Prediction of Bioactive Peptides from Chlorella sorokiniana Proteins Using Proteomic Techniques in Combination with Bioinformatics Analyses


Abstract
:
1. Introduction
2. Results and Discussion
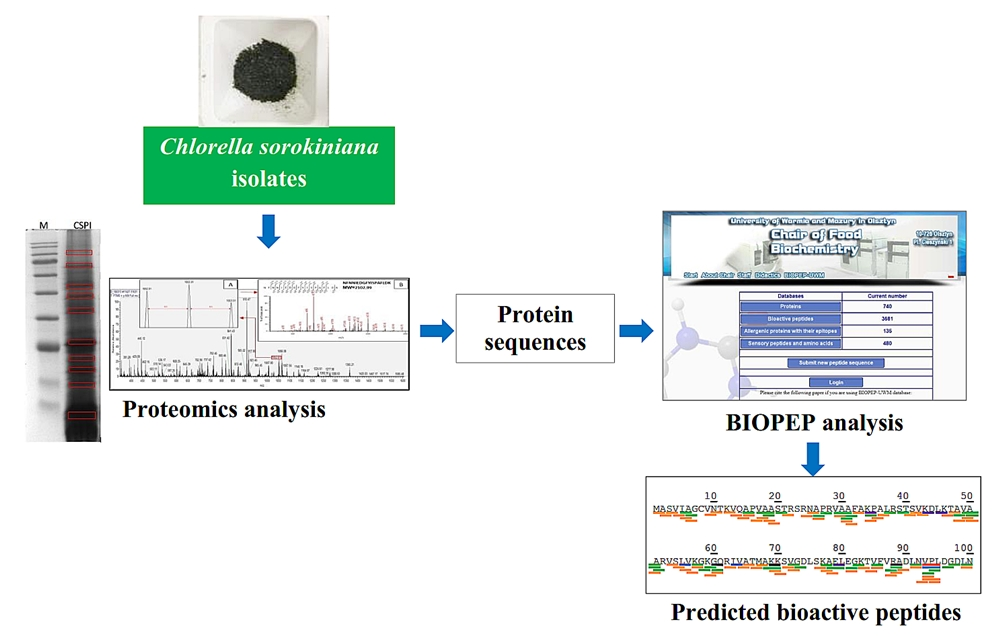
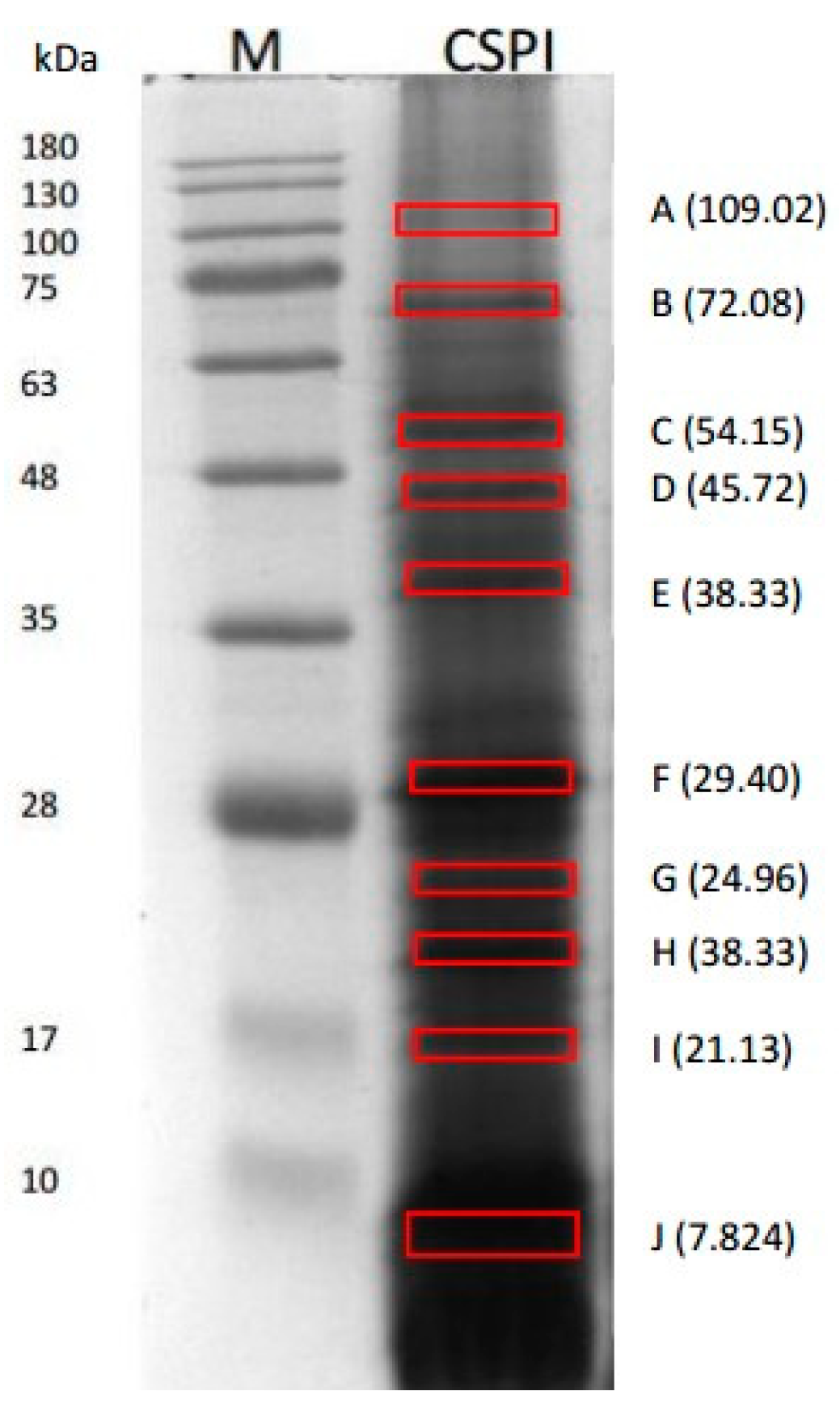
2.1. Identified Proteins of C. sorokiniana
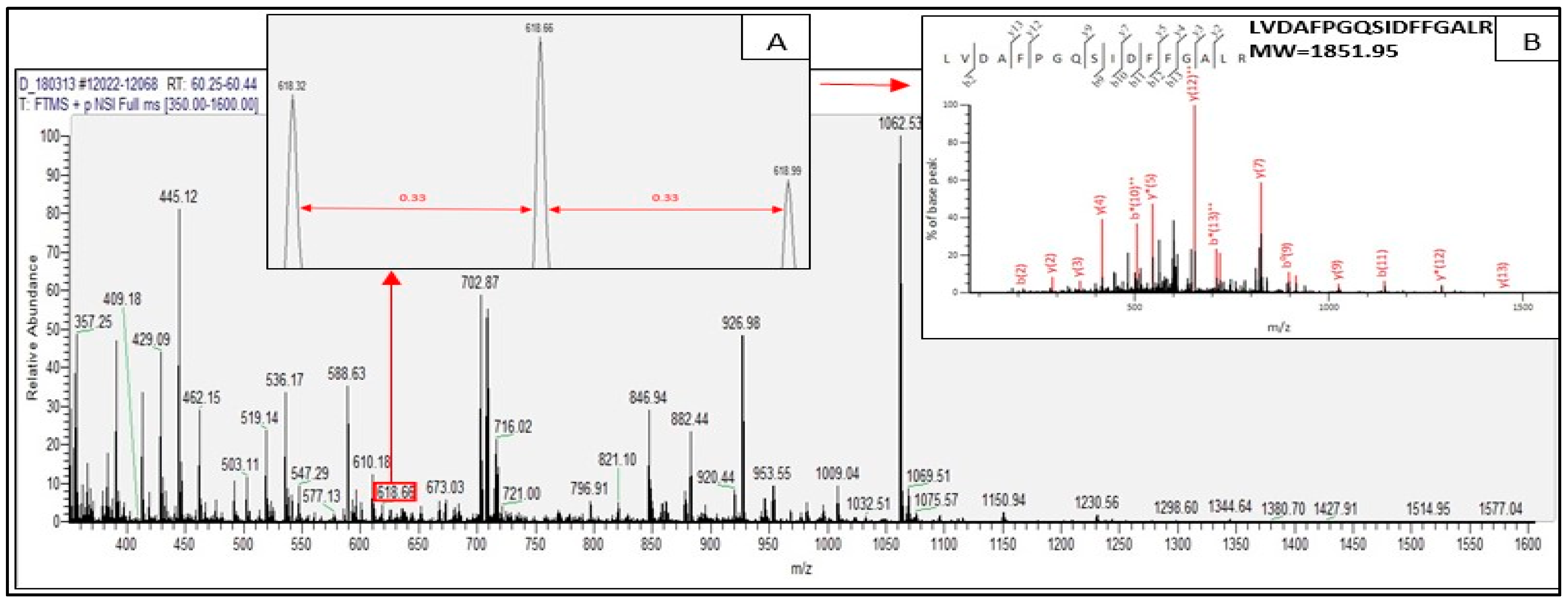
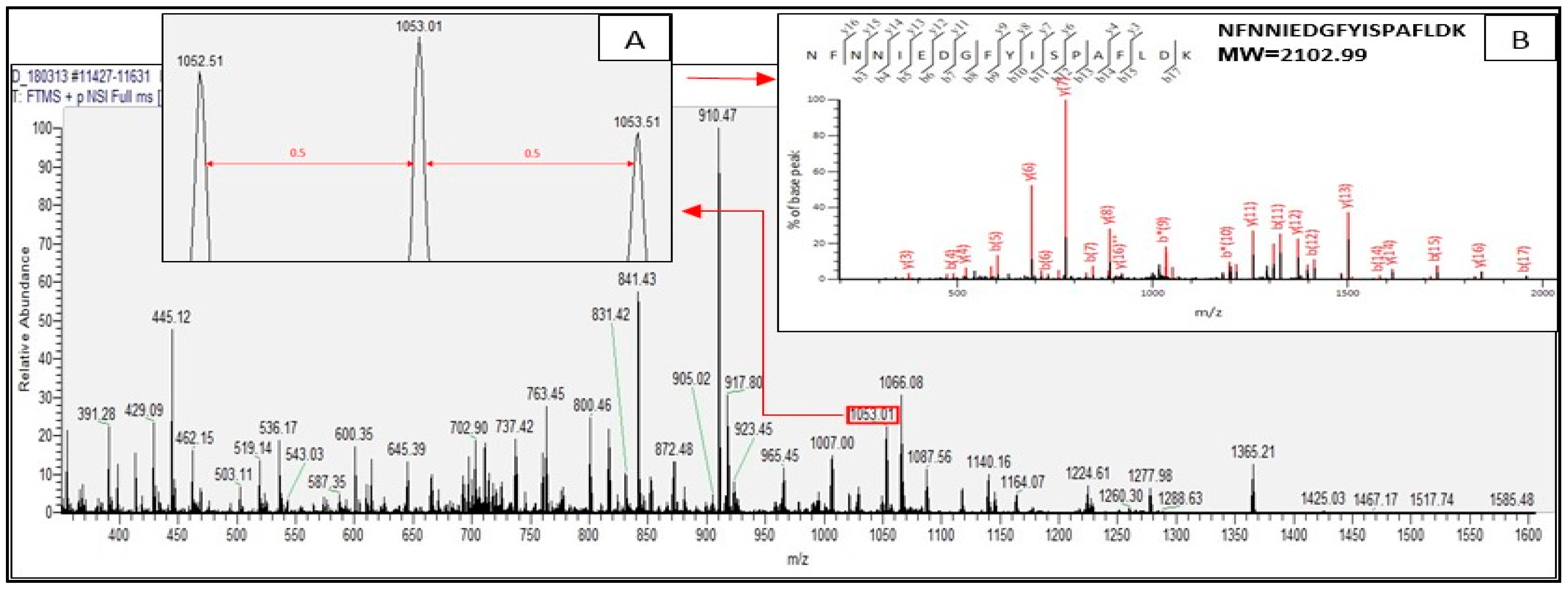
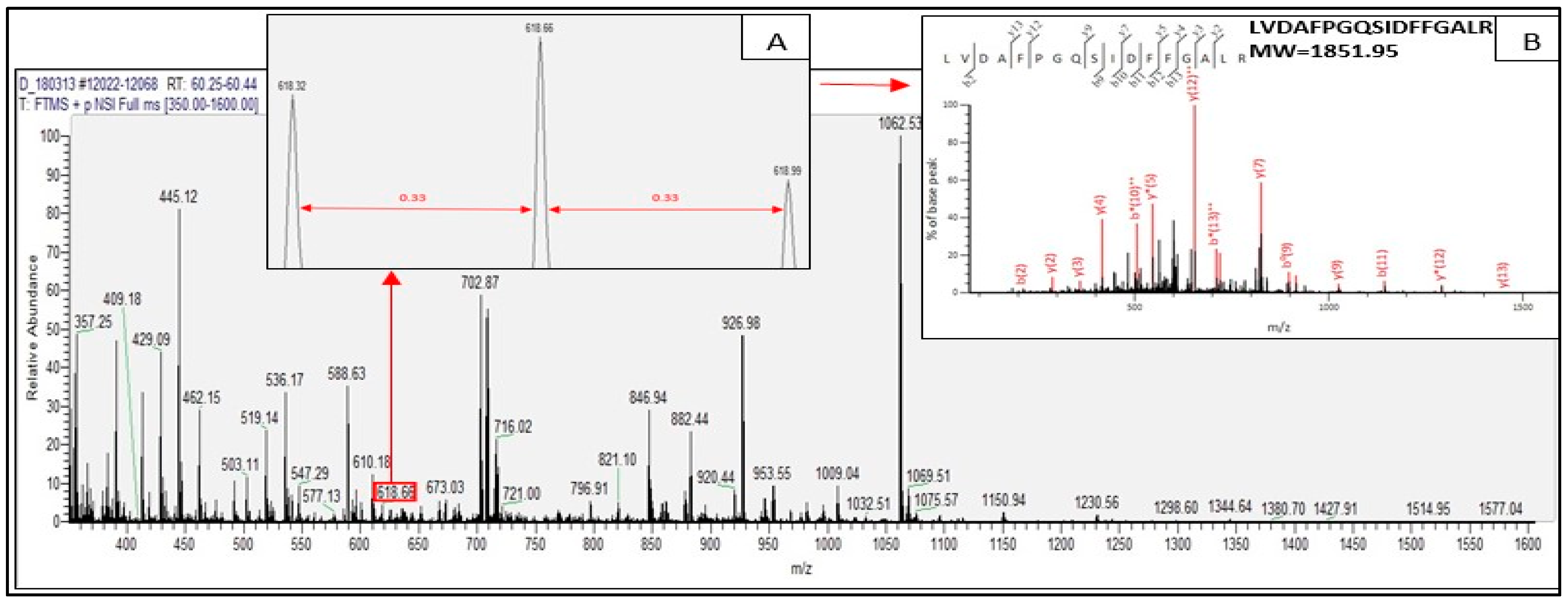
2.2. Identified Tryptic Peptides from C. sorokiniana Proteins
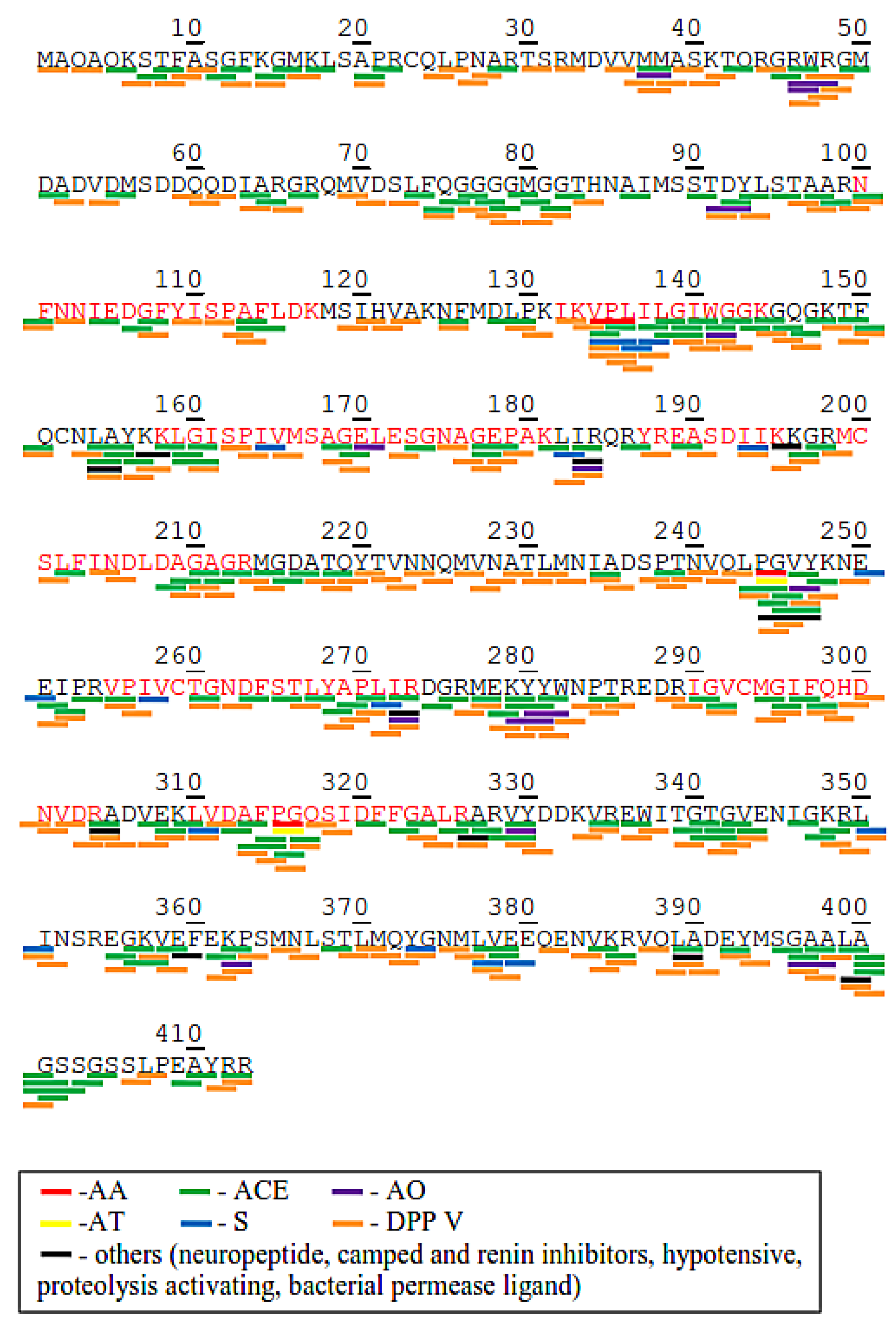
2.3. Potential Bioactive Peptides from Identified Proteins in C. sorokiniana
2.4. Prediction of Potential Bioactive Peptides after Protease Cleavage using BIOPEP-UWM Tool
3. Materials and Methods
3.1. Materials
3.2. Protein Isolation
3.3. SDS-PAGE Analysis
3.4. Proteomics Techniques
3.4.1. In-Gel Tryptic Digestion
3.4.2. Nanoliquid Chromatography–Nanoelectrospray Ionization Tandem Mass Spectrometry (NanoLC–nanoESI MS/MS) Analysis
3.4.3. Tandem MS Data Analysis of Proteins and Peptide Identification
3.4.4. In Silico Analysis of Bioactive Peptides and Enzyme Cleavages using BIOPEP-UWM Database Tools
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Spolaore, P.; Joannis-Cassan, C.; Duran, E.; Isambert, A. Commercial applications of microalgae. J. Biosci. Bioeng. 2006, 101, 87–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mubarak, M.; Shaija, A.; Suchithra, T. A review on the extraction of lipid from microalgae for biodiesel production. Algal Res. 2015, 7, 117–123. [Google Scholar] [CrossRef]
- Schwenzfeier, A.; Wierenga, P.A.; Gruppen, H. Isolation and characterization of soluble protein from the green microalgae Tetraselmis sp. Bioresour. Technol. 2011, 102, 9121–9127. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.; Chen, X.; Wu, J.; Zhou, Y.; Qian, Y.; Fang, M.; Xie, J.; Wei, D. Dipeptidyl peptidase IV inhibitory peptides from Chlorella vulgaris: In silico gastrointestinal hydrolysis and molecular mechanism. Eur. Food Res. Technol. 2017, 243, 1739–1748. [Google Scholar] [CrossRef]
- Kralovec, J.; Metera, K.; Kumar, J.; Watson, L.; Girouard, G.; Guan, Y.; Carr, R.; Barrow, C.; Ewart, H. Immunostimulatory principles from Chlorella pyrenoidosa—Part 1: Isolation and biological assessment in vitro. Phytomedicine 2007, 14, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Reyna-Martinez, R.; Gomez-Flores, R.; López-Chuken, U.; Quintanilla-Licea, R.; Caballero-Hernandez, D.; Rodríguez-Padilla, C.; Beltrán-Rocha, J.C.; Tamez-Guerra, P. Antitumor activity of Chlorella sorokiniana and Scenedesmus sp. microalgae native of Nuevo León State, México. PeerJ 2018, 6, e4358. [Google Scholar] [CrossRef] [PubMed]
