Extracellular Proteolytic Activity and Amino Acid Production by Lactic Acid Bacteria Isolated from Malaysian Foods

 , , ,
, , ,
Abstract
:1. Introduction
2. Results and Discussion
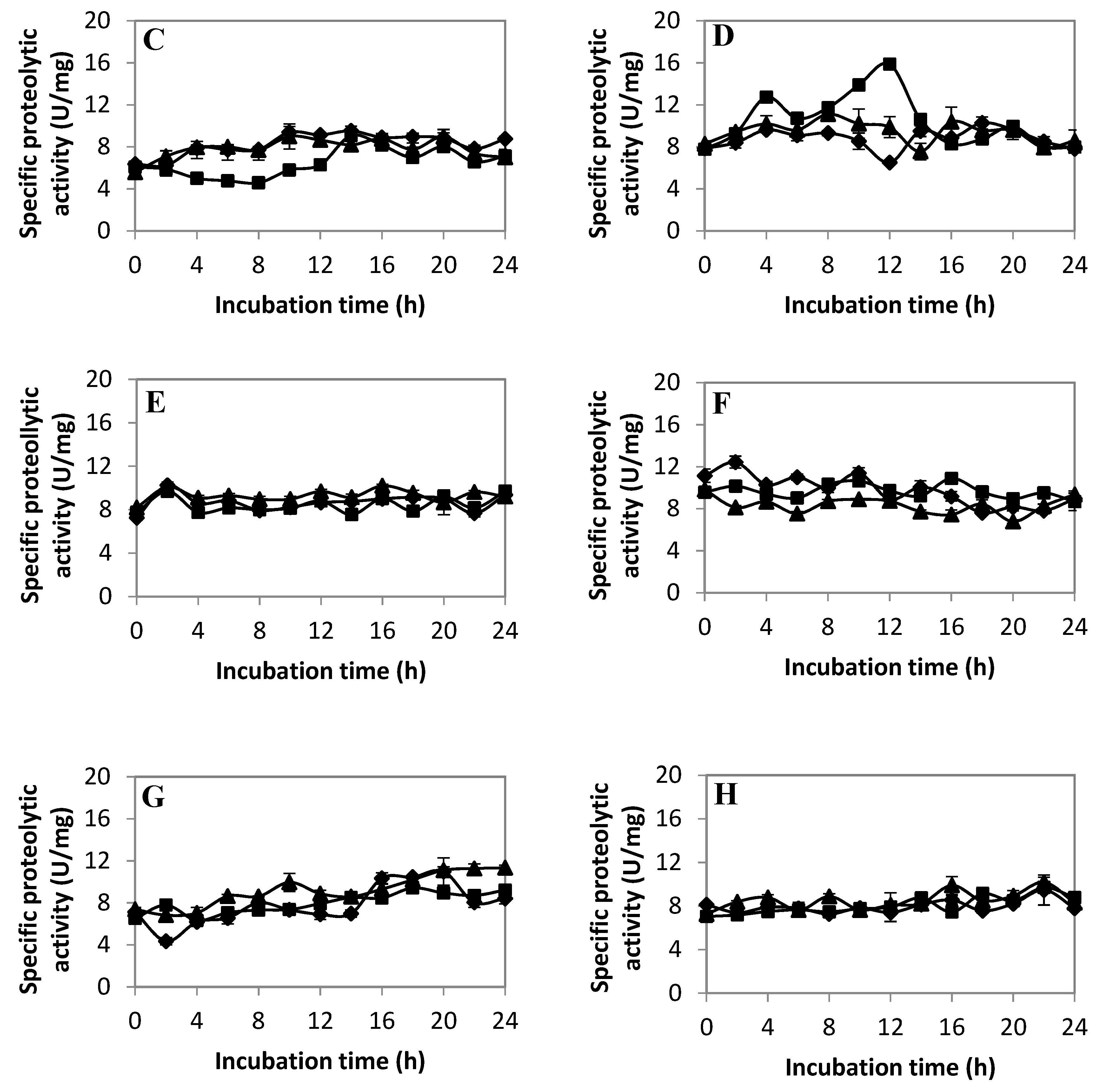
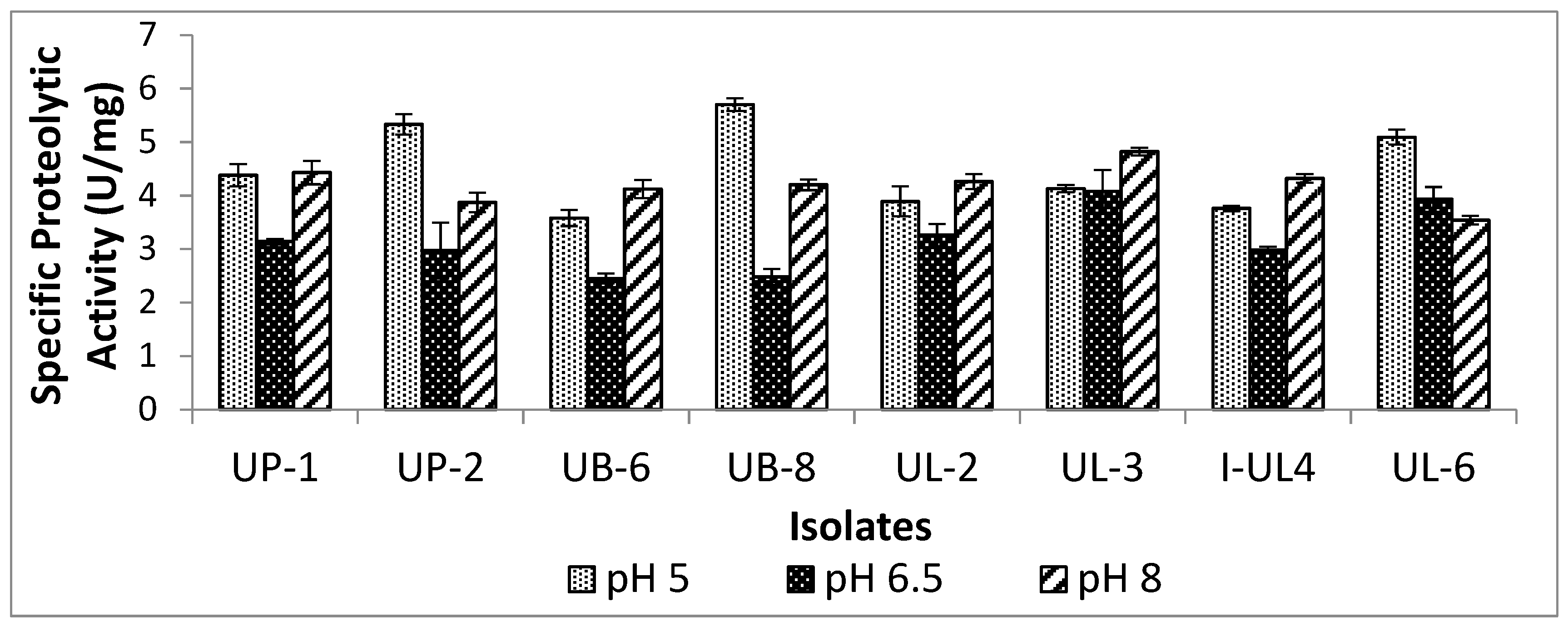
2.1. Proteolytic Activity of Lactic Acid Bacteria (LAB)
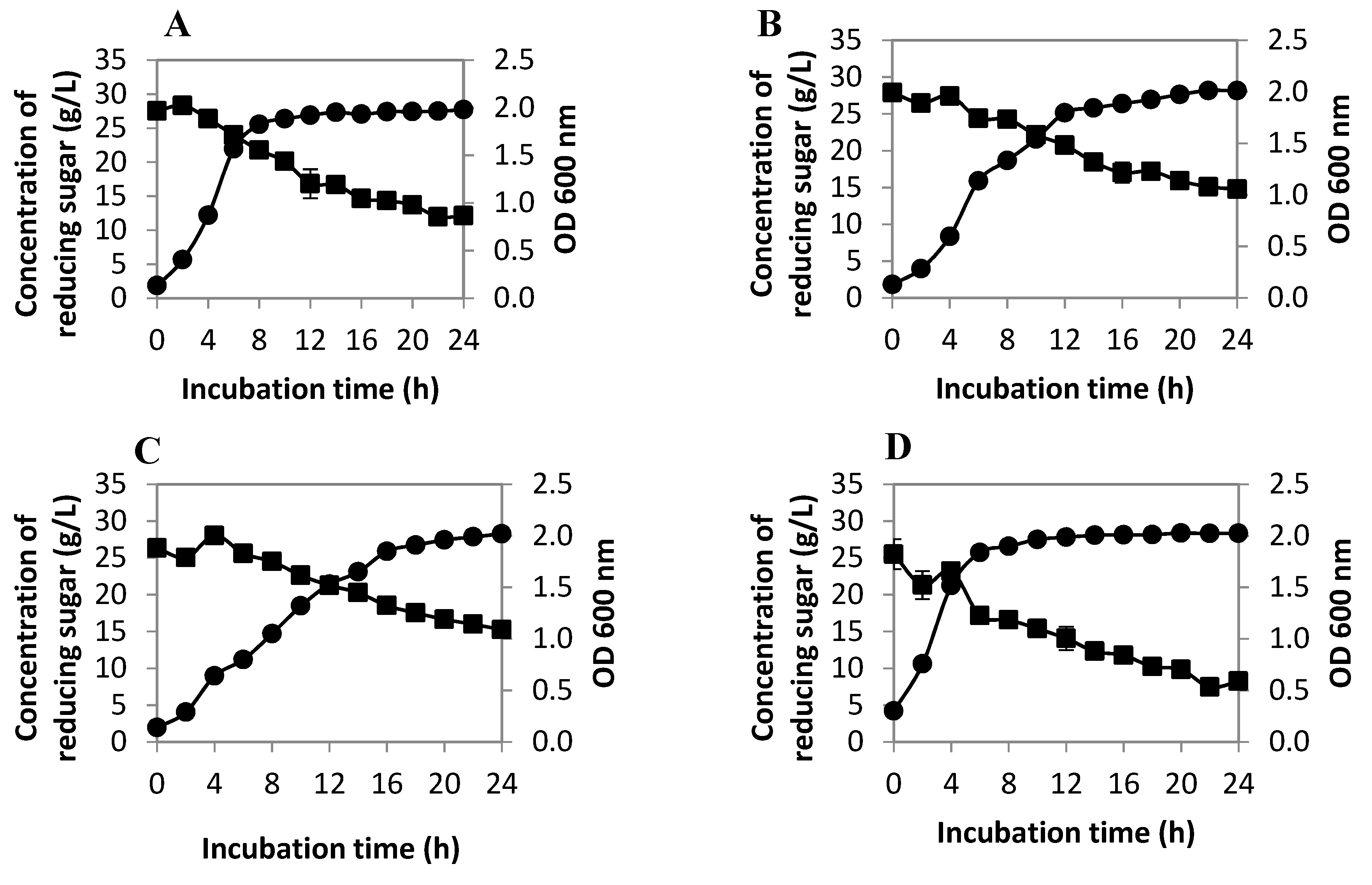
2.2. Production of Amino Acids
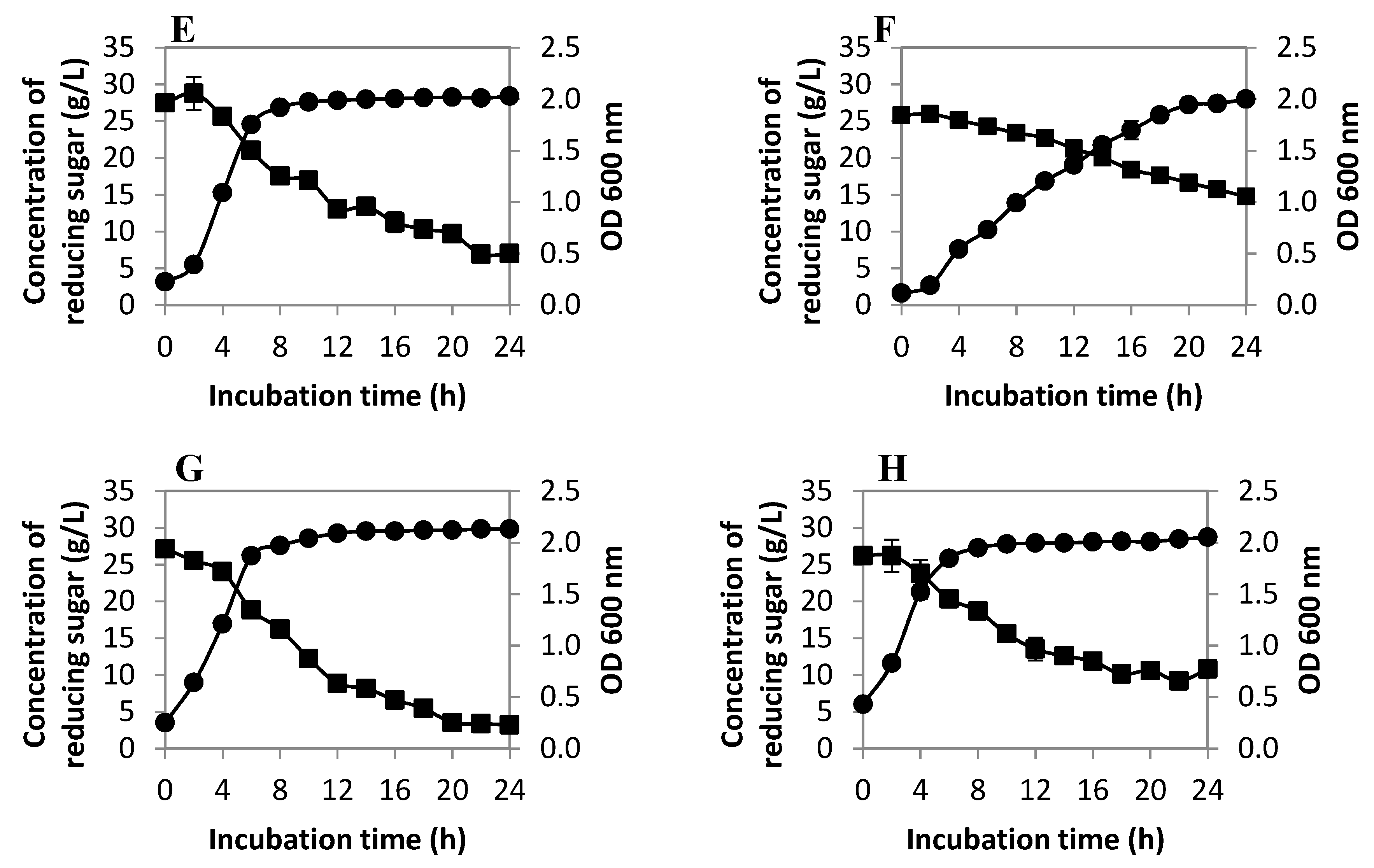
2.2.1. Cell Population and Reducing Sugar Utilization
2.2.2. Amino Acid Production Profile
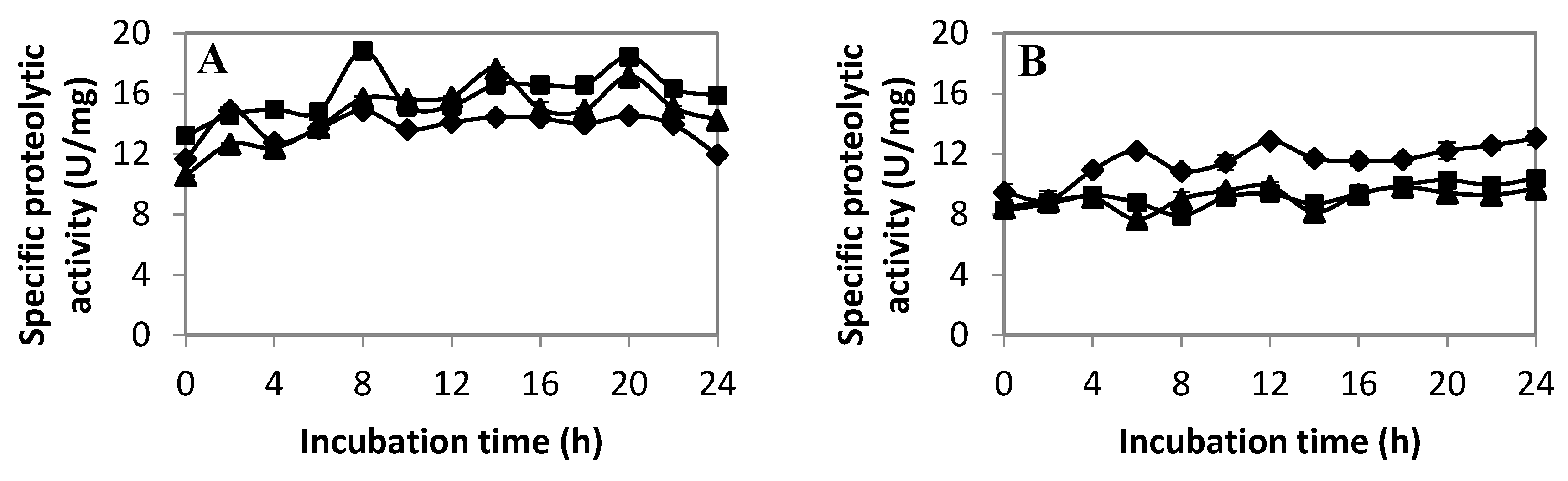
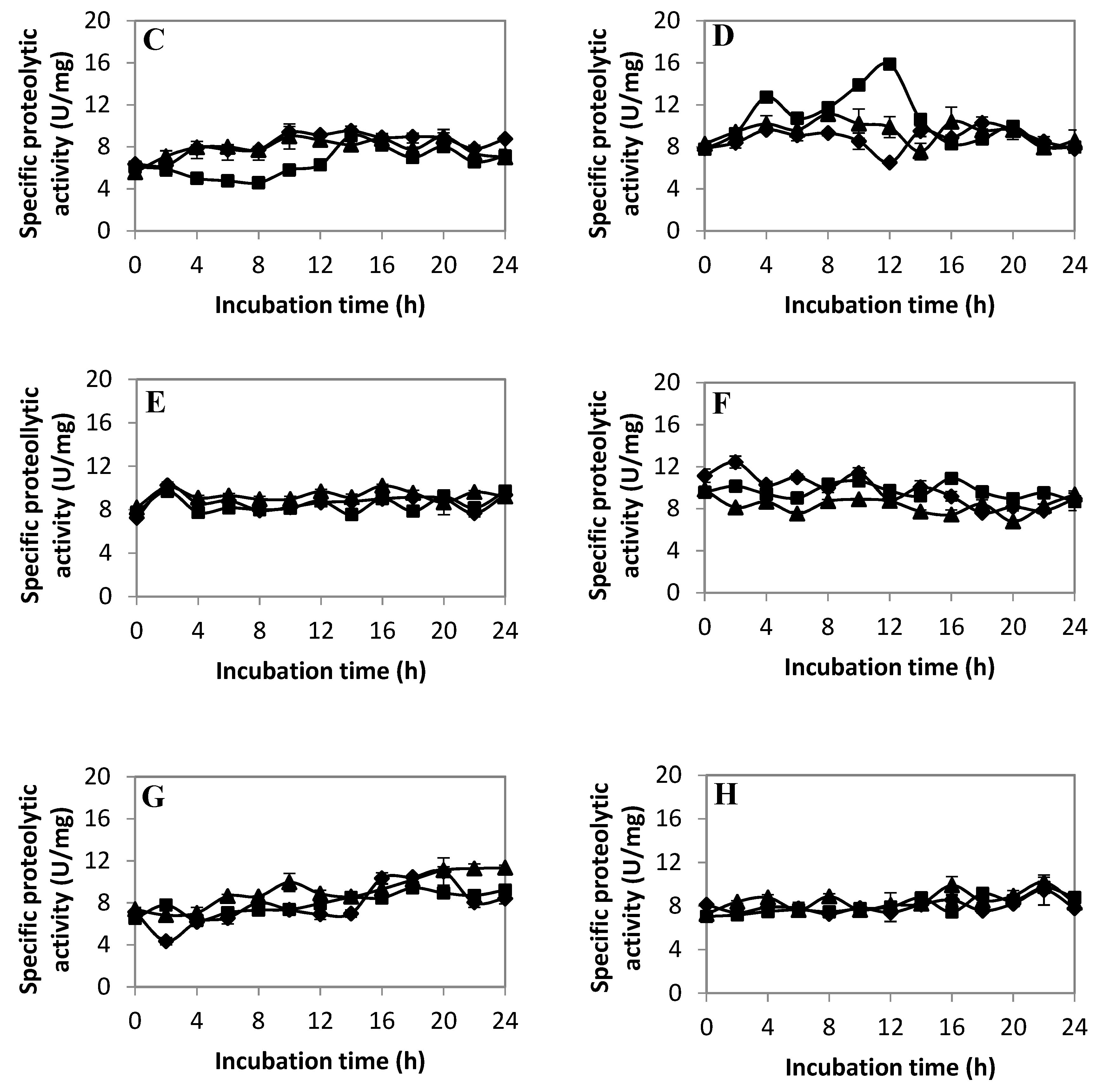
2.2.3. Extracellular Proteolytic Activity Profile of LAB
3. Materials and Methods
3.1. Bacterial Growth Conditions
3.2. Effect of pH on the Extracellular Proteolytic Activity of LAB
3.3. Determination of Proteolytic Activity
3.4. Determination of Solubilised Protein
3.5. Determination of the Amino Acid Production Profile of LAB
3.5.1. Determination of Cell Population
3.5.2. Determination of Residual Reducing Sugar
3.5.3. Determination of Amino Acids
3.6. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AA | Amino acid |
| LAB | Lactic acid bacteria |
| GRAS | Generally recognized as safe |
| MRS | de Man, Rogosa and Sharpe |
| TCA | Trichloroacetic acid |
| OD600nm | Optical density at 600 nm |
| DNS | Dinitrosalicylic acid |
| OPA | o-phthalaldehyde |
| FMOC | 9-fluorenylmethyl chloroformate |
| ANOVA | Analysis of variance |
References
- Andrea, B.; Pierandrea, T. Nuclear magnetic resonance of amino acids, peptides, and proteins. In Amino Acids, Peptides and Proteins in Organic Chemistry, Analysis and Function of Amino Acids and Peptides; Hughes, A.B., Ed.; John Wiley and Sons, Inc.: Weinheim, Germany, 2012; Volume 5, pp. 97–153. ISBN 978-3-527-32104-9. [Google Scholar]
- Jakubke, H.D.; Sewald, N. Peptides from A to Z: A Concise Encyclopedia; WILEY-VCH Verlag GmBH & Co. KGaA: Weinheim, Germany, 2008; ISBN 978-3-527-62117-0. [Google Scholar]
- Wu, G. Functional amino acids in growth, reproduction, and health. Adv. Nutr. 2010, 1, 31–37. [Google Scholar] [CrossRef]
- Boisen, S.; Hvelplund, T.; Weisbjerg, M. Ideal amino acid profiles as a basis for feed protein evaluation. Livest. Prod. Sci. 2000, 64, 239–251. [Google Scholar] [CrossRef]
- Dersjant-Li, Y.; Peisker, M. A review on recent findings on amino acids requirements in poultry studies. Iran. J. Appl. Anim. Sci. 2011, 52, 73–79. [Google Scholar]
- Novak, C.; Yakout, H.M.; Scheideler, S.E. The effect of dietary protein level and total sulfur amino acid: Lysine ratio on egg production parameters and egg yield in Hy-Line W-98 hens. Poult. Sci. 2006, 85, 2195–2206. [Google Scholar] [CrossRef]
- Wu, G. Amino acids: Metabolism, functions and nutrition. Amino Acids 2009, 37, 1–17. [Google Scholar] [CrossRef]
- Berri, C.; Besnard, J.; Relandeau, C. Increasing dietary lysine increases final pH and decreases drip loss of broiler breast meat. Poult. Sci. 2008, 87, 480–484. [Google Scholar] [CrossRef] [PubMed]
- Emmert, J.; Baker, D. Use of the ideal protein concept for precision formulation of amino acid levels in broiler diets. J. Appl. Poult. Res. 1997, 6, 462–470. [Google Scholar] [CrossRef]
- Hermann, T. Industrial production of amino acid by coryneform bacteria. J. Biotechnol. 2003, 104, 155–172. [Google Scholar] [CrossRef]
- Leuchtenberger, W.; Huthmacher, K.; Drauz, K. Biotechnological production of amino acids and derivatives: Current status and prospects. Appl. Microbiol. Biotechnol. 2005, 1, 1–8. [Google Scholar] [CrossRef]
- Ikeda, M. Amino acid production processes. In Advances in Biochemical Engineering/Biotechnology; Scheper, T., Faurie, R., Thommel, J., Eds.; Springer: New York, NY, USA, 2003; Volume 79, pp. 1–35. ISBN 978-3-540-45989-7. [Google Scholar]
- Kinoshita, S.; Ukada, S.; Shimono, M. Studies on the amino acid fermentation. J. Gen. Appl. Microbiol. 1957, 3, 193–205. [Google Scholar] [CrossRef]
- Becker, J.; Zelder, O.; Hafner, S.; Schroder, H.; Wittmann, C. From zero to hero-design based systems metabolic engineering of Corynebacterium glutamicum for L-lysine production. Metab. Eng. 2011, 13, 159–168. [Google Scholar] [CrossRef] [PubMed]
- El-Nemr, M.; Mostafa, H.E. Screening of potential infants lactobacilli isolates for amino acids production. Afr. J. Microbiol. Res. 2010, 4, 226–232. [Google Scholar] [CrossRef]
- Simova, E.; Simov, Z.; Beshkova, D.; Frengova, G.; Dimitrov, Z.; Spasov, Z. Amino acid profiles of lactic acid bacteria, isolated from kefir grains and kefir starter made from them. Int. J. Food Microbiol. 2006, 107, 112–123. [Google Scholar] [CrossRef]
- Axelsson, L. Lactic acid bacteria: Classification and physiology. In Lactic Acid Bacteria: Microbiological and Functional Aspects, 3rd ed.; Salminen, S., Wright, A.V., Ouwehand, A., Eds.; Marcel Dekker, Inc.: New York, NY, USA, 1998; pp. 1–72. ISBN 9780824701338. [Google Scholar]
- Leroy, F.; Vuyst, L. Lactic acid bacteria as functional starter cultures for the food fermentation industry. Trends Food Sci. Technol. 2004, 15, 67–78. [Google Scholar] [CrossRef]
- Ong, L.; Henriksson, A.; Shah, N.P. Proteolytic pattern and organic acid profiles of probiotic ceddar cheese as influenced by probiotics strains of Lactobacillus acidophillus, Lb. paracasei, Lb. casei or Bifidobacterium sp. Int. Dairy J. 2007, 17, 67–78. [Google Scholar] [CrossRef]
- Niven, G.W.; Knight, D.J.; Mulholland, F. Changes in the concentrations of free amino acid in milk during growth of Lactococcus lactis indicate biphasic nitrogen metabolism. J. Dairy Res. 1998, 65, 101–107. [Google Scholar] [CrossRef]
- Mills, O.E.; Thomas, T.D. Nitrogen sources for growth of lactic streptococci in milk. N. Z. J. Dairy Sci. 1981, 16, 43–55. [Google Scholar]
- Law, B.; Kolstad, J. Proteolytic system in lactic acid bacteria. Antonie van Leeuwenhoek 1983, 49, 225–245. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Bayjanov, J.; Renckens, B.; Nauta, A.; Roland, J. The proteolytic system of lactic acid bacteria revisited: A genomic comparison. BMC Genom. 2010, 11, 36. [Google Scholar] [CrossRef] [PubMed]
- Kok, J. Genetics of the proteolytic system of lactic acid bacteria. FEMS Microbiol. Rev. 1990, 87, 15–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mierau, I.; Kunji, E.; Venema, G.; Kok, J. Casein and peptide degradation in lactic acid bacteria. Biotechnol. Genet. Eng. Rev. 1997, 14, 279–301. [Google Scholar] [CrossRef]
- Savijoki, K.; Ingmer, H.; Varmanen, P. Proteolytic system of lactic acid bacteria. Appl. Microbiol. Biotechnol. 2006, 71, 394–406. [Google Scholar] [CrossRef]
