Effect of Ultraviolet-C Radiation and Melatonin Stress on Biosynthesis of Antioxidant and Antidiabetic Metabolites Produced in In Vitro Callus Cultures of Lepidium sativum L.

 , , ,
, , ,  and
and 
Abstract
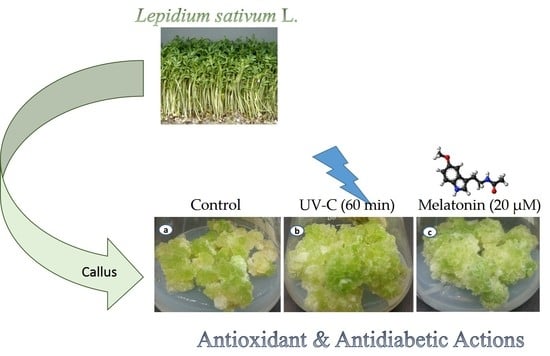
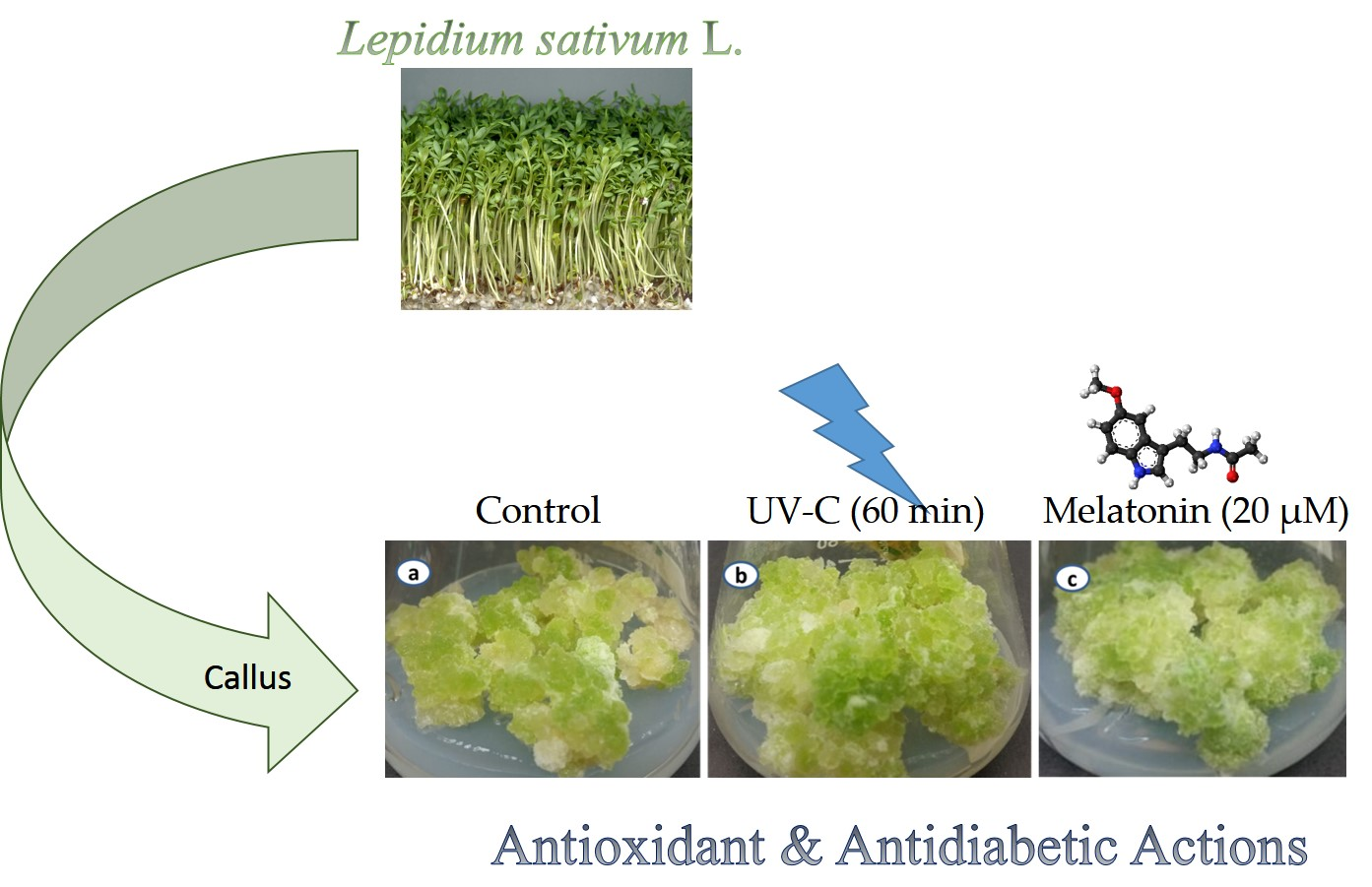
:
1. Introduction
2. Results and Discussion
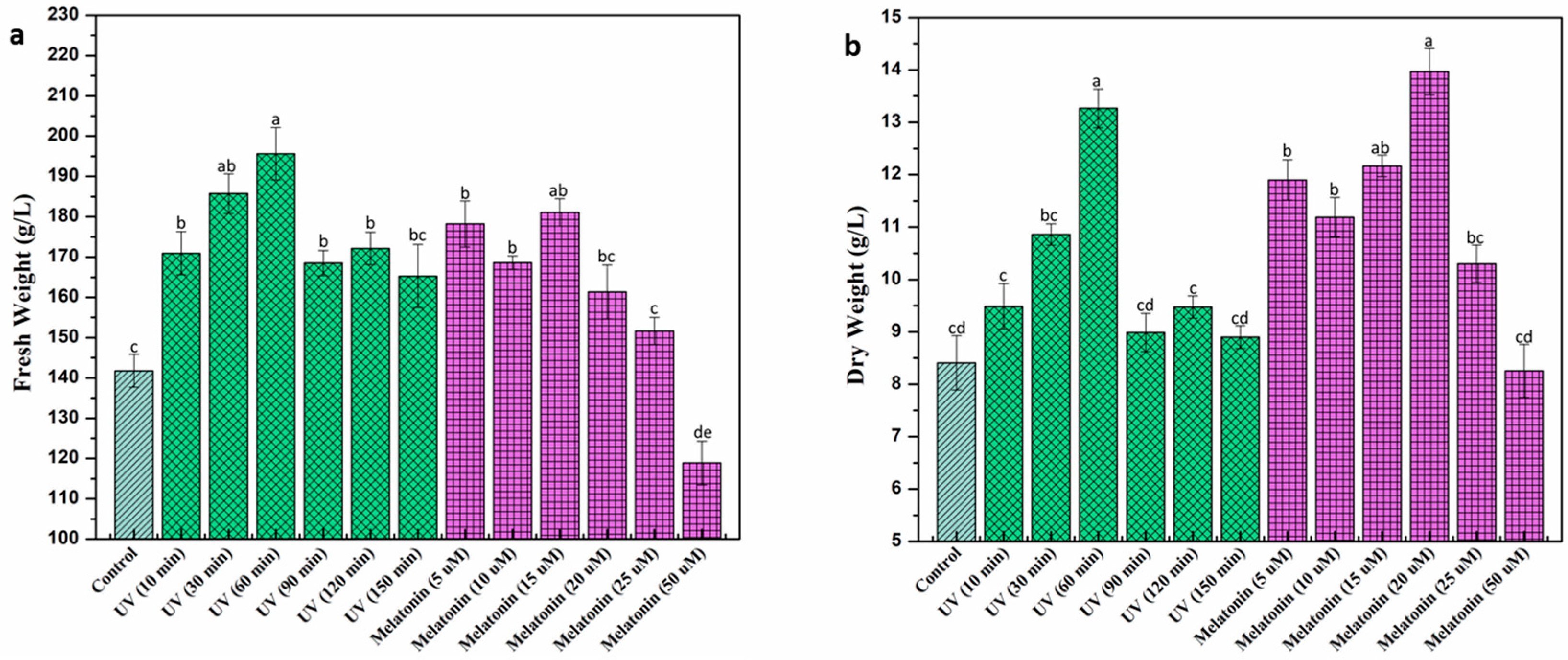
2.1. Biomass Accumulation under UV-C and Melatonin Treatment
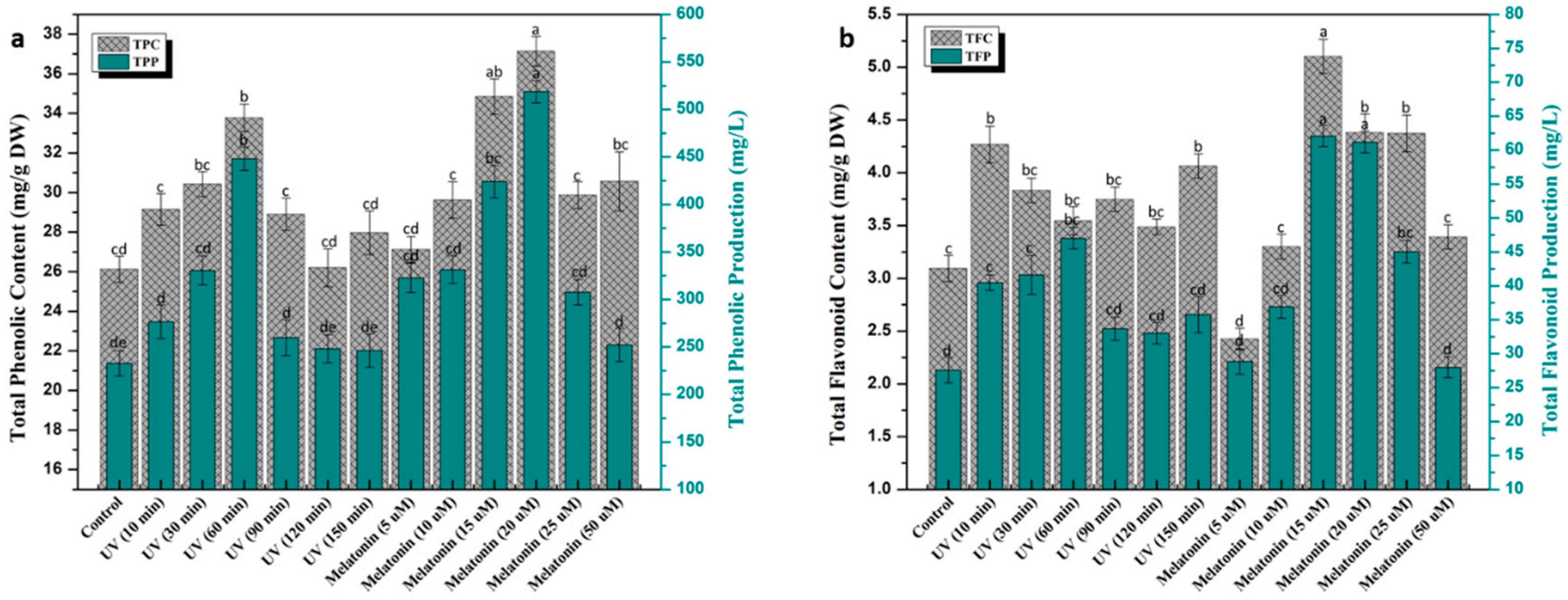
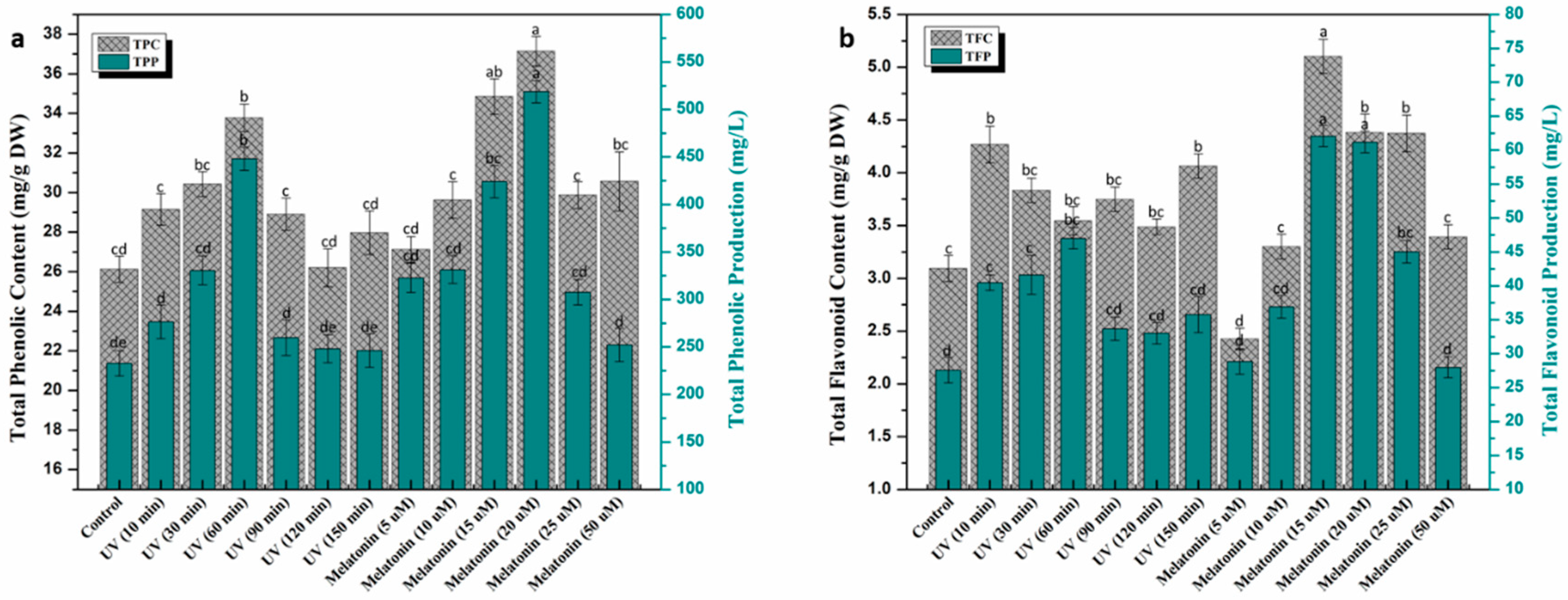
2.2. Trends in Phenolic and Flavonoid Production under UV-C and Melatonin Treatments
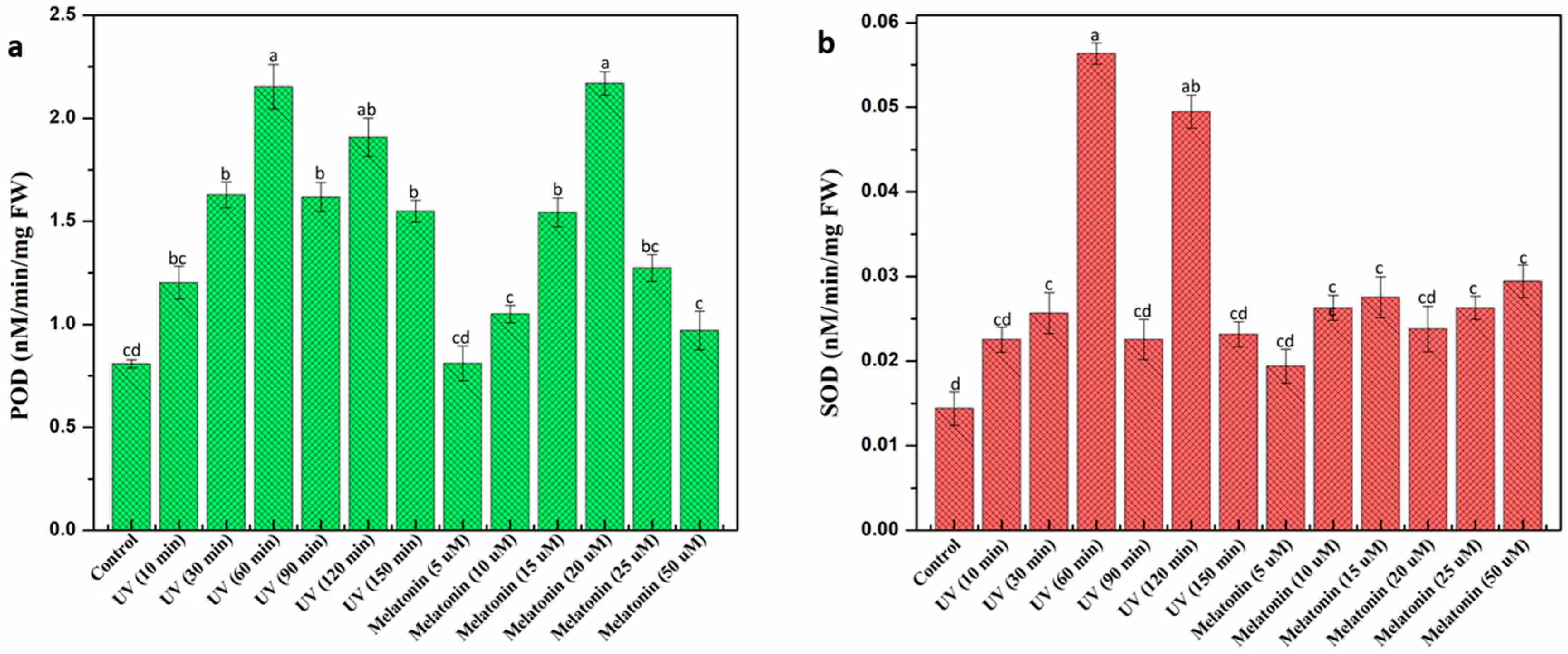
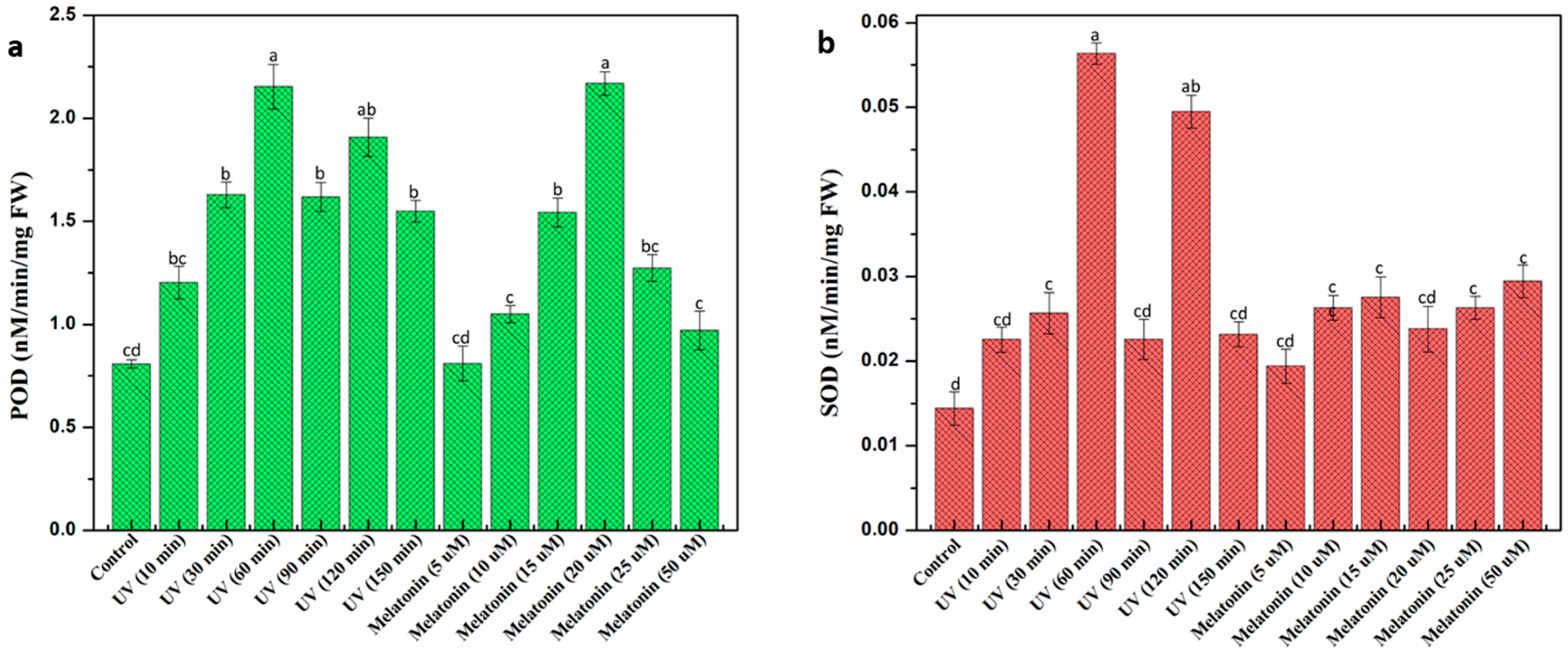
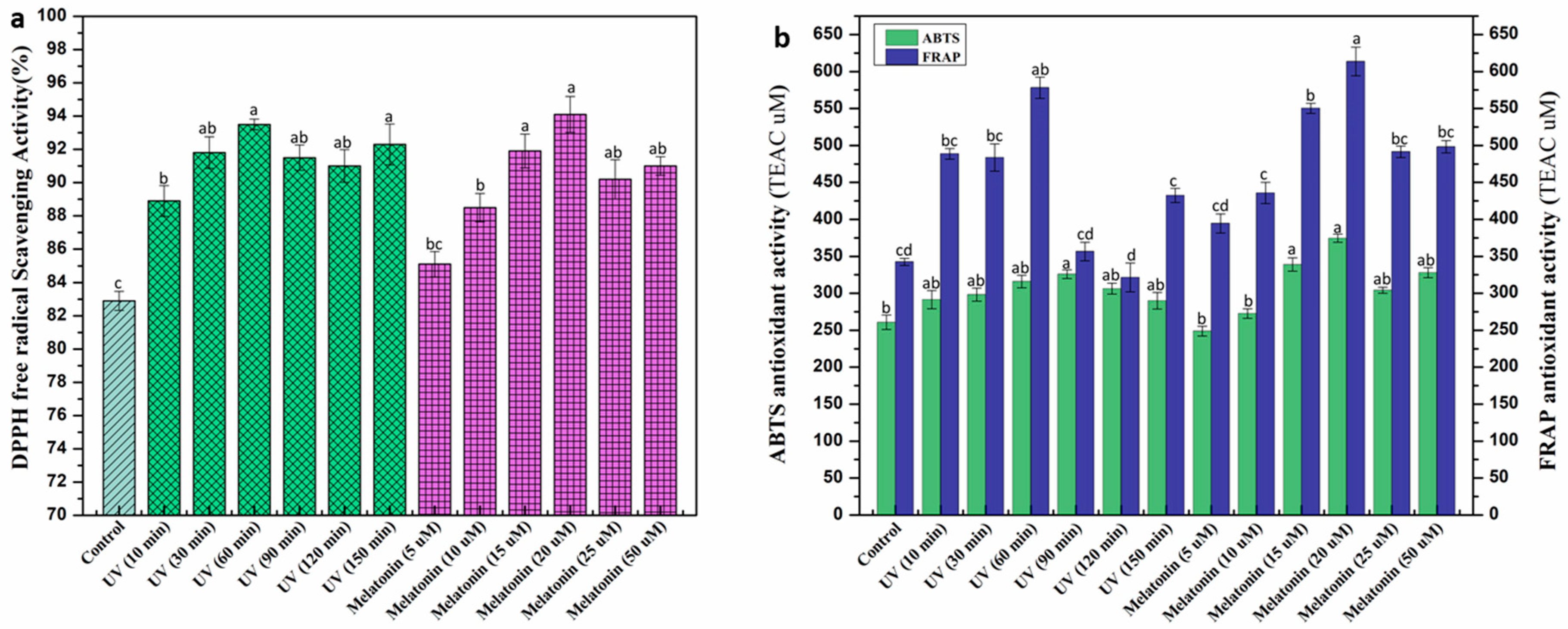
2.3. Effect of UV-C and Melatonin on In Vitro Antioxidant Activities of L. sativum Callus
2.4. Effect of Elicitors on Antidiabetic Potential of L. sativum Callus Culture
2.5. Quantification of Polyphenols Profile in L. sativum Callus Culture via HPLC
3. Materials and Methods
3.1. Chemicals
3.2. Seed Germination and Callus Induction of L. sativum
3.3. Elicitors Treatment on Callus Culture
3.3.1. UV-C Treatment
3.3.2. Melatonin Treatment
3.4. Sample Extraction
3.5. Estimation of Secondary Metabolites Accumulation
3.5.1. Qualitative Phenolic and Flavonoids Estimation
3.5.2. Quantitative Estimation of Polyphenols via HPLC
3.6. In Vitro Antioxidant Activities
3.6.1. DPPH Free Radical Scavenging Activity
3.6.2. Ferric Reducing Antioxidant Power (FRAP) Assay
3.6.3. Antioxidant ABTS Assay
3.6.4. Anti-AGEs Formation Activity
3.7. Estimation of Antidiabetic Potential
3.7.1. α-Glucosidase Inhibition
3.7.2. α-Amylase Inhibition
3.8. In Vitro Antioxidant Enzymatic Activities
3.8.1. Peroxidase Activity (POD)
3.8.2. Superoxide Dismutase Activity (SOD)
3.9. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Nadkarni, K. The Indian Materia Medica, with Ayurvedic, Unani and Home Remedies; Revised and Enlarged by AK Nadkarni; Popular Prakashan: Bombay, India, 1954; pp. 947–948. [Google Scholar]
- Nadkarni, K.; Nadkarni, A. Indian Materia Medica; Popular Prakashan Pvt. Ltd.: Bombay, India, 1976; Volume 1, 799p. [Google Scholar]
- Doke, S.; Guha, M. Garden cress (Lepidium sativum L.) seed—An important medicinal source: A Review. J. Nat. Prod. Plant Resour. 2014, 4, 69–80. [Google Scholar]
- Rehman, N.-U.; Khan, A.-U.; Alkharfy, K.M.; Gilani, A.-H. Pharmacological basis for the medicinal use of Lepidium sativum in airways disorders. Evid. Based Complement. Altern. Med. 2012. [Google Scholar] [CrossRef]
- Ahsan, S.; Tariq, M.; Ageel, M.; Alyahya, M.; Shah, A. Studies on some herbal drugs used in fracture healing. Int. J. Crude Drug Res. 1989, 27, 235–239. [Google Scholar] [CrossRef]
- Rahimi, R.; Shams-Ardekani, M.R.; Abdollahi, M. A review of the efficacy of traditional Iranian medicine for inflammatory bowel disease. World J. Gastroenterol. WJG 2010, 16, 4504. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Tyagi, A.K.; Kaur, H. Cancer modulation by glucosinolates: A review. Curr. Sci. 2000, 79, 1665–1671. [Google Scholar]
- Divisi, D.; Di Tommaso, S.; Salvemini, S. Diet and cancer. Acta Bio Med. Atenei Parm. 2006, 77, 118–123. [Google Scholar]
- Diwakar, B.; Dutta, P.; Lokesh, B.; Naidu, K. Bio-availability and metabolism of n-3 fatty acid rich garden cress (Lepidium sativum) seed oil in albino rats. Prostaglandins Leukot. Essent. Fat. Acids 2008, 78, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Gokavi, S.S.; Malleshi, N.G.; Guo, M. Chemical composition of garden cress (Lepidium sativum) seeds and its fractions and use of bran as a functional ingredient. Plant Foods Hum. Nutr. 2004, 59, 105–111. [Google Scholar] [CrossRef]
