C1 Compound Biosensors: Design, Functional Study, and Applications

 , , and
, , and
Abstract
:1. Introduction
2. Results and Discussion
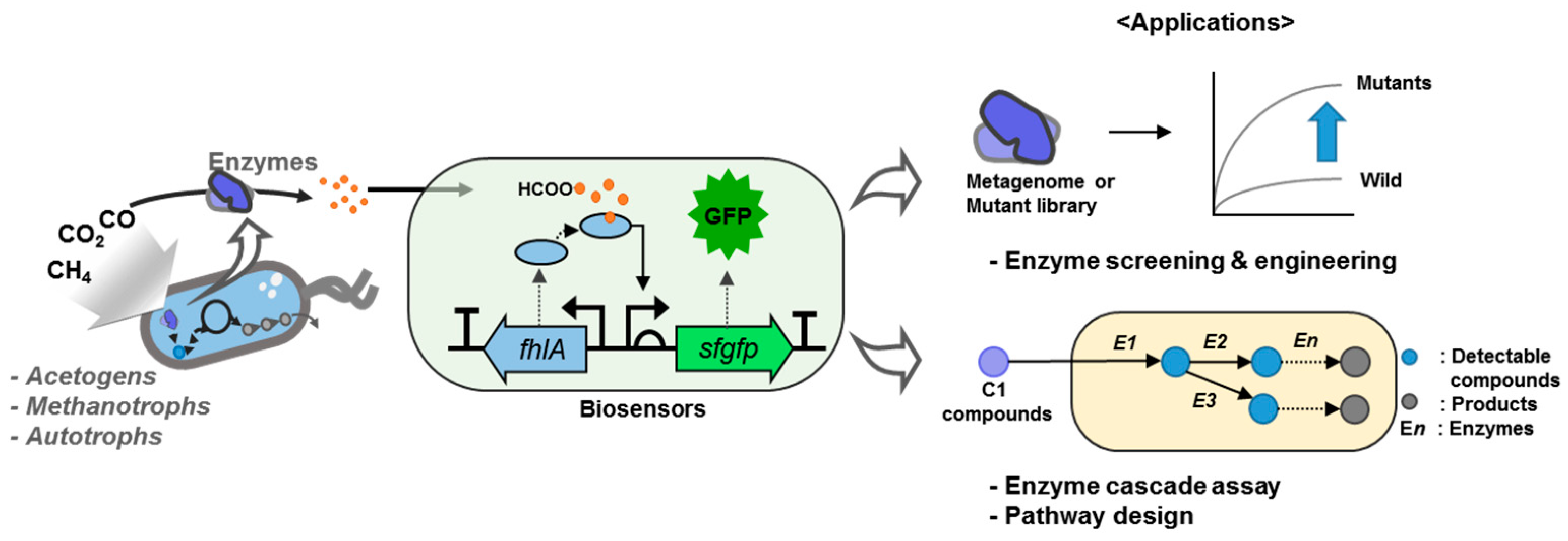
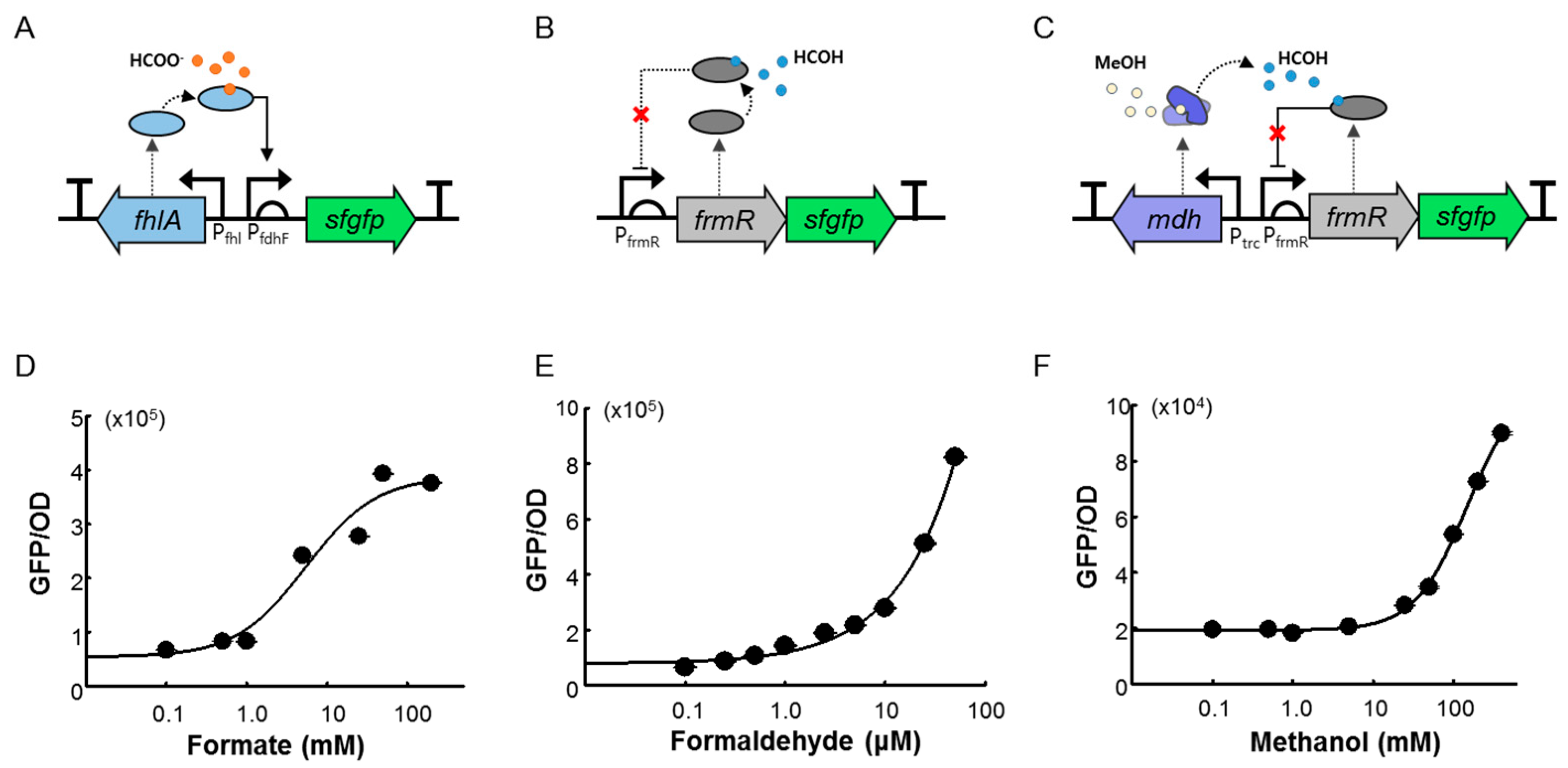
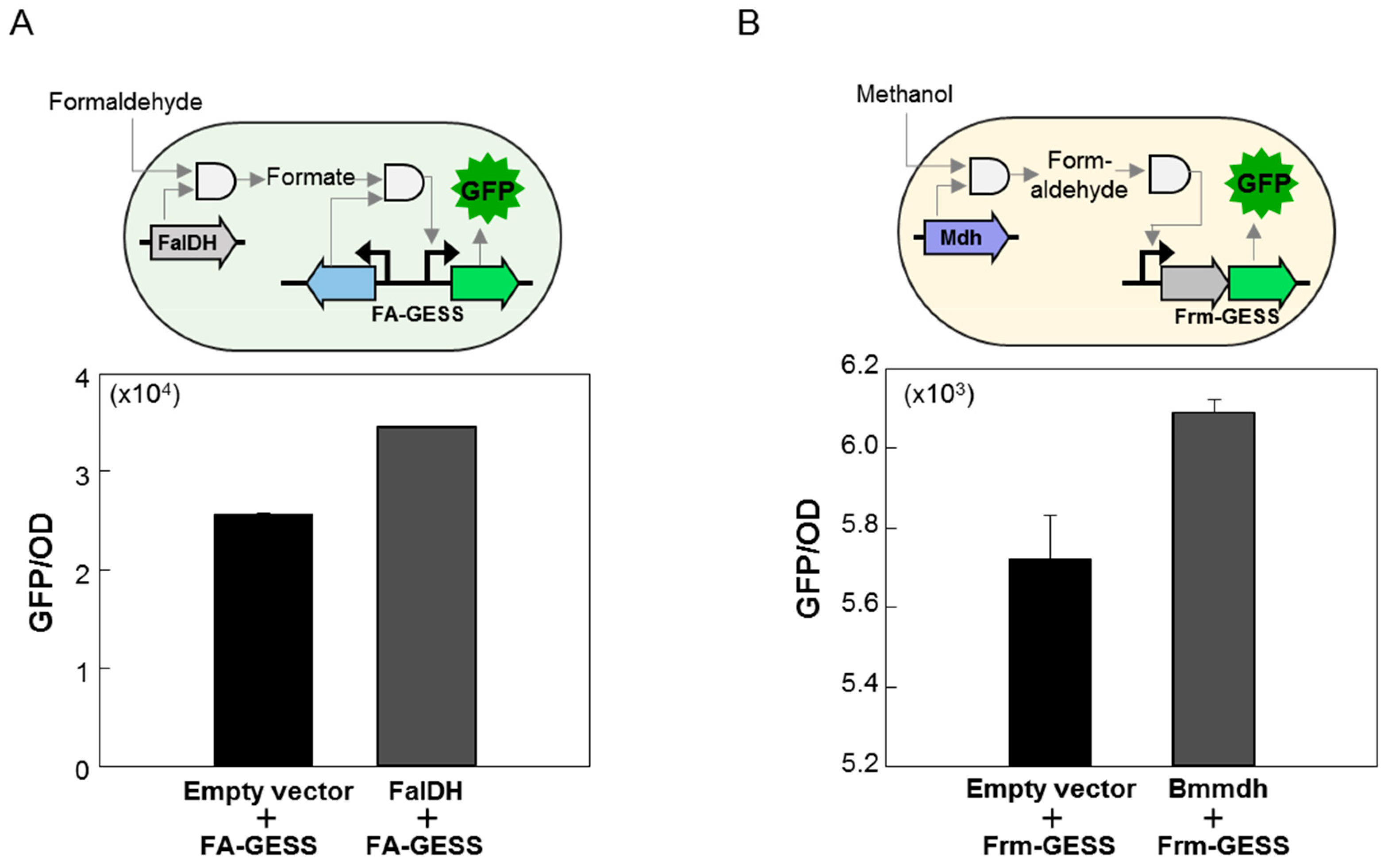
2.1. Design and Construction of the C1 Biosensors
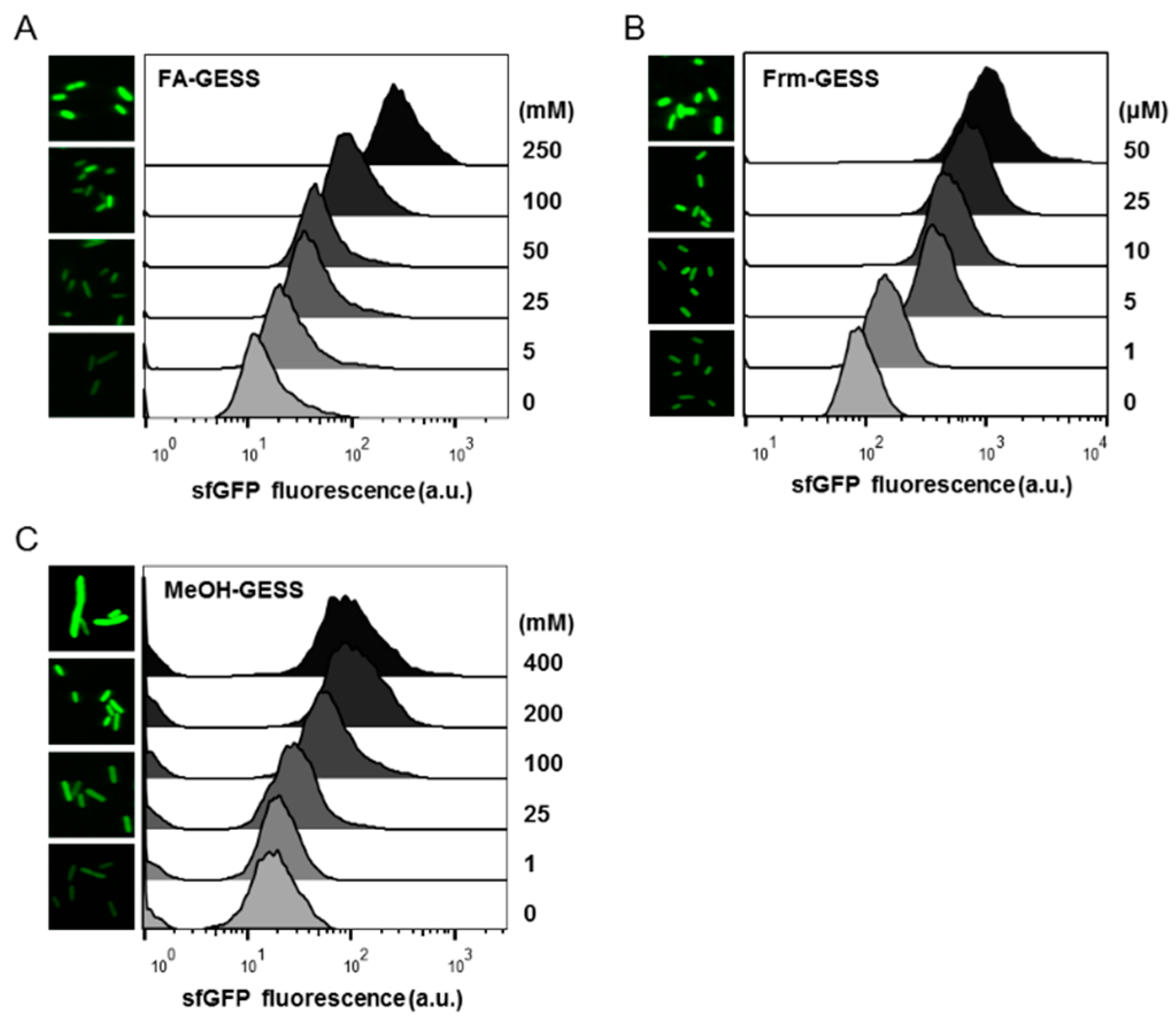
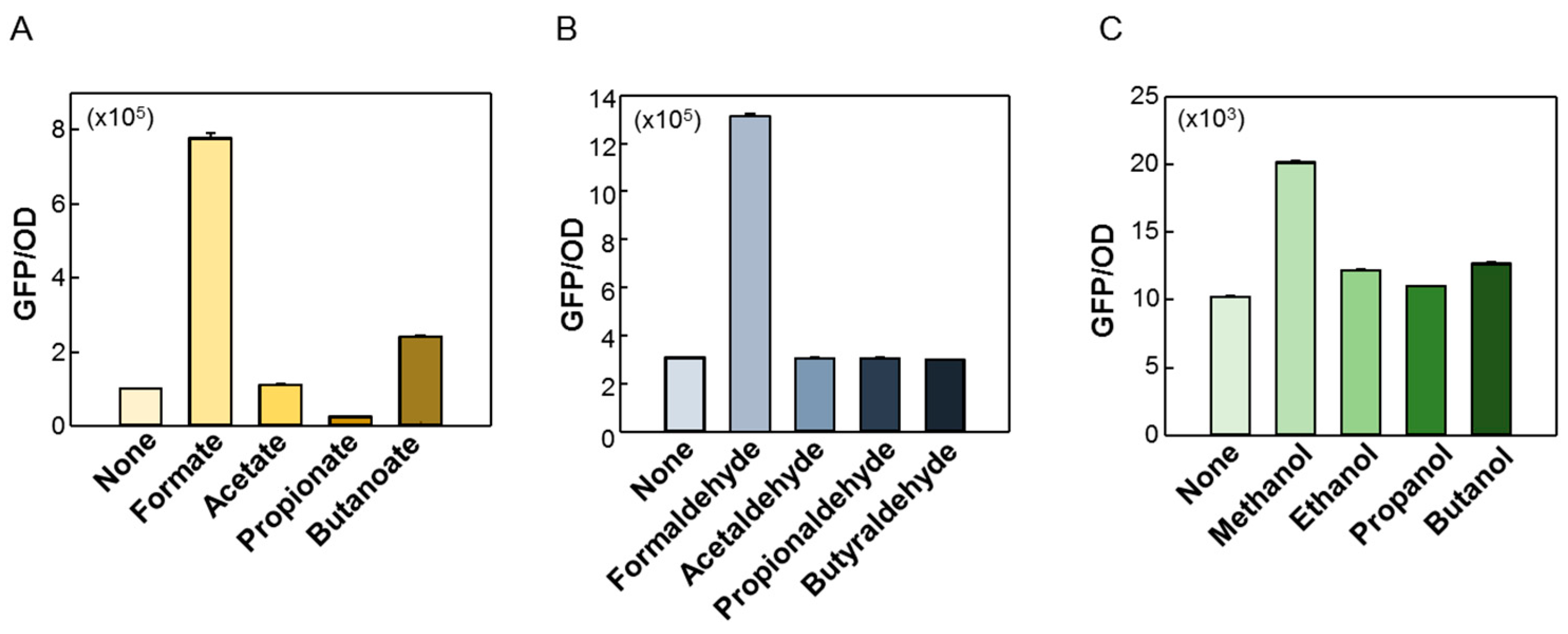
2.2. Quantitative Responses of FA-GESS, Frm-GESS, and MeOH-GESS
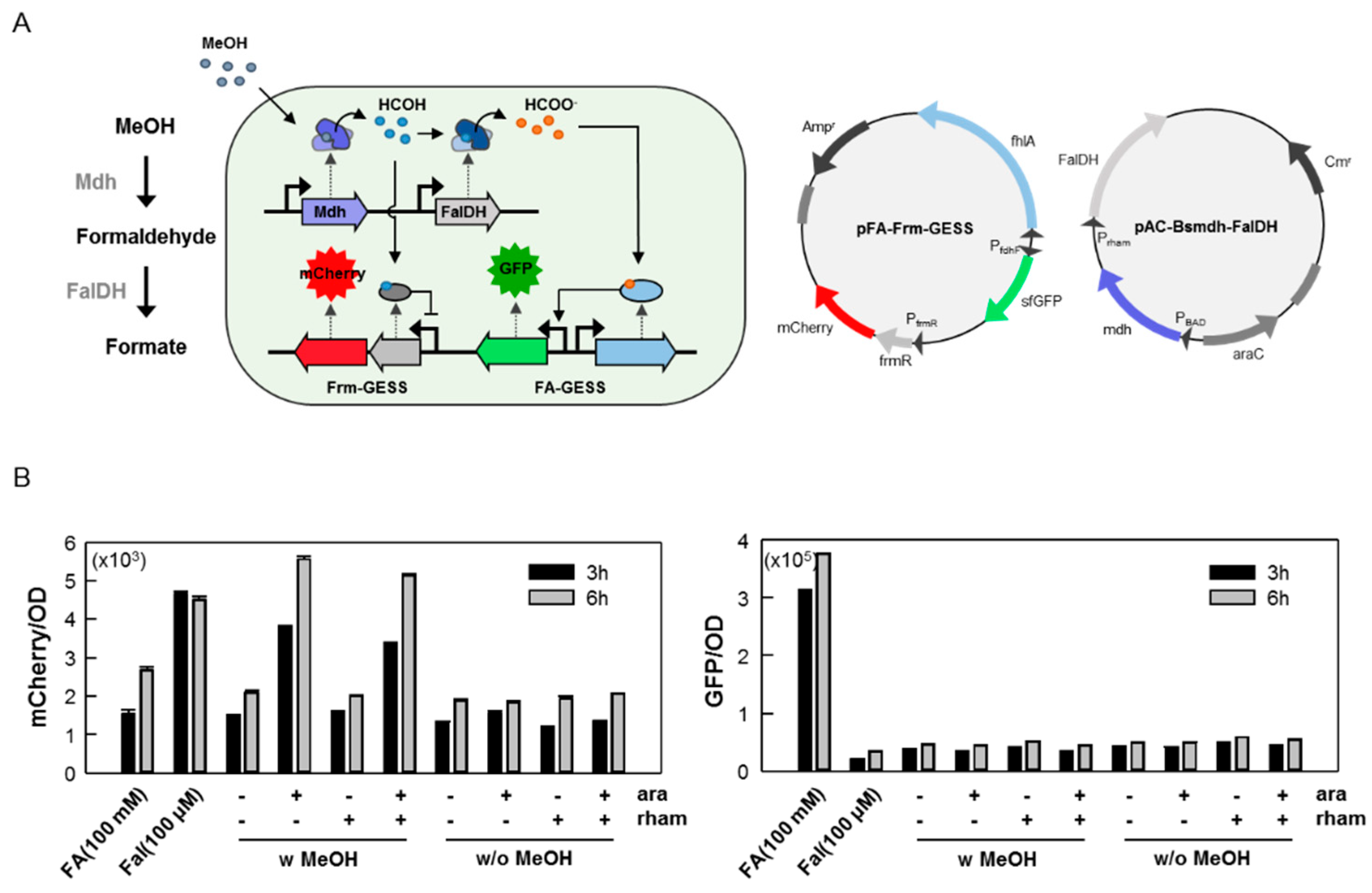
2.3. Use of the C1 Biosensor to Detect C1-converting Enzyme Activities
2.4. Perspective of C1-Detectable Biosensors
3. Methods
3.1. Materials
3.2. Construction of Formate, Formaldehyde, and Methanol Biosensors
3.3. Fluorescence-Based Functional Analysis of C1 Biosensors
3.4. Measurement of the Formaldehyde Dehydrogenase and Methanol Dehydrogenase Activity Using FA-GESS and Frm-GESS
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| Mdh | Methanol dehydrogenase |
| FalDH | Formaldehyde dehydrogenase |
| Bmmdh | Methanol dehydrogenase from B. methanolicus |
| Bsmdh | Methanol dehydrogenase from B. stearothermophilus |
| FalDH | Formaldehyde dehydrogenase from P. putida |
| MMOs | Methane monooxygenase |
| IPTG | Isopropyl β-d-1-thiogalactopyranoside |
| TF | Transcription factor |
References
- Strong, P.J.; Xie, S.; Clarke, W.P. Methane as a resource: Can the methanotrophs add value? Environ. Sci. Technol. 2015, 49, 4001–4018. [Google Scholar] [CrossRef] [PubMed]