- Gobbetti, M.; Lanciotti, R.; De Angelis, M.; Corbo, M.R.; Massini, R.; Fox, P.F. Study of the effects of temperature, pH and NaCl on the peptidase activities of non-starter lactic acid bacteria (NSLAB) by quadratic response surface methodology. Int. Dairy J. 1999, 9, 865–875. [Google Scholar] [CrossRef]
- Kunji, E.; Mierau, I.; Hagfing, A.; Poolman, B.; Konings, W. The proteolytic system of lactic acid bacteria. Antonie van Leeuwenhoek 1996, 70, 187–221. [Google Scholar] [CrossRef]
- Juillard, V.; Le Bars, D.; Kunji, E.; Konings, W.; Gripon, J.; Richard, J. Oligopeptides are the main source of nitrogen for Lactococcus lactis during growth in milk. J. Appl. Environ. Microbiol. 1995, 61, 3024–3030. [Google Scholar] [CrossRef]
- Dalmis, U.; Soyer, A. Effect of processing methods and starter culture (Staphylococcus xylosus and Pediococcus pentosaceus) on proteolytic changes in Turkish sausages (sucuk) during ripening and storage. Meat Sci. 2008, 80, 345–354. [Google Scholar] [CrossRef]
- Essid, I.; Medini, M.; Hassouna, M. Technological and safety properties of Lactobacillus plantarum strains isolated from a Tunisian traditional salted meat. Meat Sci. 2009, 81, 203–208. [Google Scholar] [CrossRef]
- Simitsopoulou, M.; Choli-papadopoulou, T.; Alichanidis, E. Purification and partial characterization of a tripeptidase from Pediococcus pentosaceus K9.2. J. Appl. Environ. Microbiol. 1997, 63, 4872–4876. [Google Scholar]
- Fadda, S.; Sanz, Y.; Vignolo, G.; Aristoy, M.; Oliver, G.; Toldra, F. Characterization of muscle sarcoplasmic and myofibrillar protein hydrolysis caused by Lactobacillus plantarum. Appl. Environ. Microbiol. 1999, 65, 3540–3546. [Google Scholar]
- Fadda, S.; Sanz, Y.; Vignolo, G.; Aristoy, M.; Oliver, G.; Toldra, F. Hydrolysis of pork muscle sarcoplasmic proteins by Lactobacillus curvatus and Lactobacillus sakei. Appl. Environ. Microbiol. 1999, 65, 578–890. [Google Scholar]
- Vafopoulou-Mastrojiannaki, A.; Litopoulou-Tzanetaki, E.; Tzanetakis, N. Proteinase, peptidase and esterase activity of crude cell-free extracts of Pediococcus pentosaceus isolated from cheese. Food Sci. Technol. 1994, 27, 342–346. [Google Scholar] [CrossRef]
- Becker, J.; Christoph, W. Bio-based production of chemicals, materials and fuels- Corynebacterium glutamicum as versatile cell factory. Curr. Opin. Biotechnol. 2012, 23, 631–640. [Google Scholar] [CrossRef]
- Moosavi-Nasab, M.; Ansari, S.; Montazer, Z. Fermentative production of lysine by Corynebacterium glutamicum from different carbon sources. Iran Agric. Res. 2007, 25, 99–106. [Google Scholar] [CrossRef]
- Wendisch, V.F.; Jorge, J.M.; Perez-García, F.; Sgobba, E. Updates on industrial production of amino acids using Corynebacterium glutamicum. World J. Microbiol. Biotechnol. 2016, 32, 105. [Google Scholar] [CrossRef] [PubMed]
- Chopin, A. Organization and regulation of genes for amino acid biosynthesis in lactic acid bacteria. FEMS Microbiol. Rev. 1993, 12, 21–37. [Google Scholar] [CrossRef]
- Sriphochanart, W.; Skolpap, W.; Scharer, J.M.; Moo-Young, M.; Douglas, P.L. Effect of amino acid requirements on the growth and lactic acid production of Pediococcus acidilactici culture. Afr. J. Microbiol. Res. 2011, 5, 3815–3822. [Google Scholar] [CrossRef]
- Yin, Y.; Yao, K.; Liu, Z.; Gong, M.; Ruan, Z.; Deng, D.; Tan, B.; Liu, Z.; Wu, G. Supplementing L-leucine to a low-protein diet increases tissue protein synthesis in weanling pigs. Amino Acids 2010, 39, 1477–1486. [Google Scholar] [CrossRef]
- Kim, J.Y.; Burghardt, R.C.; Wu, G.; Johnson, G.A.; Spencer, T.E.; Bazer, F.W. Select nutrients in the ovine uterine lumen. VII. Effects of arginine, leucine, glutamine, and glucose on trophectoderm cell signalling, proliferation, and migration. Biol. Reprod. 2011, 84, 62–69. [Google Scholar] [CrossRef]
- Cunin, R.; Nicolas, G.; Andre, P.; Victor, S. Biosynthesis and metabolism of arginine in bacteria. Microbiol. Rev. 1986, 50, 314–352. [Google Scholar] [PubMed]
- Morishita, T.; Deguchi, Y.; Yajima, M.; Sakurai, T.; Yura, T. Multiple nutritional requirements of Lactobacilli: Genetic lesions affecting amino acid biosynthetic pathways. J. Bacteriol. 1981, 148, 64–71. [Google Scholar]
- Yvon, M.; Berthelot, S.; Gripon, J. Adding α-ketoglutarate to semi-hard cheese curd highly enhances the conversion of amino acids to aroma compounds. Int. Dairy J. 1998, 8, 889–898. [Google Scholar] [CrossRef]
- Ganzle, M.; Vermeulen, N.; Vogel, R. Carbohydrate, peptide and lipid metabolism of lactic acid bacteria in sourdough. Food Microbiol. 2007, 24, 128–138. [Google Scholar] [CrossRef]
- Pritchard, G.G.; Coolbear, T. The physiology and biochemistry of the proteolytic system in lactic acid bacteria. FEMS Microbiol. Rev. 1993, 12, 179–206. [Google Scholar] [CrossRef]
- Thu, T.V.; Foo, H.L.; Loh, T.C.; Bejo, M.H. Inhibitory activity and organic acid concentrations of metabolite combinations produced by various strains of Lactobacillus plantarum. Afr. J. Biotechnol. 2011, 10, 1359–1363. [Google Scholar] [CrossRef]
- Thung, T.Y. Isolation and Purification of Proteolytic Enzyme Produced by Lactic Acid Bacteria from Budu and Bambangan. Master’s Thesis, Universiti Putra Malaysia, Serdang, Malaysia, 2012. [Google Scholar]
- Wang, S.; Wang, C.; Huang, T. Microbial reclamation of squid pen for the production of a novel extracellular serine protease by Lactobacillus paracasei subsp. paracasei TKU012. Bioresour. Technol. 2008, 41, 1545–1552. [Google Scholar] [CrossRef]
- Klaenhammer, T.R.; Barrangou, R.; Buck, B.L.; Azcarate-Peril, M.A.; Altermann, E. Genomic features of lactic acid bacteria effecting bioprocessing and health. FEMS Microbiol. Rev. 2005, 29, 393–409. [Google Scholar] [CrossRef] [Green Version]
- Nissen-Meyer, J.; Sletten, K. Purification and characterization of the free form of the lactococcal extracellular proteinase and its autoproteolytic cleavage products. Microbiology. 1991, 137, 1611–1618. [Google Scholar] [CrossRef] [Green Version]
- Flambard, B.; Helinck, S.; Richard, J.; Juillard, V. The contribution of caseins to the amino acid supply for Lactococcus lactis depends on the type of cell envelope proteinase. Appl. Environ. Microbiol. 1998, 64, 1991–1996. [Google Scholar]
- Lim, Y.S. Isolation of bacteriocinogenic lactic acid bacteria and purification of selected bacteriocins from traditional fermented foods. Master’s Thesis, Universiti Putra Malaysia, Serdang, Malaysia, 2003. [Google Scholar]
- Kareem, K.Y.; Foo, H.L.; Loh, T.C.; Ooi, M.F.; Samsudin, A. Inhibitory Activity of Postbiotic produced by strains of Lactobacillus plantarum using reconstituted media supplemented with inulin. Gut Pathog. 2014, 6, 23. [Google Scholar] [CrossRef]