- Güvenç, A.; Okada, Y.; Akkol, E.K.; Duman, H.; Okuyama, T.; Çalış, İ. Investigations of anti-inflammatory, antinociceptive, antioxidant and aldose reductase inhibitory activities of phenolic compounds from Sideritis brevibracteata. Food Chem. 2010, 118, 686–692. [Google Scholar] [CrossRef]
- Hudaib, M.; Mohammad, M.; Bustanji, Y.; Tayyem, R.; Yousef, M.; Abuirjeie, M.; Aburjai, T. Ethnopharmacological survey of medicinal plants in Jordan, Mujib Nature Reserve and surrounding area. J. Ethnopharmacol. 2008, 120, 63–71. [Google Scholar] [CrossRef]
- Jänicke, R.U.; Sprengart, M.L.; Wati, M.R.; Porter, A.G. Caspase-3 is required for DNA fragmentation and morphological changes associated with apoptosis. J. Biol. Chem. 1998, 273, 9357–9360. [Google Scholar] [CrossRef] [PubMed]
- Conforti, F.; Ioele, G.; Statti, G.; Marrelli, M.; Ragno, G.; Menichini, F. Antiproliferative activity against human tumor cell lines and toxicity test on Mediterranean dietary plants. Food Chem. Toxicol. 2008, 46, 3325–3332. [Google Scholar] [CrossRef] [PubMed]
- Hardman, W.E.; Avula, C.R.; Fernandes, G.; Cameron, I.L. Three percent dietary fish oil concentrate increased efficacy of doxorubicin against MDA-MB 231 breast cancer xenografts. Clin. Cancer Res. 2001, 7, 2041–2049. [Google Scholar] [PubMed]
- Kassie, F.; Pool-Zobel, B.; Parzefall, W.; Knasmüller, S. Genotoxic effects of benzyl isothiocyanate, a natural chemopreventive agent. Mutagenesis 1999, 14, 595–604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kassie, F.; Laky, B.; Gminski, R.; Mersch-Sundermann, V.; Scharf, G.; Lhoste, E.; Kansmüller, S. Effects of garden and water cress juices and their constituents, benzyl and phenethyl isothiocyanates, towards benzo (a) pyrene-induced DNA damage: A model study with the single cell gel electrophoresis/Hep G2 assay. Chem. Biol. Interact. 2003, 142, 285–296. [Google Scholar] [CrossRef]
- Khan, N.; Adhami, V.M.; Mukhtar, H. Apoptosis by dietary agents for prevention and treatment of cancer. Biochem. Pharmacol. 2008, 76, 1333–1339. [Google Scholar] [CrossRef]
- Merlin, N.; Parthasarathy, V.; Santhoshkumar, T. Induction of apoptosis in human breast cancer cell line MCF-7 by phytochemicals from Gmelina asiatica. Afr. J. Biotechnol. 2010, 9, 4451–4456. [Google Scholar]
- Bhushan, M.S.; Rao, C.; Ojha, S.; Vijayakumar, M.; Verma, A. An analytical review of plants for anti diabetic activity with their phytoconstituent & mechanism of action. Int. J. Pharm. Sci. Res. 2010, 1, 29–46. [Google Scholar]
- Attia, E.S.; Amer, A.H.; Hasanein, M.A. The hypoglycemic and antioxidant activities of garden cress (Lepidium sativum L.) seed on alloxan-induced diabetic male rats. Nat. Prod. Res. 2018, 13, 1–5. [Google Scholar] [CrossRef]
- Khan, M.A.; Abbasi, B.H.; Ali, H.; Ali, M.; Adil, M.; Hussain, I. Temporal variations in metabolite profiles at different growth phases during somatic embryogenesis of Silybum marianum L. Plant Cell Tissue Organ Cult. 2015, 120, 127–139. [Google Scholar] [CrossRef]
- Matkowski, A. Plant in vitro culture for the production of antioxidants—A review. Biotechnol. Adv. 2008, 26, 548–560. [Google Scholar] [CrossRef]
- Davies, K.M.; Deroles, S.C. Prospects for the use of plant cell cultures in food biotechnology. Curr. Opin. Biotechnol. 2014, 26, 133–140. [Google Scholar] [CrossRef]
- Yang, L.; Stöckigt, J. Trends for diverse production strategies of plant medicinal alkaloids. Nat. Prod. Rep. 2010, 27, 1469–1479. [Google Scholar] [CrossRef]
