3.2. Plant Microbe Interaction for AMF Structural Development and Fusarium Incidence Inhibition
The gradual increase in the limited C-substrate availability and the profession of the pathogenic infection in our PGVC planting system radically hampered the indigenous AM development and restricted mycorrhizal infection, the formation of spores, hyphal growth, and the host-plant root expansion. These AM indicators are the key participants for intensive agroecosystems because of their absence or occurrence, and their particular affiliation with species can determine the mycorrhizal effectiveness and contribute to shaping the community composition [
40]. It has also been observed that the root colonization rate is directly associated with the soil spore density [
41]. In addition, the results suggested that the AM fungal host plants also secrete chemical factors, which attract and enhance the growth of the developing spore hyphae towards the root system [
17].
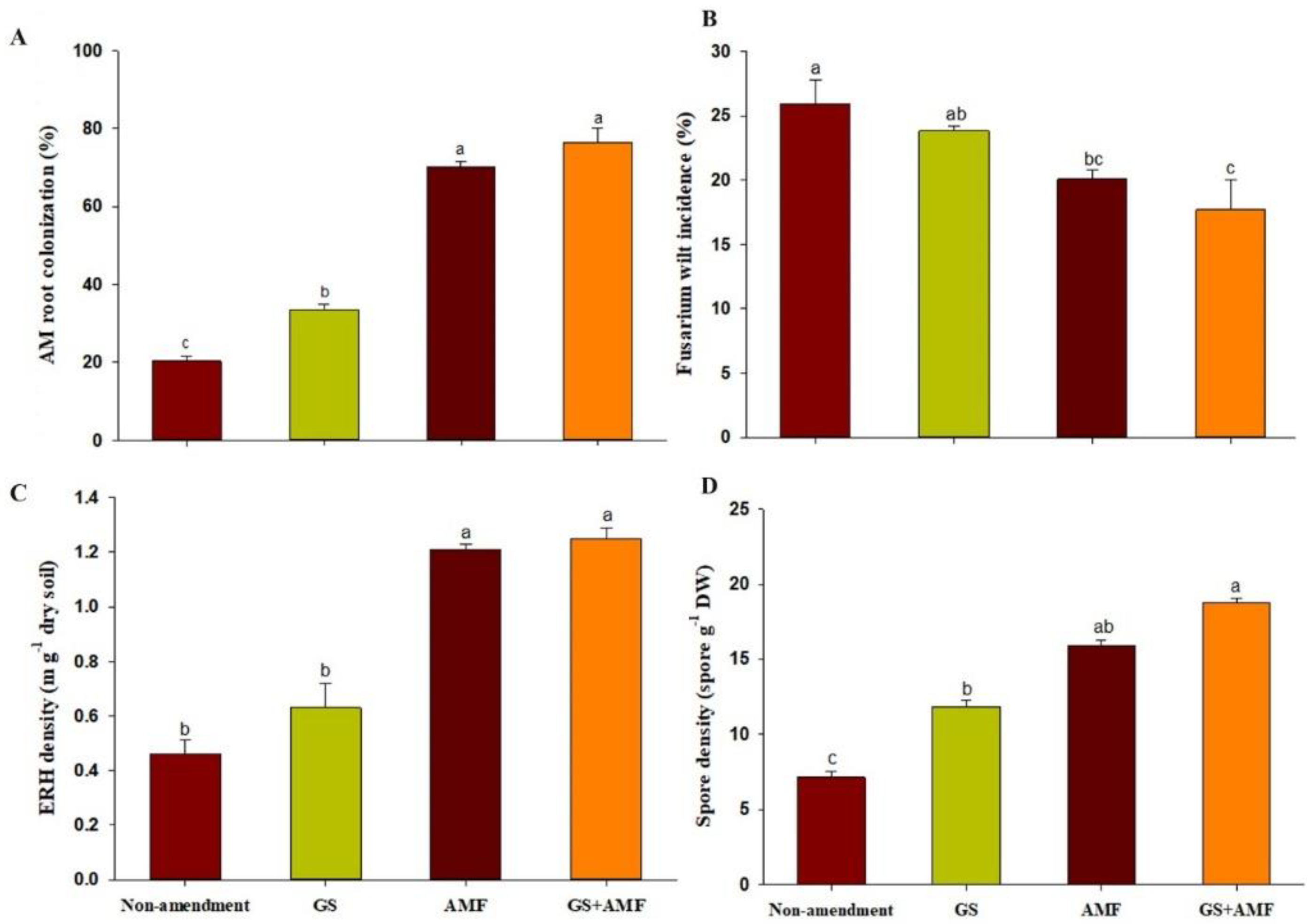
In this study, the organic substrate (GS) and AM inocula enhanced the cucumber associated AM-colonization, ERH density, spore density, and decreased the infection rate of the Fusarium wilt. The high AM root colonization, sporulation, and ERH density were identified with abundant AM species (
Table S4). The development of the
Glomaceae isolates may be generally attributed as the fastest colonizers of plant roots, and this is consistent with other studies in diversified agroecosystems [
26,
27]. External nutrient inputs, in particular, the demand for available N and AP, and the plant substrate quality could be factors responsible for the multiplication of AM growth. The host plant roots (and their associated AM fungal traits) articulate the sensitive response to the external P supply. The AM root colonization percentage may decouple from the soil in response to excessive or deficient P supply. Moreover, it was reported that the external hyphae of the AM fungi require a 4–7 times greater N concentration than that of the plant shoots and at least ten times greater than that of the roots [
23]. Subsequently, the AM fungi have a substantial N demand and entail large amounts of N for their own growth and structural development [
42].
The Glomus hyphae rely on plant-derived carbon and grow prolifically with the appropriate C-allocation through organic material and shift the structure and function of the mycorrhizal communities [
43]. The development of the mycorrhizal hyphae in this study could be an integral contribution to the AM-ecosystem services from the plant derived C decomposition and nutrient acquisition from the soil to become available to the microbiota [
