Wnt/β-Catenin Pathway Is Involved in Cadmium-Induced Inhibition of Osteoblast Differentiation of Bone Marrow Mesenchymal Stem Cells

Abstract
:1. Introduction
2. Results
2.1. Purity and Potential to Differentiate of BMMSCs
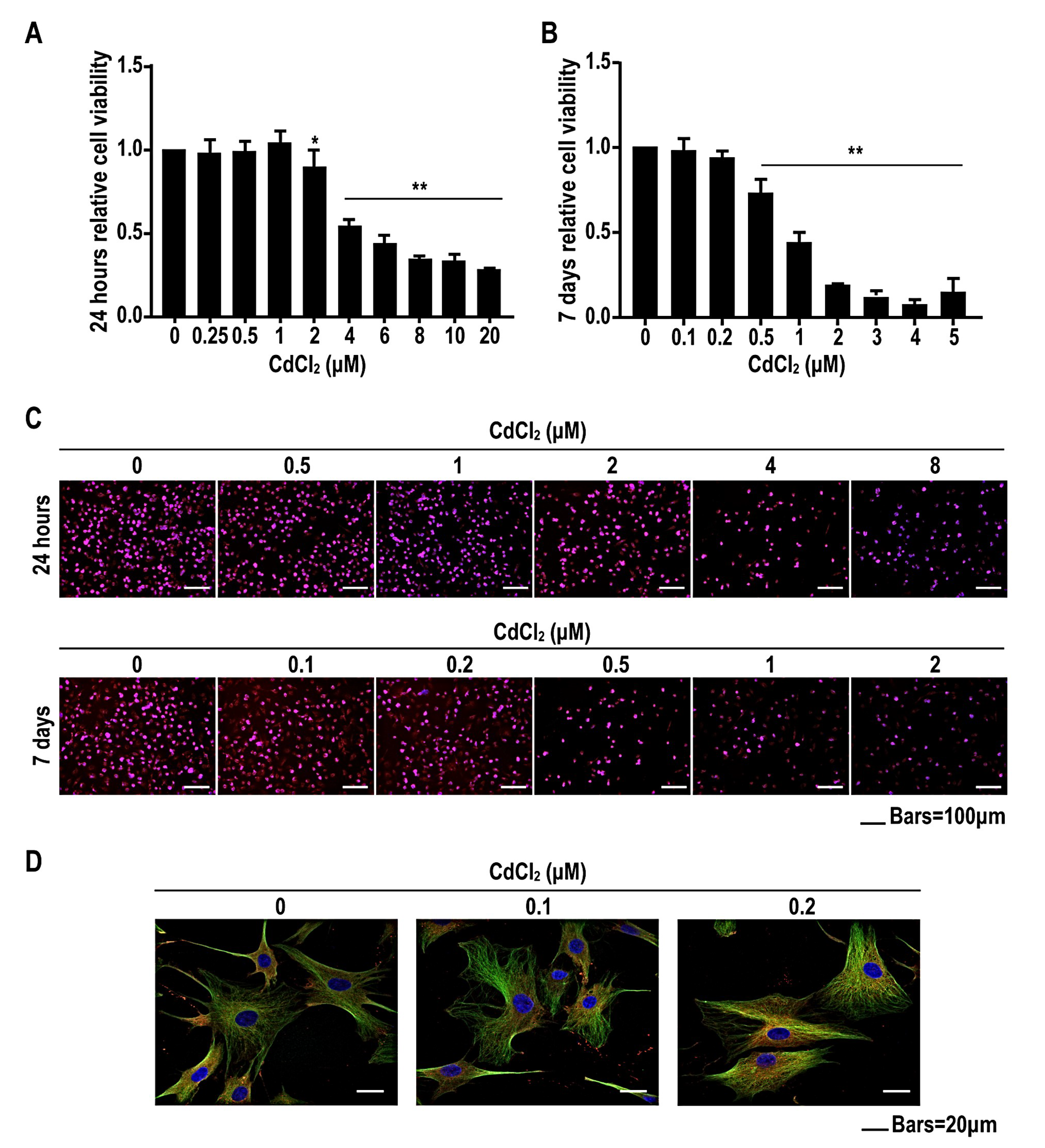
2.2. Low Concentrations of Cadmium Have No Effect on Cell Viability, but High Concentrations of Cadmium Reduce Cell Viability of BMMSCs in a Concentration-Dependent Manner
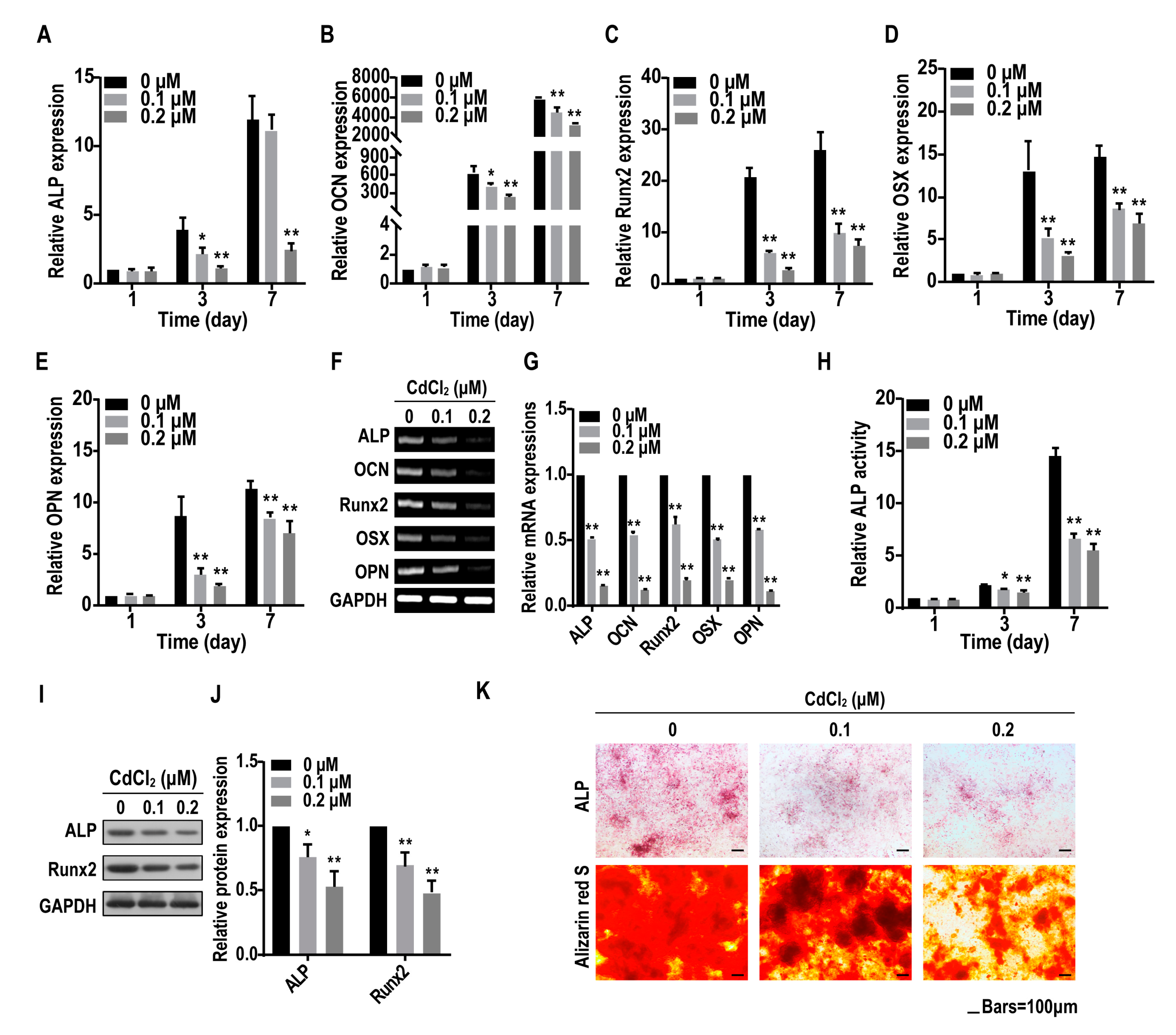
2.3. Cadmium Suppresses the Osteogenesis of BMMSCs
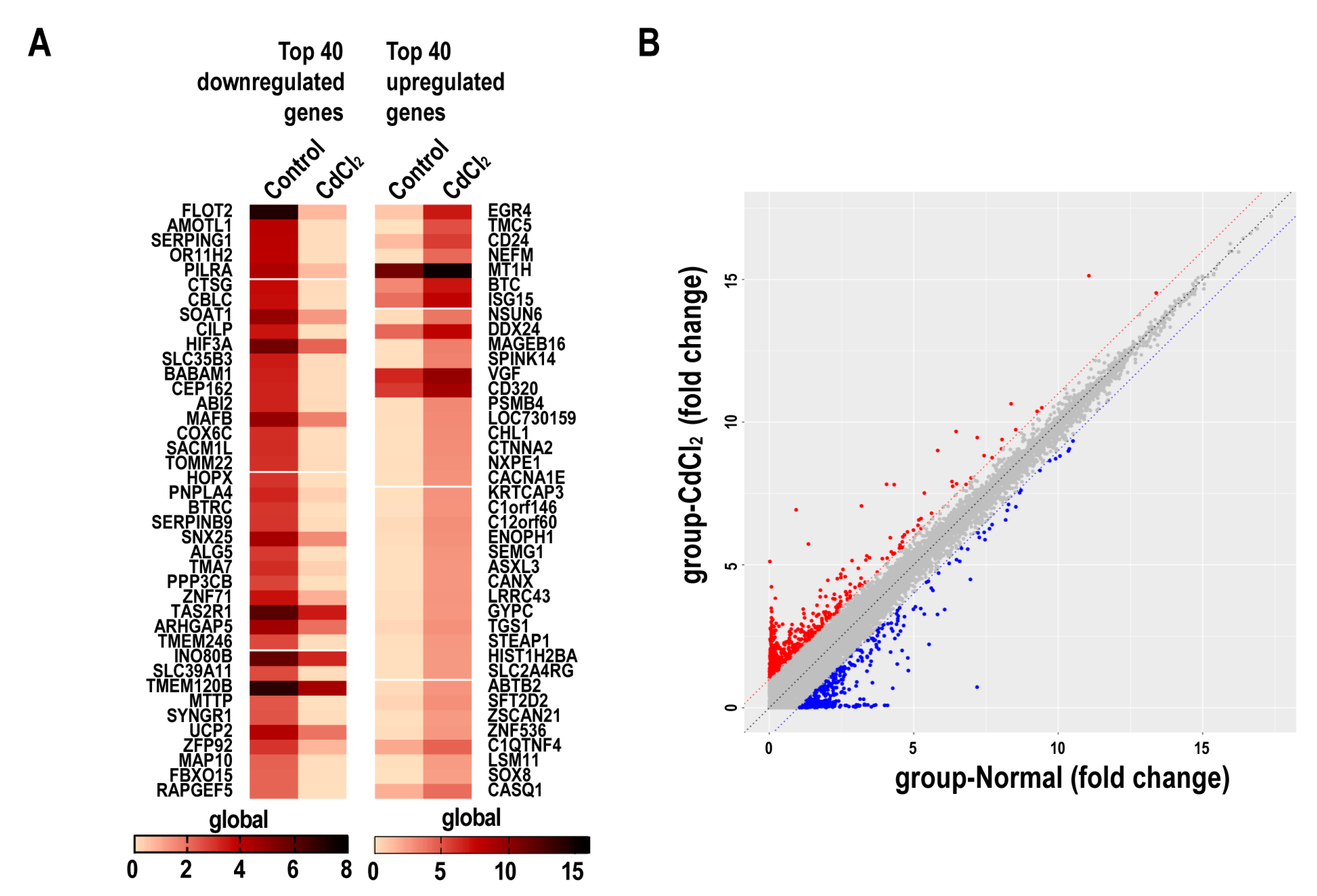
2.4. The Effects of Cadmium on mRNA Expression Profiles in BMMSCs
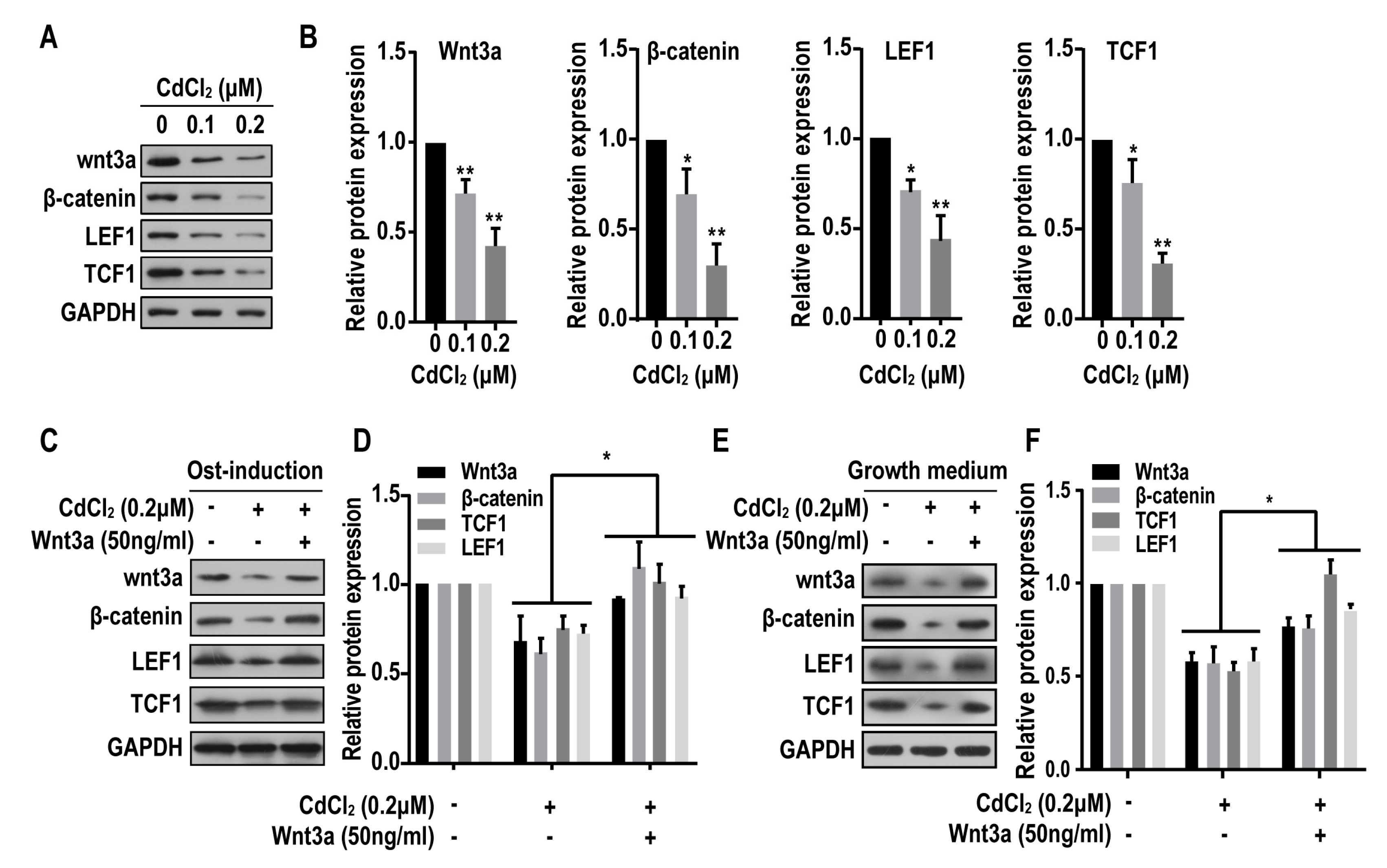
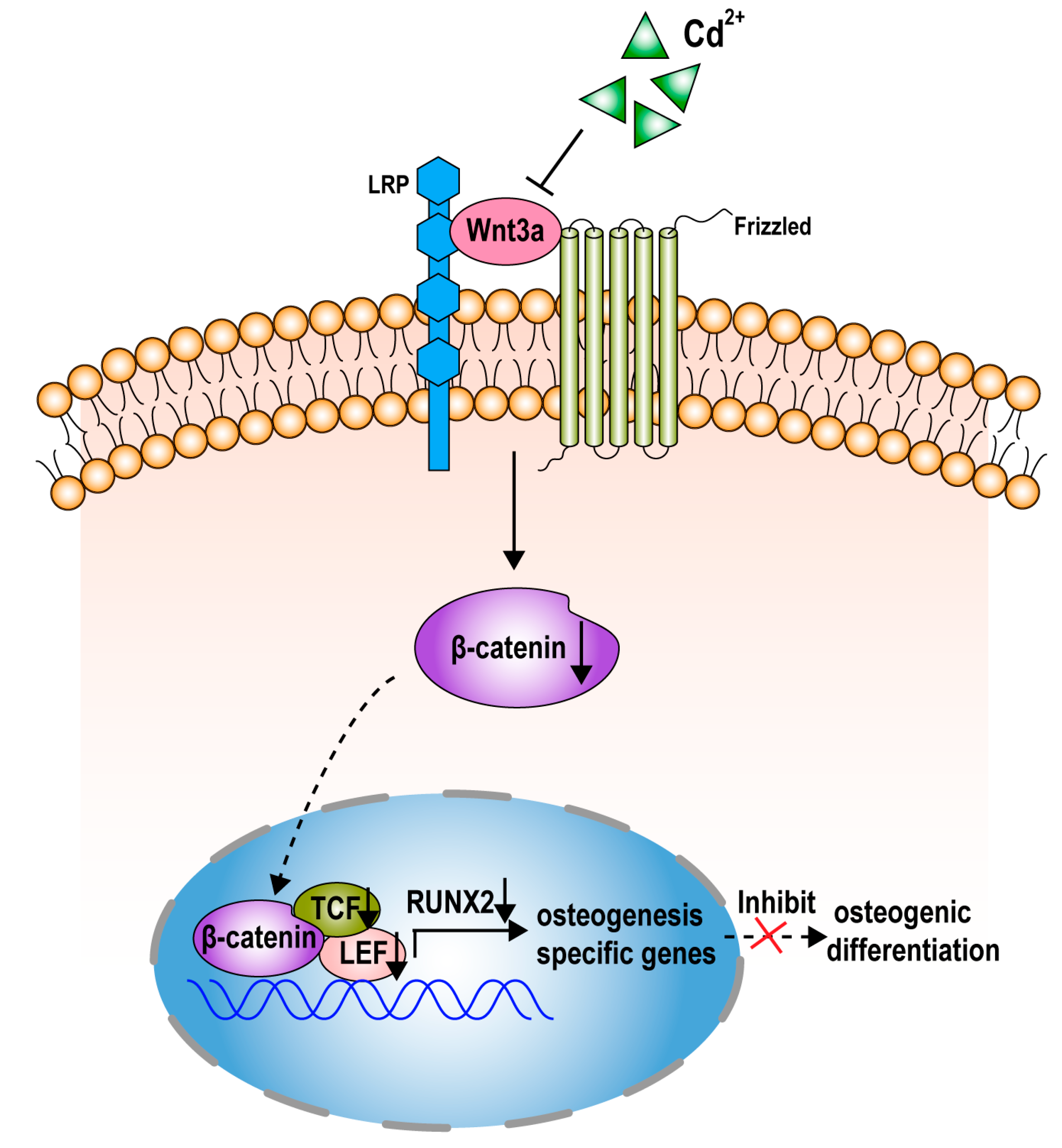
2.5. Cadmium Inhibits the Wnt/β-Catenin Pathway in BMMSCs
2.6. Wnt/β-Catenin Pathway is Involved in the Suppression of Osteogenesis Induced by Cadmuim in BMMSCs
3. Discussion
4. Materials and Methods
4.1. Isolation and Culture of Primary BMMSCs
4.2. Adipogenic Differentiation
4.3. Osteogenic Differentiation
4.4. CdCl2 Treatment
4.5. Cell Surface Marker Expression of BMMSCs
4.6. Cell Proliferation Assays
4.7. Confocal Microscopic Analysis
4.8. Oil Red O Staining
4.9. Alkaline Phosphatase (ALP) Assay
4.10. Alkaline Phosphatase (ALP) Staining
4.11. Alizarin Red S (ARS) Staining
4.12. RNA Preparation and Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
4.13. Reverse-Transcriptase Polymerase Chain Reaction (RT-PCR)
4.14. Western Blots
4.15. Microarray for Analysis of Gene Expression
4.16. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| CdCl2 | Cadmium chloride |
| BMMSCs | Bone marrow mesenchymal stem cells |
| p0 | Primary bone marrow mesenchymal stem cells |
| Adi-induction | Adipogenic induction |
| Ost-induction | Osteogenic induction |
| Wnt | Wingless-type |
| JAK-STAT | Janus kinase-Signal Transducers and Activators of Transcription |
| IGFBP7 | Insulin-like growth factor binding protein 7 |