- Yue, W.; Ming, Q.-l.; Lin, B.; Rahman, K.; Zheng, C.-J.; Han, T.; Qin, L.-P. Medicinal plant cell suspension cultures: Pharmaceutical applications and high-yielding strategies for the desired secondary metabolites. Crit. Rev. Biotechnol. 2016, 36, 215–232. [Google Scholar] [CrossRef]
- Zhao, J.; Davis, L.C.; Verpoorte, R. Elicitor signal transduction leading to production of plant secondary metabolites. Biotechnol. Adv. 2005, 23, 283–333. [Google Scholar] [CrossRef]
- Xuan, T.D.; Khanh, T.D.; Khang, D.T.; Quan, N.T.; Elzaawely, A.A. Changes in chemical composition, total phenolics and antioxidant activity of Alpinia (Alpinia zerumbet) leaves exposed to UV. Int. Lett. Nat. Sci. 2016, 55, 25–34. [Google Scholar] [CrossRef]
- Yin, X.; Singer, S.D.; Qiao, H.; Liu, Y.; Jiao, C.; Wang, H.; Li, Z.; Fei, Z.; Wang, Y.; Fan, C. Insights into the mechanisms underlying ultraviolet-C induced resveratrol metabolism in grapevine (V. amurensis Rupr.) cv.“Tonghua-3”. Front. Plant Sci. 2016, 7, 503. [Google Scholar] [CrossRef]
- Gangopadhyay, M.; Gantait, S.; Palchoudhury, S.; Ali, M.N.; Mondal, C.; Pal, A.K. UVC-priming mediated modulation of forskolin biosynthesis key genes against Macrophomina root rot of Coleus forskohlii—A tissue culture based sustainable approach. Phytochem. Lett. 2016, 17, 36–44. [Google Scholar] [CrossRef]
- Li, D.; Luo, Z.; Mou, W.; Wang, Y.; Ying, T.; Mao, L. ABA and UV-C effects on quality, antioxidant capacity and anthocyanin contents of strawberry fruit (Fragaria ananassa Duch.). Postharvest Biol. Technol. 2014, 90, 56–62. [Google Scholar] [CrossRef]
- Freitas, A.; Moldão-Martins, M.; Costa, H.S.; Albuquerque, T.G.; Valente, A.; Sanches-Silva, A. Effect of UV-C radiation on bioactive compounds of pineapple (Ananas comosus L. Merr.) by-products. J. Sci. Food Agric. 2015, 95, 44–52. [Google Scholar] [CrossRef]
- Lerner, A.B.; Case, J.D.; Takahashi, Y.; Lee, T.H.; Mori, W. Isolation of melatonin, the pineal gland factor that lightens melanocytes 1. J. Am. Chem. Soc. 1958, 80, 2587. [Google Scholar] [CrossRef]
- Tan, D.-X.; Hardeland, R.; Manchester, L.C.; Korkmaz, A.; Ma, S.; Rosales-Corral, S.; Reiter, R.J. Functional roles of melatonin in plants, and perspectives in nutritional and agricultural science. J. Exp. Bot. 2011, 63, 577–597. [Google Scholar] [CrossRef] [Green Version]
- Kolář, J.; Macháčková, I.; Eder, J.; Prinsen, E.; Van Dongen, W.; Van Onckelen, H.; Illnerová, H. Melatonin: Occurrence and daily rhythm in Chenopodium rubrum. Phytochemistry 1997, 44, 1407–1413. [Google Scholar] [CrossRef]
- Murch, S.J.; Saxena, P.K. Melatonin: A potential regulator of plant growth and development? Vitr. Cell. Dev. Biol. Plant 2002, 38, 531–536. [Google Scholar] [CrossRef]
- Arnao, M.B.; Hernández-Ruiz, J. Melatonin promotes adventitious- and lateral root regeneration in etiolated hypocotyls of Lupinus albus L. J. Pineal Res. 2007, 42, 147–152. [Google Scholar] [CrossRef]
- Pelagio-Flores, R.; Muñoz-Parra, E.; Ortiz-Castro, R.; López-Bucio, J. Melatonin regulates Arabidopsis root system architecture likely acting independently of auxin signaling. J. Pineal Res. 2012, 53, 279–288. [Google Scholar] [CrossRef]
- Wolf, K.; Kolář, J.; Witters, E.; van Dongen, W.; van Onckelen, H.; Macháčková, I. Daily profile of melatonin levels in Chenopodium rubrum L. depends on photoperiod. J. Plant Physiol. 2001, 158, 1491–1493. [Google Scholar] [CrossRef]
- Nawaz, M.A.; Huang, Y.; Bie, Z.; Ahmed, W.; Reiter, R.J.; Niu, M.; Hameed, S. Melatonin: Current status and future perspectives in plant science. Front. Plant Sci. 2016, 6, 1230. [Google Scholar] [CrossRef]