44]. The added external organic and inorganic inputs were sufficient for the proliferation of AM growth, and this can explain why the AM fungal growth was stimulated by the C-amended garlic substrate [
44].
In our cultivation system, it appears that anthropogenic activities greatly reduced fungal development, but were not completely lost following the long continuous cropping spans prior to the establishment of our experiments. We speculate that the fungal taxa were probably present in the system but grew at low undetectable levels. When suitable host plants became established under enriched organic inputs, the AM symbiotic fungi quickly rebounded, increasing root colonization and spore production rates, suggesting that the AM development became more active with exogenous AM+GS amendments over the four seasons of our experiment. Reference [
25] also elucidated the responses of AM fungal abundance and community composition simulated by organic amendments found in anthropogenically disturbed environments.
Pre-existing hyphae, germinating spores and infected root fragments are also considered to be the key players in new host establishment. It is also worth noting that the differences in AM development were the greatest within species, and this observation was primarily driven by AM colonization, mycorrhizal hyphal, and sporulation increases of a particular fungus,
Glomeraceae, known as AM-Glomus spp, indicating the recruiting preferences of the AMF within particular genera, since these results are consistent with those of previous studies [
23,
45,
46].
These consistent results indicated that the new colonization, hyphal growth, spore abundance and host-symbiont interaction of AM-Glomus largely depends on the number of fungal propagules present in the soil, compatible host-plant species, and the types of soil organic additives [
14,
47]. However, different strains of AM fungi show variable responses, and their mycorrhizal associations may shift from beneficial to detrimental for the plant growth response. This dependency of the heterogenetic response is dependent on the indigenous inoculum potential, new host-range access to compatible symbiont partners, and favorable experimental conditions [
21] that have remained largely unexplored in intensive PGVC systems. However, it is clear from this study that the addition of garlic stalk as an organic residue improved the mycorrhizal strength in the degraded environment, directly supporting the hypothesis that the effect of garlic-derived components promoted the structural development of AMF characteristic and contributed in compatible synergistic interaction.
