An Overview of Hazardous Impacts of Soil Salinity in Crops, Tolerance Mechanisms, and Amelioration through Selenium Supplementation

 ,
, 
 ,
, 
 and
and 
Abstract
:1. Introduction
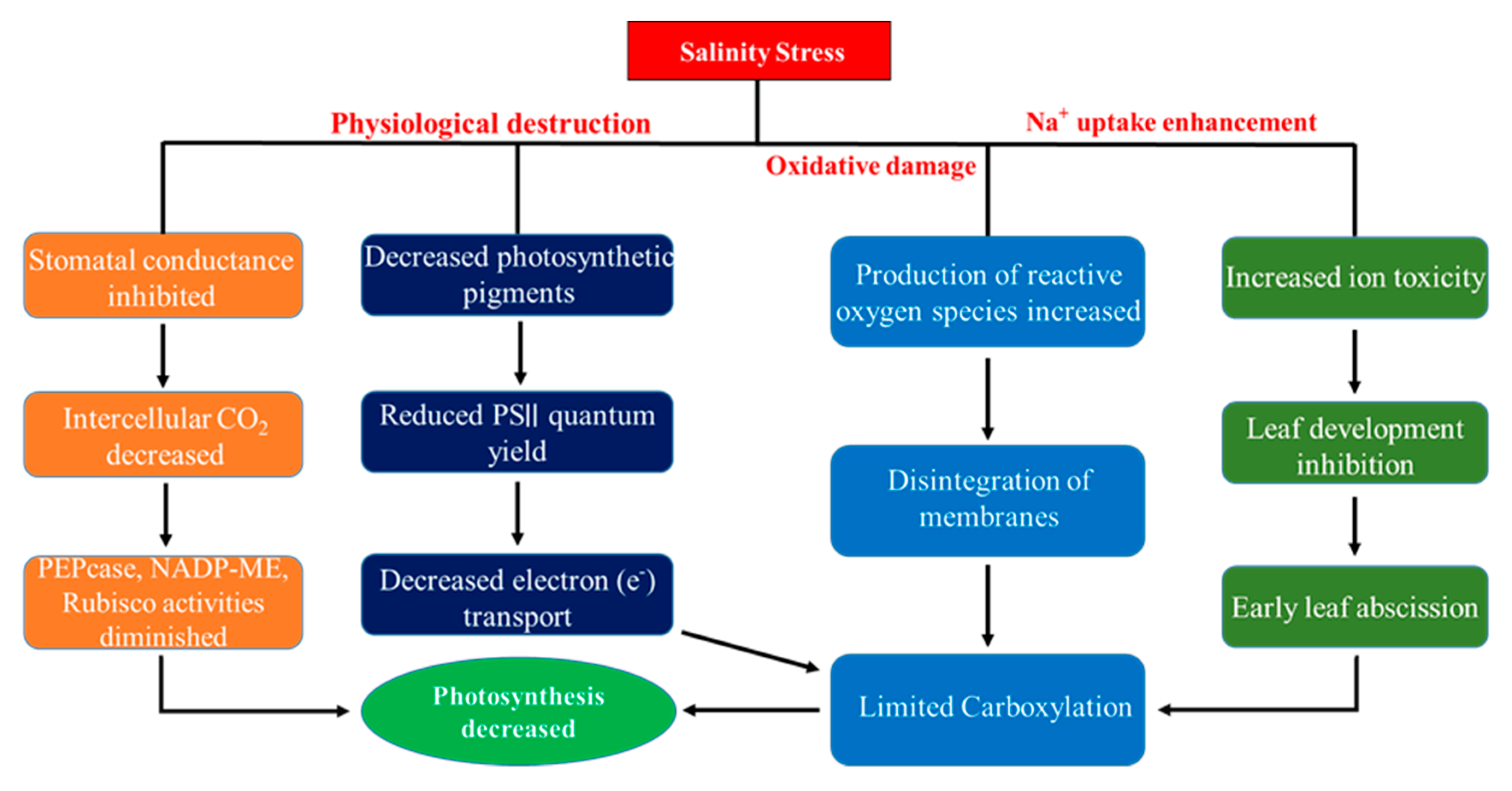
2. Hazardous Impacts of Soil Salinity in Crops
2.1. Impacts of Salinity on Plant Agronomic Traits
2.2. Impacts of Salinity on Physiological Traits
2.3. Impacts of Salinity on Enzymatic and Non-Enzymatic Antioxidants
3. Salinity Tolerance Mechanisms Adopted by Crop Plants
4. Role of Selenium under Abiotic Stresses
4.1. Selenium Speciation and Mobility in Soil
4.2. Selenium Uptake and Mobility within the Plants
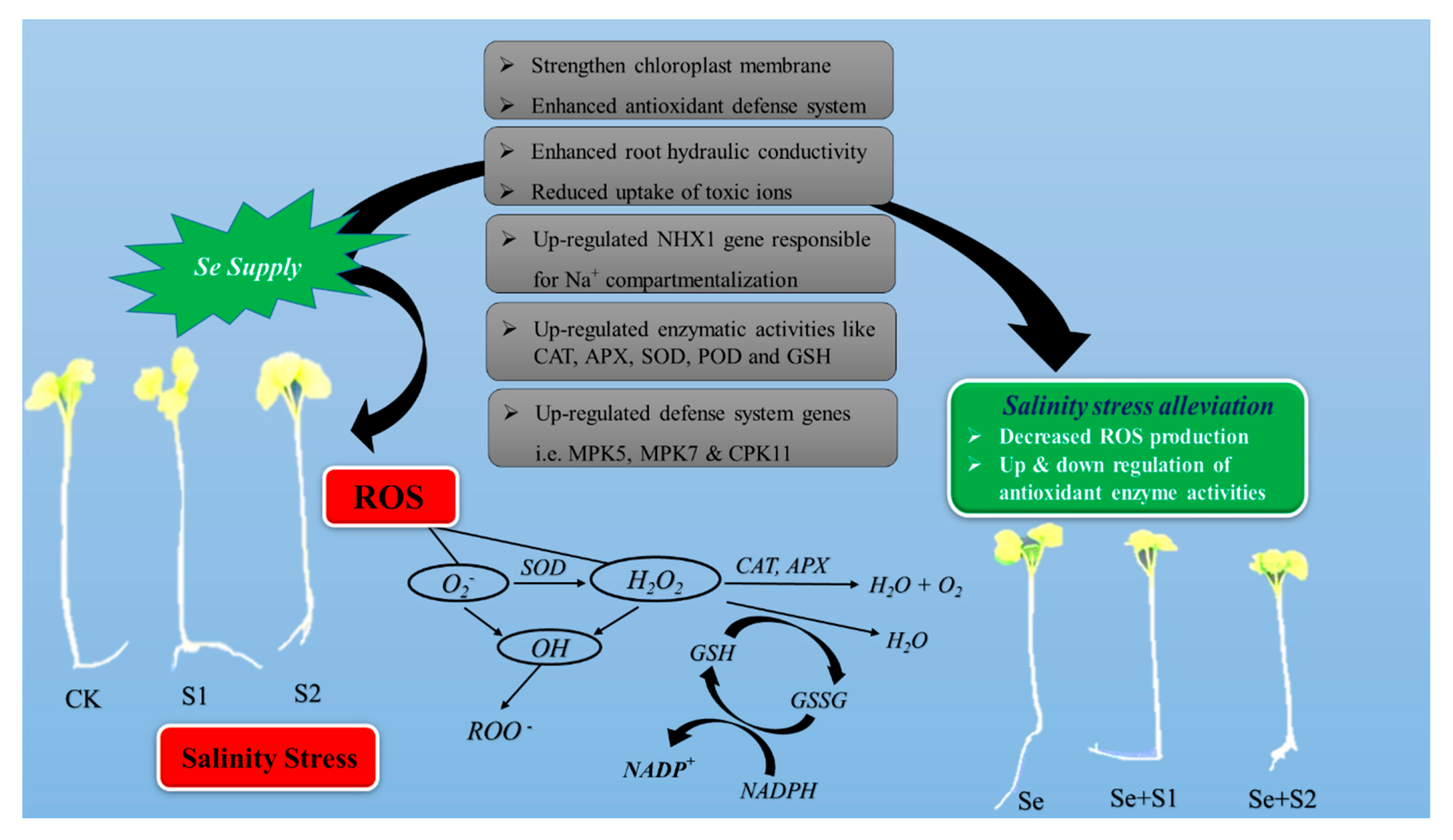
5. Selenium-Mediated Alleviation of Salinity Stress in Plants
5.1. Improvement in Agronomic Traits
5.2. Se-Mediated Improvement in Physiological Attributes
5.3. Se-Mediated Improvement in the Alleviation of ROS Effects
5.4. Se-Mediated Improvement in the Upregulation of Enzymatic and Non-Enzymatic Antioxidants
5.5. Se-Mediated Gene Expression Modifications under Salinity Stress
6. Conclusions and Future Perspectives
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| Se-NPs | Selenium-nanoparticles |
| GSH | Reduced glutathione |
| GSSG | Oxidized glutathione |
| DHAR | Dehydroascorbate reductase |
| MDHAR | Monodehydroascorbate reductase |
| GST | Glutathione S-transferase |
| GR | Glutathione reductase |
| POX | peroxidase |
| PAL | Activity of phenylalanine ammonia-lyase |
| GSH-Px | Glutathione peroxidase |
| CAT | Catalase activity |
| APX | Ascorbate peroxidase activity |
| SOD | Superoxide dismutase activity |
| POD | Peroxidase activity |
| GPX | Glutathione peroxidase activity |
| MDAR | Monodehydroascorbate reductase activity |
| RWC | Relative water contents |
| TBARS | Thiobarbituric acid reactive substances |
| NPQ | Non-photochemical quenching |
| MDA | Malondialdehyde |
| RuBPCase | Ribulose-1,5-bisphosphate-carboxylase/oxygenase content |
| SPAD | Chlorophyll content in leaves |
| H2O2 | Hydrogen peroxide |
| ATP | Adenosine triphosphate |
| NADP+ | Nicotinamide adenine dinucleotide phosphate |
| MAPK | Mitogen activated protein kinase gene |
| CPK | Calcium dependent protein kinase gen |