- Pfeifenschneider, J.; Brautaset, T.; Wendisch, V.F. Methanol as carbon substrate in the bio-economy: Metabolic engineering of aerobic methylotrophic bacteria for production of value-added chemicals. BiofuelsBioprod. Biorefin. 2017, 11, 719. [Google Scholar] [CrossRef]
- Bang, J.; Lee, S.Y. Assimilation of formic acid and CO2 by engineered Escherichia coli equipped with reconstructed one-carbon assimilation pathways. Proc. Natl. Acad. Sci. USA 2018, 115, E9271–E9279. [Google Scholar] [CrossRef]
- Yishai, O.; Lindner, S.N.; Gonzalez de la Cruz, J.; Tenenboim, H.; Bar-Even, A. The formate bio-economy. Curr. Opin. Chem. Biol. 2016, 35, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Latif, H.; Zeidan, A.A.; Nielsen, A.T.; Zengler, K. Trash to treasure: Production of biofuels and commodity chemicals via syngas fermenting microorganisms. Curr. Opin. Biotechnol. 2014, 27, 79–87. [Google Scholar] [CrossRef]
- Ragsdale, S.W. Enzymology of the wood-Ljungdahl pathway of acetogenesis. Ann. N. Y. Acad. Sci. 2008, 1125, 129–136. [Google Scholar] [CrossRef]
- Kalyuzhnaya, M.G.; Yang, S.; Rozova, O.N.; Smalley, N.E.; Clubb, J.; Lamb, A.; Gowda, G.A.; Raftery, D.; Fu, Y.; Bringel, F.; et al. Highly efficient methane biocatalysis revealed in a methanotrophic bacterium. Nat. Commun. 2013, 4, 2785. [Google Scholar] [CrossRef] [Green Version]
- Lawton, T.J.; Rosenzweig, A.C. Methane-oxidizing enzymes: An upstream problem in biological gas-to-liquids conversion. J. Am. Chem. Soc. 2016, 138, 9327–9340. [Google Scholar] [CrossRef]