- Bradford, M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Miller, G.L. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| AA (mg/L) | Incubation Time (h) | Maximum Increment | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 | 2 | 4 | 6 | 8 | 10 | 12 | 14 | 16 | 18 | 20 | 22 | 24 | Amount (mg/L) | Time (h) | |
| Asp | 44.49 ± 1.23 b | 44.59 ± 2.82 b | 48.39 ± 3.13 ab | 48.33 ± 1.64 ab | 52.18 ± 3.97 ab | 55.59 ± 1.66 a | 50.25 ± 2.55 ab | 50.06 ± 2.28 ab | 46.85 ± 1.16 b | 48.5 ± 0.71 ab | 46.1 ± 1.00 b | 49.3 ± 1.78 ab | 46.3 ± 3.10 b | 11.10 | 10 |
| Asn | 53.05 ± 2.10 g | 55.74 ± 1.61 efg | 54.57 ± 1.04 fg | 53.71 ± 0.67 g | 58.23 ± 1.50 defg | 65.98 ± 2.75 bcde | 60.54 ± 5.57 cdefg | 65.24 ± 2.14 bcdef | 74.83 ± 3.45 ab | 83.40 ± 6.42 a | 70.47 ± 3.56 bc | 67.03 ± 3.39 bcd | 62.52 ± 2.76 cdefg | 30.35 | 18 |
| Lys | 340.66 ± 9.12 a | 342.35 ± 5.89 a | 345.42 ± 7.92 a | 350.57 ± 8.65 a | 356.39 ± 9.00 a | 348.73 ± 9.91 a | 350.65 ± 3.68 a | 354.88 ± 9.22 a | 347.39 ± 5.85 a | 348.68 ± 3.76 a | 340.83 ± 8.81 a | 342.10 ± 1.96 a | 338.27 ± 4.32 a | 15.73 | 8 |
| Met | 93.67 ± 0.31 c | 99.36 ± 1.67 bc | 108.92 ± 6.91 abc | 110.77 ± 1.92 abc | 122.54 ± 8.02 a | 120.21 ± 6.95 ab | 118.05 ± 7.17 ab | 117.57 ± 7.45 ab | 114.51 ± 5.02 ab | 118.08 ± 6.34 ab | 115.52 ± 7.15 ab | 112.99 ± 8.80 abc | 113.68 ± 6.77 abc | 28.87 | 8 |
| Thr | 62.42 ± 4.41 cd | 65.36 ± 5.01 cd | 63.94 ± 4.67 cd | 57.79 ± 6.13 d | 84.38 ± 4.16 ab | 87.57 ± 3.53 a | 75.76 ± 8.23 abc | 74.50 ± 1.37 abc | 70.54 ± 1.29 bcd | 63.30 ± 6.96 cd | 64.74 ± 1.83 cd | 66.38 ± 3.05 cd | 72.30 ± 3.02 bcd | 25.15 | 10 |
| Ile | 83.71 ± 5.85 d | 102.14 ± 3.36 c | 103.96 ± 1.47 c | 114.18 ± 10.20 c | 130.18 ± 6.49 ab | 135.49 ± 5.19 a | 128.00 ± 4.26 b | 131.72 ± 4.71 ab | 129.88 ± 2.63 ab | 129.10 ± 4.07 ab | 124.81 ± 7.77 ab | 127.50 ± 2.97 b | 129.76 ± 4.25 ab | 51.78 | 10 |
| Glu | 222.01 ± 1.60 e | 220.81 ± 4.06 e | 249.61 ± 4.81 cd | 244.21 ± 5.22 d | 284.27 ± 8.25 ab | 290.55 ± 6.62 a | 273.72 ± 6.06 ab | 275.08 ± 7.96 ab | 267.30 ± 6.50 c | 280.88 ± 6.92 ab | 272.17 ± 6.08 ab | 281.05 ± 5.89 ab | 274.19 ± 8.15 ab | 68.54 | 10 |
| Pro | 40.50 ± 2.50 c | 45.84 ± 2.83 c | 47.58 ± 5.46 c | 45.77 ± 0.92 c | 54.75 ± 1.40 b | 58.50 ± 1.12 b | 55.59 ± 1.60 b | 61.03 ± 2.53 ab | 58.19 ± 1.75 ab | 62.30 ± 0.50 ab | 64.70 ± 1.06 a | 63.76 ± 2.54 a | 61.36 ± 3.12 ab | 24.20 | 20 |
| Arg | 238.25 ± 0.78 a | 223.86 ± 8.48 b | 196.86 ± 7.88 c | 174.03 ± 5.87 d | 170.99 ± 4.54 d | 151.10 ± 5.13 e | 84.79 ± 1.61 f | 57.02 ± 1.53 g | 0.00 ± 0.00 h | 0.00 ± 0.00 h | 0.00 ± 0.00 h | 0.00 ± 0.00 h | 0.00 ± 0.00 h | 0.00 | 0 |
| Ala | 180.54 ± 3.95 a | 181.39 ± 5.21 a | 184.62 ± 2.24 a | 183.95 ± 1.67 a | 194.11 ± 5.41 a | 193.95 ± 2.16 a | 192.19 ± 4.17 a | 194.12 ± 5.57 a | 189.12 ± 4.17 a | 186.65 ± 6.81 a | 186.87 ± 1.69 a | 185.51 ± 7.51 a | 184.51 ± 2.86 a | 13.58 | 14 |
| Val | 109.06 ± 8.31 a | 114.43 ± 9.13 a | 111.85 ± 3.76 a | 118.48 ± 6.94 a | 126.91 ± 8.90 a | 130.32 ± 5.04 a | 126.23 ± 1.47 a | 127.77 ± 7.72 a | 123.26 ± 6.65 a | 121.45 ± 7.06 a | 122.75 ± 5.89 a | 121.93 ± 2.28 a | 118.53 ± 7.02 a | 21.26 | 10 |
| Leu | 356.91 ± 9.43 d | 365.94 ± 4.96 cd | 374.44 ± 4.87 bcd | 379.66 ± 22.01 abcd | 381.71 ± 6.35 abcd | 393.85 ± 8.08 abc | 385.36 ± 4.21 abcd | 405.80 ± 9.19 a | 402.49 ± 1.61 ab | 407.03 ± 8.22 a | 390.77 ± 4.27 abc | 399.67 ± 6.93 ab | 384.66 ± 7.50 abcd | 50.12 | 18 |
| Phe | 152.11 ± 3.68 ab | 153.85 ± 3.11 a | 149.42 ± 2.95 ab | 135.77 ± 4.37 bc | 141.17 ± 8.82 abc | 132.20 ± 4.30 c | 132.06 ± 4.63 c | 127.67 ± 7.07 cd | 114.61 ± 5.77 de | 128.20 ± 6.25 cd | 124.42 ± 3.51 cd | 106.77 ± 5.66 ef | 93.86 ± 5.22 f | 1.74 | 2 |
| Trp | 88.33 ± 1.24 bc | 86.62 ± 3.03 bc | 84.67 ± 0.96 bc | 83.01 ± 7.91 c | 66.46 ± 0.61 a | 60.93 ± 1.94 b | 0.00 ± 0.00 d | 0.00 ± 0.00 d | 0.00 ± 0.00 d | 0.00 ± 0.00 d | 0.00 ± 0.00 d | 0.00 ± 0.00 d | 0.00 ± 0.00 d | 0.00 | 0 |
| Tyr | 80.43 ± 3.82 a | 78.98 ± 2.91 ab | 71.58 ± 1.38 abc | 69.85 ± 4.05 bc | 68.01 ± 5.02 c | 71.38 ± 3.41 abc | 66.81 ± 5.90 c | 41.07 ± 20.55 c | 22.5 ± 22.5 c | 0.00 ± 0.00 d | 0.00 ± 0.00 d | 0.00 ± 0.00 d | 0.00 ± 0.00 d | 0.00 | 0 |
| Ser | 116.21 ± 2.75 c | 119.57 ± 3.31 bc | 132.30 ± 2.23 ab | 131.71 ± 4.58 ab | 138.06 ± 5.20 a | 143.80 ± 3.87 a | 121.63 ± 2.03 bc | 108.90 ± 3.09 c | 84.25 ± 6.74 d | 61.96 ± 11.84 e | 0.00 ± 0.00 f | 0.00 ± 0.00 f | 0.00 ± 0.00 f | 27.59 | 10 |
| Gly | 75.04 ± 0.68 c | 73.66 ± 0.30 c | 74.75 ± 1.51 c | 72.24 ± 0.97 c | 97.02 ± 2.50 b | 112.44 ± 6.51 a | 104.06 ±5.14 ab | 101.75 ±2.85 ab | 102.01 ±7.35 ab | 106.13 ±2.65 ab | 101.51 ±0.80 ab | 107.85 ±5.75 ab | 105.31 ±4.56 ab | 37.39 | 10 |
| Cy2 | 122.61 ± 6.07 d | 138.17 ± 3.76 cd | 149.74 ± 1.65 bc | 145.44 ± 5.20 bc | 151.19 ± 2.31 bc | 159.70 ± 0.97 ab | 161.92 ± 9.47 ab | 158.38 ± 2.23 ab | 171.85 ± 7.38 a | 160.82 ± 4.80 ab | 159.92 ± 2.47 ab | 152.45 ± 7.89 bc | 148.60 ± 6.72 bc | 49.24 | 16 |
| AA (mg/L) | Incubation Time (h) | Maximum Increment | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 | 2 | 4 | 6 | 8 | 10 | 12 | 14 | 16 | 18 | 20 | 22 | 24 | Amount (mg/L) | Time (h) | |
| Asp | 42.00 ± 1.23 g | 42.85 ± 2.12 g | 46.42 ± 0.88 g | 52.34 ± 1.09 ef | 55.74 ± 0.97 cdef | 57.13 ± 1.34 cde | 51.49 ± 1.08 f | 60.53 ± 4.13 bc | 64.76 ± 0.40 ab | 65.83 ± 1.34 a | 55.92 ± 0.51 cdef | 55.28 ± 0.15 def | 57.77 ± 1.37 cd | 23.83 | 18 |
| Asn | 52.15 ± 0.85 a | 50.59 ± 2.88 a | 49.78 ± 1.51 ab | 48.52 ± 1.05 ab | 49.86 ± 2.50 ab | 48.21 ± 0.33 ab | 44.57 ± 0.35 b | 47.82 ± 4.24 b | 0.00 ± 0.00 c | 0.00 ± 0.00 c | 0.00 ± 0.00 c | 0.00 ± 0.00 c | 0.00 ± 0.00 c | 0 | 0 |
| Lys | 332.39± 19.03 bcd | 311.34 ± 4.88 d | 334.14 ± 25.56 bcd | 324.75 ± 7.38 bcd | 320.14 ± 0.36 cd | 306.72 ± 10.51 d | 313.18 ± 5.99 d | 335.32 ± 29.39 abcd | 364.37 ± 7.32 bc | 370.84 ± 13.51 ab | 372.07 ± 0.26 ab | 362.82 ± 21.34 abc | 383.18 ± 0.81 a | 50.79 | 24 |
| Met | 104.20 ± 5.00 f | 105.57 ± 1.61 f | 119.00 ± 4.79 e | 129.71 ± 0.33 d | 132.44 ± 2.04 d | 133.46 ± 3.76 d | 137.64 ± 1.78 cd | 139.55 ± 6.60 cd | 156.69 ± 1.29 a | 155.67 ± 1.87 a | 145.71 ± 0.87 bc | 138.99 ± 2.89 cd | 151.14 ± 0.92 a | 52.49 | 16 |
| Thr | 71.38 ± 2.08 h | 71.77 ± 2.23 h | 81.26 ± 4.11 gh | 87.87 ± 1.67 fg | 100.58 ± 0.21 de | 105.68 ± 0.30 cd | 94.94 ± 6.87 ef | 119.81 ± 8.65 ab | 127.49 ± 0.70 a | 130.16 ± 1.52 a | 120.28 ± 1.89 ab | 113.51 ± 0.44 bc | 124.87 ± 0.71 a | 58.78 | 18 |