| NADK2 | NAD kinase2 gene |
| ALMT | Aluminum-activated malate transporters |
| SULTRs | Sulfate transporters |
| γ-GK | γ-Glutamyl kinase |
| NHX | Sodium/hydrogen (Na+/H+) exchanger gene |
| PSII | Photosystem II |
| NPK | Nitrogen, phosphorous, and potassium |
References
- Gontia-Mishra, I.; Sasidharan, S.; Tiwari, S. Recent developments in use of 1-aminocyclopropane-1-carboxylate (ACC) deaminase for conferring tolerance to biotic and abiotic stress. Biotechnol. Lett. 2014, 36, 889–898. [Google Scholar] [CrossRef] [PubMed]
- Abbasi, G.H.; Akhtar, J.; Ahmad, R.; Jamil, M.; Anwar-Ul-Haq, M.; Ali, S.; Ijaz, M. Potassium application mitigates salt stress differentially at different growth stages in tolerant and sensitive maize hybrids. Plant Growth Regul. 2015, 76, 111–125. [Google Scholar] [CrossRef]
- Wallender, W.W.; Tanji, K.K. Agricultural Salinity Assessment and Management; American Society of Civil Engineers: Reston, VA, USA, 2011. [Google Scholar]
- Munns, R.; Tester, M. Mechanisms of Salinity Tolerance. Ann. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdel Latef, A.A. Changes of antioxidative enzymes in salinity tolerance among different wheat cultivars. Cereal Res. Commun. 2010, 38, 43–55. [Google Scholar] [CrossRef]
- Shabala, S.; Cuin, T.A. Potassium transport and plant salt tolerance. Physiol. Plant. 2008, 133, 651–669. [Google Scholar] [CrossRef]
- Ruiz-Lozano, J.M.; Collados, C.; Barea, J.M.; Azcón, R. Arbuscular mycorrhizal symbiosis can alleviate drought-induced nodule senescence in soybean plants. New Phytol. 2001, 151, 493–502. [Google Scholar] [CrossRef]
- Munns, R. Comparative physiology of salt and water stress. Plant Cell Environ. 2002, 25, 239–250. [Google Scholar] [CrossRef]
- Wang, W.; Vinocur, B.; Altman, A. Plant responses to drought, salinity and extreme temperatures: Towards genetic engineering for stress tolerance. Planta 2003, 218, 1–14. [Google Scholar] [CrossRef]
- Sheng, M.; Tang, M.; Chen, H.; Yang, B.; Zhang, F.; Huang, Y. Influence of arbuscular mycorrhizae on photosynthesis and water status of maize plants under salt stress. Mycorrhiza 2008, 18, 287–296. [Google Scholar] [CrossRef]