- Zhang, N.; Sun, Q.; Zhang, H.; Cao, Y.; Weeda, S.; Ren, S.; Guo, Y.-D. Roles of melatonin in abiotic stress resistance in plants. J. Exp. Bot. 2014, 66, 647–656. [Google Scholar] [CrossRef] [Green Version]
- Lei, X.Y.; Zhu, R.Y.; Zhang, G.Y.; Dai, Y.R. Attenuation of cold-induced apoptosis by exogenous melatonin in carrot suspension cells: The possible involvement of polyamines. J. Pineal Res. 2004, 36, 126–131. [Google Scholar] [CrossRef]
- Li, C.; Wang, P.; Wei, Z.; Liang, D.; Liu, C.; Yin, L.; Jia, D.; Fu, M.; Ma, F. The mitigation effects of exogenous melatonin on salinity-induced stress in Malus hupehensis. J. Pineal Res. 2012, 53, 298–306. [Google Scholar] [CrossRef]
- Wei, W.; Li, Q.-T.; Chu, Y.-N.; Reiter, R.J.; Yu, X.-M.; Zhu, D.-H.; Zhang, W.-K.; Ma, B.; Lin, Q.; Zhang, J.-S. Melatonin enhances plant growth and abiotic stress tolerance in soybean plants. J. Exp. Bot. 2014, 66, 695–707. [Google Scholar] [CrossRef] [Green Version]
- Tan, D.-X.; Manchester, L.C.; Helton, P.; Reiter, R.J. Phytoremediative capacity of plants enriched with melatonin. Plant Signal. Behav. 2007, 2, 514–516. [Google Scholar] [CrossRef]
- Gai, Q.-Y.; Jiao, J.; Luo, M.; Wang, W.; Zhao, C.-J.; Fu, Y.-J.; Ma, W. UV elicitation for promoting astragaloside production in Astragalus membranaceus hairy root cultures with transcriptional expression of biosynthetic genes. Ind. Crop. Prod. 2016, 84, 350–357. [Google Scholar] [CrossRef]
- Nawkar, G.; Maibam, P.; Park, J.; Sahi, V.; Lee, S.; Kang, C. UV-induced cell death in plants. Int. J. Mol. Sci. 2013, 14, 1608–1628. [Google Scholar] [CrossRef]
- Xu, A.; Zhan, J.-C.; Huang, W.-D. Effects of ultraviolet C, methyl jasmonate and salicylic acid, alone or in combination, on stilbene biosynthesis in cell suspension cultures of Vitis vinifera L. cv. Cabernet Sauvignon. Plant Cell Tissue Organ Cult. 2015, 122, 197–211. [Google Scholar] [CrossRef]
- Xu, A.; Zhan, J.-C.; Huang, W.-D. Combined elicitation of chitosan and ultraviolet C enhanced stilbene production and expression of chitinase and β-1, 3-glucanase in Vitis vinifera cell suspension cultures. Plant Cell Tissue Organ Cult. 2016, 124, 105–117. [Google Scholar] [CrossRef]
- Anjum, S.; Abbasi, B.H.; Doussot, J.; Favre-Réguillon, A.; Hano, C. Effects of photoperiod regimes and ultraviolet-C radiations on biosynthesis of industrially important lignans and neolignans in cell cultures of Linum usitatissimum L. (Flax). J. Photochem. Photobiol. B Biol. 2017, 167, 216–227. [Google Scholar] [CrossRef]
- Khan, T.; Ullah, M.A.; Garros, L.; Hano, C.; Abbasi, B.H. Synergistic effects of melatonin and distinct spectral lights for enhanced production of anti-cancerous compounds in callus cultures of Fagonia indica. J. Photochem. Photobiol. B Biol. 2018. [Google Scholar] [CrossRef]
- Abbasi, B.H.; Khan, M.; Guo, B.; Bokhari, S.A.; Khan, M.A. Efficient regeneration and antioxidative enzyme activities in Brassica rapa var. turnip. Plant Cell Tissue Organ Cult. 2011, 105, 337–344. [Google Scholar] [CrossRef]
- Fazal, H.; Abbasi, B.H.; Ahmad, N.; Ali, M. Elicitation of medicinally important antioxidant secondary metabolites with silver and gold nanoparticles in callus cultures of Prunella vulgaris L. Appl. Biochem. Biotechnol. 2016, 180, 1076–1092. [Google Scholar] [CrossRef]
- Sarropoulou, V.N.; Therios, I.N.; Dimassi-Theriou, K.N. Melatonin promotes adventitious root regeneration in in vitro shoot tip explants of the commercial sweet cherry rootstocks CAB-6P (Prunus cerasus L.), Gisela 6 (P. cerasus × P. canescens), and MxM 60 (P. avium × P. mahaleb). J. Pineal Res. 2012, 52, 38–46. [Google Scholar] [CrossRef]