- Sheih, I.-C.; Fang, T.J.; Wu, T.-K. Isolation and characterisation of a novel angiotensin I-converting enzyme (ACE) inhibitory peptide from the algae protein waste. Food Chem. 2009, 115, 279–284. [Google Scholar] [CrossRef]
- Suetsuna, K.; Chen, J.-R. Identification of antihypertensive peptides from peptic digest of two microalgae, Chlorella vulgaris and Spirulina platensis. Mar. Biotechnol. 2001, 3, 305–309. [Google Scholar] [CrossRef] [PubMed]
- Samarakoon, K.W.; Kwon, O.-N.; Ko, J.-Y.; Lee, J.-H.; Kang, M.-C.; Kim, D.; Lee, J.B.; Lee, J.-S.; Jeon, Y.-J. Purification and identification of novel angiotensin-I converting enzyme (ACE) inhibitory peptides from cultured marine microalgae (Nannochloropsis oculata) protein hydrolysate. J. Appl. Phycol. 2013, 25, 1595–1606. [Google Scholar] [CrossRef]
- Kang, K.-H.; Qian, Z.-J.; Ryu, B.; Kim, D.; Kim, S.-K. Protective effects of protein hydrolysate from marine microalgae Navicula incerta on ethanol-induced toxicity in HepG2/CYP2E1 cells. Food Chem. 2012, 132, 677–685. [Google Scholar] [CrossRef]
- Ko, S.-C.; Kim, D.; Jeon, Y.-J. Protective effect of a novel antioxidative peptide purified from a marine Chlorella ellipsoidea protein against free radical-induced oxidative stress. Food Chem. Toxicol. 2012, 50, 2294–2302. [Google Scholar] [CrossRef] [PubMed]
- Vo, T.-S.; Ryu, B.; Kim, S.-K. Purification of novel anti-inflammatory peptides from enzymatic hydrolysate of the edible microalgal Spirulina maxima. J. Funct. Foods 2013, 5, 1336–1346. [Google Scholar] [CrossRef]
- Sun, Y.; Chang, R.; Li, Q.; Li, B. Isolation and characterization of an antibacterial peptide from protein hydrolysates of Spirulina platensis. Eur. Food Res. Technol. 2016, 242, 685–692. [Google Scholar] [CrossRef]
- Lin, Y.-H.; Chen, G.-W.; Yeh, C.; Song, H.; Tsai, J.-S. Purification and Identification of Angiotensin I-Converting Enzyme Inhibitory Peptides and the Antihypertensive Effect of Chlorella sorokiniana Protein Hydrolysates. Nutrients 2018, 10, 1397. [Google Scholar] [CrossRef] [PubMed]
- Morgese, M.; Mhillaj, E.; Francavilla, M.; Bove, M.; Morgano, L.; Tucci, P.; Trabace, L.; Schiavone, S. Chlorella sorokiniana extract improves short-term memory in rats. Molecules 2016, 21, 1311. [Google Scholar] [CrossRef] [PubMed]