- Flowers, T.J.; Galal, H.K.; Bromham, L. Evolution of halophytes: Multiple origins of salt tolerance in land plants. Funct. Plant Biol. 2010, 37, 604–612. [Google Scholar] [CrossRef]
- Nabati, J.; Kafi, M.; Nezami, A.; Moghaddam, P.R.; Ali, M.; Mehrjerdi, M.Z. Effect of salinity on biomass production and activities of some key enzymatic antioxidants in Kochia (Kochia scoparia). Pak. J. Bot. 2011, 43, 539–548. [Google Scholar]
- Ahanger, M.A.; Agarwal, R.M. Salinity stress induced alterations in antioxidant metabolism and nitrogen assimilation in wheat (Triticum aestivum L) as influenced by potassium supplementation. Plant Physiol. Biochem. 2017, 115, 449–460. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.; Zheng, Q.; Liu, Z.; Xu, W.; Liu, L.; Zhao, G.; Long, X. Overexpression of Arabidopsis thaliana Na+/H+ antiporter gene enhanced salt resistance in transgenic poplar (Populus × euramericana “Neva”). Trees 2012, 26, 685–694. [Google Scholar] [CrossRef]
- Mittal, S.; Kumari, N.; Sharma, V. Differential response of salt stress on Brassica juncea: Photosynthetic performance, pigment, proline, D1 and antioxidant enzymes. Plant Physiol. Biochem. 2012, 54, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Gengmao, Z.; Shihui, L.; Xing, S.; Yizhou, W.; Zipan, C. The role of silicon in physiology of the medicinal plant (Lonicera japonica L.) under salt stress. Sci. Rep. 2015, 5, 12696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quintero, J.M.; Fournier, J.M.; Benlloch, M. Na+ accumulation in shoot is related to water transport in K+-starved sunflower plants but not in plants with a normal K+ status. J. Plant Physiol. 2007, 164, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Noctor, G.; Foyer, C.H. Ascorbate and Glutathione: Keeping Active Oxygen Under Control. Ann. Rev. Plant Physiol. Plant Mol. Biol. 1998, 49, 249–279. [Google Scholar] [CrossRef]
- Pitzschke, A.; Forzani, C.; Hirt, H. Reactive oxygen species signaling in plants. Antioxid. Redox Signal. 2006, 8, 1757–1764. [Google Scholar] [CrossRef]
- Farooq, M.; Hussain, M.; Wakeel, A.; Siddique, K.H. Salt stress in maize: Effects, resistance mechanisms, and management. A Rev. Agron. Sustain. Dev. 2015, 35, 461–481. [Google Scholar] [CrossRef] [Green Version]