During consecutive trials, we found that cucumber is recognized as a highly mycorrhizal-dependent vegetable crop, which has not only recolonized naturally on this cultivated soil but also can be easily newly colonized by AM fungi (colonized > 75%), and its compatible host interactions are useful to establish of AM associations. In addition, the utilization of garlic residue could ensure the sufficient carbon substrate allocation to the AM fungi. These cumulative conditions probably exert the successful developmental impact in prompt AM synergistic interactions to perform a further role in crop productivity mobilizing belowground microbiota activation [
48,
49].
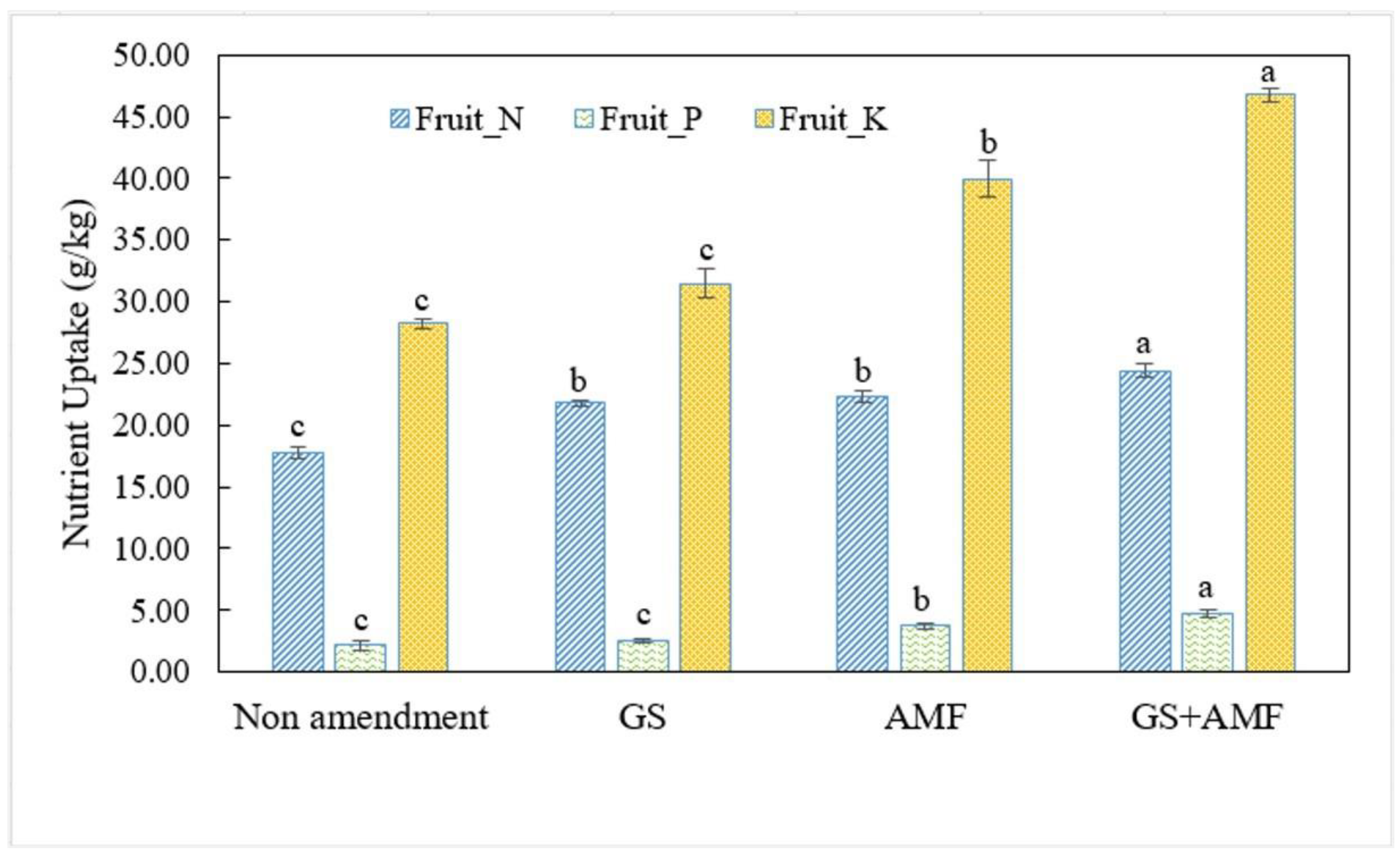
A fully developed AM-structure delivers the multiple functions in the association and facilitates the two-way movement of nutrients between the host and mutualistic fungal partner. The symbiotic association more effectively renders the host plant tolerant to various environmental stresses [
50]. In this study, we observed the additive effect between the AM fungi and organic substrates (GS) and found a close relationship between the fruit nutritional status and Chl synthesis (
Table S3). The increase in the fruit nutrient uptake under the interactive effect of GS+AM was associated with a reduction in Fusarium infection and higher rooting activity (
Table S3), suggesting that AM-inoculated seedlings distributed a favorable nutrient profile to the fruit tissue and received extended protection to endure anthropogenic stresses. A possible mechanism of increased N and K uptake and growth in GS+AM inoculated plants may be due to their effects in strengthening the source to sink capacity and resulted in a stimulation of nutrient transfer from the soil to roots and root-fruit tissue [
4]. Our results are consistent with those that found that improved P nutrition via mycorrhizal symbiosis (preferably an extension the hyphal length) caused the significant impact on fruit quality and subsequent growth expansion, which are the most important feedback of the synergistic interaction developed in the presence of organic amendments [
22].
