Effects of High Temperature on Embryological Development and Hormone Profile in Flowers and Leaves of Common Buckwheat (Fagopyrum esculentum Moench)

 , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. Embryological Analyses
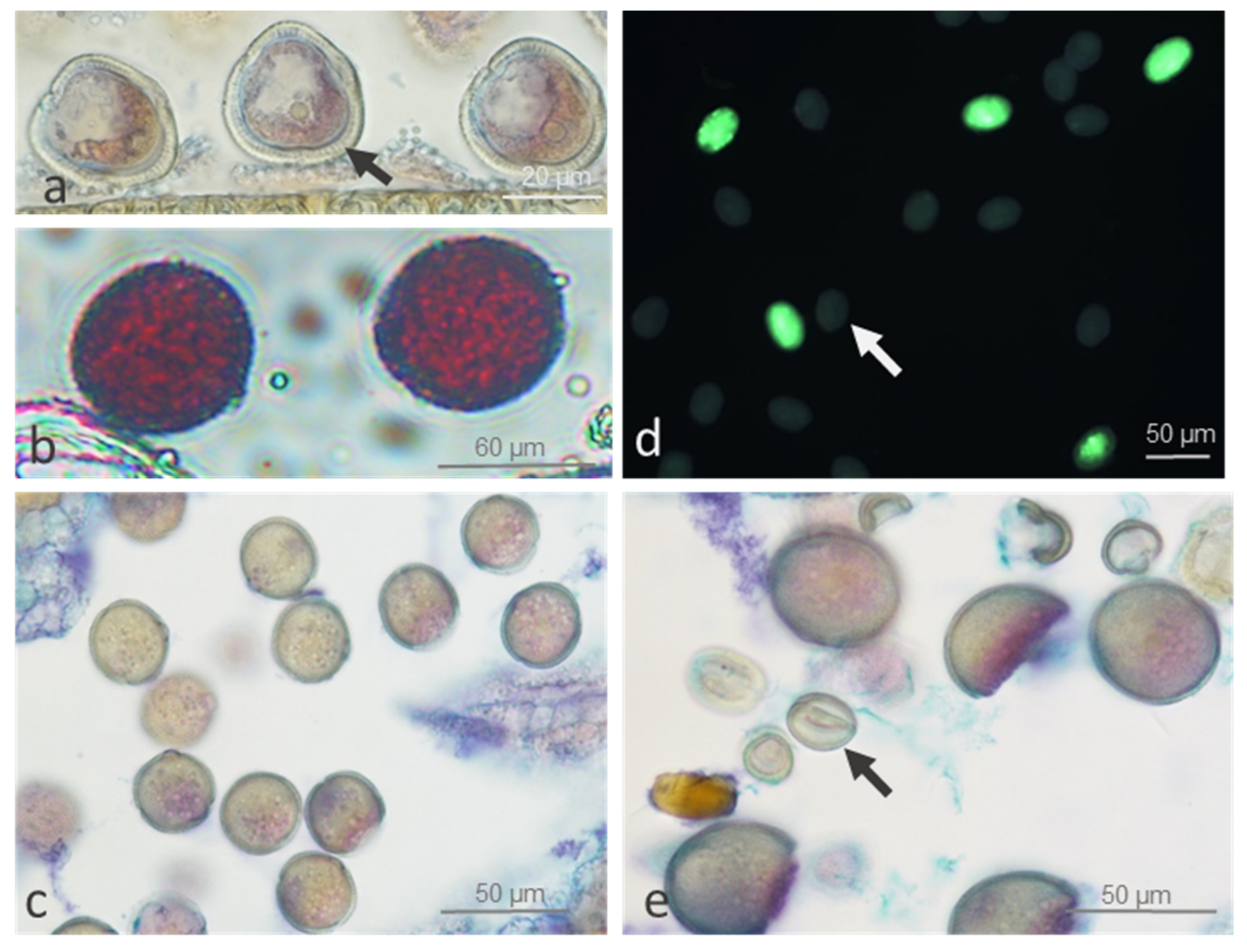
2.1.1. Pollen Viability and Development
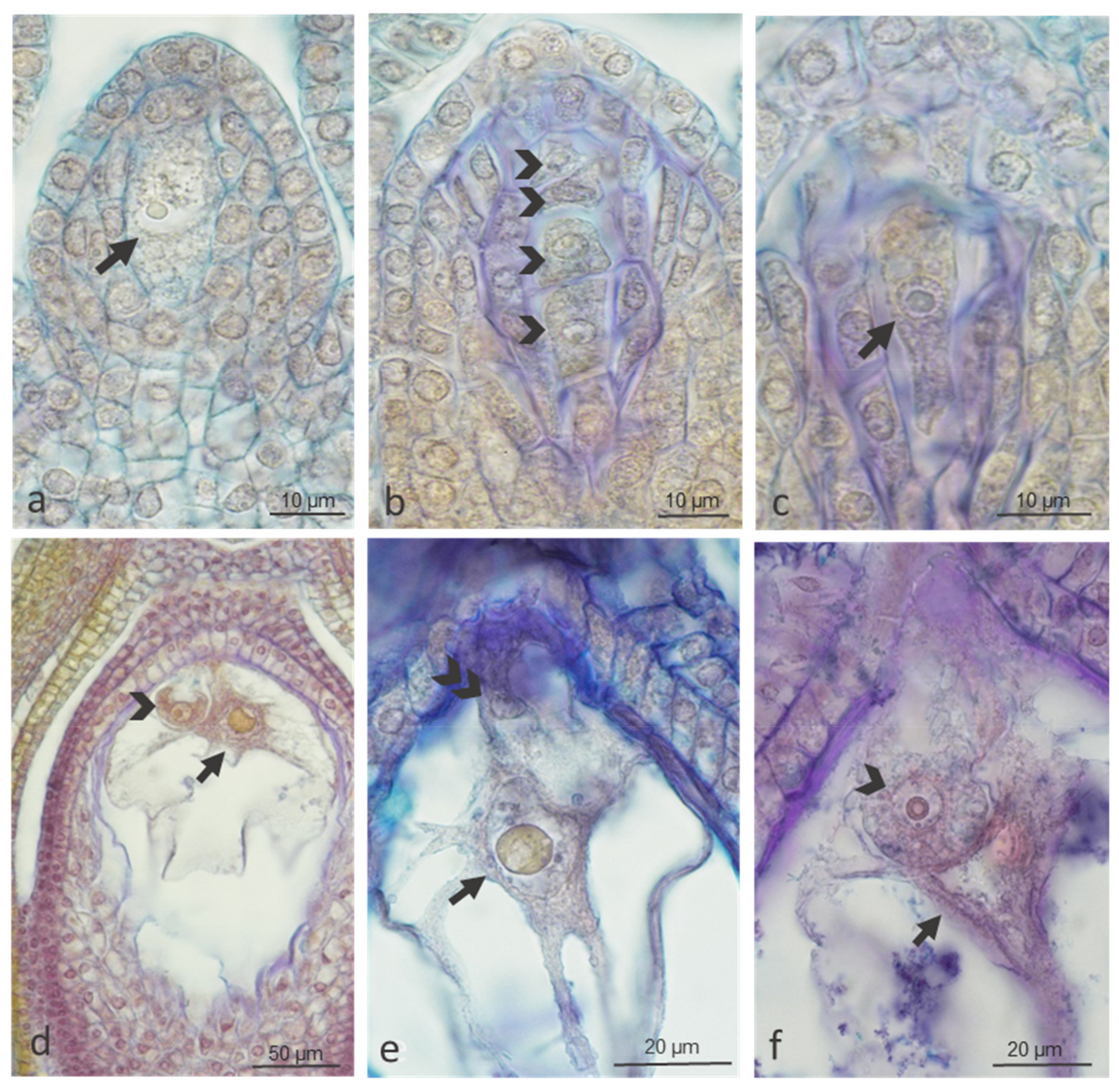
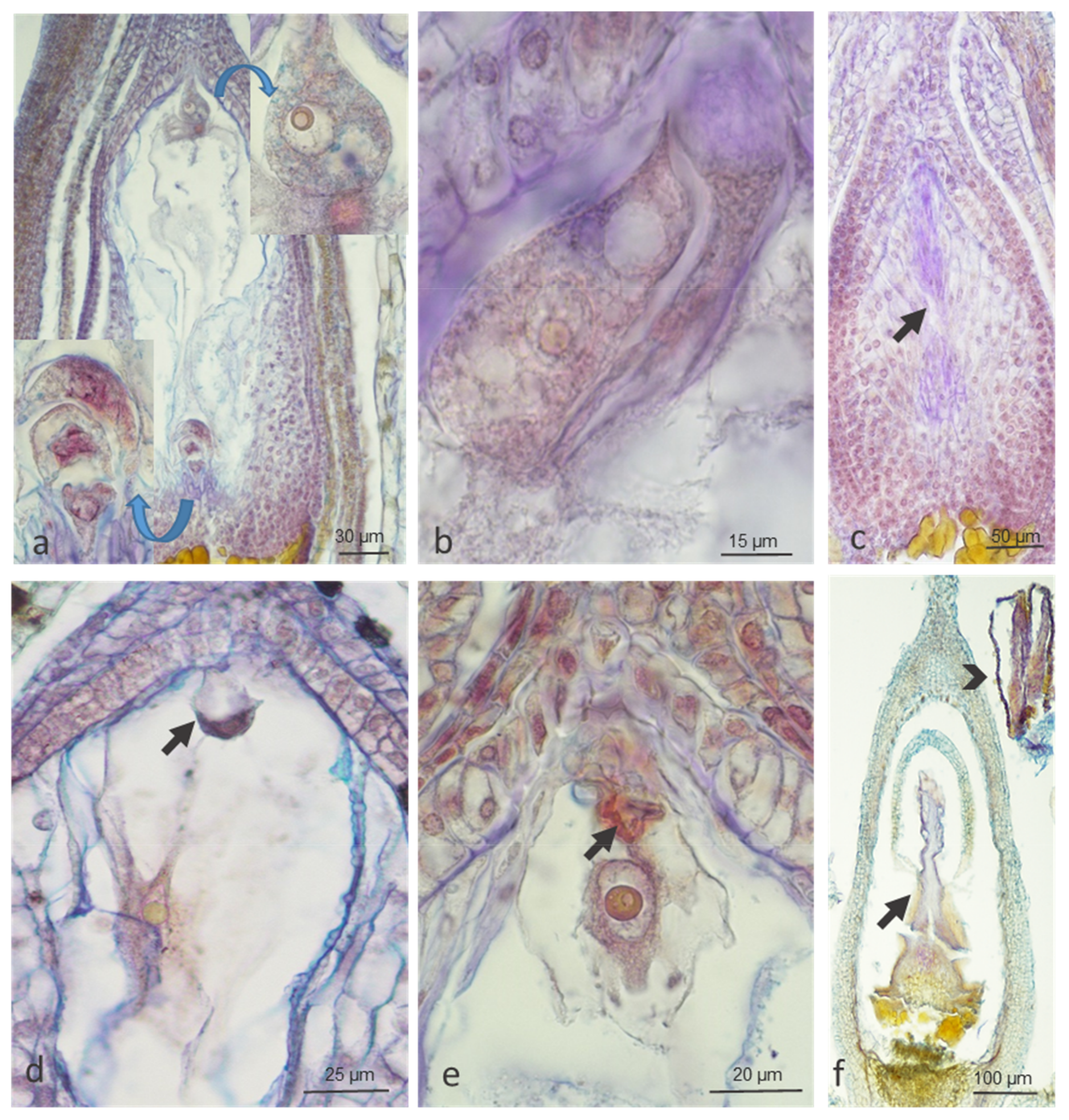
2.1.2. Ovule Development
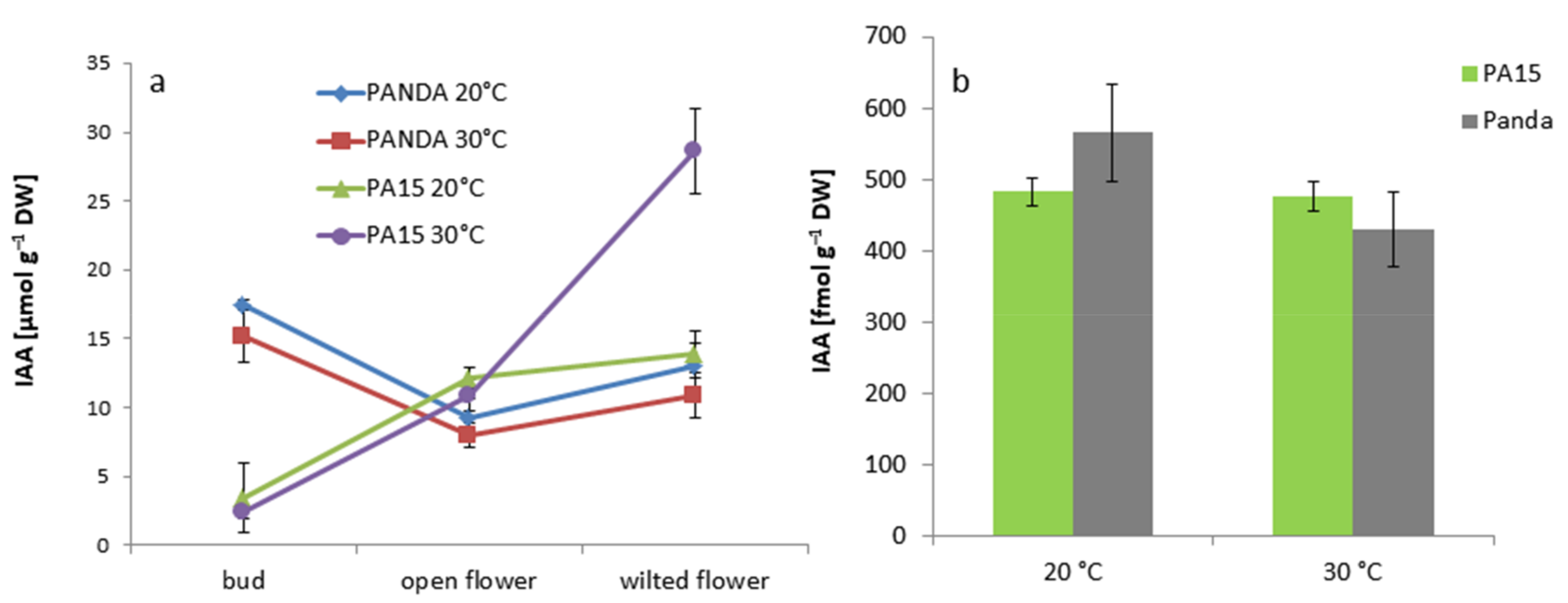
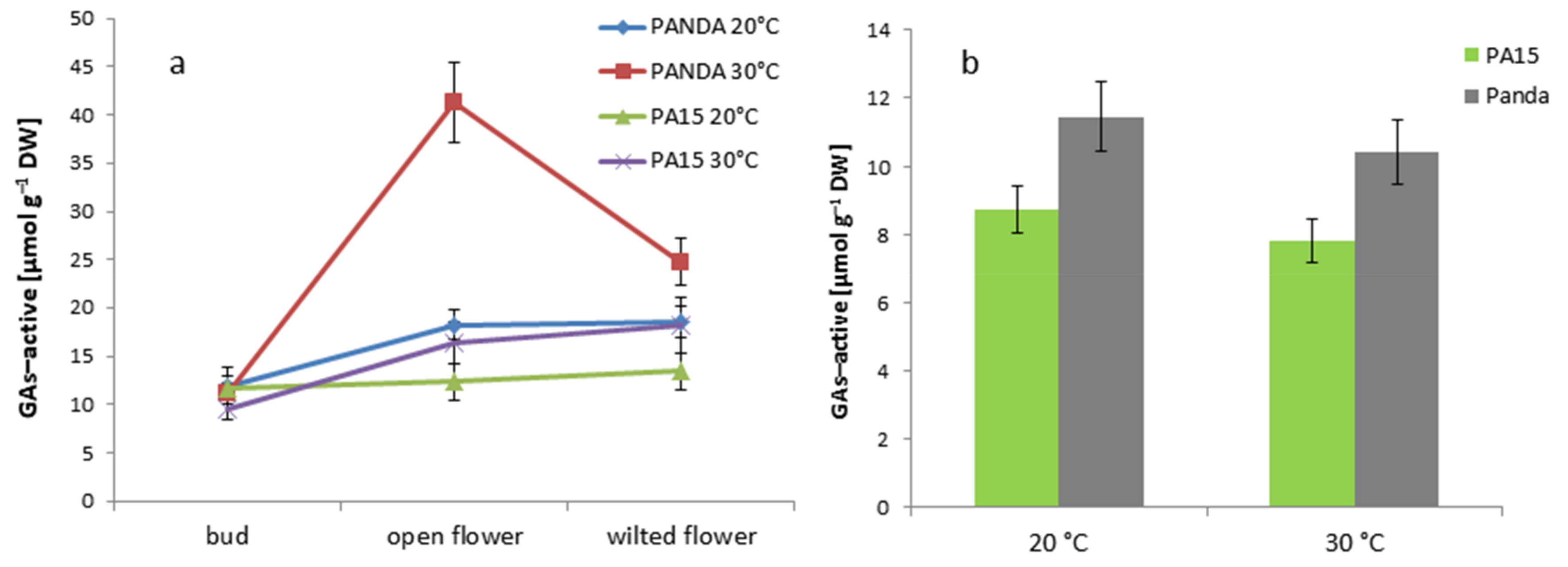
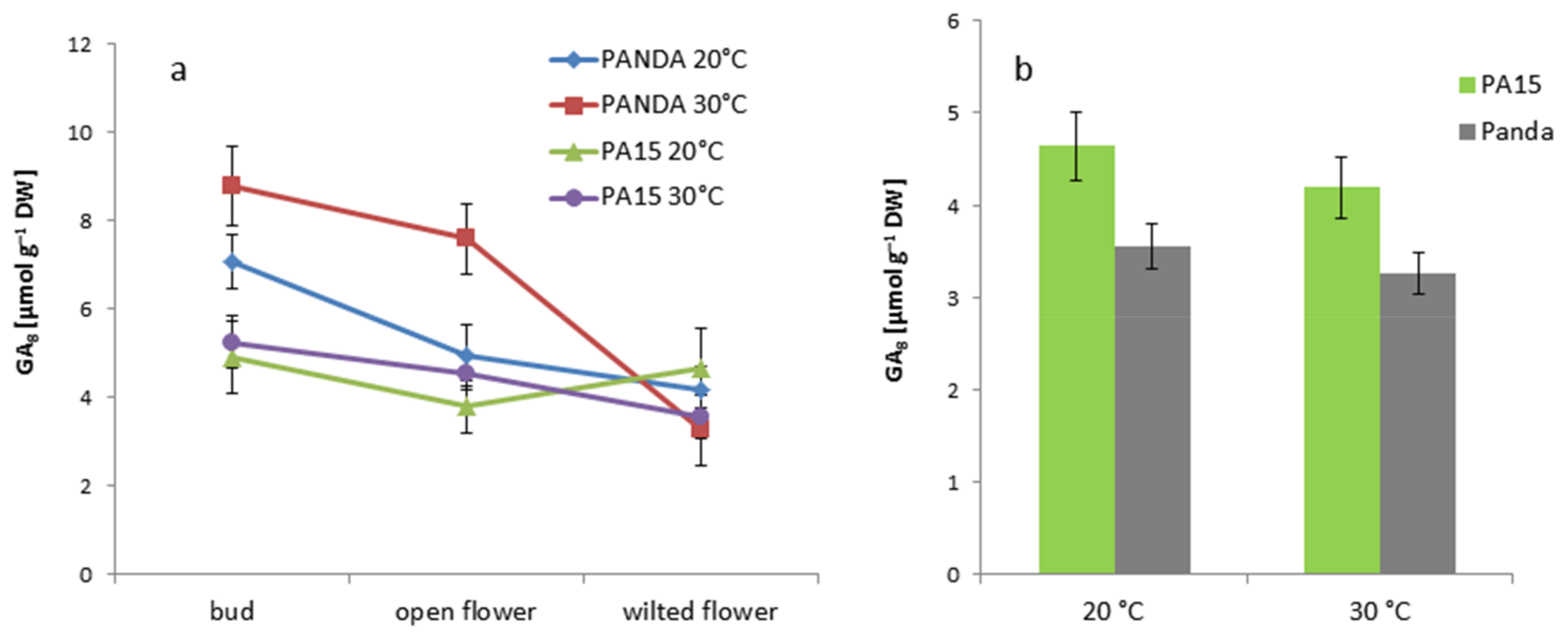
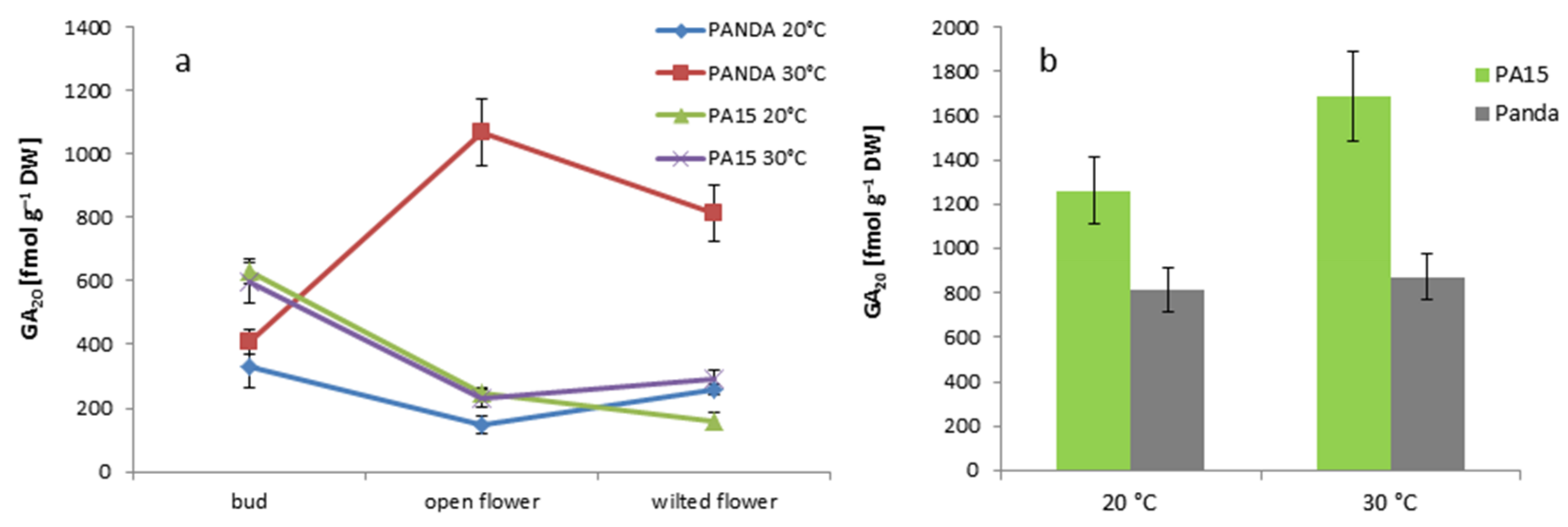
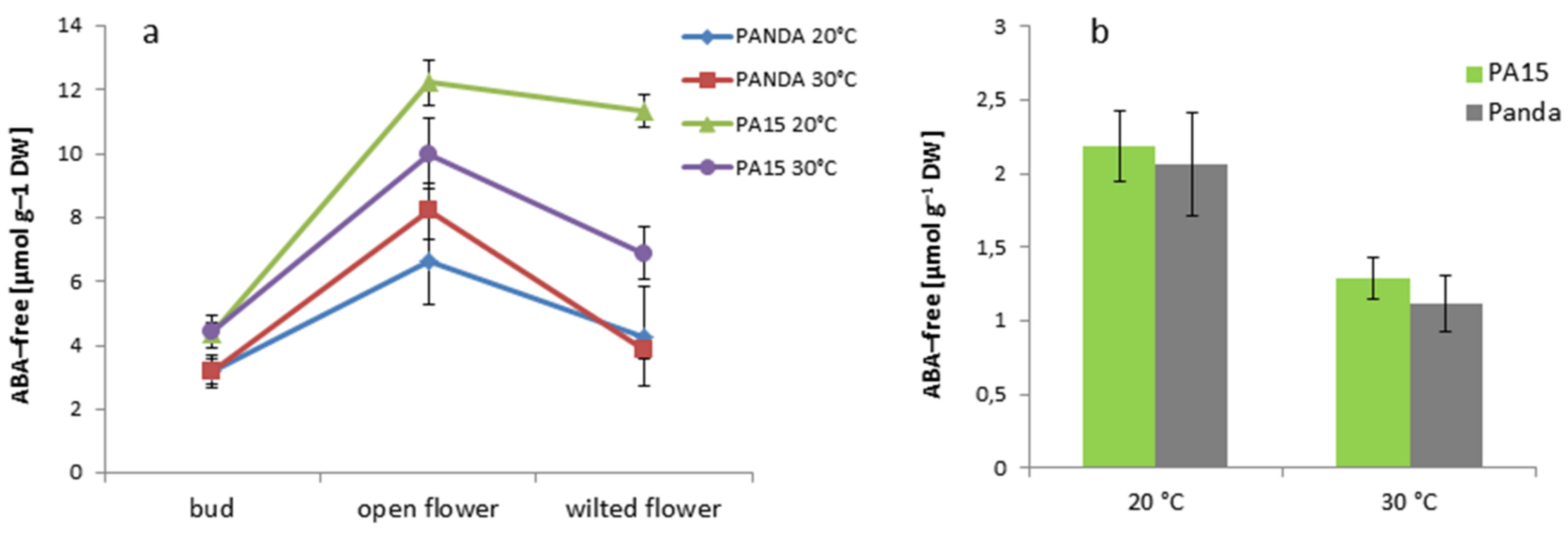
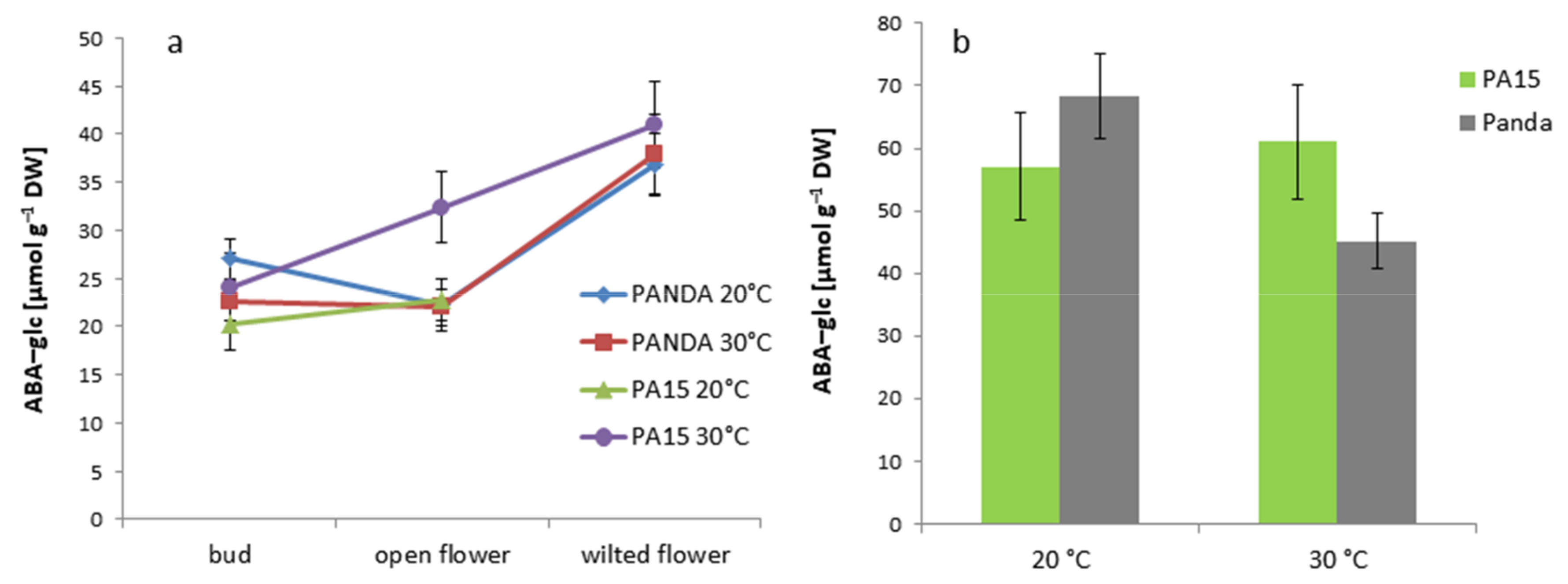
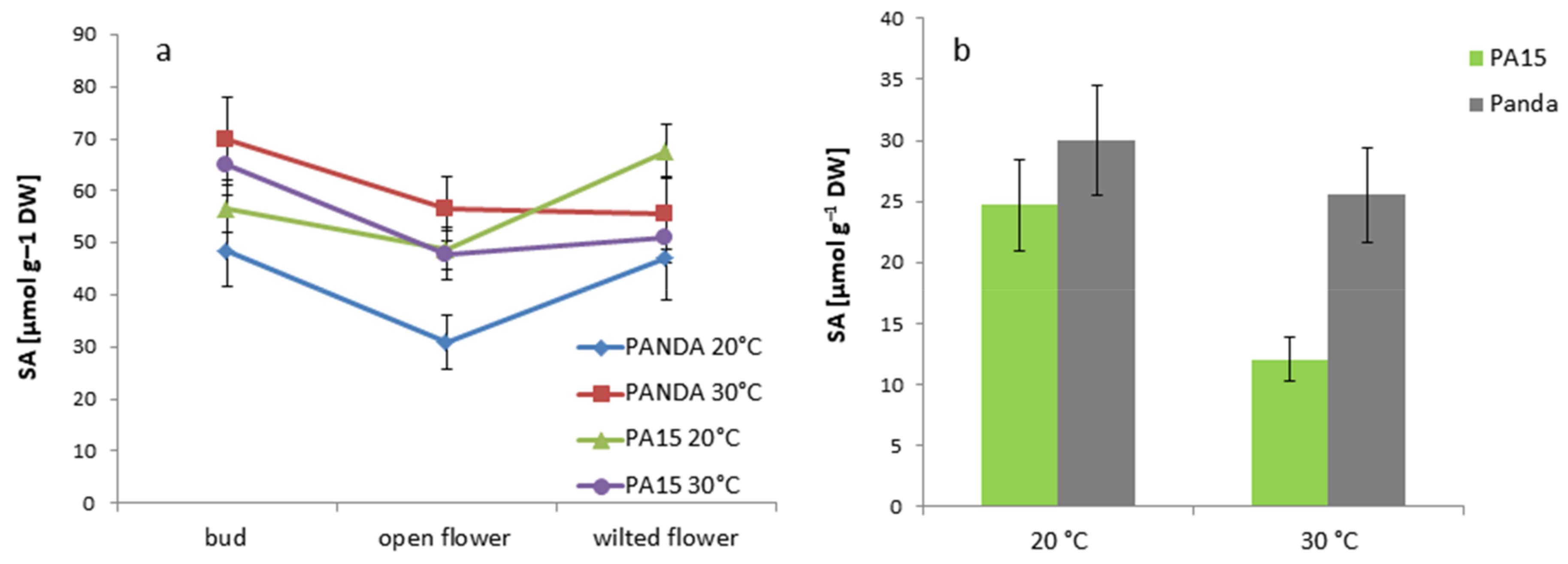
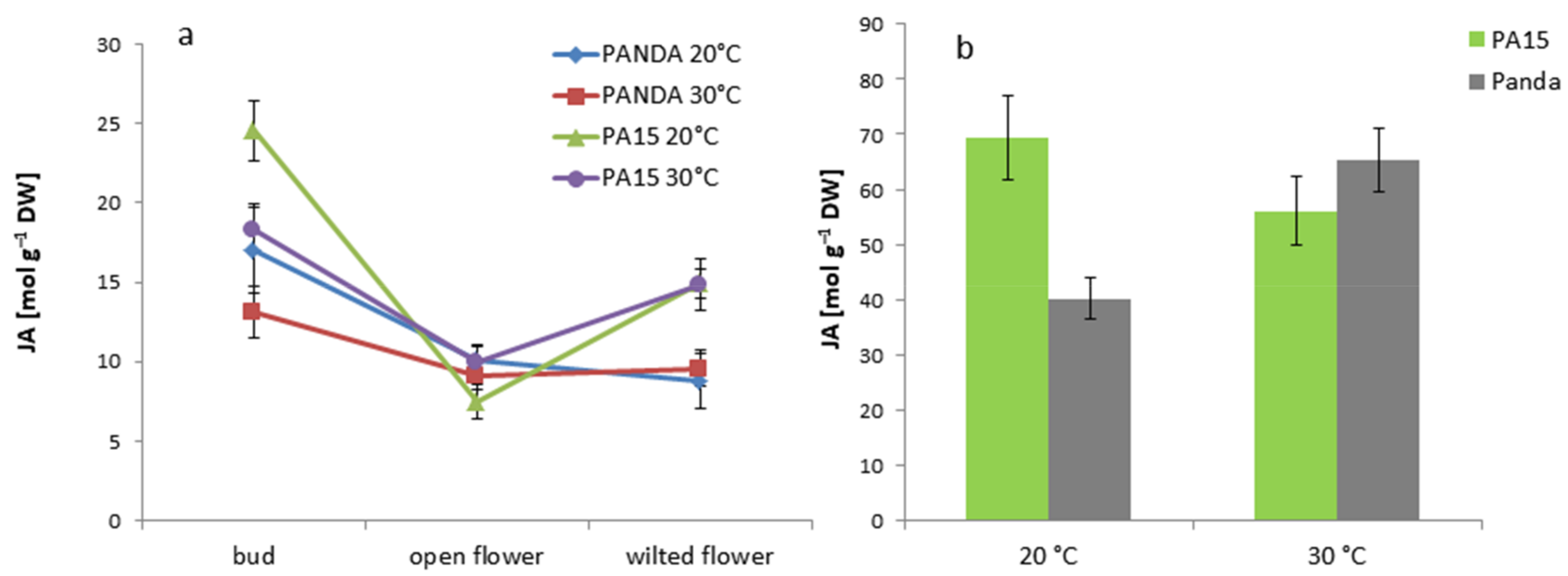
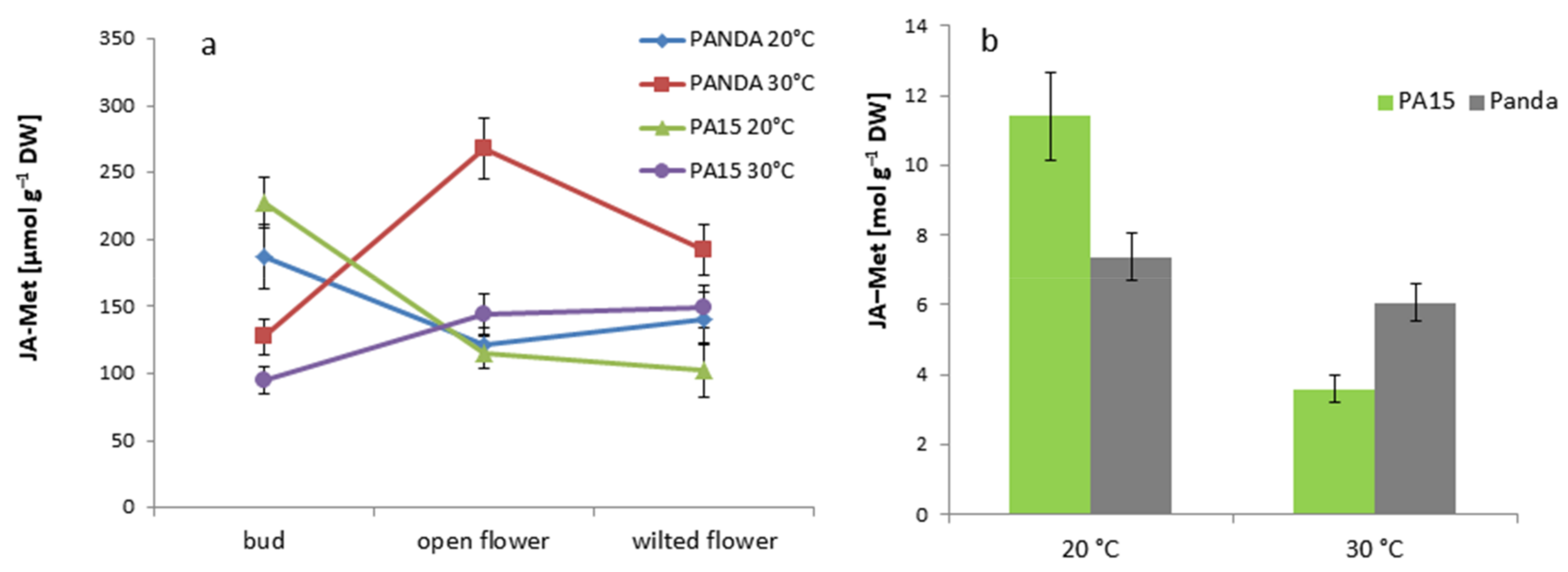
2.2. Hormonal Profile
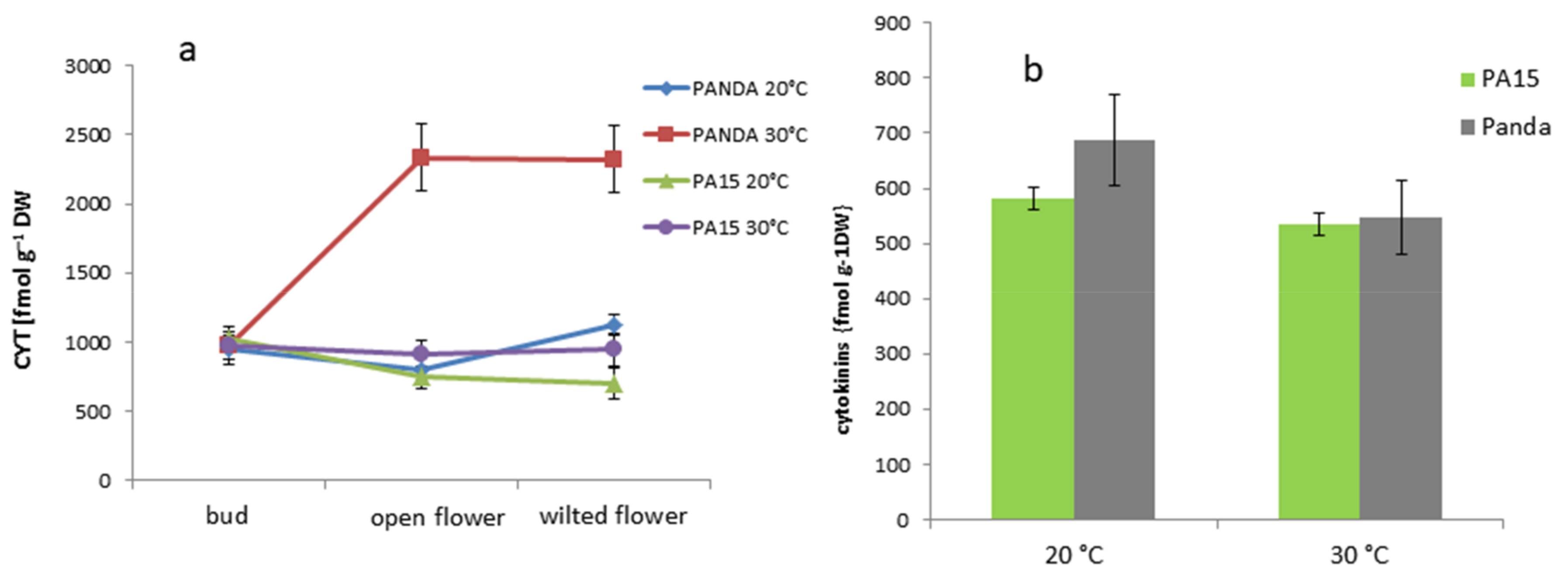
2.2.1. Total Hormone Content
2.2.2. Hormone Content in the Flowers and Leaves
2.2.3. Quantitative Ratio of the Studied Hormones
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Experimental Treatments
4.3. Measurements
4.3.1. Embryological Analyses
Pollen Viability
Ovule Development
4.3.2. Hormone Content Analysis
4.3.3. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ABA | abscisic acid |
| ABA-free | active form of abscisic acid |
| ABA-glc | abscisic acid glucosyl ester, non-active abscisic acid |
| CYT | cytokinins |
| GA | gibberellin |
| IAA | indole-3-acetic acid |
| JA | jasmonic acid |
| JA-Met | jasmonic acid methyl ester |
| SA | salicylic acid |
| ZEA | zeatin |
References
- Christa, K.; Soral-Śmietana, M. Buckwheat grains and buckwheat products—Nutritional and prophylactic value of their components—A review. Czech J. Food Sci. 2008, 26, 153–162. [Google Scholar] [CrossRef]
- Halbrecq, B.; Rommedenne, P.; Ledent, J.F. Evolution of flowering, ripening and seed set in buckwheat (Fagopyrum esculentum Moench.): Quantitative analysis. Eur. J. Agron. 2005, 23, 209–224. [Google Scholar] [CrossRef]