- Fazal, H.; Abbasi, B.H.; Ahmad, N.; Ali, M. Exogenous melatonin trigger biomass accumulation and production of stress enzymes during callogenesis in medicinally important Prunella vulgaris L. (Selfheal). Physiol. Mol. Biol. Plants 2018, 24, 1307–1315. [Google Scholar] [CrossRef]
- Urban, L.; Charles, F.; de Miranda, M.R.A.; Aarrouf, J. Understanding the physiological effects of UV-C light and exploiting its agronomic potential before and after harvest. Plant Physiol. Biochem. 2016, 105, 1–11. [Google Scholar] [CrossRef]
- Carbonell-Bejerano, P.; Diago, M.-P.; Martínez-Abaigar, J.; Martínez-Zapater, J.M.; Tardáguila, J.; Núñez-Olivera, E. Solar ultraviolet radiation is necessary to enhance grapevine fruit ripening transcriptional and phenolic responses. BMC Plant Biol. 2014, 14, 183. [Google Scholar] [CrossRef]
- Nigro, F.; Ippolito, A.; Lattanzio, V.; Di Venere, D.; Salerno, M. Effect of ultraviolet-C light on postharvest decay of strawberry. J. Plant Pathol. 2000, 82, 29–37. [Google Scholar]
- Tiecher, A.; de Paula, L.A.; Chaves, F.C.; Rombaldi, C.V. UV-C effect on ethylene, polyamines and the regulation of tomato fruit ripening. Postharvest Biol. Technol. 2013, 86, 230–239. [Google Scholar] [CrossRef]
- Khan, T.; Abbasi, B.H.; Afridi, M.S.; Tanveer, F.; Ullah, I.; Bashir, S.; Hano, C. Melatonin-enhanced biosynthesis of antimicrobial AgNPs by improving the phytochemical reducing potential of a callus culture of Ocimum basilicum L. var. thyrsiflora. RSC Adv. 2017, 7, 38699–38713. [Google Scholar]
- Adil, M.; Abbasi, B.H.; Khan, T. Interactive effects of melatonin and light on growth parameters and biochemical markers in adventitious roots of Withania somnifera L. Plant Cell Tissue Organ Cult. 2015, 123, 405–412. [Google Scholar] [CrossRef]
- Moran, J.F.; Becana, M.; Iturbe-Ormaetxe, I.; Frechilla, S.; Klucas, R.V.; Aparicio-Tejo, P. Drought induces oxidative stress in pea plants. Planta 1994, 194, 346–352. [Google Scholar] [CrossRef] [Green Version]
- Alexieva, V.; Sergiev, I.; Mapelli, S.; Karanov, E. The effect of drought and ultraviolet radiation on growth and stress markers in pea and wheat. Plant Cell Environ. 2001, 24, 1337–1344. [Google Scholar] [CrossRef] [Green Version]
- Yu, T.-W.; Anderson, D. Reactive oxygen species-induced DNA damage and its modification: A chemical investigation. Mutat. Res. Fundam. Mol. Mech. Mutagenesis 1997, 379, 201–210. [Google Scholar] [CrossRef]
- Suzuki, N.; Mittler, R. Reactive oxygen species and temperature stresses: A delicate balance between signaling and destruction. Physiol. Plant. 2006, 126, 45–51. [Google Scholar] [CrossRef] [Green Version]
- Bowler, C.; Montagu, M.V.; Inze, D. Superoxide dismutase and stress tolerance. Annu. Rev. Plant Biol. 1992, 43, 83–116. [Google Scholar] [CrossRef]
- Fridovich, I. Superoxide dismutases. Annu. Rev. Biochem. 1975, 44, 147–159. [Google Scholar]
- Meloni, D.A.; Oliva, M.A.; Martinez, C.A.; Cambraia, J. Photosynthesis and activity of superoxide dismutase, peroxidase and glutathione reductase in cotton under salt stress. Environ. Exp. Bot. 2003, 49, 69–76. [Google Scholar] [CrossRef]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Ashry, N.A.; Mohamed, H.I. Impact of secondary metabolites and related enzymes in flax resistance and or susceptibility to powdery mildew. World J. Agric. Sci. 2011, 7, 78–85. [Google Scholar]
