Mel1c Mediated Monochromatic Light-Stimulated IGF-I Synthesis through the Intracellular Gαq/PKC/ERK Signaling Pathway


Abstract
:1. Introduction
2. Results
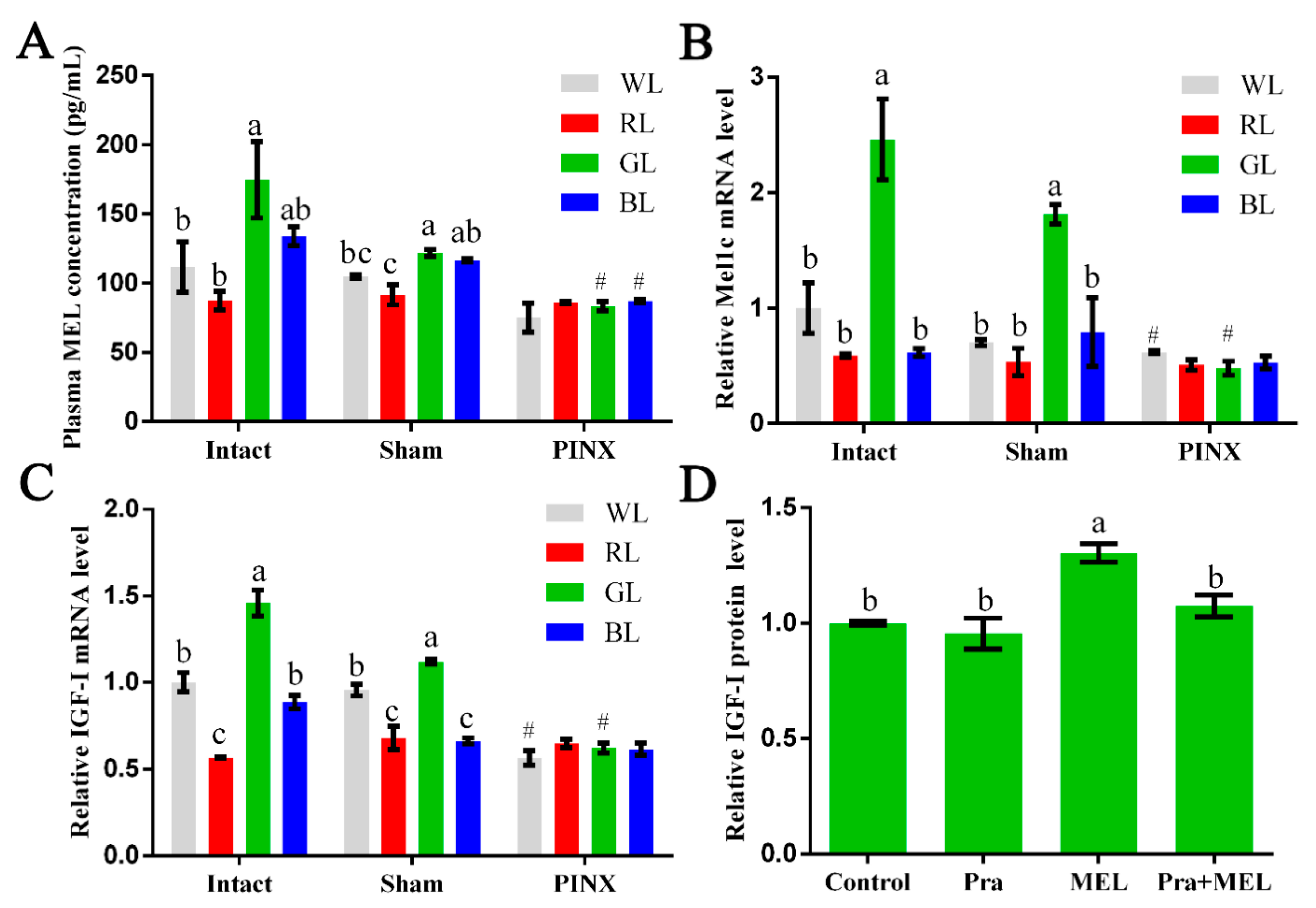
2.1. Monochromatic Green Light-Regulated Hepatic IGF-I Expression through Mel1c
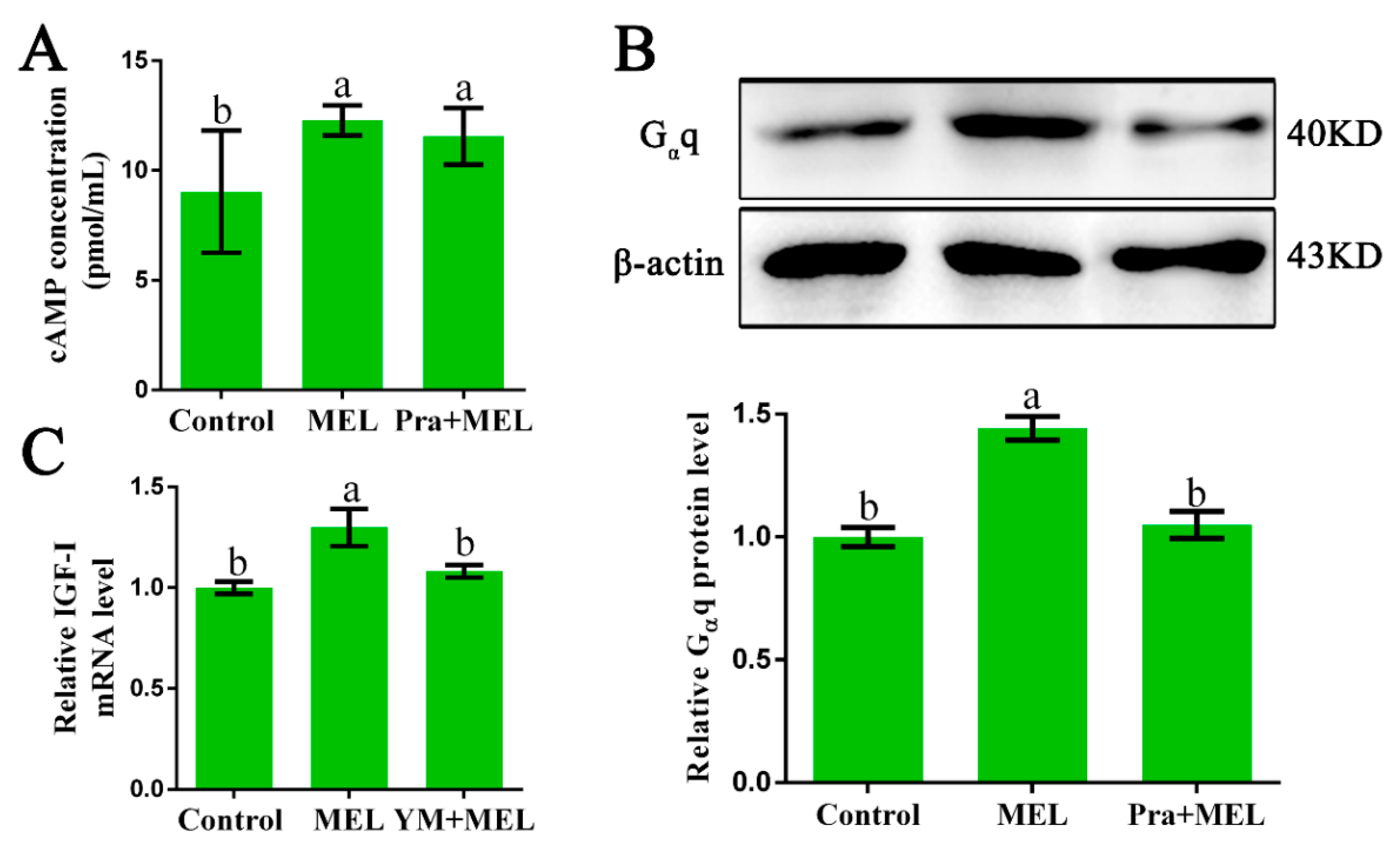
2.2. Gαq Coupled to Mel1c is Involved in MEL-Induced IGF-I Expression
2.3. Mel1c-Activated PKC through Gαq Involved in MEL-Induced IGF-I Expression
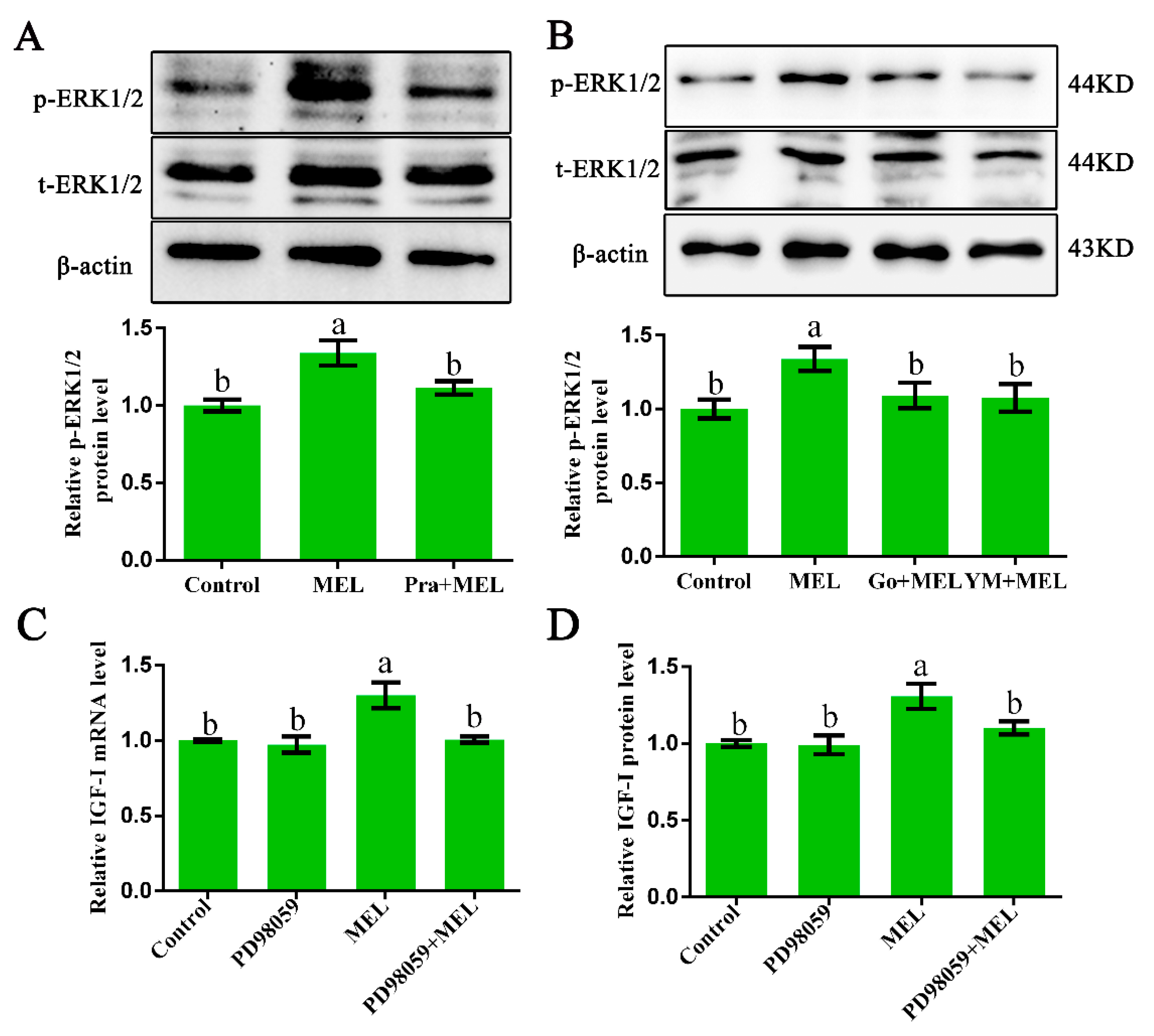
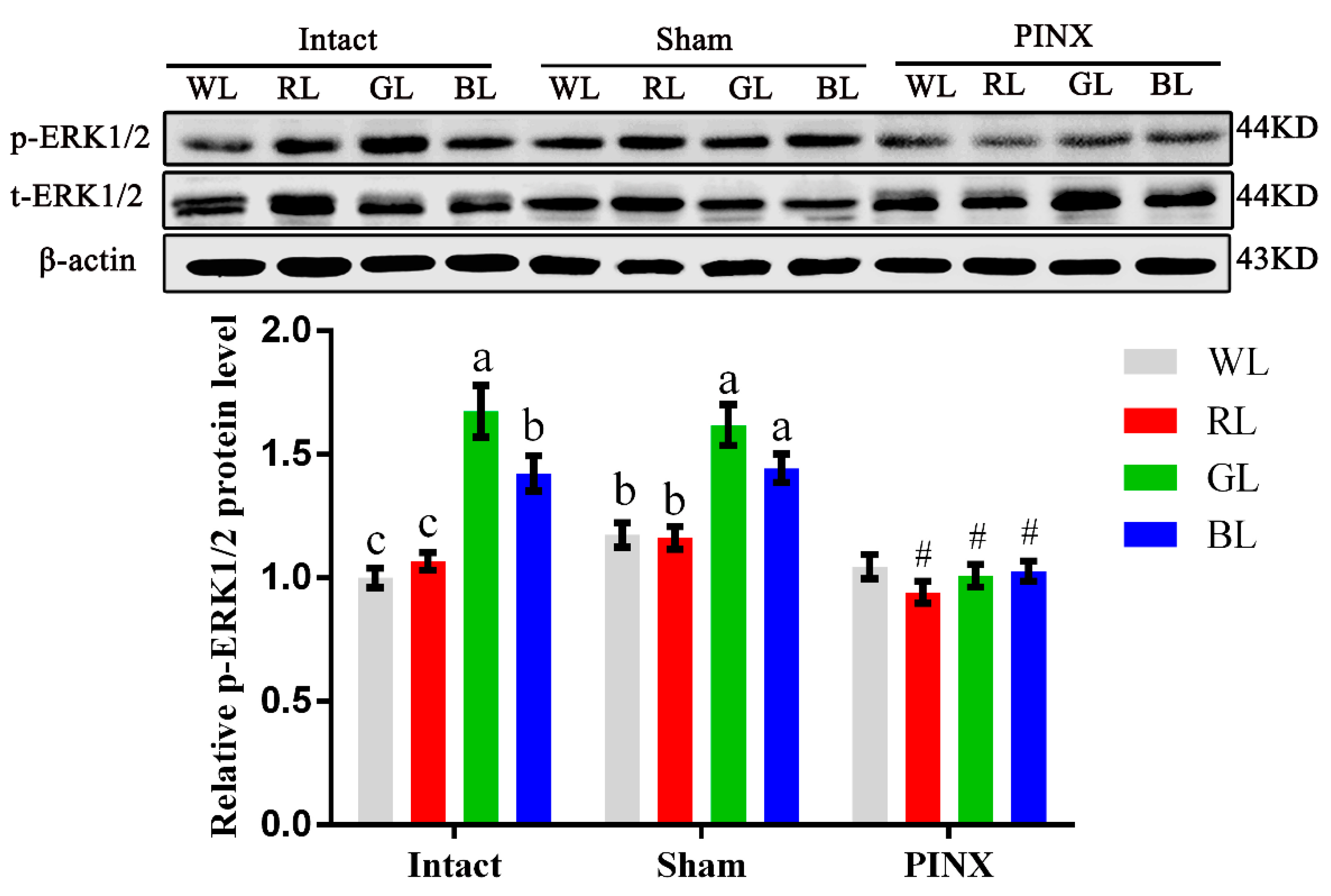
2.4. ERK1/2 Participates in Gαq-Coupled Mel1c-Mediated IGF-I Synthesis
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Animals and Treatment
4.3. Primary Hepatocytes Isolation and Culture
4.4. ELISA
4.5. Quantitative Real-Time PCR (QRT-PCR)
4.6. Western Blot Analysis
4.7. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Levenick, C.K.; Leighton, A.T. Effects of photoperiod and filtered light on growth, reproduction, and mating behavior of turkeys. 1. growth performance of two lines of males and females. Poult. Sci. 1988, 67, 1505. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, H.J.; Qiao, X.; Yue, H.Y.; Wu, S.G.; Yao, J.H.; Qi, G.H. Effect of monochromatic light stimuli during embryogenesis on muscular growth, chemical composition, and meat quality of breast muscle in male broilers. Poult. Sci. 2012, 91, 1026. [Google Scholar] [CrossRef]
- Rozenboim, I.; El Halawani, M.E.; Kashash, Y.; Piestun, Y.; Halevy, O. The effect of monochromatic photostimulation on growth and development of broiler birds. Gen. Comp. Endocrinol. 2013, 190, 214–219. [Google Scholar] [CrossRef]
- Halevy, O.; Biran, I.; Rozenboim, I. Various light source treatments affect body and skeletal muscle growth by affecting skeletal muscle satellite cell proliferation in broilers. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 1998, 120, 317–323. [Google Scholar] [CrossRef]
- Wang, T.; Dong, Y.; Wang, Z.; Cao, J.; Chen, Y. Secretion pathway of liver IGF-1 via JAK2/STAT3 in chick embryo under the monochromatic light. Growth Factors 2016, 34, 51–63. [Google Scholar] [CrossRef]
- Dishon, L.; Avitalcohen, N.; Malamud, D.; Heiblum, R.; Druyan, S.; Porter, T.E.; Gumulka, M.; Rozenboim, I. In-ovo monochromatic green light photostimulation enhances embryonic somatotropic axis activity. Poult. Sci. 2017, 96, 1884–1890. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.I.; Clemmons, D.R. Insulin-like growth factors and their binding proteins: Biological actions. Endocr. Rev. 1995, 16, 3–34. [Google Scholar] [PubMed]