- Farooq, S.; Rehman, R.U.I.; Pirzadah, T.B.; Malik, B.; Ahmad Dar, F.; Tahir, I. Cultivation, agronomic practices, and growth performance of buckwheat. In Molecular Breeding and Nutritional Aspects of Buckwheat; Meilinag, Z., Ed.; Academic Press: Oxford, UK, 2016; pp. 299–320. [Google Scholar]
- Cawoy, V.; Ledent, J.-F.; Kinet, J.-M.; Jacquemart, A.-L. Floral biology of common buckwheat (Fagopyrum esculentum Moench. Eur. J. Plant Sci. Biotechnol. 2009, 3, 1–9. [Google Scholar]
- Adachi, T. How to combine the reproductive system with biotechnology in order to overcome the breeding barrier in buckwheat. Fagopyrum 1990, 10, 7–11. [Google Scholar]
- Taylor, D.P.; Obendorf, R.L. Quantitative assessment of some factors limiting seed set in buckwheat. Crop Sci. 2001, 41, 1792–1799. [Google Scholar]
- Cawoy, V.; Deblauwe, V.; Halbrecq, B.; Ledent, J.-F.; Kinet, J.-M.; Jacquemart, A.-L. Morph diffferences and honeybee morph preference in the distylous species Fagopyrum esculentum Moench. Int. J. Plant Sci. 2006, 167, 853–861. [Google Scholar] [CrossRef]
- Słomka, A.; Michno, K.; Dubert, F.; Dziurka, M.; Kopeć, P.; Płażek, A. Embryological background of low seed set in distylous common buckwheat (Fagopyrum esculentum Moench) with biased morph ratios, and biostimulant-induced improvement of it. Crop Past. Sci. 2017, 68, 680–690. [Google Scholar] [CrossRef]
- Slawinska, J.; Obendorf, R.L. Buckwheat seed set in planta and during in vitro inflorescence culture: Evaluation of temperature and water deficit stress. Seed Sci. Res. 2001, 11, 223–233. [Google Scholar]
- Cawoy, V.; Lutts, S.; Ledent, J.-F.; Kinet, J.-M. Resource availability regulates reproductive meristem activity, developmental of reproductive structures and seed set in buckwheat (Fagopyrum esculentum). Physiol. Plant. 2007, 131, 341–453. [Google Scholar] [PubMed]