| MTT | 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide |
| EdU | 5-ethynyl-2′-deoxyuridine |
| ALP | Alkaline phosphatase |
| OCN | Osteocalcin |
| Runx2 | Runt-related transcription factor 2 |
| OSX | Osterix |
| OPN | Osteopontin |
| LEF1 | Lymphoid enhancer-binding factor 1 |
| TCF1 | T cell factor 1 |
| BMD | Bone mineral density |
| ARS | Alizarin red S |
| GO | Gene oncology |
| GSK3 | Glycogen synthase kinase 3 |
| APC | Adenomatosis polyposis coli |
References
- Staessen, J.A.; Roels, H.A.; Emelianov, D.; Kuznetsova, T.; Thijs, L.; Vangronsveld, J.; Fagard, R. Environmental exposure to cadmium, forearm bone density, and risk of fractures: Prospective population study. Public health and environmental exposure to cadmium (pheecad) study group. Lancet 1999, 353, 1140–1144. [Google Scholar] [CrossRef]
- Schutte, R.; Nawrot, T.S.; Richart, T.; Thijs, L.; Vanderschueren, D.; Kuznetsova, T.; Van Hecke, E.; Roels, H.A.; Staessen, J.A. Bone resorption and environmental exposure to cadmium in women: A population study. Environ. Health Perspect. 2008, 116, 777–783. [Google Scholar] [CrossRef]
- Lv, Y.; Wang, P.; Huang, R.; Liang, X.; Wang, P.; Tan, J.; Chen, Z.; Dun, Z.; Wang, J.; Jiang, Q.; et al. Cadmium exposure and osteoporosis: A population-based study and benchmark dose estimation in southern China. J. Bone Miner. Res. 2017, 32, 1990–2000. [Google Scholar] [CrossRef]
- Burm, E.; Ha, M.; Kwon, H.J. Association between blood cadmium level and bone mineral density reduction modified by renal function in young and middle-aged men. J. Trace Elem. Med. Biol. 2015, 32, 60–65. [Google Scholar] [CrossRef]
- Shen, Y.; Wu, L.; Qin, D.; Xia, Y.; Zhou, Z.; Zhang, X.; Wu, X. Carbon black suppresses the osteogenesis of mesenchymal stem cells: The role of mitochondria. Part. Fibre Toxicol. 2018, 15, 16. [Google Scholar] [CrossRef]