- Samuolienė, G.; Brazaitytė, A.; Urbonavičiūtė, A.; Šabajevienė, G.; Duchovskis, P. The effect of red and blue light component on the growth and development of frigo strawberries. Zemdirb. Agric. 2010, 97, 99–104. [Google Scholar]
- Djeridane, A.; Yousfi, M.; Nadjemi, B.; Boutassouna, D.; Stocker, P.; Vidal, N. Antioxidant activity of some Algerian medicinal plants extracts containing phenolic compounds. Food Chem. 2006, 97, 654–660. [Google Scholar] [CrossRef]
- Kim, D.-O.; Chun, O.K.; Kim, Y.J.; Moon, H.-Y.; Lee, C.Y. Quantification of polyphenolics and their antioxidant capacity in fresh plums. J. Agric. Food Chem. 2003, 51, 6509–6515. [Google Scholar] [CrossRef]
- Forbes, J.M.; Soldatos, G.; Thomas, M.C. Below the radar: Advanced glycation end products that detour “around the side”: Is HbA1c not an accurate enough predictor of long term progression and glycaemic control in diabetes? Clin. Biochem. Rev. /Aust. Assoc. Clin. Biochem. 2005, 26, 123. [Google Scholar]
- Jang, C.; Lim, J.H.; Park, C.W.; Cho, Y.-J. Regulator of calcineurin 1 isoform 4 (RCAN1. 4) Is overexpressed in the glomeruli of diabetic mice. Korean J. Physiol. Pharmacol. 2011, 15, 299–305. [Google Scholar] [CrossRef]
- Hunaefi, D.; Yuliana, N.; Smetanska, I.; Gruda, N. Effect of Ultraviolet and Ultrasonic on Potential Antidiabetic Activity of In Vitro Shoot Cultures of Orthosiphon Aristatus; IOP Conference Series: Earth and Environmental Science; IOP Publishing: Bristol, UK, 2018; p. 012008. [Google Scholar]
- Hano, C.; Renouard, S.; Molinié, R.; Corbin, C.; Barakzoy, E.; Doussot, J.; Lamblin, F.; Lainé, E. Flaxseed (Linum usitatissimum L.) extract as well as (+)-secoisolariciresinol diglucoside and its mammalian derivatives are potent inhibitors of α-amylase activity. Bioorganic Med. Chem. Lett. 2013, 23, 3007–3012. [Google Scholar] [CrossRef]
- Eddouks, M.; Maghrani, M.; Zeggwagh, N.-A.; Michel, J. Study of the hypoglycaemic activity of Lepidium sativum L. aqueous extract in normal and diabetic rats. J. Ethnopharmacol. 2005, 97, 391–395. [Google Scholar] [CrossRef]
- Chauhan, K.; Sharma, S.; Agarwal, N.; Chauhan, S.; Chauhan, B. A study on potential hypoglycemic and hypolipidemic effects of Lepidium Sativum (Garden Cress) in Alloxan induced diabetic rats. Am. J. Pharm. Tech. Res 2012, 2, 522–535. [Google Scholar]
- Shukla, A.; Bigoniya, P.; Srivastava, B. Hypoglycemic activity of Lepidium sativum Linn seed total alkaloid on alloxan induced diabetic rats. Res. J. Med. Plant 2012, 6, 587–596. [Google Scholar]
- Fraser, D.; Hansen, K. Making sense of advanced glycation end products and their relevance to diabetic complications. Inter Diabetes Monit 2005, 17, 1–7. [Google Scholar]
- Grillo, M.; Colombatto, S. Advanced glycation end-products (AGEs): Involvement in aging and in neurodegenerative diseases. Amino Acids 2008, 35, 29–36. [Google Scholar] [CrossRef]
- Balasundram, N.; Sundram, K.; Samman, S. Phenolic compounds in plants and agri-industrial by-products: Antioxidant activity, occurrence, and potential uses. Food Chem. 2006, 99, 191–203. [Google Scholar] [CrossRef]
- Dajas, F. Life or death: Neuroprotective and anticancer effects of quercetin. J. Ethnopharmacol. 2012, 143, 383–396. [Google Scholar] [CrossRef]
- Kashyap, D.; Sharma, A.; Tuli, H.S.; Sak, K.; Punia, S.; Mukherjee, T.K. Kaempferol—A dietary anticancer molecule with multiple mechanisms of action: Recent trends and advancements. J. Funct. Foods 2017, 30, 203–219. [Google Scholar] [CrossRef]