- Kreft, I. Buckwheat breeding prospectives. In Proceedings of the II International Symposium Buckwheat, Miyazaki, Japan, 7–10 September 1983; pp. 3–12. [Google Scholar]
- Guan, L.M.; Adachi, T. Ultrastructural changes of the mature embryo sac in buckwheat (Fagopyrum esculentum) as a result of high temperature exposure. Cytologia 1994, 59, 237–248. [Google Scholar] [CrossRef]
- Kumar, S.; Thakur, P.; Kaushal, N.; Malik, J.A.; Gaur, P.; Nayyar, H. Effect of varying high temperatures during reproductive growth on reproductive function, oxidative stress and seed yield in chickpea genotypes differing in heat sensitivity. Arch. Agron. Soil Sci. 2013, 59, 823–843. [Google Scholar] [CrossRef]
- Zinn, K.E.; Tunc-Ozdemir, M.; Harper, J.F. Temperature stress and plant sexual reproduction: Uncovering the weakest links. J. Exp. Bot. 2010, 67, 1959–1968. [Google Scholar] [CrossRef]
- Thingreingam, I.K.S.; Mitra, S.K. Understanding the pollen and ovule characters and fruit set of fruit crops in relation to temperature and genotype—A review. J. Appl. Bot. Food Qual. 2014, 87, 157–167. [Google Scholar]
- Bernier, G.; Havelange, A.; Houssa, C.; Petitjean, A.; Lejeune, P. Physiological signals that induce flowering. Plant Cell 1993, 5, 1147–1155. [Google Scholar] [CrossRef]
- McGregor, D.I. Pattern of flower and pod development in rapeseed. Can. J. Plant Sci. 1981, 61, 275–282. [Google Scholar] [CrossRef]
- Aufhammer, W.; Gotz, I.; Peter, M. Yield performance of field bean (Viciafaba L.) in relation to interactio between inflorescence at different nodes. J. Agric. Sci. 1987, 108, 479–486. [Google Scholar] [CrossRef]
- Patric, J.W. Are hormones involved in assimilate transport? In Hormone Action in Plant Development: A Critical Appraisal; Hoad, G.V., Lenton, J.R., Jackson, M.B., Actin, P.K., Eds.; Butterworths: London, UK, 1987; pp. 175–187. [Google Scholar]
- De Jong, T.J.; Klinkhamer, P.G.L. Evolutionary Ecology of Plant Reproductive Strategies; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2005. [Google Scholar]
