106. [Google Scholar] [CrossRef]
- Robinson, M.D.; Mccarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinforma. Appl. NOTE 2010, 26, 139–140. [Google Scholar] [CrossRef]
- Warnes, G.R.; Bolker, B.; Bonebakker, L.; Gentleman, R.; Liaw, W.H.A.; Lumley, T.; Maechler, M.; Magnusson, A.; Moeller, S.; Schwartz, M.V.B. gplots: Various R programming tools for plotting data. R Packag. Vers. 2009, 2, 1. [Google Scholar]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Reimand, J.; Arak, T.; Adler, P.; Kolberg, L.; Reisberg, S.; Peterson, H.; Vilo, J. g:Profiler-a web server for functional interpretation of gene lists (2016 update). Nucleic Acids Res. 2016, 44, W83–W89. [Google Scholar] [CrossRef]
- Alamancos, G.P.; Pagès, A.; Trincado, J.L.; Bellora, N.; Eyras, E. SUPPA: A super-fast pipeline for alternative splicing analysis from RNA-Seq. bioRxiv 2014, 008763. [Google Scholar] [CrossRef]
- Shen, S.; Park, J.W.; Lu, Z.-X.; Lin, L.; Henry, M.D.; Wu, Y.N.; Zhou, Q.; Xing, Y. rMATS: Robust and flexible detection of differential alternative splicing from replicate RNA-Seq data. Proc. Natl. Acad. Sci. USA 2014, 111, E5593–E5601. [Google Scholar] [CrossRef] [Green Version]
- Patro, R.; Duggal, G.; Love, M.I.; Irizarry, R.A.; Kingsford, C. Salmon provides fast and bias-aware quantification of transcript expression. Nat. Methods 2017, 14, 417–419. [Google Scholar] [CrossRef]
- Quinlan, A.R.; Hall, I.M. BEDTools: A flexible suite of utilities for comparing genomic features. Bioinformatics 2010, 26, 841–842. [Google Scholar] [CrossRef]
- Veiga, D. maser: Mapping Alternative Splicing Events to pRoteins; R Package Version 1.0.0. 2018. Available online: https://github.com/DiogoVeiga/maser (accessed on 26 March 2019).
- Wang, J.; Pan, Y.; Shen, S.; Lin, L.; Xing, Y. rMATS-DVR: rMATS discovery of differential variants in RNA. Bioinformatics 2017, 33, 2216–2217. [Google Scholar] [CrossRef]
- McKenna, A.; Hanna, M.; Banks, E.; Sivachenko, A.; Cibulskis, K.; Kernytsky, A.; Garimella, K.; Altshuler, D.; Gabriel, S.; Daly, M.; et al. The genome analysis toolkit: A MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res. 2010, 20, 1297–1303. [Google Scholar] [CrossRef]
- Wang, X.; Lu, P.; Luo, Z. GMATo: A novel tool for the identification and analysis of microsatellites in large genomes. Bioinformation 2013, 9, 541–544. [Google Scholar] [CrossRef]
- Kent, W.J. BLAT—The BLAST-Like Alignment Tool. Genome Res. 2002, 12, 656–664. [Google Scholar] [CrossRef]
- McLaren, W.; Gil, L.; Hunt, S.E.; Riat, H.S.; Ritchie, G.R.S.; Thormann, A.; Flicek, P.; Cunningham, F. The Ensembl Variant Effect Predictor. Genome Biol. 2016, 17, 122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cingolani, P.; Platts, A.; Wang, L.L.; Coon, M.; Nguyen, T.; Wang, L.; Land, S.J.; Lu, X.; Ruden, D.M. A program for annotating and predicting the effects of single nucleotide polymorphisms, SnpEff: SNPs in the genome of Drosophila melanogaster strain w1118; iso-2; iso-3. Fly (Austin) 2012, 6, 80–92. [Google Scholar] [CrossRef] [PubMed]