- Jiang, Y.; Kusama, K.; Satoh, K.; Takayama, F.; Watanabe, S.; Sakagami, H. Induction of cytotoxicity by chlorogenic acid in human oral tumor cell lines. Phytomedicine 2000, 7, 483–491. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Ullah, M.A. Trends in polyphenols accumulation in in vitro callus cultures of Lepidium sativum Linn. under different monochromatic lights. J. Photochem. Photobiol. B 2019. in revision. [Google Scholar]
- Nayyar, H.; Gupta, D. Differential sensitivity of C3 and C4 plants to water deficit stress: Association with oxidative stress and antioxidants. Environ. Exp. Bot. 2006, 58, 106–113. [Google Scholar] [CrossRef]
- Zahir, A.; Abbasi, B.H.; Adil, M.; Anjum, S.; Zia, M. Synergistic effects of drought stress and photoperiods on phenology and secondary metabolism of Silybum marianum. Appl. Biochem. Biotechnol. 2014, 174, 693–707. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Abbasi, B.H.; Khan, M.A.; Mahmood, T.; Ahmad, M.; Chaudhary, M.F.; Khan, M.A. Shoot regeneration and free-radical scavenging activity in Silybum marianum L. Plant Cell Tissue Organ Cult. 2010, 101, 371–376. [Google Scholar] [CrossRef]
- Benzie, I.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef]
- Tagliazucchi, D.; Verzelloni, E.; Bertolini, D.; Conte, A. In vitro bio-accessibility and antioxidant activity of grape polyphenols. Food Chem. 2010, 120, 599–606. [Google Scholar] [CrossRef]
- Kaewseejan, N.; Siriamornpun, S. Bioactive components and properties of ethanolic extract and its fractions from Gynura procumbens leaves. Ind. Crop. Prod. 2015, 74, 271–278. [Google Scholar] [CrossRef]
- Lagrimini, L. Plant peroxidases: Under-and over-expression in transgenic plants and physiological consequences. Plant Peroxidases 1980, 1990, 59–69. [Google Scholar]
- Giannopolitis, C.N.; Ries, S.K. Superoxide dismutases: I. Occurrence in higher plants. Plant. Physiol. 1977, 59, 309–314. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Conconcentrations | Inhibition of Advanced Glycation End Products Formation (AGE) | Antidiabetic Activity | ||
|---|---|---|---|---|---|
| Vesperlysine-Like AGEs (% Inhibition) | Pentosidine-Like AGEs (% Inhibition) | α-Glucosidase (% Inhibition) | α-Amylase (% Inhibition) | ||
| Control | TDZ+NAA | 24.28 ± 1.057 cd | 28.96 ± 1.444 d | 25.83 ± 1.057 d | 23.41 ± 1.443 e |
| UV-C (min) | 10 | 35.89 ± 0.835 bc | 52.15 ± 2.018 b | 37.86 ± 0.835 c | 47.34 ± 2.018 bc |
| 30 | 37.45 ± 1.994 bc | 51.44 ± 1.702 b | 36.93 ± 1.999 c | 46.80 ± 1.726 bc | |
| 60 | 42.62 ± 1.520 b | 57.72 ± 3.101 ab | 48.59 ± 1.528 b | 55.54 ± 0.942 b | |
| 90 | 30.18 ± 2.018 c | 38.02 ± 1.973 c | 41.55 ± 2.018 bc | 35.47 ± 1.953 d | |
| 120 | 27.11 ± 1.720 cd | 33.77 ± 1.093 cd | 26.96 ± 1.720 d | 31.75 ± 1.049 d | |
| 150 | 36.75 ± 0.998 bc | 47.51 ± 1.057 b | 32.38 ± 0.998 cd | 43.50 ± 1.033 c | |
| Melatonin (μM) | 5 | 33.45 ± 1.951 bc | 42.95 ± 0.845 bc | 27.49 ± 1.952 d | 39.49 ± 0.835 cd |
| 10 | 37.02 ± 1.048 bc | 48.07 ± 1.992 b | 33.29 ± 1.047 cd | 43.85 ± 1.839 c | |
| 15 | 46.95 ± 2.060 ab | 62.36 ± 1.711 a | 49.55 ± 2.060 b | 55.98 ± 1.528 b | |
| 20 | 52.47 ± 2.593 a | 57.01 ± 1.570 ab | 57.84 ± 2.591 a | 62.66 ± 1.720 a | |
| 25 | 31.88 ± 1.668 c | 54.87 ± 0.995 ab | 40.73 ± 1.668 bc | 49.75 ± 0.936 bc | |
| 50 | 42.46 ± 0.946 b | 55.70 ± 1.947 ab | 41.68 ± 0.942 bc | 50.46 ± 1.793 bc | |
| Treatment | Concentration | Polyphenolic Compounds (mg/g DW) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Caffeic Acid | Ferulic Acid | Vanillic Acid | p-Coumaric Acid | Sinapic Acid | Protocatechuic Acid | Chlorogenic Acid | Quercetin | Kaempferol | Total | ||