- Moe, R. Factors Affecting Flower Abortion and Malformation in Roses. Physiol. Plant. 1971, 24, 291–300. [Google Scholar] [CrossRef]
- Wang, Y.H.; Irving, H.R. Developing a model of plant hormone interactions. Plant Signal. Behav. 2011, 6, 494–500. [Google Scholar] [CrossRef]
- Beppu, K.; Ikeda, T.; Kataoke, I. Effect of high temperature exposure time during flower bud formation on the occurrence of double pistils in ‘Satohnishiki’ sweet cherry. Sci. Hort. 2001, 87, 77–84. [Google Scholar] [CrossRef]
- Seo, H.S.; Song, J.T.; Cheong, J.-J.; Lee, Y.-H.; Lee, Y.-W.; Hwang, I.; Lee, J.S.; Choi, Y.D. Jasmonic acid carboxyl methyltransferase: A key enzyme for jasmonate-regulated plant responses. Proc. Natl. Acad. Sci. USA 2001, 98, 4788–4793. [Google Scholar] [CrossRef] [PubMed]
- Guan, L.M.; Adachi, T. Ultrastructural change of organelle and cytoplasmic DNA deterioration during embryogenesis in Fagopyrum esculentum. In Current Advances in Buckwheat Research; Shinshu University Press: Matsumoto, Japan, 1995; pp. 247–253. [Google Scholar]
- Guan, L.; Chen, L.; Adachi, T. Ultrastructural studies of embryo abortion in buckwheat (Fagopyrum esculentum) as a heat-stress. Cytologia 2008, 73, 371–379. [Google Scholar] [CrossRef]
- Hedden, P.; Thomas, S.G. Gibberellin biosynthesis and its regulation. Biochem. J. 2012, 444, 11–25. [Google Scholar] [CrossRef]
- Halińska, A.; Lewak, S. Free and conjugated gibberellins in dormancy and germination of apple seeds. Physiol. Plant. 1987, 69, 523–530. [Google Scholar] [CrossRef]
- Chien, C.T.; Kuo-Huang, L.; Lin, T.P. Changes in ultrastructure and abscisic acid level, and response to applied gibberellins in Taxusmairei seeds treated with warm and cold stratification. Ann. Bot. 1998, 81, 41–47. [Google Scholar] [CrossRef]