- Talero, E.; García-Mauriño, S.; Ávila-Román, J.; Rodríguez-Luna, A.; Alcaide, A.; Motilva, V. Bioactive compounds isolated from microalgae in chronic inflammation and cancer. Mar. Drugs 2015, 13, 6152–6209. [Google Scholar] [CrossRef] [PubMed]
- Gevaert, K.; Vandekerckhove, J. Protein identification methods in proteomics. ELECTROPHORESIS Int. J. 2000, 21, 1145–1154. [Google Scholar] [CrossRef]
- Chang, Y.-W.; Alli, I.; Konishi, Y.; Ziomek, E. Characterization of protein fractions from chickpea (Cicer arietinum L.) and oat (Avena sativa L.) seeds using proteomic techniques. Food Res. Int. 2011, 44, 3094–3104. [Google Scholar] [CrossRef]
- Mazzeo, M.F.; Giulio, B.D.; Guerriero, G.; Ciarcia, G.; Malorni, A.; Russo, G.L.; Siciliano, R.A. Fish authentication by MALDI-TOF mass spectrometry. J. Agric. Food Chem. 2008, 56, 11071–11076. [Google Scholar] [CrossRef]
- Böhme, K.; Fernández-No, I.C.; Barros-Velázquez, J.; Gallardo, J.M.; Cañas, B.; Calo-Mata, P. Rapid species identification of seafood spoilage and pathogenic Gram-positive bacteria by MALDI-TOF mass fingerprinting. Electrophoresis 2011, 32, 2951–2965. [Google Scholar] [CrossRef] [PubMed]
- Huang, B.-B.; Lin, H.-C.; Chang, Y.-W. Analysis of proteins and potential bioactive peptides from tilapia (Oreochromis spp.) processing co-products using proteomic techniques coupled with BIOPEP database. J. Funct. Foods 2015, 19, 629–640. [Google Scholar] [CrossRef]
- Jao, C.-L.; Hung, C.-C.; Tung, Y.-S.; Lin, P.-Y.; Chen, M.-C.; Hsu, K.-C. The development of bioactive peptides from dietary proteins as a dipeptidyl peptidase IV inhibitor for the management of type 2 diabetes. Biomedicine 2015, 5, 14. [Google Scholar] [CrossRef]
- Minkiewicz, P.; Dziuba, J.; Iwaniak, A.; Dziuba, M.; Darewicz, M. BIOPEP database and other programs for processing bioactive peptide sequences. J. AOAC Int. 2008, 91, 965–980. [Google Scholar]
- Kumar, C.; Mann, M. Bioinformatics analysis of mass spectrometry-based proteomics data sets. FEBS Lett. 2009, 583, 1703–1712. [Google Scholar] [CrossRef] [Green Version]
- Cheung, I.W.; Nakayama, S.; Hsu, M.N.; Samaranayaka, A.G.; Li-Chan, E.C. Angiotensin-I converting enzyme inhibitory activity of hydrolysates from oat (Avena sativa) proteins by in silico and in vitro analyses. J. Agric. Food Chem. 2009, 57, 9234–9242. [Google Scholar] [CrossRef]
- Gangopadhyay, N.; Wynne, K.; O’Connor, P.; Gallagher, E.; Brunton, N.P.; Rai, D.K.; Hayes, M. In silico and in vitro analyses of the angiotensin-I converting enzyme inhibitory activity of hydrolysates generated from crude barley (Hordeum vulgare) protein concentrates. Food Chem. 2016, 203, 367–374. [Google Scholar] [CrossRef]