- Bennett, R.K.; Steinberg, L.M.; Chen, W.; Papoutsakis, E.T. Engineering the bioconversion of methane and methanol to fuels and chemicals in native and synthetic methylotrophs. Curr. Opin. Biotechnol. 2018, 50, 81–93. [Google Scholar] [CrossRef] [PubMed]
- Bennett, R.K.; Gonzalez, J.E.; Whitaker, W.B.; Antoniewicz, M.R.; Papoutsakis, E.T. Expression of heterologous non-oxidative pentose phosphate pathway from Bacillus methanolicus and phosphoglucose isomerase deletion improves methanol assimilation and metabolite production by a synthetic Escherichia coli methylotroph. Metab. Eng. 2018, 45, 75–85. [Google Scholar] [CrossRef]
- Muller, J.E.N.; Meyer, F.; Litsanov, B.; Kiefer, P.; Potthoff, E.; Heux, S.; Quax, W.J.; Wendisch, V.F.; Brautaset, T.; Portais, J.C.; et al. Engineering Escherichia coli for methanol conversion. Metab. Eng. 2015, 28, 190–201. [Google Scholar] [CrossRef] [PubMed]
- Whitaker, W.B.; Jones, J.A.; Bennett, R.K.; Gonzalez, J.E.; Vernacchio, V.R.; Collins, S.M.; Palmer, M.A.; Schmidt, S.; Antoniewicz, M.R.; Koffas, M.A.; et al. Engineering the biological conversion of methanol to specialty chemicals in Escherichia coli. Metab. Eng. 2017, 39, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Yishai, O.; Bouzon, M.; Doring, V.; Bar-Even, A. In Vivo Assimilation of One-Carbon via a Synthetic Reductive Glycine Pathway in Escherichia coli. ACS Synth. Biol. 2018, 7, 2023–2028. [Google Scholar] [CrossRef]