- Shen, Y.; Wu, L.; Wang, J.; Wu, X.; Zhang, X. The role of mitochondria in methamphetamine-induced inhibitory effects on osteogenesis of mesenchymal stem cells. Eur. J. Pharmacol. 2018, 826, 56–65. [Google Scholar] [CrossRef]
- Zhu, J.H.; Liao, Y.P.; Li, F.S.; Hu, Y.; Li, Q.; Ma, Y.; Wang, H.; Zhou, Y.; He, B.C.; Su, Y.X. Wnt11 promotes bmp9-induced osteogenic differentiation through bmps/smads and p38 mapk in mesenchymal stem cells. J. Cell Biochem. 2018, 119, 9462–9473. [Google Scholar] [CrossRef]
- Ye, C.; Chen, M.; Chen, E.; Li, W.; Wang, S.; Ding, Q.; Wang, C.; Zhou, C.; Tang, L.; Hou, W.; et al. Knockdown of foxa2 enhances the osteogenic differentiation of bone marrow-derived mesenchymal stem cells partly via activation of the erk signalling pathway. Cell Death Dis. 2018, 9, 836. [Google Scholar] [CrossRef]
- Bianco, P.; Riminucci, M.; Gronthos, S.; Robey, P.G. Bone marrow stromal stem cells: Nature, biology, and potential applications. Stem Cells 2001, 19, 180–192. [Google Scholar] [CrossRef]
- Grayson, W.L.; Bunnell, B.A.; Martin, E.; Frazier, T.; Hung, B.P.; Gimble, J.M. Stromal cells and stem cells in clinical bone regeneration. Nat. Rev. Endocrinol. 2015, 11, 140–150. [Google Scholar] [CrossRef] [Green Version]
- Lane, S.W.; Williams, D.A.; Watt, F.M. Modulating the stem cell niche for tissue regeneration. Nat. Biotechnol. 2014, 32, 795–803. [Google Scholar] [CrossRef] [Green Version]
- Murphy, W.L.; McDevitt, T.C.; Engler, A.J. Materials as stem cell regulators. Nat. Mater. 2014, 13, 547–557. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Baker, B.A.; Mou, X.; Ren, N.; Qiu, J.; Boughton, R.I.; Liu, H. Biopolymer/calcium phosphate scaffolds for bone tissue engineering. Adv. Healthc. Mater. 2014, 3, 469–484. [Google Scholar] [CrossRef]
- Gao, X.; Ge, J.; Li, W.; Zhou, W.; Xu, L. Lncrna kcnq1ot1 promotes osteogenic differentiation to relieve osteolysis via wnt/beta-catenin activation. Cell Biosci. 2018, 8, 19. [Google Scholar] [CrossRef]