In sustainable intensive agroecosystems, AM-based bio-inoculants are the most prevalent type of biocontrol agents to ameliorate plant growth and reduce the damage caused by soilborne plant pathogens. We found a co-inoculation antagonistic relationship with a cucumber root pathogen (Fusarium infection) with the AM-inoculum + GS-substrate treatments, and the observed reduction, due to the strength of the applied dual inoculants responded with a more effective induction of rooting structure, increases in root activity, and thus, a pronounced influence on the fruit quality attributes (
Table 6,
Tables S2 and S3). The pragmatic bio-efficacy of this study could plausibly explain that garlic or garlic-derived potent compounds (flavonoids, sulfur-containing compounds, phenolic, carbohydrates, and antioxidants enzymes) have not only been recognized as bio stimulants [
48,
49,
51] but have also been reported to be strong antimicrobial agents [
52,
53,
54].
The microbial-mediated decomposition of organic materials in the soil can create favorable anaerobic conditions and produce toxic metabolites, which results in the inhibition of soilborne pathogens or the immobilization of their activity [
55]. It may be possible that organic or allelopathic compounds released during residue decomposition and their bioactivity in the vicinity of the inoculated host plants may act in three manners; AM compounds that stimulate the plants, a disruption in the AM symbiosis, or action as an inducer for pathogenic fungal resistance [
19].
Indeed, it is generally suggested that various garlic tissues/organs (root exudates, garlic stalk, and garlic bulb /root extract) possessed different allelopathic potentials (+tive and -tive) depending on their mode of action, and their specificity has been verified by our previous research findings [
11,
12,
33]. However, plant chemistry, species adaptability, amount, soil type, and pertinent soil edaphic factors are key determinants to predict the derivatives of the organic materials tested [
6,
7]. By viewing the plant growth response in this study, we hypothesize that the applied garlic substrates comprised of aboveground selective components (only leaves and stalks) are compatible to adopt a particular fungal partner (
Glomus versiforme), and their putative contribution from growth promotion to plant protection were likely to be due to allelopathic regulation, rather than allelopathic interference. These findings are partially equivalent to other reports in which a synergistic co-inoculation effect through soil treatment improved the yield and fruit quality in a number of vegetable crops in potted studies under greenhouse conditions [
56,
57].