| Ile | 100.20 ± 6.31 d | 104.15 ± 2.72 d | 113.03 ± 7.91 c | 139.29 ± 4.65 b | 162.10 ± 0.35 a | 139.19 ± 3.24 b | 130.53 ± 1.65 b | 133.08 ± 8.43 b | 135.59 ± 0.24 b | 136.64 ± 2.02 b | 132.20 ± 2.04 b | 126.35 ± 0.99 b | 134.85 ± 0.58 b | 61.9 | 8 |
| Glu | 233.74 ± 3.42 f | 233.84 ± 7.56 f | 252.29 ± 13.01 ef | 262.24 ± 5.60 de | 284.71 ± 1.84 cd | 285.51 ± 3.11 cd | 272.98 ± 3.70 de | 307.21 ± 21.76 abc | 322.33 ± 0.47 ab | 331.74 ± 2.57 a | 304.21 ± 2.44 bc | 305.37 ± 0.69 bc | 320.41 ± 4.83 ab | 98 | 18 |
| Pro | 45.72 ± 2.29 d | 45.85 ± 3.00 d | 46.07 ± 2.77 d | 55.35 ± 0.88 c | 61.99 ± 0.54 bc | 66.90 ± 2.33 ab | 68.00 ± 0.88 ab | 65.40 ± 1.13 ab | 72.89 ± 1.47 a | 73.46 ± 2.49 a | 73.57 ± 5.10 a | 73.69 ± 4.51 a | 70.74 ± 0.98 a | 27.97 | 22 |
| Arg | 184.04 ± 4.80 ab | 186.04 ± 13.80 ab | 195.74 ± 0.53 a | 193.36 ± 2.92 a | 193.93 ± 0.96 a | 171.91 ± 1.64 b | 135.41 ± 4.12 c | 124.19 ± 6.78 c | 0.00 ± 0.00 d | 0.00 ± 0.00 d | 0.00 ± 0.00 d | 0.00 ± 0.00 d | 0.00 ± 0.00 d | 11.7 | 4 |
| Ala | 163.10 ± 6.42 h | 165.94 ± 10.87 gh | 178.11 ± 1.72 fgh | 183.03 ± 3.23 efg | 192.69 ± 0.44 cdef | 189.74 ± 0.19 cdef | 185.52 ± 2.86 def | 206.14 ± 16.66 bc | 216.50 ± 0.65 ab | 226.89 ± 1.54 a | 219.55 ± 1.97 ab | 205.44 ± 5.74 bcd | 199.98 ± 1.80 bcde | 63.79 | 18 |
| Val | 87.28 ± 2.24 f | 83.40 ± 1.82 f | 82.60 ± 5.53 f | 82.92 ± 0.40 f | 86.33 ± 1.47 f | 89.36 ± 2.54 ef | 89.25 ± 1.14 ef | 95.94 ± 4.00 de | 103.79 ± 0.35 bc | 108.39 ± 0.82 ab | 104.36 ± 1.77 bc | 99.77 ± 1.24 cd | 112.89 ± 1.57 a | 25.61 | 24 |
| Leu | 357.86 ± 9.06 c | 359.20 ± 7.46 c | 382.01 ± 21.38 bc | 363.62 ± 11.63 c | 387.07 ± 2.32 bc | 377.13 ± 2.02 bc | 379.77 ± 13.43 bc | 411.17 ± 32.76 ab | 437.75 ± 0.45 a | 453.28 ± 4.61 a | 434.79 ± 6.94 a | 428.33 ± 10.83 a | 452.54 ± 2.82 a | 95.42 | 18 |
| Phe | 141.52 ± 3.06 a | 133.00 ± 4.80 ab | 128.18 ± 7.82 b | 113.66 ± 0.34 c | 112.54 ± 0.24 c | 109.08 ± 2.00 c | 107.55 ± 1.53 c | 110.14 ± 7.10 c | 109.32 ± 0.25 c | 110.67 ± 1.43 c | 110.56 ± 3.09 c | 103.07 ± 3.72 c | 114.04 ± 2.69 c | 0 | 0 |
| Trp | 73.91 ± 2.13 b | 81.64 ± 4.71 a | 82.25 ± 3.84 a | 70.46 ± 0.59 b | 0.00 ± 0.00 c | 0.00 ± 0.00 c | 0.00 ± 0.00 c | 0.00 ± 0.00 c | 0.00 ± 0.00 c | 0.00 ± 0.00 c | 0.00 ± 0.00 c | 0.00 ± 0.00 c | 0.00 ± 0.00 c | 8.34 | 4 |
| Tyr | 66.93 ± 2.60 b | 74.87 ± 6.90 a | 75.52 ± 5.58 a | 60.35 ± 0.92 b | 0.00 ± 0.00 c | 0.00 ± 0.00 c | 0.00 ± 0.00 c | 0.00 ± 0.00 c | 0.00 ± 0.00 c | 0.00 ± 0.00 c | 0.00 ± 0.00 c | 0.00 ± 0.00 c | 0.00 ± 0.00 c | 8.59 | 4 |
| Ser | 116.85 ± 4.63 d | 118.2 ± 2.5 d | 130.6 ± 8.3 cd | 129.6 ± 2.6 cd | 138.5 ± 0.4 c | 137.17 ± 0.30 c | 137.91 ± 2.02 c | 153.93 ± 12.63 b | 162.60 ± 0.97 ab | 167.43 ± 1.71 ab | 160.08 ± 1.32 ab | 161.62 ± 3.78 ab | 171.12 ± 0.91 a | 54.27 | 24 |
| Gly | 61.15 ± 3.33 g | 63.02± 4.09 fg | 71.12± 1.71 def | 70.44± 1.10 efg | 81.02± 0.65 bc | 74.41± 1.10 cde | 69.79± 1.59 efg | 80.10± 7.97 bcd | 85.74 ±1.17 ab | 91.87 ± 0.61 a | 86.16 ±1.93 ab | 88.71 ±2.57 ab | 94.90 ± 1.81 a | 33.75 | 24 |
| Cy2 | 91.72 ± 13.57 f | 107.43 ± 10.22 def | 134.01 ± 1.88 bcd | 119.77 ± 4.13 cdef | 146.32 ± 6.46 abc | 125.54 ± 10.11 bcde | 95.90 ± 4.17 ef | 154.36 ± 20.80 ab | 167.60 ± 10.06 a | 173.72 ± 8.97 a | 113.75 ± 7.96 def | 116.63 ± 0.48 cdef | 126.70 ± 4.76 bcde | 82 | 18 |
| AA (mg/L) | Incubation Time (h) | Maximum Increment | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 | 2 | 4 | 6 | 8 | 10 | 12 | 14 | 16 | 18 | 20 | 22 | 24 | Amount (mg/L) | Time (h) | |
| Asp | 36.47 ± 4.50 a | 33.89 ± 0.67 abc | 29.64 ± 1.55 bcd | 36.43 ± 2.22 a | 35.22 ± 0.80 ab | 36.82 ± 1.20 a | 31.55 ± 1.71 abcd | 27.34 ± 0.47 d | 31.89 ± 0.26 abcd | 31.66± 1.03 abcd | 30.39 ± 1.97 bcd | 28.77 ± 0.14 cd | 29.18 ± 0.93 cd | 0.36 | 10 |
| Asn | 48.56 ± 2.41 a | 42.92 ± 1.69 b | 38.66 ± 3.77 bc | 37.40 ± 1.89 bc | 33.75 ± 1.83 c | 34.49 ± 2.05 c | 36.54 ± 1.76 c | 34.26 ± 0.09 c | 38.14 ± 0.43 bc | 34.79 ± 0.62 cd | 36.50 ± 0.27 cd | 34.52 ± 1.65 cd | 33.48 ± 2.04 cd | 0 | 0 |
| Lys | 280.87 ± 2.82 bcd | 257.85 ± 9.62 ef | 253.09 ± 1.11 f | 265.62 ± 4.45 cdef | 277.30 ± 3.34 bcde | 261.80 ± 13.39 def | 283.67 ± 0.28 bc | 271.29 ± 6.86 bcdef | 288.82 ± 2.28 b | 316.65 ± 5.85 a | 281.06 ± 4.69 bcd | 264.17 ± 5.72 cdef | 270.29 ± 5.03 bcdef | 35.78 | 18 |
| Met | 49.25 ± 0.06 d | 50.21 ± 0.10 cd | 54.21 ± 0.33 abc | 50.65 ± 3.23 cd | 53.08 ± 1.09 abcd | 52.04 ± 1.54 bcd | 54.29 ± 0.96 abc | 49.20 ± 0.99 d | 55.53 ± 0.47 ab | 56.42 ± 1.23 a | 53.07 ± 1.03 abcd | 53.24 ± 0.41 abcd | 52.21 ± 1.69 abcd | 7.18 | 18 |
| Thr | 56.83 ± 1.31 e | 55.64 ± 0.72 e | 59.77 ± 2.24 de | 69.04 ± 3.76 bc | 73.11 ± 5.27 abc | 66.38 ± 2.04 cd | 77.32 ± 2.52 a | 68.47 ± 0.65 c | 73.20 ± 0.19 abc | 76.71 ± 2.91 ab | 71.15 ± 1.64 abc | 77.16 ± 1.68 a | 76.63 ± 1.49 ab | 20.49 | 12 |
| Ile | 69.72 ± 3.00 c | 63.02 ± 1.76 d | 72.09 ± 1.53 bc | 77.74 ± 3.97 bc | 79.35 ± 2.63 a | 79.04 ± 0.79 a | 78.16 ± 0.79 a | 77.48 ± 0.29 bc | 83.60 ± 0.50 a | 81.57 ± 1.36 a | 81.51 ± 1.94 a | 78.36 ± 0.96 a | 77.93 ± 0.11 bc | 13.89 | 16 |
| Glu | 176.10 ± 11.28 cd | 167.47 ± 6.77 d | 181.38 ± 3.84 bcd | 206.26 ± 11.47 a | 194.23 ± 8.70 abc | 197.26 ± 4.79 ab | 197.14 ± 7.19 ab | 198.31 ± 0.68 ab | 202.49 ± 2.16 a | 195.86 ± 4.11 abc | 204.19 ± 2.73 a | 187.21 ± 2.70 abc | 192.91 ± 0.46 abc | 30.16 | 6 |
| Pro | 53.26 ± 0.50 e | 48.23 ± 0.09 f | 56.73 ± 3.86 de | 61.93 ± 1.27 abc | 65.21 ± 1.98 abc | 60.86 ± 1.43 bcd | 60.63 ± 1.14 cd | 61.49 ± 0.60 abcd | 63.90 ± 2.22 abc | 65.05 ± 0.77 abc | 62.04 ± 0.42 abc | 65.97 ± 1.10 ab | 66.34 ± 1.03 a | 13.08 | 24 |
| Arg | 151.92 ± 3.36 a | 136.53 ± 2.06 b | 138.67 ± 2.95 b | 127.31 ± 0.46 c | 81.59 ± 0.31 d | 36.66 ± 0.95 e | 3.42 ± 0.69 h | 4.22 ± 0.84 hg | 9.95 ± 3.34 f | 9.78 ± 1.35 fg | 3.73 ± 1.01 h | 4.49 ± 0.39 afgh | 4.73 ± 0.11 fgh | 0.00 | 0 |
| Ala | 119.73 ± 5.02 cd | 113.20 ± 2.87 d | 120.41 ± 2.86 cd | 120.92 ± 5.47 cd | 119.26 ± 7.08 cd | 120.77 ± 3.11 cd | 122.41 ± 2.85 cd | 127.30 ± 0.07 bc | 136.05 ± 1.93 ab | 142.69 ± 2.66 a | 138.88 ± 2.74 a | 137.85 ± 2.03 ab | 140.46 ± 0.27 a | 22.95 | 18 |
| Val | 77.42 ± 3.34 cd | 73.84 ± 1.44 d | 81.86 ± 2.36 bc | 90.22 ± 4.51 a | 91.56 ± 5.38 a | 90.92 ± 1.21 a | 92.20 ± 0.14 a | 89.65 ± 0.33 ab | 92.95 ± 0.41 a | 93.76 ± 1.56 a | 92.94 ± 2.83 a | 87.95 ± 0.29 ab | 89.18 ± 0.74 ab | 16.34 | 18 |
| Leu | 242.48 ± 5.95 ab | 217.21 ± 2.48 c | 237.33 ± 9.48 abc | 230.35 ± 2.65 bc | 232.80 ± 11.22 abc | 230.00 ± 10.01 bc | 235.80 ± 3.80 abc | 232.29 ± 2.76 bc | 254.11 ± 6.25 a | 240.59 ± 4.17 ab | 243.55 ± 6.33 ab | 240.59 ± 5.86 ab | 241.41 ± 3.22 ab | 11.63 | 16 |