- Jing, H.; Su, X.; Gao, B.; Shuai, Y.; Chen, J.; Deng, Z.; Liao, L.; Jin, Y. Epigenetic inhibition of wnt pathway suppresses osteogenic differentiation of bmscs during osteoporosis. Cell Death Dis. 2018, 9, 176. [Google Scholar] [CrossRef]
- Zhang, W.; Chen, E.; Chen, M.; Ye, C.; Qi, Y.; Ding, Q.; Li, H.; Xue, D.; Gao, X.; Pan, Z. Igfbp7 regulates the osteogenic differentiation of bone marrow-derived mesenchymal stem cells via wnt/beta-catenin signaling pathway. FASEB J. 2018, 32, 2280–2291. [Google Scholar] [CrossRef]
- Muruganandan, S.; Govindarajan, R.; McMullen, N.M.; Sinal, C.J. Chemokine-like receptor 1 is a novel wnt target gene that regulates mesenchymal stem cell differentiation. Stem Cells 2017, 35, 711–724. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, Y.; Wang, W.; Liu, Z.; Meng, J.; Han, Z. Mesenchymal stem cells modified with heme oxygenase-1 have enhanced paracrine function and attenuate lipopolysaccharide-induced inflammatory and oxidative damage in pulmonary microvascular endothelial cells. Cell Physiol. Biochem. 2018, 49, 101–122. [Google Scholar] [CrossRef]
- Zhang, T.; Li, X.H.; Zhang, D.B.; Liu, X.Y.; Zhao, F.; Lin, X.W.; Wang, R.; Lang, H.X.; Pang, X.N. Repression of coup-tfi improves bone marrow-derived mesenchymal stem cell differentiation into insulin-producing cells. Mol. Ther. Nucleic Acids 2017, 8, 220–231. [Google Scholar] [CrossRef]
- Katz, E.; Schran, H.F.; Adashi, E.Y. Successful treatment of a prolactin-producing pituitary macroadenoma with intravaginal bromocriptine mesylate: A novel approach to intolerance of oral therapy. Obstet. Gynecol. 1989, 73, 517–520. [Google Scholar]
- Chen, Q.; Shou, P.; Zheng, C.; Jiang, M.; Cao, G.; Yang, Q.; Cao, J.; Xie, N.; Velletri, T.; Zhang, X.; et al. Fate decision of mesenchymal stem cells: Adipocytes or osteoblasts? Cell Death Differ. 2016, 23, 1128–1139. [Google Scholar] [CrossRef] [PubMed]