- Krzywinski, M.; Schein, J.; Birol, I.; Connors, J.; Gascoyne, R.; Horsman, D.; Jones, S.J.; Marra, M.A. Circos: An Information Aesthetic for Comparative Genomics. Genome Res. 2009, 19, 1639–1645. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CTR1 | CTR2 | CTR3 | CTR4 | CTR5 | IUGR1 | IUGR2 | IUGR3 | IUGR4 | IUGR5 | |
|---|---|---|---|---|---|---|---|---|---|---|
| Fetal sex | female | female | female | male | male | female | female | male | male | male |
| Row reads | 64,041,868 | 76,079,178 | 55,220,598 | 73,481,438 | 57,398,092 | 43,095,034 | 56,174,786 | 36,854,904 | 42,138,704 | 41,532,984 |
| Trimmed reads | 57,087,670 | 66,388,446 | 47,395,044 | 64,759,848 | 49,765,752 | 37,894,430 | 49,348,796 | 30,969,750 | 36,987,266 | 36,422,794 |
| Mapped | 54,927,730 | 64,035,634 | 45,653,626 | 62,541,870 | 48,246,002 | 36,523,434 | 47,657,008 | 29,887,328 | 35,801,520 | 35,037,148 |
| Uniquely mapped | 50,834,558 | 53,206,552 | 41,489,480 | 60,190,958 | 44,263,500 | 33,625,168 | 42,868,410 | 28,014,300 | 32,690,420 | 32,541,876 |
| Multi-mapped | 4,082,300 | 10,817,500 | 4,156,886 | 2,342,936 | 3,974,602 | 2,891,872 | 4,780,564 | 1,868,814 | 3,104,808 | 2,488,174 |
| Too many loci | 10,872 | 11,582 | 7260 | 7976 | 7900 | 6394 | 8034 | 4214 | 6292 | 7098 |
| Gene_ID | BaseMean | log2FC | Padj | Gene Symbol |
|---|---|---|---|---|
| XLOC_048584 | 7.49193 | −2.805 | 1.05 × 10−2 | THEMIS |
| XLOC_032992 | 34.69158 | −2.784 | 2.74 × 10−2 | PTPRN |
| XLOC_028055 | 8.40907 | −2.603 | 6.26 × 10-3 | ADGRE4P |
| XLOC_050480 | 45.34250 | −2.401 | 6.47 × 10−4 | AC245427.9 |
| XLOC_031488 | 63.22940 | −2.386 | 9.55 × 10-3 | FNDC4 |
| XLOC_015723 | 52.46403 | −2.375 | 1.24 × 10−2 | TRAC |
| XLOC_050482 | 62.93612 | −2.313 | 4.71 × 10-3 | TRBC2 |
| XLOC_034173 | 7.15533 | −2.304 | 1.93 × 10−2 | SIRPG |
| XLOC_058872 | 62.88279 | −2.277 | 5.00 × 10−3 | SLC38A5 |
| XLOC_035229 | 129.54309 | −2.191 | 1.99 × 10−2 | B3GALT5 |
| XLOC_030101 | 84.31897 | −2.137 | 1.05 × 10−2 | ZAP70 |
| XLOC_000516 | 38.22445 | −2.130 | 1.02 × 10−2 | LCK |
| XLOC_017361 | 19.31762 | −2.093 | 3.33 × 10−4 | BCL11B |
| XLOC_013236 | 25.54894 | −2.078 | 2.13 × 10−2 | TESPA1 |
| XLOC_043316 | 143.34412 | −2.072 | 3.12 × 10−3 | IL7R |
| XLOC_044222 | 40.17459 | −2.033 | 1.00 × 10−2 | ITK |
| XLOC_047901 | 57.73795 | −1.817 | 2.56 × 10−3 | LTB |
| XLOC_027402 | 22.55203 | −1.804 | 1.24 × 10−2 | PINLYP |
| XLOC_039524 | 248.13131 | 1.392 | 4.71 × 10−3 | ASTE1 |
| XLOC_048872 | 4.95620 | 2.326 | 2.13 × 10−2 | AL023775.1 |
| XLOC_044458 | 371.82484 | 2.429 | 6.17 × 10−3 | BTNL9 |
| XLOC_037183 | 36.60827 | 2.447 | 5.00 × 10−3 | AC087857.1 |
| XLOC_004057 | 13.87959 | 2.524 | 2.36 × 10−2 | TCHHL1 |
| XLOC_029611 | 4.74119 | 2.629 | 1.57 × 10−2 | NA |
| XLOC_053736 | 5.04141 | 2.647 | 4.06 × 10−2 | ADAM2 |
| XLOC_007768 | 6.30889 | 2.937 | 1.34 × 10−2 | NA |
| XLOC_051256 | 14.10328 | 3.087 | 5.00 × 10−3 | PHBP5 |
| XLOC_006484 | 268.76888 | 3.304 | 9.15 × 10−4 | ARMS2 |
| Term ID | Term Name | Gene Symbol |