| Phe | 122.16 ± 2.28 a | 113.90 ± 0.42 b | 108.57 ± 2.97 bc | 111.85 ± 5.93 b | 109.23 ± 2.00 bc | 108.38 ± 3.27 bc | 101.67 ± 1.51 cd | 97.11 ± 0.97 de | 99.12 ± 0.33 ed | 92.79 ± 2.81 e | 92.57 ± 2.79 e | 79.46 ± 0.25 f | 81.96 ± 0.48 f | 0.00 | 0 |
| Trp | 54.90 ± 0.26 bc | 57.81 ± 1.49 ab | 49.89 ± 1.58 d | 49.96 ± 3.10 d | 59.65 ± 0.96 ab | 60.90 ± 1.38 a | 54.96 ± 1.85 bc | 50.87 ± 0.19 cd | 58.15 ± 2.17 ab | 60.00 ± 0.93 a | 51.63 ± 0.06 cd | 49.80 ± 0.42 d | 50.35 ± 1.93 cd | 6.00 | 10 |
| Tyr | 28.18 ± 0.05 a | 26.70 ± 0.51 a | 23.67 ± 1.06 a | 26.70 ± 0.76 a | 26.73 ± 0.08 a | 26.88 ± 0.23 a | 25.01 ± 0.11 a | 23.48 ± 0.96 a | 26.26 ± 1.13 a | 28.24 ± 1.69 a | 22.77 ± 0.12 a | 24.06 ± 1.13 a | 22.57 ± 0.13 a | 0.06 | 0 |
| Ser | 79.77 ± 2.31 e | 80.54 ± 0.92 e | 87.92 ± 0.20 cde | 86.63 ± 3.27 de | 91.20 ± 5.84 bcd | 90.98 ± 6.72 bcd | 93.75 ± 1.01 abcd | 96.30 ± 2.39 abcd | 102.43 ± 1.10 a | 100.03 ± 1.56 ab | 100.57 ± 2.91 ab | 97.01 ± 1.62 abc | 97.41 ± 3.04 abc | 22.66 | 16 |
| Gly | 44.40 ± 0.68 d | 44.15 ± 0.08 d | 50.98 ± 1.52 c | 54.04 ± 1.87 bc | 54.70 ± 4.05 bc | 53.93 ± 4.08 bc | 58.88 ± 0.87 ab | 57.45 ± 1.98 abc | 63.79 ± 1.04 a | 61.67 ± 0.95 a | 62.23 ± 2.59 a | 61.85 ± 0.99 a | 62.94 ± 0.56 a | 19.39 | 16 |
| Cy2 | 106.99 ± 4.08 bcd | 95.74 ± 1.68 d | 99.30 ± 4.86 d | 98.22 ± 3.06 d | 98.06 ± 4.41 d | 100.73 ± 4.84 d | 103.80 ± 6.07 cd | 99.19 ± 7.21 d | 119.92 ±3.94 abc | 121.73 ± 1.49 ab | 120.03 ± 9.72 abc | 120.28 ± 1.94 abc | 133.13 ± 7.15 a | 26.14 | 24 |
| AA (mg/L) | Incubation Time (h) | Maximum Increment | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 | 2 | 4 | 6 | 8 | 10 | 12 | 14 | 16 | 18 | 20 | 22 | 24 | Amount (mg/L) | Time (h) | |
| Asp | 57.98 ± 1.18 a | 54.90 ± 2.67 ab | 56.22 ± 2.63 ab | 51.70 ± 2.47 b | 55.83 ± 3.37 ab | 57.29 ± 1.77 a | 56.46 ± 1.44 ab | 0.00 ± 0.00 c | 0.00 ± 0.00 c | 0.00 ± 0.00 c | 0.00 ± 0.00 c | 0.00 ± 0.00 c | 0.00 ± 0.00 c | 0.00 | 0 |
| Asn | 75.11 ± 0.81 a | 71.18 ± 4.77 ab | 72.34 ± 0.57 ab | 63.46 ± 3.29 c | 71.01 ± 4.55 ab | 71.64 ± 0.68 ab | 65.62 ± 1.06 bc | 46.97 ± 23.51 c | 0.00 ± 0.00 d | 0.00 ± 0.00 d | 0.00 ± 0.00 d | 0.00 ± 0.00 d | 0.00 ± 0.00 d | 0.00 | 0 |
| Lys | 440.20 ± 5.70 a | 451.33 ± 8.68 a | 436.07 ± 5.94 a | 401.44 ± 4.19 bc | 393.75 ± 6.83 c | 393.91 ± 1.37 c | 396.46 ± 3.22 c | 416.32 ± 5.07 b | 392.89 ± 6.21 c | 371.98 ± 5.54 d | 353.99 ± 7.18 d | 334.04 ± 8.66 e | 363.47 ± 8.77 d | 11.13 | 4 |
| Met | 100.81 ± 0.16 d | 100.91 ± 3.06 d | 113.87 ± 1.06 bcd | 106.57 ± 2.35 d | 109.80 ± 7.81 cd | 119.96 ± 4.28 abc | 119.0 ± 2.02 abc | 128.84 ± 0.07 a | 123.90 ± 1.32 ab | 125.77 ± 4.08 ab | 132.71 ± 7.93 a | 128.99 ± 5.37 a | 126.34 ± 4.55 ab | 31.90 | 20 |
| Thr | 92.89 ± 0.78 abc | 83.18 ± 5.30 c | 91.07 ± 6.15 abc | 88.30 ± 4.65 bc | 99.57 ± 6.62 abc | 107.96 ± 0.24 a | 100.49 ± 1.24 ab | 101.80 ± 2.22 ab | 100.00 ± 5.91 abc | 94.78 ± 8.43 abc | 68.25 ± 8.10 d | 54.26 ± 2.11 d | 55.36 ± 4.82 d | 15.06 | 10 |
| Ile | 123.70 ± 2.52 c | 146.71 ± 8.04 ab | 152.87 ± 3.66 a | 146.59 ± 7.59 ab | 152.09 ± 10.01 a | 155.07 ± 3.28 a | 148.25 ± 6.33 ab | 163.48 ± 4.09 a | 153.11 ± 1.19 a | 150.79 ± 6.04 ab | 127.52 ± 5.90 c | 122.54 ± 4.61 c | 133.48 ± 3.62 bc | 39.79 | 14 |
| Glu | 244.46 ± 5.57 e | 244.70 ± 6.49 e | 251.28 ± 8.53 de | 253.62 ± 4.49 de | 256.88 ± 3.49 cde | 280.46 ± 3.55 ab | 278.00 ± 2.47 abc | 290.27 ± 9.49 a | 271.35 ± 5.30 abcd | 263.94 ± 2.42 bcde | 264.36 ± 9.63 bcde | 262.52 ± 7.72 bcde | 262.39 ± 8.84 bcde | 45.81 | 14 |
| Pro | 37.24 ± 0.31 g | 37.90 ± 0.93 g | 46.25 ± 1.35 fg | 57.19 ± 1.83 ef | 65.22 ± 2.86 de | 71.56 ± 7.06 bcd | 80.46 ± 9.69 abc | 70.61 ± 4.57 cd | 75.65 ± 3.36 abcd | 79.55 ± 2.83 abc | 86.13 ± 2.49 a | 81.12 ± 5.04 ab | 85.06 ± 1.32 abc | 48.89 | 20 |
| Arg | 289.51 ± 2.16 a | 255.66 ± 9.01 b | 252.12 ± 6.78 b | 185.52 ± 8.83 c | 134.92 ± 1.87 d | 85.91 ± 4.61 e | 0.00 ± 0.00 f | 0.00 ± 0.00 f | 0.00 ± 0.00 f | 0.00 ± 0.00 f | 0.00 ± 0.00 f | 0.00 ± 0.00 f | 0.00 ± 0.00 f | 0.00 | 0 |
| Ala | 218.94 ± 3.05 a | 217.08 ± 8.85 a | 217.19 ± 3.09 a | 193.60 ± 7.47 b | 195.88 ± 5.18 b | 183.84 ± 6.23 bc | 171.54 ± 2.78 cd | 179.86 ± 0.73 bcd | 173.01 ± 2.24 cd | 171.58 ± 3.35 cd | 164.20 ± 7.56 d | 166.71 ± 4.38 cd | 174.27 ± 7.02 cd | 0.00 | 0 |
| Val | 99.46 ± 3.79 cd | 100.54 ± 7.85 cd | 107.27 ± 7.50 bc | 107.50 ± 6.48 bc | 116.49 ± 6.91 abc | 124.67 ± 1.88 ab | 121.6 3± 7.14 ab | 129.84 ± 6.27 a | 124.2 ± 2.79 abc | 118.7 ± 7.15 ab | 84.10 ± 5.29 de | 79.27 ± 1.34 e | 88.05 ± 3.17 de | 30.38 | 14 |
| Leu | 430.99 ± 1.80 cdef | 432.12 ± 2.26 cdef | 443.27 ± 4.87 bc | 434.76 ± 4.65 bcdef | 434.55 ± 8.63 bcdef | 441.52 ± 4.94 bcd | 449.68 ± 7.16 ab | 462.72 ± 3.23 a | 440.30 ± 7.47 bcd | 438.22 ± 2.34 cdef | 425.77 ± 0.90 def | 420.06 ± 3.30 f | 423.88 ± 4.78 ef | 31.73 | 14 |
| Phe | 145.38 ± 6.60 b | 128.71 ± 8.37 bcd | 137.25 ± 8.31 bc | 111.48 ± 8.21 d | 118.73 ± 7.45 cd | 11.8.84 ± 6.10 cd | 120.73 ± 5.88 cd | 122.46 ± 4.29 cd | 117.31 ± 3.36 cd | 115.50 ± 3.69 d | 128.33 ± 3.36 bcd | 167.96 ± 2.28 a | 183.25 ± 7.87 a | 37.87 | 24 |
| Trp | 124.92 ± 1.82 a | 115.80 ± 6.82 a | 120.21 ± 4.47 a | 109.97 ± 3.29 ab | 121.31 ± 10.91 a | 117.66 ± 5.71 a | 117.69 ± 4.14 a | 123.80 ± 2.09 a | 119.80 ± 2.11 a | 112.42 ± 4.65 ab | 100.15 ± 4.93 b | 0.00 ± 0.00 c | 0.00 ± 0.00 c | 0.00 | 0 |
| Tyr | 83.04 ± 1.57 a | 82.59 ± 4.87 a | 83.90 ± 3.31 a | 78.11 ± 3.61 a | 79.92 ± 5.90 a | 78.70 ± 3.61 a | 51.04 ± 25.53 a | 0.00 ± 0.00 b | 0.00 ± 0.00 b | 0.00 ± 0.00 b | 0.00 ± 0.00 b | 0.00 ± 0.00 b | 0.00 ± 0.00 b | 0.87 | 4 |
| Ser | 138.54 ± 2.09 ab | 134.52 ± 7.06 abc | 140.82 ± 1.51 a | 127.12 ± 8.08 ab | 120.16 ± 9.67 bc | 121.88 ± 0.83 bc | 115.87 ± 4.43 c | 119.26 ± 3.16 c | 117.68 ± 3.24 c | 119.35 ± 2.13 c | 127.32 ± 7.52 abc | 134.03 ± 5.55 abc | 132.45± 8.09 abc | 2.27 | 4 |
| Gly | 83.53 ± 1.85 d | 77.90 ± 6.94 d | 86.73 ± 3.26 cd | 80.31 ± 5.00 d | 100.67 ± 9.01 bc | 101.07 ± 3.93 bc | 100.36 ± 1.90 bc | 101.20 ± 3.00 bc | 97.73 ± 6.10 bc | 107.92 ± 3.04 b | 126.46 ± 0.82 a | 124.11 ± 2.94 a | 122.66 ± 4.18 a | 42.94 | 20 |
| Cy2 | 176.31 ± 6.49 a | 172.49 ± 7.9 a | 173.99 ± 6.76 a | 172.53 ± 7.20 a | 173.27 ± 9.76 a | 171.17 ± 4.10 a | 174.64 ± 6.07 a | 173.67 ± 7.17 a | 176.47 ± 7.48 a | 178.63 ± 8.11 a | 190.63 ± 8.67 a | 185.97 ± 3.73 a | 178.97 ± 9.35 a | 14.32 | 20 |
| AA (mg/L) | Incubation Time (h) | Maximum Increment | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 | 2 | 4 | 6 | 8 | 10 | 12 | 14 | 16 | 18 | 20 | 22 | 24 | Amount (mg/L) | Time (h) | |