- Lafarga, T.; O’Connor, P.; Hayes, M. Identification of novel dipeptidyl peptidase-IV and angiotensin-I-converting enzyme inhibitory peptides from meat proteins using in silico analysis. Peptides 2014, 59, 53–62. [Google Scholar] [CrossRef]
- Waghmare, A.G.; Salve, M.K.; LeBlanc, J.G.; Arya, S.S. Concentration and characterization of microalgae proteins from Chlorella pyrenoidosa. Bioresour. Bioprocess. 2016, 3, 16. [Google Scholar] [CrossRef]
- Rosenberg, J.N.; Kobayashi, N.; Barnes, A.; Noel, E.A.; Betenbaugh, M.J.; Oyler, G.A. Comparative analyses of three Chlorella species in response to light and sugar reveal distinctive lipid accumulation patterns in the microalga C. sorokiniana. PLoS ONE 2014, 9, e92460. [Google Scholar] [CrossRef]
- Watson, H.; Walker, N.; Shaw, P.; Bryant, T.; Wendell, P.; Fothergill, L.; Perkins, R.; Conroy, S.; Dobson, M.; Tuite, M. Sequence and structure of yeast phosphoglycerate kinase. EMBO J. 1982, 1, 1635–1640. [Google Scholar] [CrossRef]
- Banks, R.; Blake, C.; Evans, P.; Haser, R.; Rice, D.; Hardy, G.; Merrett, M.; Phillips, A. Sequence, structure and activity of phosphoglycerate kinase: A possible hinge-bending enzyme. Nature 1979, 279, 773–777. [Google Scholar] [CrossRef]
- Shevchenko, A.; Tomas, H.; Havli, J.; Olsen, J.V.; Mann, M. In-gel digestion for mass spectrometric characterization of proteins and proteomes. Nat. Protoc. 2006, 1, 2856–2860. [Google Scholar] [CrossRef]
- Wysocki, V.H.; Resing, K.A.; Zhang, Q.; Cheng, G. Mass spectrometry of peptides and proteins. Methods 2005, 35, 211–222. [Google Scholar] [CrossRef]
- Power, O.; Nongonierma, A.B.; Jakeman, P.; FitzGerald, R.J. Food protein hydrolysates as a source of dipeptidyl peptidase IV inhibitory peptides for the management of type 2 diabetes. Proc. Nutr. Soc. 2014, 73, 34–46. [Google Scholar] [CrossRef]
- Boots, J.-W.P. Protein Hydrolysate Enriched in Peptides Inhibiting DPP IV and Their Use. U.S. Patent No. 8,273,710, 25 September 2012. [Google Scholar]
- Lambeir, A.-M.; Durinx, C.; Scharpé, S.; De Meester, I. Dipeptidyl-peptidase IV from bench to bedside: An update on structural properties, functions, and clinical aspects of the enzyme DPP IV. Crit. Rev. Clin. Lab. Sci. 2003, 40, 209–294. [Google Scholar] [CrossRef]
- Liu, R.; Zhou, L.; Zhang, Y.; Sheng, N.-J.; Wang, Z.-K.; Wu, T.-Z.; Wang, X.-Z.; Wu, H. Rapid Identification of Dipeptidyl Peptidase-IV (DPP IV) Inhibitory Peptides from Ruditapes philippinarum Hydrolysate. Molecules 2017, 22, 1714. [Google Scholar] [CrossRef]