3.3. Plant Microbe Interaction for the AM Community Composition and Cucumber Productivity Feedback
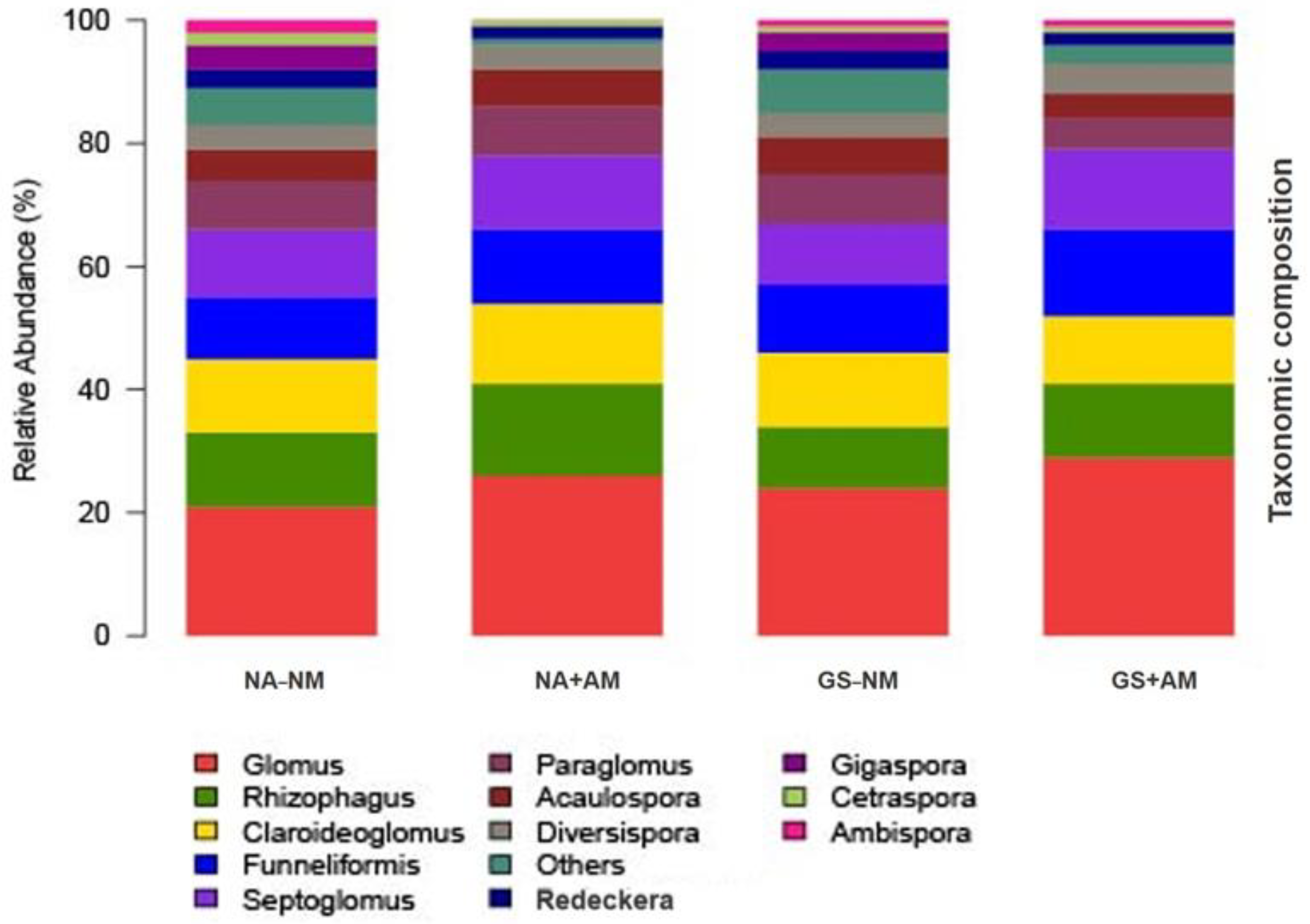
The fungal community identified in this study predominantly assessed from AM-Glomeromycotan sequences using Illumina MiSeq approach (
Figure S1) suggests that the AMF have the capacity to interact with others soil microbial communities during litter decomposition [
29]. The predominant
Glomus taxa served as a resilient ecological indicator species with widespread co-occurrence profiling across all the soil samples. These results are consistent with and confirm a previous study that the
Glomus species were the most abundant in the AMF assemblage, and their enrichment in the soil acts as a mycorrhizal developer that enabled the host plants to develop an effective AM symbiosis [
24]. The occurrence of
Glomus as the dominant members in the AMF assemblage among those of other genera could be influenced by particular factors. The related investigation revealed that the species that belong to the
Glomus genus could usually emerge with large numbers of spore production and hyphal fragments, which can rapidly colonize and extensively spread onto the AM-infected roots of plants. The influence of these factors may be the reason for the dominance of the Glomus genus in this study [