| Asp | 43.30 ± 1.48 a | 44.28 ± 2.44 a | 43.04 ± 0.64 a | 40.77 ± 3.58 a | 40.60 ± 1.84 a | 39.40 ± 1.44 a | 40.49 ± 1.15 a | 28.81 ± 14.41 a | 0.00 ± 0.00 b | 0.00 ± 0.00 b | 0.00 ± 0.00 b | 0.00 ± 0.00 b | 0.00 ± 0.00 b | 0.98 | 2 |
| Asn | 53.15 ± 2.24 a | 51.34 ± 2.53 a | 43.29 ± 1.29 b | 40.84 ± 4.11 b | 43.70 ± 2.84 b | 27.72 ± 13.86 b | 0.00 ± 0.00 c | 0.00 ± 0.00 c | 0.00 ± 0.00 c | 0.00 ± 0.00 c | 0.00 ± 0.00 c | 0.00 ± 0.00 c | 0.00 ± 0.00 c | 0.00 | 0 |
| Lys | 208.29 ± 5.04 a | 214.11 ± 6.76 a | 211.15 ± 6.87 a | 196.66 ± 4.33 a | 203.43 ± 8.84 a | 193.46 ± 7.98 a | 208.04 ± 9.82 a | 202.63 ± 1.18 a | 208.495 ± 9.31 a | 162.31 ± 9.21 b | 149.98 ± 8.49 bc | 141.98 ± 3.14 bc | 136.19 ± 8.04 c | 5.83 | 2 |
| Met | 48.46 ± 2.31 ab | 54.49 ± 3.54 a | 49.73 ± 1.97 ab | 54.65 ± 4.96 a | 58.38 ± 2.46 a | 54.12 ± 3.74 a | 53.38 ± 1.81 ab | 57.36 ± 1.98 a | 55.17 ± 2.24 a | 52.67 ± 6.63 ab | 50.90 ± 2.81 ab | 43.45 ± 1.76 b | 43.17 ± 1.02 b | 9.92 | 8 |
| Thr | 35.36 ± 4.39 d | 37.70 ± 5.61 cd | 43.99 ± 1.25 abcd | 53.92 ± 5.43 a | 53.73 ± 1.86 a | 52.73 ± 4.06 a | 50.39 ± 1.81 ab | 47.55 ± 2.51 abc | 54.34 ± 0.62 a | 47.91 ± 4.59 abc | 44.35 ± 0.99 abcd | 39.37 ± 0.23 cd | 40.14 ± 1.35 bcd | 18.98 | 16 |
| Ile | 59.05 ± 2.40 c | 66.67 ± 4.36 abc | 75.21 ± 6.56 ab | 78.43 ± 6.11 a | 74.38 ± 3.84 ab | 75.65 ± 6.39 a | 77.86 ± 4.71 a | 77.92 ± 2.94 a | 78.61 ± 4.48 a | 68.17 ± 7.85 abc | 60.18 ± 3.44 bc | 54.69 ± 0.16 c | 54.19 ± 1.40 c | 19.56 | 16 |
| Glu | 109.60 ± 4.73 g | 133.19 ± 7.69 f | 143.16± 1.87 def | 16.24 ± 0.47 abc | 175.41 ± 4.96 ab | 176.24 ± 2.19 a | 161.60 ± 8.24 abc | 158.48 ± 7.69 bcd | 174.43 ± 3.61 ab | 150.76 ± 7.46 cde | 145.5 8± 0.89 cdef | 139.28 ± 6.16 ef | 133.42 ± 1.95 f | 66.64 | 10 |
| Pro | 73.95 ± 3.41 cd | 68.84 ± 2.81 d | 74.54 ± 2.33 cd | 79.52 ± 3.94 cd | 89.80 ± 5.34 bc | 101.18 ± 8.00 ab | 101.38 ± 7.05 ab | 98.32 ± 4.60 ab | 107.03 ± 9.79 ab | 110.28 ± 1.77 a | 108.34 ± 4.18 ab | 102.92 ± 9.83 ab | 107.88 ± 1.54 ab | 36.33 | 18 |
| Arg | 141.29 ± 3.16 b | 156.59 ± 6.03 a | 147.95 ± 3.15 ab | 150.39 ± 6.79 ab | 139.18 ± 6.34 b | 83.14 ± 7.84 c | 65.65 ± 1.60 d | 55.41 ± 1.87 d | 0.00 ± 0.00 e | 0.00 ± 0.00 e | 0.00 ± 0.00 e | 0.00 ± 0.00 e | 0.00 ± 0.00 e | 15.31 | 2 |
| Ala | 114.10 ± 1.08 a | 123.10 ± 6.82 a | 125.31 ± 3.44 a | 128.66 ± 7.56 a | 117.87 ± 5.62 a | 90.83 ± 6.05 a | 87.06 ± 6.78 b | 86.58 ± 5.53 b | 91.88 ± 5.98 b | 83.37 ± 7.70 b | 81.82 ± 0.21 b | 76.79 ± 2.54 b | 78.77 ± 2.13 b | 14.56 | 6 |
| Val | 69.79 ± 0.84 bc | 72.43 ± 4.78 bc | 68.45 ± 4.76 c | 84.83 ± 8.13 abc | 94.68 ± 6.71 a | 87.23 ± 9.50 abc | 92.78 ± 8.68 a | 80.43 ± 4.25 abc | 88.61 ± 4.64 ab | 83.23 ± 7.81 abc | 88.72 ± 2.56 ab | 82.88 ± 1.07 abc | 81.16 ± 1.15 abc | 24.92 | 8 |
| Leu | 223.27 ± 9.04 e | 274.81 ± 3.48 bc | 269.09 ± 8.90 bc | 262.78 ± 8.73 bc | 277.75 ± 5.61 bc | 285.27 ± 6.25 ab | 302.91 ± 7.02 a | 302.62 ± 9.85 a | 301.61 ± 8.67 a | 254.14 ± 9.35 cd | 234.91 ± 9.63 de | 219.65 ± 3.95 e | 225.93 ± 5.83 e | 79.64 | 12 |
| Phe | 97.42 ± 0.23 bcd | 108.24 ± 8.07 ab | 108.10 ± 7.32 ab | 108.48 ± 9.44 ab | 110.57 ± 9.43 ab | 117.38 ± 3.08 a | 115.54 ± 6.00 ab | 103.55 ± 2.89 ab | 102.59 ± 3.47 ab | 85.64 ± 4.64 cde | 80.84 ± 4.23 de | 75.89 ± 1.21 e | 77.26 ± 2.07 e | 19.96 | 10 |
| Trp | 84.93 ± 3.59 ab | 84.61 ± 1.83 ab | 85.82 ± 1.36 a | 81.61 ± 5.27 ab | 84.30 ± 1.39 ab | 80.23 ± 0.64 ab | 52.31 ± 2.26 b | 0.00 ± 0.00 c | 0.00 ± 0.00 c | 0.00 ± 0.00 c | 0.00 ± 0.00 c | 0.00 ± 0.00 c | 0.00 ± 0.00 c | 0.90 | 4 |
| Tyr | 79.29 ± 2.54 a | 78.88 ± 0.35 ab | 79.37 ± 1.47 a | 77.60 ± 7.73 ab | 74.00 ± 5.74 abcd | 75.49 ± 2.52 abc | 74.11 ± 2.59 abcd | 77.91 ± 4.83 ab | 76.21 ± 2.57 abc | 66.93 ± 5.01 bcde | 65.18 ± 1.46 cde | 63.07 ± 0.91 de | 59.45 ± 1.41 e | 0.09 | 4 |
| Ser | 39.85 ± 2.08 b | 50.8 ± 5.40 ab | 55.59 ± 2.22 a | 52.74 ± 7.98 a | 50.01 ± 3.96 ab | 51.37 ± 3.48 ab | 47.88 ± 2.73 ab | 49.45 ± 2.68 ab | 53.45 ± 3.14 a | 47.44 ± 5.11 ab | 45.96 ± 0.82 ab | 43.04 ± 1.70 ab | 46.91 ± 0.50 ab | 15.74 | 4 |
| Gly | 35.07 ± 2.23 d | 42.47 ± 3.26 cd | 43.12 ± 1.24 cd | 51.84 ± 9.00 abc | 56.02 ± 5.56 abc | 54.89 ± 4.55 abc | 55.57 ± 3.64 abc | 57.41± 2.93 ab | 64.94 ± 3.31 a | 55.60 ± 6.30 abc | 53.85 ± 0.19 abc | 47.27 ± 2.35 bcd | 51.46 ± 0.79 abc | 29.87 | 16 |
| Cy2 | 120.10 ± 7.89 cde | 127.24 ± 3.26 cd | 107.54 ± 2.98 e | 123.89 ± 8.17 cde | 131.89 ± 5.12 bcd | 127.89 ± 5.00 bcd | 134.96 ± 5.79 abc | 126.14 ± 2.36 cd | 151.49 ± 9.58 a | 137.09 ± 8.00 abc | 146.08 ± 2.14 ab | 113.83 ± 3.54 de | 127.56 ± 2.89 bcd | 31.39 | 16 |
| AA (mg/L) | Incubation Time (h) | Maximum Increment | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 | 2 | 4 | 6 | 8 | 10 | 12 | 14 | 16 | 18 | 20 | 22 | 24 | Amount (mg/L) | Time (h) | |
| Asp | 62.02 ± 1.33 a | 61.29 ± 3.42 a | 60.27 ± 2.06 a | 61.23 ± 0.89 a | 61.56 ± 2.36 a | 62.11 ± 7.02 a | 61.81 ± 2.51 a | 62.79 ± 3.85 a | 60.95 ± 2.39 a | 62.13 ± 0.95 a | 60.65 ± 2.95 a | 60.35 ± 0.94 a | 60.24 ± 2.10 a | 0.77 | 14 |
| Asn | 76.51 ± 3.89 a | 77.15 ± 2.63 a | 75.96 ± 1.67 a | 75.72 ± 1.47 a | 76.33 ± 2.28 a | 75.74 ± 8.06 a | 76.98 ± 2.33 a | 76.52 ± 4.95 a | 76.14 ± 6.90 a | 76.60 ± 0.72 a | 75.24 ± 5.43 a | 76.17 ± 1.81 a | 76.40 ± 0.77 a | 0.64 | 2 |
| Lys | 367.47 ± 7.30 a | 340.81 ± 10.05 ab | 328.24 ± 1.66 b | 336.36 ± 9.21 ab | 323.94 ± 5.35 b | 311.66 ± 10.51 b | 320.94 ± 7.37 b | 315.93 ± 4.08 b | 316.68 ± 3.82 b | 320.71 ± 5.49 b | 311.92 ± 34.43 b | 318.66 ± 7.01 b | 328.29 ± 4.99 b | 0.00 | 0 |
| Thr | 70.78 ± 1.11 a | 70.77 ± 2.33 a | 64.77 ± 3.75 ab | 61.15 ± 2.72 abc | 65.38 ± 2.40 ab | 61.81 ± 6.89 abc | 57.20 ± 2.52 bc | 5.7.84 ± 1.11 bc | 57.05 ± 3.75 bc | 58.11 ± 2.40 bc | 53.31 ± 2.51 c | 59.99 ± 1.08 bc | 58.28 ± 0.40 bc | 0.00 | 0 |
| Met | 106.52 ± 0.68 c | 111.07 ± 2.97 bc | 105.76 ± 1.52 c | 116.59 ± 4.07 abc | 119.22 ± 7.06 ab | 120.48 ± 7.60 ab | 124.61 ± 0.42 a | 126.33 ± 3.95 a | 125.74 ± 4.46 a | 12.8.45 ± 2.63 a | 126.58 ± 1.67 a | 128.03 ± 3.22 a | 129.72 ± 3.87 a | 23.21 | 24 |
| Ile | 99.15 ± 4.71 f | 116.86 ± 3.65 e | 120.81 ± 1.22 de | 133.82 ± 3.33 bc | 150.22 ± 1.30 a | 145.91 ± 4.15 a | 149.50 ± 0.99 a | 142.32 ± 1.55 ab | 125.08 ± 2.39 cde | 126.95 ± 6.94 cde | 121.33 ± 5.38 de | 129.83 ± 5.60 cd | 143.38 ± 2.12 ab | 51.07 | 8 |
| Glu | 228.52 ± 4.89 cd | 234.65 ± 8.34 bcd | 225.80 ± 8.34 d | 237.21 ± 8.94 bcd | 254.74 ± 8.18 abcd | 260.23 ± 24.95 ab | 269.48 ± 4.44 a | 256.69 ± 4.85 abcd | 249.90 ± 8.93 abcd | 251.99 ± 3.29 abcd | 245.09 ± 0.50 abcd | 248.68 ± 6.62 abcd | 258.82 ± 1.18 abc | 40.96 | 12 |
| Pro | 50.25 ± 1.12 g | 50.52 ± 0.21 g | 52.03 ± 0.34 g | 52.75 ± 0.40 g | 62.31 ± 0.70 f | 67.25 ± 0.45 e | 71.58 ± 0.92 d | 70.91 ± 0.26 d | 74.10 ± 1.21 c | 77.91 ± 0.77 b | 79.29 ± 0.49 b | 82.08 ± 1.97 a | 81.95 ± 0.51 a | 31.83 | 22 |