- Kwon, K.K.; Lee, D.-H.; Kim, S.J.; Choi, S.-L.; Rha, E.; Yeom, S.-J.; Subhadra, B.; Lee, J.; Jeong, K.J.; Lee, S.-G. Evolution of enzymes with new specificity by high-throughput screening using DmpR-based genetic circuits and multiple flow cytometry rounds. Sci. Rep. 2018, 8, 2659. [Google Scholar] [CrossRef] [Green Version]
- Choi, S.L.; Rha, E.; Lee, S.J.; Kim, H.; Kwon, K.; Jeong, Y.S.; Rhee, Y.H.; Song, J.J.; Kim, H.S.; Lee, S.G. Toward a generalized and high-throughput enzyme screening system based on artificial genetic circuits. ACS Synth. Biol. 2014, 3, 163–171. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Rha, E.; Seong, W.; Yeom, S.J.; Lee, D.H.; Lee, S.G. A cell-cell communication-based screening system for novel microbes with target enzyme activities. ACS Synth. Biol. 2016, 5, 1231–1238. [Google Scholar] [CrossRef] [PubMed]
- Yeom, S.J.; Kim, M.; Kwon, K.K.; Fu, Y.; Rha, E.; Park, S.H.; Lee, H.; Kim, H.; Lee, D.H.; Kim, D.M.; et al. A synthetic microbial biosensor for high-throughput screening of lactam biocatalysts. Nat. Commun. 2018, 9, 5053. [Google Scholar] [CrossRef]
- Fu, Y.; Yeom, S.J.; Kwon, K.K.; Hwang, J.; Kim, H.; Woo, E.J.; Lee, D.H.; Lee, S.G. Structural and functional analyses of the cellulase transcription regulator CelR. FEBS Lett. 2018, 592, 2776–2785. [Google Scholar] [CrossRef]
- Kwon, K.K.; Yeom, S.J.; Lee, D.H.; Jeong, K.J.; Lee, S.G. Development of a novel cellulase biosensor that detects crystalline cellulose hydrolysis using a transcriptional regulator. Biochem. Biophys. Res. Commun. 2018, 495, 1328–1334. [Google Scholar] [CrossRef]