- Aghighi Shahverdi, M.; Omidi, H.; Tabatabaei, S.J. Plant growth and steviol glycosides as affected by foliar application of selenium, boron, and iron under NaCl stress in Stevia rebaudiana Bertoni. Ind. Crops Prod. 2018, 125, 408–415. [Google Scholar] [CrossRef]
- Iqbal, M.; Hussain, I.; Liaqat, H.; Ashraf, M.A.; Rasheed, R.; Rehman, A.U. Exogenously applied selenium reduces oxidative stress and induces heat tolerance in spring wheat. Plant Physiol. Biochem. 2015, 94, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Sieprawska, A.; Kornaś, A.; Filek, M. Involvement of selenium in protective mechanisms of plants under environmental stress conditions—Review. Acta Biol. Crac. Ser. Bot. 2015, 57, 9–20. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Hossain, M.A.; Fujita, M. Selenium-induced up-regulation of the antioxidant defense and methylglyoxal detoxification system reduces salinity-induced damage in rapeseed seedlings. Biol. Trace Elem. Res. 2011, 143, 1704–1721. [Google Scholar] [CrossRef] [PubMed]
- Versini, A.; Di Tullo, P.; Aubry, E.; Bueno, M.; Thiry, Y.; Pannier, F.; Castrec-Rouelle, M. Influence of Se concentrations and species in hydroponic cultures on Se uptake, translocation and assimilation in non-accumulator ryegrass. Plant Physiol. Biochem. 2016, 108, 372–380. [Google Scholar] [CrossRef]
- Babalar, M.; Mohebbi, S.; Zamani, Z.; Askari, M.A. Effect of foliar application with sodium selenate on selenium biofortification and fruit quality maintenance of Starking Delicious apple during storage. J. Sci. Food Agric. 2019, 99, 5149–5156. [Google Scholar] [CrossRef]
- Subramanyam, K.; Du Laing, G.; Van Damme, E.J.M. Sodium selenate treatment using a combination of seed priming and foliar spray alleviates salinity stress in rice. Front. Plant Sci. 2019, 10, 116. [Google Scholar] [CrossRef] [Green Version]
- Elkelish, A.A.; Soliman, M.H.; Alhaithloul, H.A.; El-Esawi, M.A. Selenium protects wheat seedlings against salt stress-mediated oxidative damage by up-regulating antioxidants and osmolytes metabolism. Plant Physiol. Biochem. 2019, 137, 144–153. [Google Scholar] [CrossRef]