| Arg | 186.26 ± 6.63 a | 177.25 ± 7.49 ab | 177.56 ± 1.55 ab | 178.02 ± 17.62 ab | 170.27 ± 10.76 ab | 151.44 ± 10.96 bc | 133.47 ± 2.92 c | 28.1 ± 28.1 d | 0.00 ± 0.00 d | 0.00 ± 0.00 d | 0.00 ± 0.00 d | 0.00 ± 0.00 d | 0.00 ± 0.00 d | 0.00 | 0 |
| Ala | 193.89 ± 3.22 ab | 188.45 ± 5.76 b | 186.47 ± 3.06 b | 188.76 ± 3.15 b | 193.35 ± 6.54 b | 199.21 ± 8.51 ab | 199.89 ± 2.79 ab | 198.50 ± 3.67 ab | 195.47 ± 8.94 ab | 189.69 ± 1.63 b | 195.45 ± 0.80 ab | 202.00 ± 3.28 ab | 209.46 ± 1.00 a | 15.57 | 24 |
| Val | 78.85 ± 1.65 c | 84.08 ± 0.85 c | 82.82 ± 4.24 c | 91.75 ± 3.78 b | 98.16 ± 3.02 ab | 98.05 ± 3.47 ab | 102.03 ± 1.70 a | 97.27 ± 1.41 ab | 95.78 ± 3.10 ab | 100.75 ± 2.23 a | 95.81 ± 0.79 ab | 99.20 ± 3.51 ab | 103.39 ± 1.00 a | 24.54 | 24 |
| Leu | 361.65± 26.99 e | 406.78 ± 5.18 d | 409.97 ± 7.52 cd | 415.55 ± 8.64 bcd | 422.73 ± 3.19 abcd | 442.52 ± 6.54 abc | 449.45 ± 6.65 a | 444.06 ± 6.79 ab | 417.34 ± 9.89 abcd | 421.27 ± 6.03 abcd | 439.83 ± 8.29 abc | 426.01 ± 3.08 abcd | 435.13 ± 1.37 abcd | 87.80 | 12 |
| Phe | 121.39 ± 4.97 a | 115.87 ± 3.79 ab | 112.87 ± 0.62 abc | 118.34 ± 4.65 a | 115.55 ± 0.20 ab | 105.51 ± 0.37 bc | 110.20 ± 4.18 abc | 103.73 ± 4.66 c | 92.51 ± 2.10 d | 93.31 ± 4.90 d | 90.56 ± 5.22 d | 91.50 ± 1.78 d | 89.74 ± 1.10 d | 0.00 | 0 |
| Tyr | 81.40 ± 1.31 ab | 77.76 ± 2.18 b | 77.20 ± 2.41 b | 77.07 ± 1.19 b | 64.45 ± 32.63 a | 60.86 ± 32.53 b | 57.38 ± 28.76 b | 51.42 ± 25.71 b | 25.78 ± 25.78 c | 0.00 ± 0.00 c | 0.00 ± 0.00 c | 0.00 ± 0.00 c | 0.00 ± 0.00 c | 0.00 | 0 |
| Trp | 123.86 ± 4.74 a | 116.10 ± 2.91 abc | 116.42 ± 1.72 abc | 107.48 ± 2.26 c | 120.78 ± 6.55 ab | 83.15 ± 44.36 bc | 0.00 ± 0.00 d | 0.00 ± 0.00 d | 0.00 ± 0.00 d | 0.00 ± 0.00 d | 0.00 ± 0.00 d | 0.00 ± 0.00 d | 0.00 ± 0.00 d | 0.00 | 0 |
| Ser | 115.91 ± 1.46 de | 114.70 ± 4.83 e | 112.19 ± 1.6 e | 119.61 ± 3.2 de | 128.27± 6.59 cde | 125.69 ± 9.25 cde | 135.37 ± 4.76 bcd | 129.04 ± 4.95 cde | 127.03 ± 7.07 cde | 131.53 ± 1.75 cde | 144.44 ± 8.05 abc | 150.31 ± 8.74 ab | 161.43 ± 6.28 a | 45.52 | 24 |
| Gly | 89.67 ± 5.90 de | 85.66 ± 4.57 e | 94.94 ± 2.01 de | 90.33 ± 4.15 de | 98.44 ± 8.06 cde | 97.25 ± 8.53 cde | 108.25 ± 8.25 bcd | 104.17 ± 5.06 bcde | 108.72 ± 4.19 bcd | 116.42 ± 5.98 bc | 116.66 ± 4.46 bc | 135.69 ± 6.59 a | 121.28 ± 5.93 ab | 46.02 | 22 |
| Cy2 | 163.78 ± 4.44 ab | 159.38 ± 6.14 ab | 153.75 ± 10.34 b | 158.12 ± 3.27 ab | 168.81 ± 11.24 ab | 175.34 ± 21.02 ab | 184.80 ± 8.32 ab | 179.93 ± 7.02 ab | 193.40 ± 18.37 ab | 186.42 ± 16.46 ab | 193.74 ± 20.43 ab | 204.47 ± 14.54 a | 201.38 ± 18.08 a | 40.69 | 22 |
| AA (mg/L) | Incubation Time (h) | Maximum Increment | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 | 2 | 4 | 6 | 8 | 10 | 12 | 14 | 16 | 18 | 20 | 22 | 24 | Amount (mg/L) | Time (h) | |
| Asp | 40.89 ± 0.29 b | 40.57 ± 1.91 b | 41.61 ± 1.26 b | 47.47 ± 1.12 a | 39.66 ± 0.46 b | 35.93 ± 0.46 c | 32.47 ± 1.00 d | 0.00 ± 0.00 e | 0.00 ± 0.00 e | 0.00 ± 0.00 e | 0.00 ± 0.00 e | 0.00 ± 0.00 e | 0.00 ± 0.00 e | 6.58 | 6 |
| Asn | 48.90 ± 0.40 b | 50.20 ± 0.59 a | 40.99 ± 0.03 c | 49.15 ± 0.68 b | 0.00 ± 0.00 d | 0.00 ± 0.00 d | 0.00 ± 0.00 d | 0.00 ± 0.00 d | 0.00 ± 0.00 d | 0.00 ± 0.00 d | 0.00 ± 0.00 d | 0.00 ± 0.00 d | 0.00 ± 0.00 d | 1.30 | 2 |
| Lys | 357.18 ± 1.82 a | 350.95 ± 8.79 ab | 351.82 ± 4.79 ab | 359.05 ± 16.93 a | 343.97 ± 2.91 abc | 328.77 ± 5.03 bcd | 333.41 ± 3.81 bc | 325.66 ± 10.47 cd | 321.30 ± 1.45 cde | 324.01 ± 1.37 cde | 305.18 ± 6.89 def | 295.97 ± 9.95 f | 301.17 ± 3.05 ef | 1.87 | 6 |
| Met | 91.27 ± 0.66 c | 93.79 ± 2.93 bc | 94.83 ± 1.74 abc | 95.83 ± 1.82 abc | 99.87 ± 1.44 a | 96.93 ± 2.06 abc | 97.81 ± 1.26 ab | 95.55 ± 2.36 abc | 96.10 ± 0.89 abc | 94.69 ± 1.05 abc | 91.30 ± 0.03 c | 91.61± 0.87 c | 91.60 ± 2.34 c | 8.61 | 8 |
| Thr | 66.37 ± 3.39 b | 62.55 ± 5.40 b | 63.53 ± 4.71 b | 83.05 ± 2.25 a | 65.15 ± 6.01 b | 63.78 ± 2.52 b | 57.71 ± 4.21 bc | 47.71 ± 1.70 cd | 47.7 ± 0.78 cd | 43.47 ± 1.19 d | 41.79 ± 1.47 d | 38.97 ± 0.93 d | 38.52 ± 2.77 d | 16.67 | 6 |
| Ile | 87.50 ± 0.90 g | 89.89 ± 1.87 fg | 92.77 ± 0.07 f | 112.95 ± 0.73 a | 103.58 ± 2.16 bc | 102.47 ± 1.85 cd | 107.08 ± 1.45 b | 101.0 ± 0.92 cde | 103.8 ± 1.47 bc | 101.9 ± 1.48 d | 98.3 ± 1.38 de | 97.3 ± 1.18 e | 98.7 ± 1.32 de | 25.45 | 6 |
| Glu | 215.74 ± 1.50 g | 215.52 ± 5.70 g | 220.18 ± 2.23 fg | 253.11 ± 3.51 a | 248.47 ± 6.16 ab | 243.75 ± 4.89 abc | 248.81 ± 3.49 ab | 238.37 ± 25.1 bcde | 241.0 ± 1.85 bcd | 232.9 ± 2.58 cde | 228.9 ± 5.13 ef | 229.5 ± 1.60 def | 227.2 ± 2.28 ef | 37.38 | 6 |
| Pro | 36.49 ± 0.59 c | 36.72 ± 0.11 c | 35.95 ± 0.53 c | 37.27 ± 3.06 c | 37.78 ± 1.47 c | 39.37 ± 1.12 bc | 45.46 ± 0.74 a | 42.75 ± 1.15 ab | 43.02 ± 1.25 ab | 44.74 ± 0.93 a | 43.90 ± 1.62 a | 44.38 ± 1.50 a | 42.55 ± 0.78 ab | 8.97 | 12 |
| Arg | 203.69 ± 1.60 abc | 208.44 ± 1.26 ab | 210.51 ± 2.14 a | 203.07± 6.41 abc | 200.21 ± 2.81 bcd | 192.06 ± 2.98 def | 195.50 ± 1.51 cde | 193.80 ± 3.01 cde | 195.70 ± 1.98 cde | 189.61 ± 1.05 ef | 176.24 ± 6.19 h | 173.33 ± 2.99 h | 183.62 ± 0.78 fg | 6.81 | 4 |
| Ala | 165.93 ± 1.56 b | 165.34 ± 1.21 b | 165.65 ± 0.98 b | 176.29 ± 2.07 a | 162.34 ± 0.84 b | 147.74 ± 2.27 c | 145.05 ± 2.55 cd | 137.86 ± 1.55 d | 136.66 ± 1.08 d | 125.33 ± 1.79 e | 116.43 ± 7.23 f | 117.07 ± 4.67 f | 115.21 ± 1.54 f | 10.37 | 6 |
| Val | 111.28 ± 0.91 d | 112.01 ± 1.76 d | 113.96 ± 1.70 d | 126.04± 1.10 abc | 125.31 ± 1.18 bc | 124.29 ± 2.62 bc | 131.39 ± 2.00 a | 127.34 ± 2.61 ab | 126.14± 0.37 abc | 122.83 ± 2.81 bc | 122.15 ± 1.05 bc | 120.30 ± 1.59 c | 121.16 ± 2.26 c | 20.11 | 12 |
| Leu | 346.55 ± 5.00 bcde | 349.15 ± 4.25 bcd | 356.35 ± 0.32 ab | 365.34 ± 6.26 a | 351.18 ± 2.97 bc | 336.63 ± 2.35 cdef | 339.97 ± 4.10 cdef | 332.21 ± 2.11 efg | 336.02 ± 4.34 def | 329.73 ± 3.58 fgh | 318.26 ± 8.97 gh | 315.71 ± 5.90 h | 321.26 ± 2.66 gh | 18.79 | 6 |
| Phe | 159.89 ± 0.91 ab | 160.54 ± 1.36 b | 163.07 ± 0.80 a | 154.53 ± 4.45 bc | 154.95 ± 3.52 bc | 148.13 ± 2.02 de | 152.58 ± 2.21 cd | 147.77 ± 0.35 de | 145.02 ± 0.86 e | 143.72 ± 0.80 e | 137.28 ± 1.93 f | 136.25 ± 1.17 f | 136.85 ± 2.05 f | 3.21 | 4 |
| Trp | 80.23 ± 00.67 a | 77.49 ± 0.68 b | 77.21 ± 1.69 b | 78.35 ± 1.55 ab | 72.37 ± 0.26 c | 70.36 ± 0.49 c | 72.02 ± 0.88 c | 0.00 ± 0.00 d | 0.00 ± 0.00 d | 0.00 ± 0.00 d | 0.00 ± 0.00 d | 0.00 ± 0.00 d | 0.00 ± 0.00 d | 0.00 | 0 |
| Tyr | 65.84 ± 0.41 a | 62.03 ± 0.58 abc | 64.05 ± 0.55 ab | 55.23 ± 3.89 def | 62.28 ± 1.21 abc | 57.77 ± 0.99 cd | 56.92 ± 1.12 cde | 55.88 ± 1.25 de | 58.14 ± 0.74 bcd | 53.38 ± 0.97 def | 51.00 ± 3.52 ef | 49.68 ± 3.35 f | 51.09 ± 0.45 ef | 0.00 | 0 |