- Yang, X.A.; Zweifach, A. Temperature-Dependent Expression of a CFP-YFP FRET Diacylglycerol Sensor Enables Multiple-Read Screening for Compounds That Affect C1 Domains. SLAS Discov. 2019. [Google Scholar] [CrossRef] [PubMed]
- Dai, B.; Wang, L.W.Y.; Yu, G.; Huang, X. Single-Cell Nanometric Coating Towards Whole-Cell-Based Biodevices and Biosensors. Chem. Sel. 2018, 3, 7208–7221. [Google Scholar] [CrossRef]
- Wang, H.; Gunsalus, R.P. Coordinate regulation of the Escherichia coli formate dehydrogenase fdnGHI and fdhF genes in response to nitrate, nitrite, and formate: Roles for NarL and NarP. J. Bacteriol. 2003, 185, 5076–5085. [Google Scholar] [CrossRef] [PubMed]
- Osman, D.; Piergentili, C.; Chen, J.; Sayer, L.N.; Uson, I.; Huggins, T.G.; Robinson, N.J.; Pohl, E. The Effectors and Sensory Sites of Formaldehyde-responsive Regulator FrmR and Metal-sensing Variant. J. Biol. Chem. 2016, 291, 19502–19516. [Google Scholar] [CrossRef] [PubMed]
- Libis, V.; Delepine, B.; Faulon, J.-L. Expanding biosensing abilities through computer-aided design of metabolic pathways. ACS Synth. Biol. 2016, 5, 1076–1085. [Google Scholar] [CrossRef]
- Denby, K.J.; Iwig, J.; Bisson, C.; Westwood, J.; Rolfe, M.D.; Sedelnikova, S.E.; Higgins, K.; Maroney, M.J.; Baker, P.J.; Chivers, P.T.; et al. The mechanism of a formaldehyde-sensing transcriptional regulator. Sci. Rep. 2016, 6, 38879. [Google Scholar] [CrossRef] [Green Version]