| Control | TDZ+NAA | 0.323 ± 0.01 cd | 0.224 ± 0.007 bc | 0.076 ± 0.003 c | 0.129 ± 0.034 b | 0.044 ± 0.009 bc | 0.037 ± 0.0021 c | 2.86 ± 0.63 de | 7.58 ± 0.941 d | 2.64 ± 0.573 e | 13.94 |
| UV-C (min) | 10 | 0.682 ± 0.02 b | 0.462 ± 0.04 ab | 0.140 ± 0.005 ab | 0.158 ± 0.025 a | 0.065 ± 0.007 ab | 0.049 ± 0.0047 b | 5.24 ± 0.84 bc | 15.43 ± 1.843 b | 5.42 ± 1.053 bc | 27.67 |
| 30 | 0.675 ± 0.009 b | 0.458 ± 0.027 ab | 0.135 ± 0.019 b | 0.129 ± 0.019 b | 0.063 ± 0.003 ab | 0.048 ± 0.0085 b | 5.17 ± 0.93 bc | 15.26 ± 1.909 b | 5.36 ± 0.593 bc | 27.32 | |
| 60 | 0.806 ± 0.06 ab | 0.545 ± 0.017 a | 0.158 ± 0.006 a | 0.135 ± 0.064 ab | 0.071 ± 0.005 a | 0.052 ± 0.0031 ab | 6.04 ± 0.48 ab | 18.13 ± 2.015 ab | 6.37 ± 1.953 ab | 32.33 | |
| 90 | 0.506 ± 0.03 bc | 0.345 ± 0.013 b | 0.103 ± 0.004 bc | 0.077 ± 0.004 c | 0.052 ± 0.001 b | 0.041 ± 0.0016 bc | 4.04 ± 0.41 cd | 11.56 ± 0.931 c | 4.05 ± 0.351 d | 20.78 | |
| 120 | 0.448 ± 0.024 c | 0.307 ± 0.038 b | 0.099 ± 0.001 c | 0.122 ± 0.03 b | 0.052 ± 0.004 b | 0.042 ± 0.0063 bc | 3.70 ± 0.52 cd | 10.32 ± 0.683 cd | 3.61 ± 0.683 de | 18.72 | |
| 150 | 0.625 ± 0.043 b | 0.425 ± 0.047 ab | 0.130 ± 0.049 b | 0.066 ± 0.005 c | 0.062 ± 0.008 ab | 0.047 ± 0.0074 b | 4.87 ± 0.48 bc | 14.19 ± 0.946 bc | 4.98 ± 0.395 c | 25.40 | |
| Melatonin (μM) | 5 | 0.563 ± 0.053 bc | 0.384 ± 0.042 b | 0.123 ± 0.031 b | 0.155 ± 0.029 a | 0.060 ± 0.004 ab | 0.047 ± 0.0023 b | 4.48 ± 0.17 c | 12.85 ± 1.042 c | 4.50 ± 0.445 cd | 23.19 |
| 10 | 0.629 ± 0.062 b | 0.427 ± 0.009 ab | 0.132 ± 0.002 b | 0.175 ± 0.006 a | 0.063 ± 0.002 ab | 0.048 ± 0.0019 b | 4.90 ± 0.52 bc | 14.28 ± 2.19 bc | 5.01 ± 1.053 c | 25.69 | |
| 15 | 0.809 ± 0.061 ab | 0.548 ± 0.053 a | 0.175 ± 0.035 a | 0.140 ± 0.018 ab | 0.079 ± 0.006 a | 0.057 ± 0.0067 a | 6.15 ± 0.201 ab | 18.26 ± 0.538 ab | 6.41 ± 1.539 ab | 32.65 | |
| 20 | 0.915 ± 0.028 a | 0.617 ± 0.073 a | 0.168 ± 0.02 a | 0.116 ± 0.004 b | 0.073 ± 0.007 a | 0.053 ± 0.0043 ab | 6.72 ± 0.42 a | 20.48 ± 2.638 a | 7.21 ± 0.213 a | 36.35 | |
| 25 | 0.719 ± 0.083 b | 0.487 ± 0.041 ab | 0.145 ± 0.018 ab | 0.082 ± 0.001 bc | 0.067 ± 0.009 ab | 0.050 ± 0.0064 ab | 5.48 ± 0.106 b | 16.24 ± 1.503 b | 5.70 ± 0.573 b | 28.99 | |
| 50 | 0.730 ± 0.042 b | 0.494 ± 0.098 ab | 0.147 ± 0.007 ab | 0.082 ± 0.003 bc | 0.067 ± 0.001 ab | 0.050 ± 0.0028 ab | 5.55 ± 0.113 b | 16.47 ± 1.548 b | 5.79 ± 0.443 b | 29.39 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ullah, M.A.; Tungmunnithum, D.; Garros, L.; Drouet, S.; Hano, C.; Abbasi, B.H. Effect of Ultraviolet-C Radiation and Melatonin Stress on Biosynthesis of Antioxidant and Antidiabetic Metabolites Produced in In Vitro Callus Cultures of Lepidium sativum L. Int. J. Mol. Sci. 2019, 20, 1787. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20071787
Ullah MA, Tungmunnithum D, Garros L, Drouet S, Hano C, Abbasi BH. Effect of Ultraviolet-C Radiation and Melatonin Stress on Biosynthesis of Antioxidant and Antidiabetic Metabolites Produced in In Vitro Callus Cultures of Lepidium sativum L. International Journal of Molecular Sciences. 2019; 20(7):1787. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20071787
Chicago/Turabian StyleUllah, Muhammad Asad, Duangjai Tungmunnithum, Laurine Garros, Samantha Drouet, Christophe Hano, and Bilal Haider Abbasi. 2019. "Effect of Ultraviolet-C Radiation and Melatonin Stress on Biosynthesis of Antioxidant and Antidiabetic Metabolites Produced in In Vitro Callus Cultures of Lepidium sativum L." International Journal of Molecular Sciences 20, no. 7: 1787. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20071787