- Yun, H.M.; Park, K.R.; Quang, T.H.; Oh, H.; Hong, J.T.; Kim, Y.C.; Kim, E.C. 2,4,5-trimethoxyldalbergiquinol promotes osteoblastic differentiation and mineralization via the bmp and wnt/beta-catenin pathway. Cell Death Dis. 2015, 6, e1819. [Google Scholar] [CrossRef]
- Bennett, C.N.; Ross, S.E.; Longo, K.A.; Bajnok, L.; Hemati, N.; Johnson, K.W.; Harrison, S.D.; MacDougald, O.A. Regulation of wnt signaling during adipogenesis. J. Biol. Chem. 2002, 277, 30998–31004. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Z.; Li, Q.; Luo, S.; Liu, Z.; Luo, D.; Zhang, B.; Zhang, D.; Rao, P.; Xiao, J. Ppargamma and wnt signaling in adipogenic and osteogenic differentiation of mesenchymal stem cells. Curr. Stem Cell Res. Ther. 2016, 11, 216–225. [Google Scholar] [CrossRef]
- Park, H.W.; Kim, Y.C.; Yu, B.; Moroishi, T.; Mo, J.S.; Plouffe, S.W.; Meng, Z.; Lin, K.C.; Yu, F.X.; Alexander, C.M.; et al. Alternative wnt signaling activates yap/taz. Cell 2015, 162, 780–794. [Google Scholar] [CrossRef]
- Bhattacharyya, M.H. Cadmium osteotoxicity in experimental animals: Mechanisms and relationship to human exposures. Toxicol. Appl. Pharmacol. 2009, 238, 258–265. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Wang, G.; Li, X.; Gan, C.; Zhu, G.; Jin, T.; Wang, Z. Environmental level of cadmium exposure stimulates osteoclasts formation in male rats. Food Chem. Toxicol. 2013, 60, 530–535. [Google Scholar] [CrossRef] [PubMed]
- Tomaszewska, E.; Dobrowolski, P.; Winiarska-Mieczan, A.; Kwiecien, M.; Tomczyk, A.; Muszynski, S.; Radzki, R. Alteration in bone geometric and mechanical properties, histomorphometrical parameters of trabecular bone, articular cartilage, and growth plate in adolescent rats after chronic co-exposure to cadmium and lead in the case of supplementation with green, black, red and white tea. Environ. Toxicol. Pharmacol. 2016, 46, 36–44. [Google Scholar]
- Dermience, M.; Lognay, G.; Mathieu, F.; Goyens, P. Effects of thirty elements on bone metabolism. J. Trace Elem. Med. Biol. 2015, 32, 86–106. [Google Scholar] [CrossRef]