- Ohlsson, C.; Mohan, S.; Sjögren, K.; Tivesten, A.; Isgaard, J.; Isaksson, O.; Jansson, J.O.; Svensson, J. The role of liver-derived insulin-like growth factor-I. Endocr. Rev. 2009, 30, 494–535. [Google Scholar] [CrossRef]
- Bai, X.; Wang, Y.; Wang, Z.; Cao, J.; Dong, Y.; Chen, Y. In ovo exposure to monochromatic lights affect posthatch muscle growth and satellite cell proliferation of chicks: Role of IGF-1. Growth Factors 2016, 34, 107–118. [Google Scholar] [CrossRef]
- Liu, W.; Wang, Z.; Chen, Y. Effects of monochromatic light on developmental changes in satellite cell population of pectoral muscle in broilers during early posthatch period. Anat. Rec. (Hoboken). 2010, 293, 1315–1324. [Google Scholar] [CrossRef]
- Haldar, C.; Ahmad, R. Photoimmunomodulation and melatonin. J. Photochem. Photobiol. B 2010, 98, 107–117. [Google Scholar] [CrossRef]
- Park, S.Y.; Walker, J.J.; Johnson, N.W.; Zhao, Z.; Lightman, S.L.; Spiga, F. Constant light disrupts the circadian rhythm of steroidogenic proteins in the rat adrenal gland. Mol. Cell Endocrinol. 2013, 371, 114–123. [Google Scholar] [CrossRef]
- Claustrat, B.; Brun, J.; Chazot, G. The basic physiology and pathophysiology of melatonin. Sleep Med. Rev. 2005, 9, 11–24. [Google Scholar] [CrossRef]
- Pandi-Perumal, S.R.; Srinivasan, V.; Maestroni, G.J.; Cardinali, D.P.; Poeggeler, B.; Hardeland, R. Melatonin: Nature’s most versatile biological signal. FEBS. J. 2006, 273, 2813–2838. [Google Scholar] [CrossRef]
- Jin, E.; Jia, L.; Li, J.; Yang, G.; Wang, Z.; Cao, J.; Chen, Y. Effect of monochromatic light on melatonin secretion and arylalkylamine N -Acetyltransferase mRNA expression in the retina and pineal gland of broilers. Anat. Rec. 2011, 294, 1233–1241. [Google Scholar] [CrossRef]
- Li, S.; Cao, J.; Wang, Z.; Dong, Y.; Wang, W.; Chen, Y. Melatonin mediates monochromatic light-induced Insulin-like growth factor 1 secretion of chick liver: Involvement of membrane receptors. Photochem Photobiol. 2016, 92, 595–603. [Google Scholar] [CrossRef] [PubMed]
- Shiu, S.Y.; Ng, N.; Pang, S.F. A molecular perspective of the genetic relationships of G-protein coupled melatonin receptor subtypes. J. Pineal Res. 1996, 20, 198–204. [Google Scholar] [CrossRef] [PubMed]