- Ganesh, I.; Vidhya, S.; Eom, G.T.; Hong, S.H. Construction of Methanol-Sensing Escherichia coli by the Introduction of a Paracoccus denitrificans MxaY-Based Chimeric Two-Component System. J. Microbiol. Biotechnol. 2017, 27, 1106–1111. [Google Scholar]
- Rohlhill, J.; Sandoval, N.R.; Papoutsakis, E.T. Sort-seq approach to engineering a formaldehyde-inducible promoter for dynamically regulated Escherichia coli growth on methanol. ACS Synth. Biol. 2017, 6, 1584–1595. [Google Scholar] [CrossRef] [PubMed]
- Woolston, B.M.; Roth, T.; Kohale, I.; Liu, D.R.; Stephanopoulos, G. Development of a formaldehyde biosensor with application to synthetic methylotrophy. Biotechnol. Bioeng. 2018, 115, 206–215. [Google Scholar] [CrossRef]
- Hanson, R.S.; Hanson, T.E. Methanotrophic bacteria. Microbiol. Rev. 1996, 60, 439–471. [Google Scholar]
- Netto, C.G.C.M.; Andradea, L.H.; Toma, H.E. Association of Pseudomonas putida formaldehyde dehydrogenase with superparamagnetic nanoparticles: An effective way of improving the enzyme stability, performance and recycling. New J. Chem. 2015, 39, 2162–2167. [Google Scholar] [CrossRef]
- Oppenheimer, N.J.; Henehan, G.T.; Huete-Perez, J.A.; Ito, K. P. putida formaldehyde dehydrogenase. An alcohol dehydrogenase masquerading as an aldehyde dehydrogenase. Adv. Exp. Med. Biol. 1997, 414, 417–423. [Google Scholar]