- Wang, H.; Zhu, G.; Shi, Y.; Weng, S.; Jin, T.; Kong, Q.; Nordberg, G.F. Influence of environmental cadmium exposure on forearm bone density. J. Bone Miner. Res. 2003, 18, 553–560. [Google Scholar] [CrossRef]
- Chen, Z.; Luo, Q.; Lin, C.; Kuang, D.; Song, G. Simulated microgravity inhibits osteogenic differentiation of mesenchymal stem cells via depolymerizing f-actin to impede taz nuclear translocation. Sci. Rep. 2016, 6, 30322. [Google Scholar] [CrossRef] [PubMed]
- Tao, K.; Xiao, D.; Weng, J.; Xiong, A.; Kang, B.; Zeng, H. Berberine promotes bone marrow-derived mesenchymal stem cells osteogenic differentiation via canonical wnt/beta-catenin signaling pathway. Toxicol. Lett. 2016, 240, 68–80. [Google Scholar] [CrossRef] [PubMed]
- Qiu, J.; Li, D.; Mou, X.; Li, J.; Guo, W.; Wang, S.; Yu, X.; Ma, B.; Zhang, S.; Tang, W.; et al. Effects of graphene quantum dots on the self-renewal and differentiation of mesenchymal stem cells. Adv. Healthc. Mater. 2016, 5, 702–710. [Google Scholar] [CrossRef] [PubMed]
- Akrami, R.; Jacobsen, A.; Hoell, J.; Schultz, N.; Sander, C.; Larsson, E. Comprehensive analysis of long non-coding rnas in ovarian cancer reveals global patterns and targeted DNA amplification. PLoS ONE 2013, 8, e80306. [Google Scholar] [CrossRef] [PubMed]
- Grafe, I.; Alexander, S.; Peterson, J.R.; Snider, T.N.; Levi, B.; Lee, B.; Mishina, Y. Tgf-beta family signaling in mesenchymal differentiation. Cold Spring Harb. Perspect. Biol. 2018, 1, a022202. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.; Li, Y.; Huang, T.; Yu, Z.; Ma, K.; Yang, M.; Liu, Q.; Pan, H.; Wang, H.; Wang, J.; et al. Runx2/osterix and zinc uptake synergize to orchestrate osteogenic differentiation and citrate containing bone apatite formation. Adv. Sci. 2018, 5, 1700755. [Google Scholar] [CrossRef]