- Jarzynka, M.J.; Passey, D.K.; Ignatius, P.F.; Melan, M.A.; Radio, N.M.; Jockers, R.; Rasenick, M.M.; Brydon, L.; Witt-Enderby, P.A. Modulation of melatonin receptors and G-protein function by microtubules. J. Pineal Res. 2010, 41, 324–336. [Google Scholar] [CrossRef]
- Shiu, S.Y.; Pang, B.; Tam, C.W.; Yao, K.M. Signal transduction of receptor-mediated antiproliferative action of melatonin on human prostate epithelial cells involves dual activation of Gα(s) and Gα(q) proteins. J. Pineal Res. 2010, 49, 301–311. [Google Scholar] [CrossRef]
- Zheng, Y. G protein control of microtubule assembly. Annu. Rev. Cell Dev. Biol. 2004, 20, 867–894. [Google Scholar] [CrossRef]
- Hepler, J.R.; Gilman, A.G. G proteins. Trends. Biochem. Sci. 1992, 17, 383–387. [Google Scholar] [CrossRef]
- Dhanasekaran, N.; Heasley, L.E.; Johnson, G.L. G protein-coupled receptor systems involved in cell growth and oncogenesis. Endocr. Rev. 1995, 16, 259. [Google Scholar] [CrossRef]
- Dhanasekaran, N.; Prasad, M.V. G protein subunits and cell proliferation. Biol. Signals Recept. 1998, 7, 109. [Google Scholar] [CrossRef]
- Rozengurt, E. Signal transduction pathways in the mitogenic response to G protein-coupled neuropeptide receptor agonists. J. Cell. Physiol. 1998, 177, 507–517. [Google Scholar] [CrossRef]
- Goldsmith, Z.G.; Dhanasekaran, D.N. G protein regulation of MAPK networks. Oncogene 2007, 26, 3122–3142. [Google Scholar] [CrossRef]
- Simon, M.I.; Strathmann, M.P.; Gautam, N. Diversity of G proteins in signal transduction. Science 1991, 252, 802–808. [Google Scholar] [CrossRef]
- Zhang, L.; Cao, J.; Wang, Z.; Dong, Y.; Chen, Y. Melatonin modulates monochromatic light-induced GHRH expression in the hypothalamus and GH secretion in chicks. Acta. Histochem. 2016, 118, 286–292. [Google Scholar] [CrossRef]
- Rozenboim, I.; Piestun, Y.; Mobarkey, N.; Barak, M.; Hoyzman, A.; Halevy, O. Monochromatic light stimuli during embryogenesis enhance embryo development and posthatch growth. Poult. Sci. 2004, 83, 1413. [Google Scholar] [CrossRef]
- Allen, R.E.; Merkel, R.A.; Young, R.B. Cellular aspects of muscle growth: Myogenic cell proliferation. J. Anim. Sci. 1979, 49, 115. [Google Scholar] [CrossRef]
- Schaeffer, H.J.; Sirotkin, A.V. Melatonin and serotonin regulate the release of insulin-like growth factor-I, oxytocin and progesterone by cultured human granulosa cells. Exp. Clin. Endocrinol. Diabetes. 1997, 105, 109–112. [Google Scholar] [CrossRef]