- Shu, K.; Chen, Q.; Wu, Y.; Liu, R.; Zhang, H.; Wang, S.; Tang, S.; Yang, W.; Xie, Q. Abscisic Acid-Insensitive 4 negatively regulates flowering through directly promoting Arabidopsis FLOWERING LOCUS C transcription. J. Exp. Bot. 2016, 67, 195–205. [Google Scholar] [CrossRef] [PubMed]
- Riboni, M.; Test, A.R.; Galbiati, M.; Tonelli, C.; Conti, L. ABA-dependent control of GIGANTEA signalling enables drought escape via up-regulation of FLOWERING LOCUS T in Arabidopsis thaliana. J. Exp. Bot. 2016, 67, 6309–6322. [Google Scholar] [CrossRef]
- Aneja, M.; Gianfagna, T.; Ng, E. The roles of abscisic acid and ethylene in the abscission and senescence of Cocoa flowers. Plant Growth Reg. 1999, 27, 149–155. [Google Scholar] [CrossRef]
- Ferrante, A.; Vernieri, P.; Tognoni, F.; Serra, G. Changes in abscisic acid and flower pigments during floral senescence of Petunia. Biol. Plant. 2006, 50, 581–585. [Google Scholar] [CrossRef]
- Panavas, T.; Rubinstein, B.; Walker, E.L. Possible involvement of abscisic acid in senescence of daylily petals. J. Exp. Bot. 1998, 49, 198–1997. [Google Scholar] [CrossRef]
- Reid, M.S. The role of ethylene in flower senescence. Acta Hort. 1989, 261, 156–170. [Google Scholar] [CrossRef]
- Zhong, Y.; Ciafré, C. Role of ABA in ethylene-independent Iris flower senescence. In Proceedings of the International Conference on Food Engineering and Biotechnology, Bangkok, Thailand, 7–9 May 2011; IACSIT Press: Singapore, 2011; pp. 261–266. [Google Scholar]
- Shu, K.; Zhou, W.; Chen, F.; Luo, X.; Yang, W. Abscisic acid and gibberellins antagonistically mediate plant development and abiotic stress responses. Front. Plant Sci. 2018, 9, 416. [Google Scholar] [CrossRef] [PubMed]
- Miransari, M.; Smith, D.L. Plant hormones and seed germination. Environ. Exp. Bot. 2014, 99, 110–121. [Google Scholar] [CrossRef]
- Franklin, K.A.; Lee, S.H.; Patel, D.; Vinod Kumar, S.; Spartz, A.K.; Gu, C.; Ye, S.; Yu, P.; Breen, G.; Cohen, J.D.; et al. Phytochrome-Interacting Factor 4 (PIF4) regulates auxin biosynthesis at high temperature. Proc. Natl. Acad. Sci. USA 2011, 108, 20231–20235. [Google Scholar] [CrossRef] [PubMed]