58]. Therefore, these structural improvements assist the profusion and proliferation of
Glomus genus members in a degraded ecosystem, and the appearance of this developmental phenomenon is also the result of adaptation to the local ecological environment.
Similarly, a remarkable difference in AM fungal diversity was detected after the final spring cultivation of cucumber in 2018 (
Table 7). The mycorrhizal seedlings under the GS-substrate significantly induced the Glomus-OTUs richness and diversity (Shannon) that were greatly suppressed in the continuous cropping and NA-NM treatments [
18,
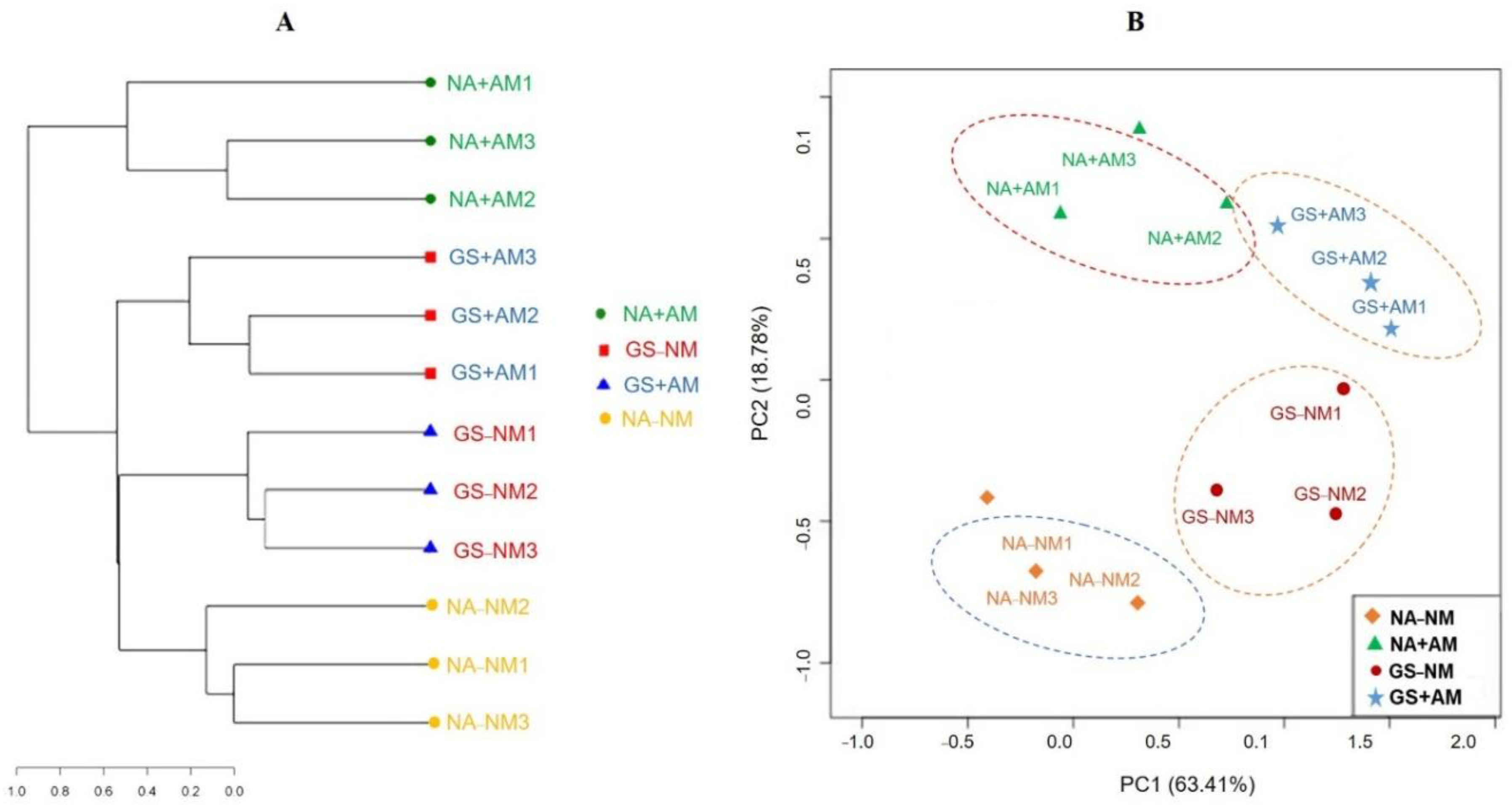
59]. In addition, the OTU-based AM composition showed that the soil samples from all the treatments at the same sampling site clustered together, indicating similarities in the AMF community composition by hierarchical cluster analysis results (
Figure 5A). Variation among the samples from different treatments separated by the PCA analysis indicated that the continuous addition of the GS+AM treatments led to divergences in the microbial community structure and shaped the trajectory of succession in distinctive manners (