- Schuster, C.; Gauer, F.; Guerrero, H.; Lakhdar-Ghazal, N.; Pevet, P.; Masson-Pevet, M. Photic regulation of mt1 melatonin receptors in the Siberian hamster pars tuberalis and suprachiasmatic nuclei: Involvement of the circadian clock and intergeniculate leaflet. J. Neuroendocrinol. 2000, 12, 207–216. [Google Scholar] [CrossRef]
- Schuster, C.; Gauer, F.; Malan, A.; Recio, J.; Pévet, P.; Masson-Pévet, M. The circadian clock, light/dark cycle and melatonin are differentially involved in the expression of daily and photoperiodic variations in mt(1) melatonin receptors in the Siberian and Syrian hamsters. Neuroendocrinology 2001, 74, 55–68. [Google Scholar] [CrossRef]
- Masson-Pévet, M.; Gauer, F.; Schuster, C.; Guerrero, H.Y. Photic regulation of mt(1) melatonin receptors and 2-iodomelatonin binding in the rat and Siberian hamster. Biol. Signals Recept. 2000, 9, 188–196. [Google Scholar] [CrossRef]
- Kumar Yadav, S.; Haldar, C.; Kumar Singh, S.; Dash, D. Melatonin regulates splenocytes proliferation via IP3-dependent intracellular Ca2+ release in seasonally breeding bird, Perdicula asiatica. J. Recept. Signal Transduct. Res. 2014, 34, 233–240. [Google Scholar] [CrossRef]
- Bielawski, K.; Bielawska, A.; Sosnowska, K.; Miltyk, W.; Winnicka, K.; Pałka, J. Novel amidine analogue of melphalan as a specific multifunctional inhibitor of growth and metabolism of human breast cancer cells. Biochem. Pharmacol. 2006, 72, 320–331. [Google Scholar] [CrossRef] [PubMed]
- Mayo, J.C.; Hevia, D.; Quiros-Gonzalez, I.; Rodriguez-Garcia, A.; Gonzalez-Menendez, P.; Cepas, V.; Gonzalez-Pola, I.; Sainz, R.M. IGFBP3 and MAPK/ERK signaling mediates melatonin-induced antitumor activity in prostate cancer. J. Pineal Res. 2017, 62, e12373. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Liu, J.; Ren, L.; Wei, J.; Zhang, F.; Li, Y.; Guo, C.; Duan, J.; Sun, Z.; Zhou, X. Silica nanoparticles induce abnormal mitosis and apoptosis via PKC-δ mediated negative signaling pathway in GC-2 cells of mice. Chemosphere 2018, 208, 942–950. [Google Scholar] [CrossRef] [PubMed]
- Williams, S.R.; Son, D.S.; Terranova, P.F. Protein kinase C δ is activated in mouse ovarian surface epithelial cancer cells by 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD). Toxicology 2004, 195, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, V.G.; Gonçalves, G.L.; Costa-Pessoa, J.M.; Thieme, K.; Lins, B.B.; Casare, F.A.M.; de Ponte, M.C.; Camara, N.O.S.; Oliveira-Souza, M. Angiotensin II-induced podocyte apoptosis is mediated by endoplasmic reticulum stress/PKC-δ/p38 MAPK pathway activation and trough increased Na + /H + exchanger isoform 1 activity. BMC Nephrol. 2018, 19, 179. [Google Scholar] [CrossRef]