- Jiang, C.; Zu, C.; Lu, D.; Zheng, Q.; Shen, J.; Wang, H.; Li, D. Effect of exogenous selenium supply on photosynthesis, Na+ accumulation and antioxidative capacity of maize (Zea mays L.) under salinity stress. Sci. Rep. 2017, 7, 42039. [Google Scholar] [CrossRef] [Green Version]
- Diao, M.; Ma, L.; Wang, J.; Cui, J.; Fu, A.; Liu, H.Y. Selenium Promotes the Growth and Photosynthesis of Tomato Seedlings Under Salt Stress by Enhancing Chloroplast Antioxidant Defense System. J. Plant Growth Regul. 2014, 33, 671–682. [Google Scholar] [CrossRef]
- Kusvuran, S.; Kiran, S.; Ellialtioglu, S.S. Antioxidant Enzyme Activities and Abiotic Stress Tolerance Relationship in Vegetable Crops. In Abiotic and Biotic Stress in Plants–Recent Advances and Future Perspectives; IntechOpen: London, UK, 2016. [Google Scholar]
- Munns, R. Physiological processes limiting plant growth in saline soils: Some dogmas and hypotheses. Plant Cell Environ. 1993, 16, 15–24. [Google Scholar] [CrossRef]
- Evelin, H.; Kapoor, R.; Giri, B. Arbuscular mycorrhizal fungi in alleviation of salt stress: A review. Ann. Bot. 2009, 104, 1263–1280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hajiboland, R.; Aliasgharzad, N.; Laiegh, S.F.; Poschenrieder, C. Colonization with arbuscular mycorrhizal fungi improves salinity tolerance of tomato (Solanum lycopersicum L.) plants. Plant Soil 2010, 331, 313–327. [Google Scholar] [CrossRef]
- Hussain, M.; Park, H.W.; Farooq, M.; Jabran, K.; Lee, D.J. Morphological and physiological basis of salt resistance in different rice genotypes. Int. J. Agric. Biol. 2013, 15, 113–118. [Google Scholar]
- Hachicha, M.; Kahlaoui, B.; Khamassi, N.; Misle, E.; Jouzdan, O. Effect of electromagnetic treatment of saline water on soil and crops. J. Saudi Soc. Agric. Sci. 2018, 17, 154–162. [Google Scholar] [CrossRef]
- Hamada, A.M. Alleviation of the adverse effects of NaCl on germination, seedling, growth and metabolic activities of maize plants by calcium salts. Bull. Fac. Sci. Assiut Univ. 1995, 24, 211–220. [Google Scholar]
- Lee, D.G.; Park, K.W.; An, J.Y.; Sohn, Y.G.; Ha, J.K.; Kim, H.Y.; Bae, D.W.; Lee, K.H.; Kang, N.J.; Lee, B.H.; et al. Proteomics analysis of salt-induced leaf proteins in two rice germplasms with different salt sensitivity. Can. J. Plant Sci. 2011, 91, 337–349. [Google Scholar] [CrossRef]