Figure 5B), which were consistent with these previous studies [
60,
61].
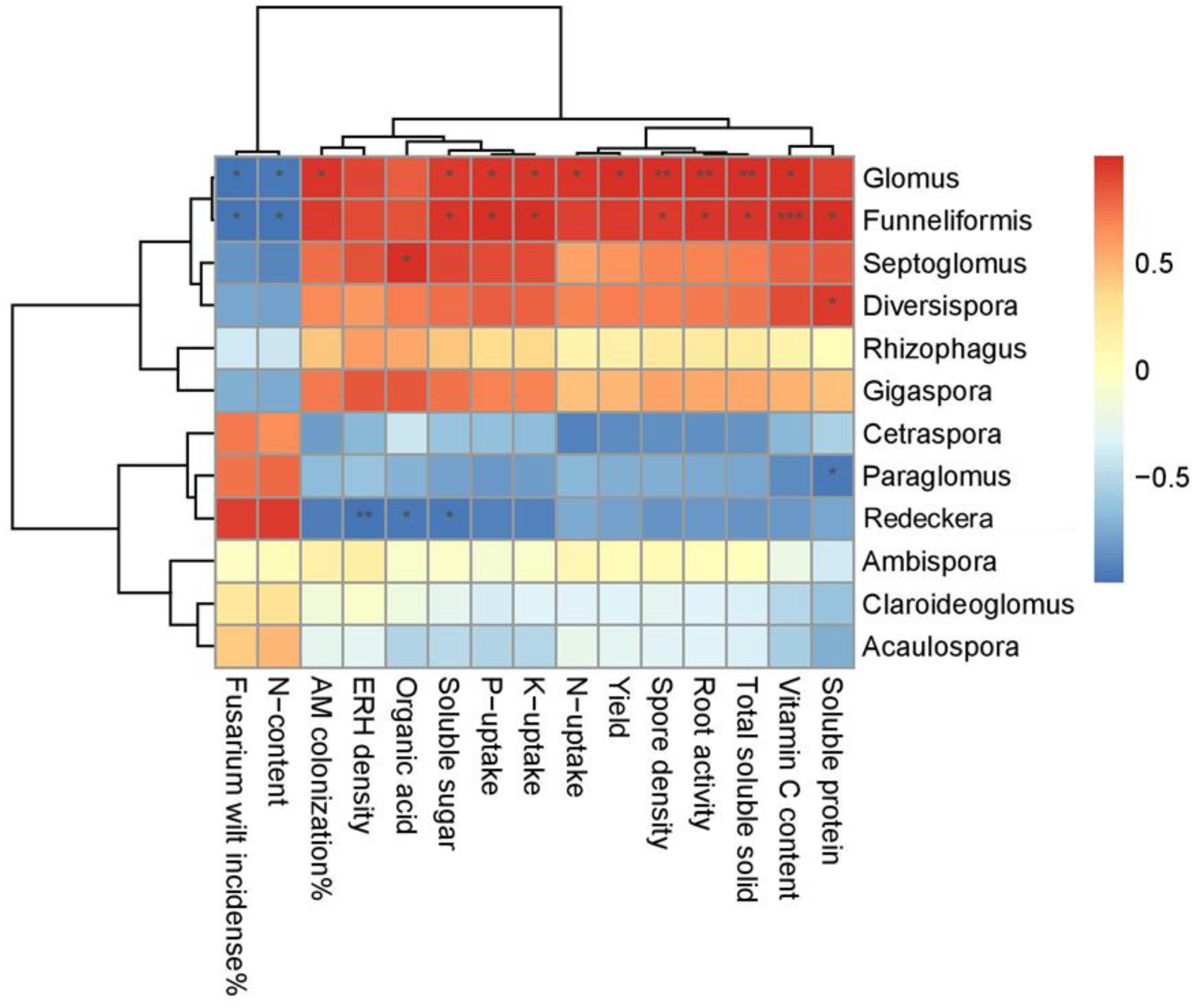
In this study, the fully developed structure of AM-Glomus was found to be a key ecological driver influencing plant growth and crop productivity. The cucumber yield associated with the Glomus taxon, and therefore, the dominant community composition, significantly increases the seasonal fruit yield, indicating the presence of intrinsic connections of the Glomus-AM symbiosis among biomass production, AM structural improvement and cucumber development. This observation is parallel with other results and implies that the taxonomic associated Glomus community composition and diversity have influenced the crop yield in diversified cropping systems [
59]. In addition, this research also confirmed that there are significant relationships between the AMF development, Glomus community structure and fruit quality attributes (
Figure 6). The increases in the fruit TSS, SS, OA, VC, and SP were positively associated [
62], while the Fusarium suppression and reduction in nitrate concentration were negatively correlated with the enriched Glomus taxon [
63]. These interactions were most likely attributable to the fact that the greatest colonization, spore germination and hyphal growth of Glomus species followed by
Funneliformis can directly exert an impact on fruit productivity traits, which may be stimulated by the favorable mutualism induced by the treatments applied [
4,
16].
Others causation of the AM community composition and diversity changes, due to the addition of fertilizer supplements, soil biochemical and seasonality impact may contribute to modify the AM fungal community [
28]. However, we are uncertain about discriminating the particular influence in this context being beyond the scope of the study. Nevertheless, we found that direct response of growth productivity improvements and community composition changes to these treatments could be an indirect response from the soil modification and successive seasonal impact. Despite the role of these elements, our data suggested that the treatments applied are the sole factor responsible for the shift of the AM fungal community, and the garlic-AM symbiosis may have superseded the influence of these factors on the AM fungal diversity. Thus, derived feedbacks from the AM and organic substrate are the primary implications for local PGVC to understand the composition, diversity, and productivity.








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}