- Adornetto, A.; Pagliara, V.; Renzo, G.D.; Arcone, R. Polychlorinated biphenyls impair dibutyryl cAMP-induced astrocytic differentiation in rat C6 glial cell line. Febs. Open Bio. 2013, 3, 459–466. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Jia, J.; Lei, C.; Ji, L.; Chen, X.; Sang, H.; Xiong, L. Cannabinoid receptor CB1 is involved in nicotine-induced protection against Abeta1-42 neurotoxicity in HT22 cells. J. Mol. Neurosci. 2015, 55, 778–787. [Google Scholar] [CrossRef] [PubMed]
- Renzi, A.; Glaser, S.; Demorrow, S.; Mancinelli, R.; Meng, F.; Franchitto, A.; Venter, J.; White, M.; Francis, H.; Han, Y. Melatonin inhibits cholangiocyte hyperplasia in cholestatic rats by interaction with MT1 but not MT2 melatonin receptors. Am. J. Physiol. Gastrointest. Liver Physiol. 2011, 301, 634–643. [Google Scholar] [CrossRef] [PubMed]
- Yung, L.Y.; Tsim, S.T.; Wong, Y.H. Stimulation of cAMP accumulation by the cloned Xenopus melatonin receptor through G i and G z proteins. FEBS. Lett. 1995, 372, 99. [Google Scholar] [CrossRef]
- Chan, A.S.; Lai, F.P.; Lo, R.K.; Voyno-Yasenetskaya, T.A.; Stanbridge, E.J.; Wong, Y.H. Melatonin mt1 and MT2 receptors stimulate c-Jun N-terminal kinase via pertussis toxin-sensitive and -insensitive G proteins. Cell. Signal. 2002, 14, 249–257. [Google Scholar] [CrossRef]
- Tam, C.W.; Shiu, S.Y. Functional interplay between melatonin receptor-mediated antiproliferative signaling and androgen receptor signaling in human prostate epithelial cells: Potential implications for therapeutic strategies against prostate cancer. J. Pineal Res. 2011, 51, 297–312. [Google Scholar] [CrossRef] [PubMed]
- Frey, U.H.; Lieb, W.; Erdmann, J.; Savidou, D.; Heusch, G.; Leineweber, K.; Jakob, H.; Hense, H.W.; Lowel, H.; Brockmeyer, N.H.; et al. Characterization of the GNAQ promoter and association of increased Gq expression with cardiac hypertrophy in humans. Eur. Heart J. 2008, 29, 888–897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kucukkaya, B.; Arslan, D.O.; Kan, B. Role of G proteins and ERK activation in hemin-induced erythroid differentiation of K562 cells. Life Sci. 2006, 78, 1217–1224. [Google Scholar] [CrossRef] [PubMed]
- Davis, M.G.; Kawai, Y.; Arinze, I.J. Involvement of Gialpha2 in sodium butyrate-induced erythroblastic differentiation of K562 cells. Biochem J. 2000, 346, 455–461. [Google Scholar] [CrossRef] [PubMed]