- de Vries, G.E.; Arfman, N.; Terpstra, P.; Dijkhuizen, L. Cloning, expression, and sequence analysis of the Bacillus methanolicus C1 methanol dehydrogenase gene. J. Bacteriol. 1992, 174, 5346–5353. [Google Scholar] [CrossRef]
- Cheng, F.; Tang, X.L.; Kardashliev, T. Transcription factor-based biosensors in high-throughput screening: Advances and applications. Biotechnol. J. 2018, 13, e1700648. [Google Scholar] [CrossRef]
- Meyer, A.J.; Segall-Shapiro, T.H.; Glassey, E.; Zhang, J.; Voigt, C.A. Escherichia coli “Marionette” strains with 12 highly optimized small-molecule sensors. Nat. Chem. Biol. 2019, 15, 196–204. [Google Scholar] [CrossRef]
- Loeschcke, A.; Thies, S. Pseudomonas putida-a versatile host for the production of natural products. Appl. Microbiol. Biotechnol. 2015, 99, 6197–6214. [Google Scholar] [CrossRef]
- Sheehan, M.C.; Bailey, C.J.; Dowds, B.C.; McConnell, D.J. A new alcohol dehydrogenase, reactive towards methanol, from Bacillus stearothermophilus. Biochem. J. 1988, 252, 661–666. [Google Scholar] [CrossRef]
- Vick, J.E.; Johnson, E.T.; Choudhary, S.; Bloch, S.E.; Lopez-Gallego, F.; Srivastava, P.; Tikh, I.B.; Wawrzyn, G.T.; Schmidt-Dannert, C. Optimized compatible set of BioBrick vectors for metabolic pathway engineering. Appl. Microbiol. Biotechnol. 2011, 92, 1275–1286. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Description | Reference |
|---|---|---|
| Strain | ||
| E. coli DH5α | Cloning and protein expression | NEB |
| Plasmid | ||
| pFA-GESS | AmpR, PfhlA-FhlR-PfdhF-sfgfp | This work |
| pFrm-GESS | AmpR, Pfrm-frmR-sfgfp | This work |