- Stavang, J.A.; Gallego-Bartolomé, J.; Gómez, M.D.; Yoshida, S.; Asami, T.; Olsen, J.E.; García-Martínez, J.L.; Alabadí, D.; Blázquez, M.A. Hormonal regulation of temperature-induced growth in Arabidopsis. Plant J. 2009, 60, 589–601. [Google Scholar] [CrossRef] [PubMed]
- Nordström, A.; Tarkowski, P.; Tarkowska, D.; Norbaek, R.; Åstot, C.; Dolezal, K.; Sandberg, G. Auxin regulation of cytokinin biosynthesis in Arabidopsis thaliana: A factor of potential importance for auxin–cytokinin-regulated development. Proc. Natl. Acad. Sci. USA 2004, 101, 8039–8044. [Google Scholar] [CrossRef] [PubMed]
- Dafni, A.; Firmage, D. Pollen viability and longevity: Practical, ecological and evolutionary implications. Plant Syst. Evol. 2000, 222, 113–132. [Google Scholar] [CrossRef]
- Singh, R.J. Plant Cytogenetics, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2003. [Google Scholar]
- Gerlach, D. Zarys Mikrotechniki Botanicznej; Państwowe Wydawnictwo Rolnicze i Leśne: Warszawa, Poland, 1972. (In Polish) [Google Scholar]
- Płażek, A.; Dubert, F.; Kopeć, P.; Dziurka, M.; Kalandyk, A.; Pastuszak, J.; Wolko, B. Seed hydropriming and smoke water significantly improve low-temperature germination of Lupinusangustifolius L. Int. J. Mol. Sci. 2018, 19, 992. [Google Scholar] [CrossRef]
- Dziurka, M.; Janeczko, A.; Juhász, C.; Gullner, G.; Oklestková, J.; Novák, O.; Saja, D.; Skoczowski, A.; Tóbiász, J.; Barna, B. Local and systemic hormonal responses in pepper leaves during compatible and incompatible pepper-tobamovirus interactions. Plant Physiol. Biochem. 2016, 109, 355–364. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Organ | cv. ‘Panda’ | PA15 | ||
|---|---|---|---|---|
| 20 °C | 30 °C | 20 °C | 30 °C | |
| Buds | 100.0 | 57.7 * | 83.3 | 79.5 |
| Open flowers | 88.2 | 44.4 * | 77.8 | 26.7 * |