- Syrovatkina, V.; Alegre, K.O.; Dey, R.; Huang, X.Y. Regulation, Signaling, and Physiological Functions of G-Proteins. J. Mol. Biol. 2016, 428, 3850–3868. [Google Scholar] [CrossRef] [PubMed]
- Strange, P.G. Signaling mechanisms of GPCR ligands. Curr. Opin. Drug Discov. Devel. 2008, 11, 196–202. [Google Scholar]
- Duc, N.M.; Kim, H.R.; Chung, K.Y. Structural mechanism of G protein activation by G protein-coupled receptor. Eur. J. Pharmacol. 2015, 763, 214–222. [Google Scholar] [CrossRef]
- Frank, M.; Thümer, L.; Lohse, M.J.; Bünemann, M. G Protein activation without subunit dissociation depends on a G{alpha}(i)-specific region. J. Biol. Chem. 2005, 280, 24584–24590. [Google Scholar] [CrossRef] [PubMed]
- Liu, A.M.; Ho, M.K.; Wong, C.S.; Chan, J.H.; Pau, A.H.; Wong, Y.H. Galpha(16/z) chimeras efficiently link a wide range of G protein-coupled receptors to calcium mobilization. J. Biomol. Screen. 2003, 8, 39. [Google Scholar] [CrossRef] [PubMed]
- Pingitore, A.; Chambers, E.S.; Hill, T.; Maldonado, I.R.; Liu, B.; Bewick, G.; Morrison, D.J.; Preston, T.; Wallis, G.A.; Tedford, C. The diet-derived short chain fatty acid propionate improves beta-cell function in humans and stimulates insulin secretion from human islets in vitro. Diabetes Obes. Metab. 2017, 19, 257. [Google Scholar] [CrossRef]
- Wang, J.; Carrillo, J.J.; Lin, H.V. GPR142 agonists stimulate glucose-dependent insulin secretion via Gq-dependent signaling. PLoS ONE 2016, 11, e0154452. [Google Scholar] [CrossRef]
- Baba, K.; Benleulmi-Chaachoua, A.; Journé, A.S.; Kamal, M.; Guillaume, J.L.; Dussaud, S.; Gbahou, F.; Yettou, K.; Liu, C.; Contreras-Alcantara, S.; et al. Heteromeric MT1/MT2 melatonin receptors modulate photoreceptor function. Sci. Signal. 2013, 6, ra89. [Google Scholar] [CrossRef]
- Ahmed, R.; Mahavadi, S.; Al-Shboul, O.; Bhattacharya, S.; Grider, J.R.; Murthy, K.S. Characterization of signaling pathways coupled to melatonin receptors in gastrointestinal smooth muscle. Regul. Pept. 2013, 184, 96–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sotovega, E.; Meza, I.; Ramírezrodríguez, G.; Benitezking, G. Melatonin stimulates calmodulin phosphorylation by protein kinase C. J. Pineal Res. 2004, 37, 98–106. [Google Scholar] [CrossRef]
- Suzuki, Y.; Zhang, H.; Saito, N.; Kojima, I.; Urano, T.; Mogami, H. Glucagon-like peptide 1 activates protein kinase C through Ca2+-dependent activation of phospholipase C in insulin-secreting cells. J. Biol. Chem. 2006, 281, 28499–28507. [Google Scholar] [CrossRef] [PubMed]