|---|---|---|
| GO:0050851 | antigen receptor-mediated signaling pathway | ITK, ZAP70, TESPA1, THEMIS, LCK |
| GO:0050852 | T cell receptor signaling pathway | ITK, ZAP70, TESPA1, THEMIS, LCK |
| GO:0007159 | leukocyte cell-cell adhesion | SIRPG, ZAP70, TESPA1, IL7R, LCK |
| GO:1903037 | regulation of leukocyte cell-cell adhesion | SIRPG, ZAP70, TESPA1, IL7R, LCK |
| GO:0045321 | leukocyte activation | SIRPG, ITK, ZAP70, BCL11B, TESPA1, IL7R, THEMIS, LCK |
| GO:0046649 | lymphocyte activation | SIRPG, ITK, ZAP70, BCL11B, TESPA1, IL7R, THEMIS, LCK |
| GO:0042110 | T cell activation | SIRPG, ITK, ZAP70, BCL11B, TESPA1, IL7R, THEMIS, LCK |
| GO:0002520 | immune system development | ITK, ZAP70, BCL11B, TESPA1, IL7R, THEMIS, LCK, LTB |
| GO:0048534 | hematopoietic or lymphoid organ development | ITK, ZAP70, BCL11B, TESPA1, IL7R, THEMIS, LCK, LTB |
| GO:0030097 | hemopoiesis | ITK, ZAP70, BCL11B, TESPA1, IL7R, THEMIS, LCK, |
| GO:0022409 | positive regulation of cell-cell adhesion | SIRPG, ZAP70, TESPA1, IL7R, LCK |
| GO:1903039 | positive regulation of leukocyte cell-cell adhesion | SIRPG, ZAP70, TESPA1, IL7R, LCK |
| GO:0050863 | regulation of T cell activation | SIRPG, ZAP70, TESPA1, IL7R, LCK |
| GO:0002521 | leukocyte differentiation | ITK, ZAP70, BCL11B, TESPA1, IL7R, THEMIS, LCK |
| GO:0030098 | lymphocyte differentiation | ITK, ZAP70, BCL11B, TESPA1, IL7R, THEMIS, LCK |
| GO:0030217 | T cell differentiation | ITK, ZAP70, BCL11B, TESPA1, IL7R, THEMIS, LCK |
| GO:0045058 | T cell selection | ZAP70, BCL11B, THEMIS |
| GO:0043368 | positive T cell selection | ZAP70, BCL11B, THEMIS |
| GO:0033077 | T cell differentiation in thymus | ZAP70, BCL11B, TESPA1, IL7R |
| GO:0051251 | positive regulation of lymphocyte activation | SIRPG, ZAP70, TESPA1, IL7R, LCK |
| GO:0050870 | positive regulation of T cell activation | SIRPG, ZAP70, TESPA1, IL7R, LCK |
| GO:0004715 | non-membrane spanning protein tyrosine kinase activity | ITK, ZAP70, LCK |
| KEGG:04064 | NF-kappa B signaling pathway | ZAP70, LCK, LTB |
| KEGG:04660 | T cell receptor signaling pathway | ITK, ZAP70, LCK |
| KEGG:05340 | Primary immunodeficiency | ZAP70, IL7R, LCK |
| Gene Symbol | AS Type | ΔPSI | p-value | AS Genomic Coordinates |
|---|---|---|---|---|
| TFPI | AF | 0.35943 | 0 | 2:187503770-187513616:187513641:187503770-187554200:187554492:- |
| TFPI | AF | 0.33763 | 1.349 × 10−2 | 2:187503770-187513616:187513653:187503770-187554200:187554492:- |
| TFPI | AF | −0.23995 | 3.853 × 10−2 | 2:187503770-187513616:187519860:187503770-187554200:187554438:- |
| WARS | AF | 0.69358 | 4.496 × 10−2 | 14:100369258-100375283:100375473:100369258-100376260:100376308:- |
| PSG6 | AF | −0.20069 | 2.922 × 10−2 | 19:42910858-42916125:42916398:42910858-42917729:42917887:- |
| C1QTNF1 | AF | −0.31195 | 3.996 × 10−2 | 17:79022934:79023088-79043955:79025663:79025935-79043955:+ |
| C1QTNF1 | AF | −0.21092 | 4.396 × 10−2 | 17:79024070:79024494-79043955:79025663:79025935-79043955:+ |
| C1QTNF1 | AF | −0.33645 | 4.396 × 10−2 | 17:79024261:79024494-79046555:79043593:79044123-79046555:+ |