| Ser | 114.44 ± 1.49 b | 113.36 ± 0.16 b | 113.00 ± 0.35 b | 139.32 ± 3.04 a | 0.00 ± 0.00 c | 0.00 ± 0.00 c | 0.00 ± 0.00 c | 0.00 ± 0.00 c | 0.00 ± 0.00 c | 0.00 ± 0.00 c | 0.00 ± 0.00 c | 0.00 ± 0.00 c | 0.00 ± 0.00 c | 24.88 | 6 |
| Gly | 67.04 ± 0.90 cde | 63.40 ± 0.33 e | 64.15 ± 0.16 de | 78.20 ± 2.95 ab | 71.15 ± 0.30 de | 73.11 ± 0.70 abcde | 76.37 ± 0.95 abc | 77.07 ± 1.38 abc | 82.19 ± 2.46 a | 76.98 ± 2.50 abc | 73.99 ± 7.97 abcd | 75.49 ± 5.15 abc | 78.38 ± 2.58 ab | 15.15 | 16 |
| Cy2 | 128.81 ± 5.42 a | 100.84 ± 3.63 bc | 101.21 ± 5.18 bc | 113.18 ± 2.96 ab | 93.72 ± 0.08 bcd | 87.01± 1.97 cde | 84.13 ± 3.50 cde | 74.43 ± 2.45 de | 87.54 ± 1.46 cde | 78.56 ± 11.14 de | 72.66 ± 9.04 de | 69.01 ± 16.07 e | 76.44 ± 2.62 de | 0.00 | 0 |
| AA (mg/L) | Incubation Time (h) | Maximum Increment | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 | 2 | 4 | 6 | 8 | 10 | 12 | 14 | 16 | 18 | 20 | 22 | 24 | Amount (mg/L) | Time (h) | |
| Asp | 47.12 ± 3.94 a | 46.56 ± 6.06 a | 45.80 ± 2.38 a | 46.37 ± 3.48 a | 47.64 ± 5.93 a | 46.88 ± 7.20 a | 46.23 ± 6.17 a | 47.92 ± 5.75 a | 45.75 ± 3.70 a | 47.64 ± 7.27 a | 46.58 ± 5.69 a | 48.90 ± 2.22 a | 45.43 ± 4.88 a | 1.79 | 22 |
| Asn | 53.18 ± 4.44 a | 49.43 ± 4.11 a | 55.91 ± 3.12 a | 51.47 ± 5.69 a | 56.47 ± 4.61 a | 50.82 ± 7.45 a | 51.27 ± 4.47 a | 52.78 ± 8.23 a | 50.03 ± 5.57 a | 57.47 ± 9.13 a | 57.41 ± 8.16 a | 58.98 ± 2.70 a | 52.40 ± 7.12 a | 5.80 | 22 |
| Lys | 369.72 ± 7.13 a | 311.54 ± 1.07 b | 307.32 ± 7.88 b | 275.70 ± 7.99 c | 270.01 ± 5.27 c | 240.95 ± 4.78 d | 272.85 ± 5.47 c | 235.75 ± 5.25 d | 203.09 ± 8.77 e | 236.22 ± 4.36 d | 239.71 ± 3.74 d | 252.87 ± 2.55 d | 236.28 ± 6.19 d | 0.00 | 0 |
| Met | 97.00 ± 2.02 ab | 96.03 ± 8.26 ab | 100.09 ± 8.74 ab | 102.82 ± 3.50 ab | 111.37 ± 7.46 a | 101.72 ± 3.09 ab | 108.77 ± 12.08 a | 106.36 ± 3.31 a | 83.41 ± 0.58 b | 59.69 ± 8.51 c | 59.20 ± 8.47 c | 60.78 ± 1.62 c | 54.33 ± 2.51 c | 14.37 | 8 |
| Thr | 71.61 ± 4.82 a | 70.04 ± 6.00 a | 70.70± 8.20 a | 71.24 ± 7.18 a | 66.06 ± 8.86 ab | 65.60 ± 5.35 ab | 61.36 ± 1.20 ab | 62.82 ± 3.09 ab | 59.46 ± 0.68 ab | 55.39 ± 9.26 ab | 55.10 ± 4.28 ab | 52.68 ± 5.25 ab | 48.93 ± 3.91 b | 0.00 | 0 |
| Ile | 113.02 ± 2.43 bc | 99.46 ± 7.20 c | 102.88 ± 4.70 c | 99.30 ± 8.63 c | 104.65 ± 6.45 c | 126.34 ± 1.42 ab | 129.62 ± 4.81 a | 102.43 ± 4.83 c | 78.81 ± 2.42 d | 72.54 ± 7.05 d | 75.60 ± 4.79 d | 74.53 ± 1.43 d | 67.85 ± 2.03 d | 16.60 | 12 |
| Glu | 205.26 ± 2.95 cde | 194.95 ± 5.30 e | 205.46± 9.36 cde | 234.70 ± 6.66 ab | 244.13 ± 9.57 a | 200.55 ± 3.22 de | 230.05 ± 9.16 ab | 216.88 ± 4.14 bcd | 203.30 ± 7.06 cde | 222.50 ± 5.17 bc | 226.22 ± 2.93 ab | 216.45± 6.08 bcd | 191.70 ± 4.15 e | 38.87 | 8 |
| Pro | 47.65 ± 2.36 d | 46.02 ± 2.03 d | 45.23 ± 3.80 d | 46.34 ± 1.08 d | 47.41 ± 1.28 d | 56.14 ± 7.79 d | 60.25 ± 6.48 d | 122.09 ± 3.86 c | 125.27 ± 6.64 c | 149.27 ± 4.72 b | 158.85 ± 5.05 ab | 163.71 ± 8.46 ab | 169.53 ± 6.47 a | 121.88 | 24 |
| Arg | 226.39 ± 3.50 a | 216.13 ± 4.53 b | 208.92 ± 7.25 b | 164.05 ± 5.45 c | 118.73 ± 5.21 d | 0.00 ± 0.00 e | 0.00 ± 0.00 e | 0.00 ± 0.00 e | 0.00 ± 0.00 e | 0.00 ± 0.00 e | 0.00 ± 0.00 e | 0.00 ± 0.00 e | 0.00 ± 0.00 e | 0.00 | 0 |
| Ala | 176.49 ± 8.07 a | 165.51 ± 6.97 a | 156.90 ± 7.09 a | 157.70 ± 5.94 a | 156.33 ± 6.14 a | 119.49 ± 9.41 bc | 134.15 ± 5.89 b | 123.49 ± 9.01 bc | 106.18 ± 5.22 cd | 125.95 ± 5.36 bc | 130.30 ± 6.51 b | 120.63 ± 0.87 bc | 92.74 ± 4.52 d | 0.00 | 0 |
| Val | 56.46 ± 2.33 cd | 53.42 ± 6.11 d | 59.24 ± 8.72 bcd | 63.58 ± 1.92 abcd | 81.39 ± 3.76 a | 64.53 ± 6.78 abcd | 62.98 ± 1.45 bcd | 68.59 ± 7.37 abcd | 67.50 ± 3.64 abcd | 74.98 ± 9.06 ab | 74.44 ± 3.57 abc | 76.13 ± 5.80 ab | 75.82 ± 1.14 ab | 24.93 | 8 |
| Leu | 392.77 ± 1.94 a | 344.25 ± 3.80 b | 330.83 ± 7.26 bc | 301.26 ± 6.73 d | 295.62 ± 9.41 d | 260.28 ± 4.88 e | 256.08 ± 7.71 e | 267.76 ± 7.25 e | 254.69 ± 3.47 e | 255.55 ± 6.34 e | 269.14 ± 9.42 e | 313.59 ± 9.04 cd | 270.71 ± 5.85 e | 0.00 | 0 |
| Phe | 84.74 ± 2.38 cde | 76.96 ± 8.91 e | 84.15 ± 4.09 de | 99.94 ± 7.23 bcd | 99.46 ± 4.53 bcd | 113.35 ± 8.62 ab | 124.66 ± 3.68 a | 102.93 ± 6.92 bcd | 90.41 ± 4.16 cde | 104.67 ± 9.03 bc | 101.71 ± 3.96 bcd | 102.26 ± 3.02 bcd | 97.32 ± 5.60 bcd | 39.92 | 12 |
| Trp | 83.33 ± 6.13 a | 82.09 ± 4.04 a | 86.71 ± 3.29 a | 85.54 ± 7.92 a | 82.30 ± 12.60 a | 61.62 ± 0.96 a | 56.67 ± 6.22 a | 0.00 ± 0.00 b | 0.00 ± 0.00 b | 0.00 ± 0.00 b | 0.00 ± 0.00 b | 0.00 ± 0.00 b | 0.00 ± 0.00 b | 3.39 | 4 |
| Tyr | 79.4 ± 5.71 a | 66.45 ± 9.00 b | 75.92 ± 0.47 b | 78.36 ± 2.49 b | 78.12 ± 2.68 b | 73.46 ± 2.71 b | 76.19 ± 2.76 b | 77.74 ± 1.06 b | 73.35 ± 1.05 b | 77.61 ± 2.93 b | 77.06 ± 4.57 b | 76.48 ± 1.32 b | 70.95 ± 4.89 ab | 0.00 | 0 |
| Ser | 110.30 ± 5.27 a | 90.27 ± 9.41 ab | 93.66 ± 8.79 ab | 89.92 ± 7.66 ab | 89.72 ± 5.42 ab | 62.44 ± 8.25 c | 83.77 ± 6.90 b | 60.53 ± 9.00 c | 55.75 ± 2.37 c | 63.20 ± 2.11 c | 61.84 ± 4.82 c | 54.90 ± 2.38 c | 54.95 ± 6.49 c | 0.00 | 0 |
| Gly | 62.36 ± 5.0 d | 93.92 ± 5.72 c | 94.10 ± 6.73 c | 122.11 ± 2.40 ab | 133.42 ± 6.17 a | 111.73 ± 6.88 bc | 108.26 ± 8.35 bc | 107.08 ± 4.73 bc | 100.14 ± 8.62 c | 99.12 ± 8.55 c | 74.08 ± 7.18 d | 68.58 ± 4.53 d | 60.51 ± 2.28 d | 71.06 | 8 |
| Cy2 | 155.90 ± 2.81 bcd | 170.93 ± 3.80 ab | 160.30 ± 3.68 bcd | 166.23 ± 7.44 abc | 180.94 ± 8.66 a | 160.21 ± 6.26 bcd | 158.18 ± 9.73 bcd | 164.89 ± 6.88 abc | 136.12 ± 1.21 ef | 148.29 ± 1.88 cde | 130.73 ± 1.63 ef | 144.15 ± 7.63 de | 124.15 ± 7.82 f | 25.04 | 8 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Toe, C.J.; Foo, H.L.; Loh, T.C.; Mohamad, R.; Abdul Rahim, R.; Idrus, Z. Extracellular Proteolytic Activity and Amino Acid Production by Lactic Acid Bacteria Isolated from Malaysian Foods. Int. J. Mol. Sci. 2019, 20, 1777. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20071777
Toe CJ, Foo HL, Loh TC, Mohamad R, Abdul Rahim R, Idrus Z. Extracellular Proteolytic Activity and Amino Acid Production by Lactic Acid Bacteria Isolated from Malaysian Foods. International Journal of Molecular Sciences. 2019; 20(7):1777. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20071777
Chicago/Turabian StyleToe, Cui Jin, Hooi Ling Foo, Teck Chwen Loh, Rosfarizan Mohamad, Raha Abdul Rahim, and Zulkifli Idrus. 2019. "Extracellular Proteolytic Activity and Amino Acid Production by Lactic Acid Bacteria Isolated from Malaysian Foods" International Journal of Molecular Sciences 20, no. 7: 1777. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20071777