- Lacroix, I.M.; Li-Chan, E.C. Evaluation of the potential of dietary proteins as precursors of dipeptidyl peptidase (DPP)-IV inhibitors by an in silico approach. J. Funct. Foods 2012, 4, 403–422. [Google Scholar] [CrossRef]
- Nongonierma, A.B.; FitzGerald, R.J. An in silico model to predict the potential of dietary proteins as sources of dipeptidyl peptidase IV (DPP IV) inhibitory peptides. Food Chem. 2014, 165, 489–498. [Google Scholar] [CrossRef]
- Nongonierma, A.B.; Mooney, C.; Shields, D.C.; FitzGerald, R.J. In silico approaches to predict the potential of milk protein-derived peptides as dipeptidyl peptidase IV (DPP IV) inhibitors. Peptides 2014, 57, 43–51. [Google Scholar] [CrossRef]
- Kim, Y.K.; Chung, B.H. A novel angiotensin-I-converting enzyme inhibitory peptide from human αs1-casein. Biotechnol. Lett. 1999, 21, 575–578. [Google Scholar] [CrossRef]
- Kohmura, M.; Nio, N.; Kubo, K.; Minoshima, Y.; Munekata, E.; Ariyoshi, Y. Inhibition of angiotensin-converting enzyme by synthetic peptides of human β-casein. Agric. Biol. Chem. 1989, 53, 2107–2114. [Google Scholar] [CrossRef]
- Li, C.H.; Matsui, T.; Matsumoto, K.; Yamasaki, R.; Kawasaki, T. Latent production of angiotensin I-converting enzyme inhibitors from buckwheat protein. J. Pept. Sci. 2002, 8, 267–274. [Google Scholar] [CrossRef]
- Maruyama, S.; Mitachi, H.; Awaya, J.; Kurono, M.; Tomizuka, N.; Suzuki, H. Angiotensin I-converting enzyme inhibitory activity of the C-terminal hexapeptide of αs1-casein. Agric. Biol. Chem. 1987, 51, 2557–2561. [Google Scholar] [CrossRef]
- Miyoshi, S.; Ishikawa, H.; Kaneko, T.; Fukui, F.; Tanaka, H.; Maruyama, S. Structures and activity of angiotensin-converting enzyme inhibitors in an α-zein hydrolysate. Agric. Biol. Chem. 1991, 55, 1313–1318. [Google Scholar] [CrossRef]
- Li, G.-H.; Le, G.-W.; Shi, Y.-H.; Shrestha, S. Angiotensin I–converting enzyme inhibitory peptides derived from food proteins and their physiological and pharmacological effects. Nutr. Res. 2004, 24, 469–486. [Google Scholar] [CrossRef]
- Abdelhedi, O.; Nasri, R.; Jridi, M.; Mora, L.; Oseguera-Toledo, M.E.; Aristoy, M.-C.; Amara, I.B.; Toldrá, F.; Nasri, M. In silico analysis and antihypertensive effect of ACE-inhibitory peptides from smooth-hound viscera protein hydrolysate: Enzyme-peptide interaction study using molecular docking simulation. Process Biochem. 2017, 58, 145–159. [Google Scholar] [CrossRef]