| S100A13 | AF | −0.22921 | 4.795 × 10−2 | 1:153626533-153626951:153627048:153626533-153628121:153628180:- |
| S100A13 | AF | −0.20534 | 4.795 × 10−2 | 1:153626533-153627139:153627268:153626533-153628121:153628180:- |
| PSG6 | AL | −0.20069 | 2.922 × 10−2 | 19:42906636:42907176-42910580:42907725:42907854-42910580:- |
| TFPI | MX | −0.21823 | 3.853 × 10−2 | 2:187503770-187513616:187513646-187554200:187503770-187520542:187520613-187554200:- |
| TFPI | SE | 0.12683 | 3.853 × 10−2 | 2:187503770-187529364:187529485-187554200:- |
| EVI5 | SE | −0.30817 | 4.795 × 10−2 | 1:92636336-92663420:92663452-92665939:- |
| PSME2 | SE | −0.28553 | 4.895 × 10−2 | 14:24145772-24146208:24146240-24146534:- |
| ADGRG6 | SE | −0.28155 | 3.497 × 10−2 | 6:142438364-142440908:142440953-142443337:+ |
| SIN3B | SE | 0.09338 | 4.595 × 10−2 | 19:16862559-16862884:16862979-16863680:+ |
| SCEL | SE | 0.35527 | 4.412 × 10−2 | 13:77591460-77593514:77593573-77597545:+ |
| C1QTNF1 | SE | 0.23831 | 4.396 × 10−2 | 17:79024494-79043955:79044123-79046555:+ |
| PSAP | SE | −0.09818 | 2.597 × 10−2 | 10:71822007-71823888:71823896-71825837:- |
| SNV Localization | Number of SNV Annotations | Percentage Ratio |
|---|---|---|
| DOWNSTREAM | 3179 | 31.38% |
| EXON | 2386 | 23.55% |
| INTERGENIC | 10 | 0.10% |
| INTRON | 1528 | 15.08% |
| SPLICE_SITE_ACCEPTOR | 1 | 0.01% |
| SPLICE_SITE_REGION | 54 | 0.53% |
| UPSTREAM | 1369 | 13.51% |
| UTR_3_PRIME | 1260 | 12.44% |
| UTR_5_PRIME | 345 | 3.41% |
| Chr | Site | Quality | FDR | Fraction Difference | Gene Name | Strand | Location | Subst. |
|---|---|---|---|---|---|---|---|---|
| 19 | 45394416 | 357.14 | 9.99 × 10−5 | 0.122 | PPP1R13L | - | Intron | A>G |
| 2 | 37100515 | 373.16 | 1.06 × 10−5 | 0.106 | EIF2AK2 | - | 3UTR | A>G |
| 2 | 218237842 | 275.35 | 7.53 × 10−4 | 0.178 | ARPC2 | + | Intron | A>G |
| X | 150767753 | 438.58 | 2.39 × 10−4 | 0.047 | CD99L2 | - | 3UTR | A>G |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Majewska, M.; Lipka, A.; Paukszto, L.; Jastrzebski, J.P.; Szeszko, K.; Gowkielewicz, M.; Lepiarczyk, E.; Jozwik, M.; Majewski, M.K. Placenta Transcriptome Profiling in Intrauterine Growth Restriction (IUGR). Int. J. Mol. Sci. 2019, 20, 1510. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20061510
Majewska M, Lipka A, Paukszto L, Jastrzebski JP, Szeszko K, Gowkielewicz M, Lepiarczyk E, Jozwik M, Majewski MK. Placenta Transcriptome Profiling in Intrauterine Growth Restriction (IUGR). International Journal of Molecular Sciences. 2019; 20(6):1510. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20061510
Chicago/Turabian StyleMajewska, Marta, Aleksandra Lipka, Lukasz Paukszto, Jan Pawel Jastrzebski, Karol Szeszko, Marek Gowkielewicz, Ewa Lepiarczyk, Marcin Jozwik, and Mariusz Krzysztof Majewski. 2019. "Placenta Transcriptome Profiling in Intrauterine Growth Restriction (IUGR)" International Journal of Molecular Sciences 20, no. 6: 1510. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20061510