| Wilted flowers | 83.3 | 80.0 | 81.3 | 78.9 |
| Hormone | Buds | Open Flowers | Wilted Flowers | Leaves |
|---|---|---|---|---|
| CYT | 0.98 ± 0.08 b | 1.20 ± 0.11 a | 1.27 ± 0.12 a | 0.59 ± 0.06 c |
| IAA | 9.63 ± 1.72 b | 10.07 ± 1.51 b | 16.63 ± 2.50 a | 0.49 ± 0.07 c |
| GAs-active | 11.08 ± 0.99 c | 22.10 ± 2.32 a | 18.74 ± 2.06 b | 10.76 ± 1.61 c |
| GA8 | 2.39 ± 1.15 b | 6.45 ± 0.97 a | 5.22 ± 0.78 a | 1.16 ± 0.17 c |
| GA9 | 6.50 ± 0.98 c | 13.86 ± 1.52 a | 10.27 ± 1.02 b | 6.73 ± 0.84 c |
| GA20 | 0.49 ± 0.06 a | 0.42 ± 0.06 a | 0.31 ± 0.05 b | 3.91 ± 0.99 ab |
| ABA-free | 3.71 ± 0.53 c | 9.27 ± 1.39 a | 6.60 ± 0.99 b | 1.66 ± 0.25 d |
| ABA-glc | 23.51 ± 3.90 c | 24.97 ± 3.75 c | 36.27 ± 5.44 b | 57.91 ± 8.69 a |
| SA | 59.60 ± 6.88 a | 46.04 ± 6.91 a | 55.34 ± 8.30 a | 23.06 ± 3.46 b |
| JA | 19.21 ± 2.89 b | 9.41 ± 1.41 c | 11.30 ± 1.70 c | 57.76 ± 8.70 a |
| JA-Met | 0.16 ± 0.01 b | 0.16 ± 0.02 b | 0.15 ± 0.02 b | 7.11 ± 1.07 a |
| Active Gibberellin | Buds and Flowers | Leaves | ||
|---|---|---|---|---|
| cv. ‘Panda’ | Strain PA15 | cv. ‘Panda’ | Strain PA15 | |
| GA1 | 30.6 ± 4.7 a | 24.3 ± 3.6 a | 16.7 ± 2.0 b | 18.7 ± 2.0 b |
| GA3 | 18.3 ± 2.7 a | 17.1 ± 2.6 a | 20.7 ± 2.4 a | 17.2 ± 2.0 a |
| GA4 | 5.1 ± 0.5 a | 4.5 ± 0.5 b | 5.7 ± 0.7 a | 4.7 ± 0.5 b |
| GA5 | 8.6 ± 1.0 c | 8.3 ± 1.0 c | 11.3 ± 1.3 bc | 15.1 ± 1.8 a |
| GA6 | 36.6 ± 5.3 b | 44.4 ± 7.7 a | 43.6 ± 5.2 a | 42.5 ± 5.1 a |
| GA7 | 0.9 ± 0.1 b | 1.5 ± 0.1 b | 2.1 ± 0.2 ab | 1.8 ± 0.2 a |
| Strain/Cultivar | Temp. | Organ | |||
|---|---|---|---|---|---|
| Buds | Open Flowers | Wilted Flowers | Leaves | ||
| GAs/ABA-free | |||||
| PA15 | 20 | 2.77 ± 0.33 bB | 1.12 ± 1.00 cC | 1.19 ± 0.09 dC | 4.57 ± 0.51 bA |
| 30 | 3.56 ± 0.42 aC | 3.08 ± 0.27 bC | 4.44 ± 0.49 bB | 9.19 ± 1.09 aA | |
| ‘Panda’ | 20 | 2.15 ± 0.25 cB | 1.64 ± 0.15 cC | 2.69 ± 0.24 cB | 5.86 ± 0.65 bA |