- Bleakley, S.; Hayes, M.; O’Shea, N.; Gallagher, E.; Lafarga, T. Predicted Release and Analysis of Novel ACE-I, Renin, and DPP IV Inhibitory Peptides from Common Oat (Avena sativa) Protein Hydrolysates Using in Silico Analysis. Foods 2017, 6, 108. [Google Scholar] [CrossRef]
- Muhammad, S.A.; Fatima, N. In silico analysis and molecular docking studies of potential angiotensin-converting enzyme inhibitor using quercetin glycosides. Pharmacogn. Mag. 2015, 11, S123–S126. [Google Scholar] [CrossRef]
- Paiva, L.; Lima, E.; Neto, A.I.; Baptista, J. Isolation and characterization of angiotensin I-converting enzyme (ACE) inhibitory peptides from Ulva rigida C. Agardh protein hydrolysate. J. Funct. Foods 2016, 26, 65–76. [Google Scholar] [CrossRef]
- Kang, K.-H.; Qian, Z.-J.; Ryu, B.; Kim, S.-K. Characterization of growth and protein contents from microalgae Navicula incerta with the investigation of antioxidant activity of enzymatic hydrolysates. Food Sci. Biotechnol. 2011, 20, 183–191. [Google Scholar] [CrossRef]
- Sheih, I.-C.; Wu, T.-K.; Fang, T.J. Antioxidant properties of a new antioxidative peptide from algae protein waste hydrolysate in different oxidation systems. Bioresour. Technol. 2009, 100, 3419–3425. [Google Scholar] [CrossRef]
- Parimi, N.S.; Singh, M.; Kastner, J.R.; Das, K.C.; Forsberg, L.S.; Azadi, P. Optimization of protein extraction from Spirulina platensis to generate a potential co-product and a biofuel feedstock with reduced nitrogen content. Front. Energy Res. 2015, 3, 30. [Google Scholar] [CrossRef]
- Markwell, M.A.K.; Haas, S.M.; Bieber, L.; Tolbert, N. A modification of the Lowry procedure to simplify protein determination in membrane and lipoprotein samples. Anal. Biochem. 1978, 87, 206–210. [Google Scholar] [CrossRef]
- Schägger, H.; Von Jagow, G. Tricine-sodium dodecyl sulfate-polyacrylamide gel electrophoresis for the separation of proteins in the range from 1 to 100 kDa. Anal. Biochem. 1987, 166, 368–379. [Google Scholar] [CrossRef]
- Rosenfeld, J.; Capdevielle, J.; Guillemot, J.C.; Ferrara, P. In-gel digestion of proteins for internal sequence analysis after one-or two-dimensional gel electrophoresis. Anal. Biochem. 1992, 203, 173–179. [Google Scholar] [CrossRef]
- Perkins, D.N.; Pappin, D.J.; Creasy, D.M.; Cottrell, J.S. Probability-based protein identification by searching sequence databases using mass spectrometry data. ELECTROPHORESIS Int. J. 1999, 20, 3551–3567. [Google Scholar] [CrossRef]