- Wang, G.; Zheng, L.; Zhao, H.; Miao, J.; Sun, C.; Ren, N.; Wang, J.; Liu, H.; Tao, X. In vitro assessment of the differentiation potential of bone marrow-derived mesenchymal stem cells on genipin-chitosan conjugation scaffold with surface hydroxyapatite nanostructure for bone tissue engineering. Tissue Eng. Part A 2011, 17, 1341–1349. [Google Scholar] [CrossRef]
- Li, J.; Mou, X.; Qiu, J.; Wang, S.; Wang, D.; Sun, D.; Guo, W.; Li, D.; Kumar, A.; Yang, X.; et al. Surface charge regulation of osteogenic differentiation of mesenchymal stem cell on polarized ferroelectric crystal substrate. Adv. Healthc. Mater. 2015, 4, 998–1003. [Google Scholar] [CrossRef] [PubMed]
- Langdahl, B.; Ferrari, S.; Dempster, D.W. Bone modeling and remodeling: Potential as therapeutic targets for the treatment of osteoporosis. Ther. Adv. Musculoskelet Dis. 2016, 8, 225–235. [Google Scholar] [CrossRef] [PubMed]
- Knight, M.N.; Karuppaiah, K.; Lowe, M.; Mohanty, S.; Zondervan, R.L.; Bell, S.; Ahn, J.; Hankenson, K.D. R-spondin-2 is a wnt agonist that regulates osteoblast activity and bone mass. Bone Res. 2018, 6, 24. [Google Scholar] [CrossRef]
- Muruganandan, S.; Roman, A.A.; Sinal, C.J. Adipocyte differentiation of bone marrow-derived mesenchymal stem cells: Cross talk with the osteoblastogenic program. Cell Mol. Life Sci. 2009, 66, 236–253. [Google Scholar] [CrossRef] [PubMed]
- Reya, T.; Clevers, H. Wnt signalling in stem cells and cancer. Nature 2005, 434, 843–850. [Google Scholar] [CrossRef] [PubMed]
- Kato, M.; Patel, M.S.; Levasseur, R.; Lobov, I.; Chang, B.H.; Glass, D.A., 2nd; Hartmann, C.; Li, L.; Hwang, T.H.; Brayton, C.F.; et al. Cbfa1-independent decrease in osteoblast proliferation, osteopenia, and persistent embryonic eye vascularization in mice deficient in lrp5, a wnt coreceptor. J. Cell Biol. 2002, 157, 303–314. [Google Scholar] [CrossRef] [PubMed]
- Pandur, P.; Maurus, D.; Kuhl, M. Increasingly complex: New players enter the wnt signaling network. Bioessays 2002, 24, 881–884. [Google Scholar] [CrossRef] [PubMed]