| 30 | 3.77 ± 0.45 aC | 6.68 ± 0.62 aB | 6.52 ± 0.71 aB | 10.82 ± 1.18 aA | |
| IAA/ABA-free | |||||
| PA15 | 20 | 0.82 ± 0.09 bB | 1.10 ± 0.08 bB | 2.52 ± 0.28 cA | 0.22 ± 0.02 bA |
| 30 | 5.55 ± 0.66 aA | 1.56 ± 0.09 aC | 3.07 ± 0.39 bB | 0.45 ± 0.05 aD | |
| ‘Panda’ | 20 | 0.59 ± 0.07 cC | 1.08 ± 0.08 bB | 3.94 ± 0.41 aA | 0.28 ± 0.03 bD |
| 30 | 5.05 ± 0.49 aA | 1.07 ± 0.08 bD | 3.03 ± 0.29 bC | 0.41 ± 0.05 aB | |
| IAA/CYT | |||||
| PA15 | 20 | 3.29 ± 0.34 cB | 17.19 ± 1.72 aA | 19.89 ± 1.97 bA | 0.87 ± 0.08 aC |
| 30 | 18.02 ± 2.1 aA | 10.29 ± 1.08 cC | 13.94 ± 1.45 cB | 0.92 ± 0.09 aD | |
| ‘Panda’ | 20 | 2.65 ± 2.58 dC | 13.91 ± 1.35 bB | 24.77 ± 2.67 aA | 0.87 ± 0.09 aD |
| 30 | 15.75 ± 1.74 bA | 4.94 ± 0.52 dC | 6.92 ± 0.68 dB | 0.92 ± 0.09 aD | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Płażek, A.; Słomka, A.; Kopeć, P.; Dziurka, M.; Hornyák, M.; Sychta, K.; Pastuszak, J.; Dubert, F. Effects of High Temperature on Embryological Development and Hormone Profile in Flowers and Leaves of Common Buckwheat (Fagopyrum esculentum Moench). Int. J. Mol. Sci. 2019, 20, 1705. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20071705
Płażek A, Słomka A, Kopeć P, Dziurka M, Hornyák M, Sychta K, Pastuszak J, Dubert F. Effects of High Temperature on Embryological Development and Hormone Profile in Flowers and Leaves of Common Buckwheat (Fagopyrum esculentum Moench). International Journal of Molecular Sciences. 2019; 20(7):1705. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20071705
Chicago/Turabian StylePłażek, Agnieszka, Aneta Słomka, Przemysław Kopeć, Michał Dziurka, Marta Hornyák, Klaudia Sychta, Jakub Pastuszak, and Franciszek Dubert. 2019. "Effects of High Temperature on Embryological Development and Hormone Profile in Flowers and Leaves of Common Buckwheat (Fagopyrum esculentum Moench)" International Journal of Molecular Sciences 20, no. 7: 1705. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20071705