| pMeOH-GESS | AmpR, Pfrm-frmR-sfgfp, Ptrc-Bsmdh | This work |
| pET-trc | KanR, trc promoter | This work |
| pET-Bmmdh | KanR, Ptrc-Bmmdh | This work |
| pET-Faldh | KanR, Ptrc-Faldh | This work |
| pFA-Frm-GESS | AmpR, FA-GESS, Frm-GESS module | This work |
| pAC-Bsmdh-Faldh | CmR, PBAD-Bsmdh, Prham-Faldh | This work |
| Name | Minimal Detection Concentration (mM) | Operational Range (mM) | Fold-Change |
|---|---|---|---|
| FA-GESS | 1.0 | 1.0–250 | 7 |
| Frm-GESS | 1.0 × 10−3 | 1.0–50 × 10−3 | 12 |
| MeOH-GESS | 25 | 5–400 | 5 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, J.-Y.; Sung, B.H.; Oh, S.-H.; Kwon, K.K.; Lee, H.; Kim, H.; Lee, D.-H.; Yeom, S.-J.; Lee, S.-G. C1 Compound Biosensors: Design, Functional Study, and Applications. Int. J. Mol. Sci. 2019, 20, 2253. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20092253
Lee J-Y, Sung BH, Oh S-H, Kwon KK, Lee H, Kim H, Lee D-H, Yeom S-J, Lee S-G. C1 Compound Biosensors: Design, Functional Study, and Applications. International Journal of Molecular Sciences. 2019; 20(9):2253. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20092253
Chicago/Turabian StyleLee, Jin-Young, Bong Hyun Sung, So-Hyung Oh, Kil Koang Kwon, Hyewon Lee, Haseong Kim, Dae-Hee Lee, Soo-Jin Yeom, and Seung-Goo Lee. 2019. "C1 Compound Biosensors: Design, Functional Study, and Applications" International Journal of Molecular Sciences 20, no. 9: 2253. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20092253