- Pruitt, K.D.; Tatusova, T.; Maglott, D.R. NCBI reference sequences (RefSeq): A curated non-redundant sequence database of genomes, transcripts and proteins. Nucleic Acids Res. 2006, 35, D61–D65. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein Name | Accession Number (NCBI) | Score | Sequence Coverage (%) | Length (AA) | Molecular Weights from NCBI Database (kDa) | Molecular Weights (kDa) Estimated from SDS–PAGE |
|---|---|---|---|---|---|---|
| Chloroplast Rubisco Activase | AEL29575.1 | 323 | 31% | 567 | 45.68 | 45.72 (D) |
| 109.02 (A) | ||||||
| 72.08 (B) | ||||||
| 38.33 (E) | ||||||
| 29.04 (F) | ||||||
| 24.96 (G) | ||||||
| 21.13 (H) | ||||||
| 16.59 (I) | ||||||
| 7.82 (J) | ||||||
| 50S Ribosomal Protein L7/L12 (Chloroplast) | YP_009020879.1 | 72 | 18% | 130 | 13.58 | 7.82 (J) |
| Phosphoglycerate Kinase | AKP17751.1 | 250 | 22% | 465 | 49.13 | 45.72 (D) |
| 109.02 (A) | ||||||
| 72.08 (B) | ||||||
| 54.15 (C) | ||||||
| 38.33 (E) | ||||||
| 29.04 (F) | ||||||
| 24.96 (G) | ||||||
| 21.13 (H) | ||||||
| 16.59 (I) | ||||||
| 7.82 (J) | ||||||
| Fe-superoxide Dismutase | AHD25899.1 | 82 | 8% | 236 | 26.41 | 24.96 (G) |
| Heat Shock Protein 70 | AKP17750.1 | 128 | 29% | 652 | 71.55 | 72.08 (B) |
| 109.02 (A) | ||||||
| 54.15 (C) | ||||||
| 38.33 (E) | ||||||
| ATP Synthase Subunit Beta (Chloroplast) | YP_009020893.1 | 906 | 49% | 481 | 51.833 | 54.15 (C) |
| elongation factor 2, partial | BAE48222.1 | 120 | 4% | 816 | 90.54 | 45.72 (D) |
| 7.82 (J) | ||||||
| V-type H+ ATPase Subunit A, partial | BAE48224.1 | 61 | 4% | 596 | 65.47 | 7.82 (J) |
| Protein Name | Number of Bioactive Peptides | ||||||
|---|---|---|---|---|---|---|---|
| AC | AO | AA | AT | S | R | DPP | |
| Chloroplast Rubisco Activase | 187 (0.329) | 18 (0.032) | 3 (0.005) | 2 (0.004) | 14 (0.25) | 3 (0.005) | 250 (0.441) |
| 50s Ribosomal Protein L7/L12 (Chloroplast) | 59 (0.454) | 8 (0.062) | 1 (0.008) | 1 (0.008) | 8 (0.062) | 1 (0.008) | 83 (0.638) |
| Phosphoglycerate Kinase | 224 (0.482) | 23 (0.049) | 5 (0.011) | 4 (0.009) | 33 (0.071) | 6 (0.013) | 297 (0.639) |
| Fe-Superoxide Dismutase | 107 (0.453) | 18 (0.076) | - | - | 5 (0.021) | - | 156 (0.661) |
| Heat Shock Protein 70 | 276 (0.423) | 30 (0.046) | 4 (0.006) | 4 (0.006) | 41 (0.064) | 4 (0.006) | 446 (0.684) |
| ATP Synthase Subunit Beta (Chloroplast) | 204 (0.424) | 20 (0.042) | 5 (0.010) | 5 (0.010) | 25 (0.052) | 6 (0.12) | 332 (0.690) |
| Protease | Chloroplast Rubisco Activase | Phosphoglycerate Kinase | Heat Shock Protein 70 | ATP Synthase Subunit Beta | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| AC | AO | S | DP | AC | AO | S | DP | AC | AO | S | DP | AC | AO | S | DP | |
| Pepsin (EC 3.4.23.1) | 31 | 2 | 2 | 48 | 29 | - | 5 | 51 | 42 | 11 | 1 | 78 | 44 | 6 | 9 | 66 |