- Aida, Y.; Kurihara, H.; Kato, K. Wnt3a promotes differentiation of human bone marrow-derived mesenchymal stem cells into cementoblast-like cells. In Vitro Cell Dev. Biol. Anim. 2018, 54, 468–476. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative pcr and the 2(-delta delta c(t)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GAPDH | 5′-GGCACAGTCAAGGCTGAGAATG-3′ |
| 5′-ATGGTGGTGAAGACGCCAGTA-3′ | |
| ALP | 5′-GGGACTGGTACTCGGACAAT-3′ |
| 5′-GGCCTTCTCATCCAGTTCAT-3′ | |
| OCN | 5′-CATGAGGACCCTCTCTCTGC-3′ |
| 5′-TGGACATGAAGGCTTTGTCA-3′ | |
| Runx2 | 5′-GCACCCAGCCCATAATAGA-3′ |
| 5′-TTGGAGCAAGGAGAACCC-3′ | |
| OSX | 5′-AGCGACCACTTGAGCAAACAT-3′ |
| 5′-GCGGCTGATTGGCTTCTTCT-3′ | |
| OPN | 5′-CTTTCACTCCAATCGTCCCTAC-3′ |
| 5′-CTGCCCTTTCCGTTGTTGTC-3′ |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, L.; Wei, Q.; Lv, Y.; Xue, J.; Zhang, B.; Sun, Q.; Xiao, T.; Huang, R.; Wang, P.; Dai, X.; et al. Wnt/β-Catenin Pathway Is Involved in Cadmium-Induced Inhibition of Osteoblast Differentiation of Bone Marrow Mesenchymal Stem Cells. Int. J. Mol. Sci. 2019, 20, 1519. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20061519
Wu L, Wei Q, Lv Y, Xue J, Zhang B, Sun Q, Xiao T, Huang R, Wang P, Dai X, et al. Wnt/β-Catenin Pathway Is Involved in Cadmium-Induced Inhibition of Osteoblast Differentiation of Bone Marrow Mesenchymal Stem Cells. International Journal of Molecular Sciences. 2019; 20(6):1519. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20061519
Chicago/Turabian StyleWu, Lu, Qinzhi Wei, Yingjian Lv, Junchao Xue, Bo Zhang, Qian Sun, Tian Xiao, Rui Huang, Ping Wang, Xiangyu Dai, and et al. 2019. "Wnt/β-Catenin Pathway Is Involved in Cadmium-Induced Inhibition of Osteoblast Differentiation of Bone Marrow Mesenchymal Stem Cells" International Journal of Molecular Sciences 20, no. 6: 1519. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20061519