- Jacobo, S.M.; Guerra, M.L.; Hockerman, G.H. Cav1.2 and Cav1.3 are differentially coupled to glucagon-like peptide-1 potentiation of glucose-stimulated insulin secretion in the pancreatic beta-cell line INS-1. J. Pharmacol. Exp. Ther. 2009, 331, 724–732. [Google Scholar] [CrossRef]
- Shigeto, M.; Ramracheya, R.; Tarasov, A.I.; Cha, C.Y.; Chibalina, M.V.; Hastoy, B.; Philippaert, K.; Reinbothe, T.; Rorsman, N.; Salehi, A.; et al. GLP-1 stimulates insulin secretion by PKC-dependent TRPM4 and TRPM5 activation. J. Clin. Invest. 2015, 125, 4714–4728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luttrell, L.M. ‘Location, location, location’: Activation and targeting of MAP kinases by G protein-coupled receptors. J. Mol. Endocrinol. 2003, 30, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Osmond, R.I.; Sheehan, A.; Borowicz, R.; Barnett, E.; Harvey, G.; Turner, C.; Brown, A.; Crouch, M.F.; Dyer, A.R. GPCR screening via ERK 1/2: A novel platform for screening G protein-coupled receptors. J. Biomol. Screen. 2005, 10, 730. [Google Scholar] [CrossRef] [PubMed]
- Reindl, K.M.; Kittilson, J.D.; Bergan, H.E.; Sheridan, M.A. Growth hormone-stimulated insulin-like growth factor-1 expression in rainbow trout (Oncorhynchus mykiss) hepatocytes is mediated by ERK, PI3K-AKT, and JAK-STAT. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2011, 301, R236. [Google Scholar] [CrossRef]
- Carbajo-Pescador, S.; García-Palomo, A.; Martín-Renedo, J.; Piva, M.; González-Gallego, J.; Mauriz, J.L. Melatonin modulation of intracellular signaling pathways in hepatocarcinoma HepG2 cell line: Role of the MT1 receptor. J. Pineal Res. 2011, 51, 463–471. [Google Scholar] [CrossRef]
- Wang, T.; Liu, B.; Guan, Y.; Gong, M.; Zhang, W.; Pan, J.; Liu, Y.; Liang, R.; Yuan, Y.; Ye, L. Melatonin inhibits the proliferation of breast cancer cells induced by bisphenol A via targeting estrogen receptor-related pathways. Thorac. Cancer. 2018, 9, 368–375. [Google Scholar] [CrossRef]
- Castro, L.M.; Gallant, M.; Niles, L.P. Novel targets for valproic acid: Up-regulation of melatonin receptors and neurotrophic factors in C6 glioma cells. J. Neurochem. 2005, 95, 1227–1236. [Google Scholar] [CrossRef] [PubMed]