| Thermolysin (EC 3.4.24.27) | 12 | - | - | 14 | 20 | 4 | - | 23 | 21 | - | - | 31 | 20 | 1 | - | 25 |
| Bromelain (EC 3.4.22.32) | 26 | 3 | 1 | 36 | 25 | 8 | 3 | 41 | 36 | 3 | 2 | 50 | 20 | 2 | 4 | 37 |
| Chymotrypsin A (EC 3.4.21.1) | 5 | 1 | 1 | 10 | 9 | 1 | 2 | 11 | 8 | 2 | 2 | 15 | 7 | 4 | 2 | 10 |
| Chymotrypsin C (EC 3.4.21.2) | 17 | 3 | 1 | 16 | 14 | - | 2 | 19 | 20 | 6 | 2 | 35 | 17 | 3 | 4 | 25 |
| Pancreatic elastase (EC 3.4.21.36) | 16 | 2 | - | 31 | 27 | 5 | - | 49 | 21 | 2 | - | 34 | 19 | 2 | - | 41 |
| Papain (EC 3.4.22.2) | 31 | 3 | 1 | 35 | 27 | 6 | 2 | 31 | 40 | 4 | 4 | 50 | 27 | 1 | 4 | 43 |
| Proteinase K (EC 3.4.21.67) | 12 | 1 | - | 20 | 17 | 1 | 1 | 19 | 23 | 5 | - | 41 | 15 | 1 | - | 28 |
| Trypsin (EC 3.4.21.4) | 5 | - | - | 3 | 3 | 1 | - | 3 | 5 | - | - | 2 | 3 | - | - | 3 |
| Ficin (EC 3.4.22.3) | 27 | 7 | 2 | 37 | 22 | 8 | 3 | 33 | 26 | 4 | 3 | 35 | 22 | 7 | 1 | 28 |
| Cathepsin (EC 3.4.21.20) | 4 | 1 | 1 | 6 | 7 | 1 | 1 | 6 | 6 | 2 | 1 | 7 | 5 | 3 | 1 | 6 |
| Subtisilin (EC 3.4.21.62) | 10 | 4 | 2 | 14 | 10 | 3 | 4 | 18 | 9 | 5 | 7 | 24 | 12 | 2 | 6 | 23 |
| Pepsin + Trypsin | 28 | 4 | 2 | 55 | 33 | 1 | 5 | 56 | 36 | 2 | 10 | 66 | 40 | 6 | 9 | 62 |
| Pepsin + Trysin + Chymotrypsin A | 31 | 4 | 1 | 40 | 33 | 1 | 5 | 54 | 34 | 1 | 10 | 63 | 38 | 5 | 9 | 61 |
| Pepsin + Trysin + Chymotrypsin C | 28 | 4 | 1 | 33 | 35 | 1 | 4 | 54 | 41 | 2 | 10 | 70 | 38 | 5 | 9 | 51 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tejano, L.A.; Peralta, J.P.; Yap, E.E.S.; Panjaitan, F.C.A.; Chang, Y.-W. Prediction of Bioactive Peptides from Chlorella sorokiniana Proteins Using Proteomic Techniques in Combination with Bioinformatics Analyses. Int. J. Mol. Sci. 2019, 20, 1786. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20071786
Tejano LA, Peralta JP, Yap EES, Panjaitan FCA, Chang Y-W. Prediction of Bioactive Peptides from Chlorella sorokiniana Proteins Using Proteomic Techniques in Combination with Bioinformatics Analyses. International Journal of Molecular Sciences. 2019; 20(7):1786. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20071786
Chicago/Turabian StyleTejano, Lhumen A., Jose P. Peralta, Encarnacion Emilia S. Yap, Fenny Crista A. Panjaitan, and Yu-Wei Chang. 2019. "Prediction of Bioactive Peptides from Chlorella sorokiniana Proteins Using Proteomic Techniques in Combination with Bioinformatics Analyses" International Journal of Molecular Sciences 20, no. 7: 1786. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20071786