- Tocharus, C.; Puriboriboon, Y.; Junmanee, T.; Tocharus, J.; Ekthuwapranee, K.; Govitrapong, P. Melatonin enhances adult rat hippocampal progenitor cell proliferation via ERK signaling pathway through melatonin receptor. Neuroscience 2014, 275, 314–321. [Google Scholar] [CrossRef]
- Cao, J.; Liu, W.; Wang, Z.; Xie, D.; Jia, L.; Chen, Y. Green and blue monochromatic lights promote growth and development of broilers via stimulating testosterone secretion and myofiber growth. J. Appl. Poult. Res. 2008, 17, 211–218. [Google Scholar] [CrossRef]
- Karaganis, S.P.; Bartell, P.A.; Shende, V.R.; Moore, A.F.; Cassone, V.M. Modulation of metabolic and clock gene mRNA rhythms by pineal and retinal circadian oscillators. Gen. Comp. Endocrinol. 2009, 161, 179–192. [Google Scholar] [CrossRef]
- Yamanaka, N.; Kitani, H.; Mikami, O.; Nakajima, Y.; Miura, K. Serum-free culture of adult chicken hepatocytes; morphological and biochemical characterisation. Res. Vet. Sci. 1997, 62, 233–237. [Google Scholar] [CrossRef]
- Fraslin, J.M.; Touquette, L.; Douaire, M.; Menezo, Y.; Guillemot, J.C.; Mallard, J. Isolation and long-term maintenance of differentiated adult chicken hepatocytes in primary culture. In Vitro Cell Dev. Biol. 1992, 28, 615–620. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Primer Sequences (5′-3′) | Product Size (bp) | Accession No. |
|---|---|---|---|
| IGF-I | F: TTCTTGAAGGTGAAGATGCACAC | 217 | NM_001004384.2 |
| R: TTCCCTTGTGGTGTAAGCGT | |||
| Mel1c | F: AGA TAA GTG GGT TCC TGA TGG | 237 | NM_205361.1 |
| R: GCA AAG GTG CAA GAG TAA ATC | |||
| GAPDH | F: ATC ACA GCC ACA CAG AAG ACG | 124 | NM_204305 |
| R:TGA CTT TCC CCA CAG CCT TA |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ning, S.; Wang, Z.; Cao, J.; Dong, Y.; Chen, Y. Mel1c Mediated Monochromatic Light-Stimulated IGF-I Synthesis through the Intracellular Gαq/PKC/ERK Signaling Pathway. Int. J. Mol. Sci. 2019, 20, 1682. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20071682
Ning S, Wang Z, Cao J, Dong Y, Chen Y. Mel1c Mediated Monochromatic Light-Stimulated IGF-I Synthesis through the Intracellular Gαq/PKC/ERK Signaling Pathway. International Journal of Molecular Sciences. 2019; 20(7):1682. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20071682
Chicago/Turabian StyleNing, Shujie, Zixu Wang, Jing Cao, Yulan Dong, and Yaoxing Chen. 2019. "Mel1c Mediated Monochromatic Light-Stimulated IGF-I Synthesis through the Intracellular Gαq/PKC/ERK Signaling Pathway" International Journal of Molecular Sciences 20, no. 7: 1682. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20071682