- Ibrahim, E.A. Seed priming to alleviate salinity stress in germinating seeds. J. Plant Physiol. 2016, 192, 38–46. [Google Scholar] [CrossRef]
- Memon, S.A.; Hou, X.; Wang, L.J. Morphological analysis of salt stress response of pak choi. Electron. J. Environ. Agric. Food Chem. 2010, 9, 1. [Google Scholar]
- Hanafy, M.S.; El-Banna, A.; Schumacher, H.M.; Jacobsen, H.J.; Hassan, F.S. Enhanced tolerance to drought and salt stresses in transgenic faba bean (Vicia faba L.) plants by heterologous expression of the PR10a gene from potato. Plant Cell Rep. 2013, 32, 663–674. [Google Scholar] [CrossRef]
- Kapoor, K.; Srivastava, A. Assessment of Salinity Tolerance of Vigna mungo Var. Pu-19 Using ex vitro and in vitro Methods. Asian J. Biotechnol. 2010, 2, 73–85. [Google Scholar] [CrossRef] [Green Version]
- Anwar-ul-Haq, M.; Akram, S.; Akhtar, J.; Saqib, M.; Saqib, Z.A.; Abbasi, G.H.; Jan, M. Morpho-physiological characterization of sunflower genotypes (Helianthus annuus L.) under saline condition. Pak. J. Agric. Sci. 2013, 50, 49–54. [Google Scholar]
- Mallahi, T.; Saharkhiz, M.J.; Javanmardi, J. Salicylic acid changes morpho-physiological attributes of feverfew (Tanacetum parthenium L.) under salinity stress. Acta Ecol. Sin. 2018, 38, 351–355. [Google Scholar] [CrossRef]
- Qados, A.M.S.A. Effect of salt stress on plant growth and metabolism of bean plant Vicia faba (L.). J. Saudi Soc. Agric. Sci. 2011, 10, 7–15. [Google Scholar]
- Hajiboland, R.; Norouzi, F.; Poschenrieder, C. Growth, physiological, biochemical and ionic responses of pistachio seedlings to mild and high salinity. Trees Struct. Funct. 2014, 28, 1065–1078. [Google Scholar] [CrossRef]
- Ahmadi, F.I.; Karimi, K.; Struik, P.C. Effect of exogenous application of methyl jasmonate on physiological and biochemical characteristics of Brassica napus L. cv. Talaye under salinity stress. S. Afr. J. Bot. 2018, 115, 5–11. [Google Scholar] [CrossRef]
- Shi, Q.; Ding, F.; Wang, X.; Wei, M. Exogenous nitric oxide protects cucumber roots against oxidative stress induced by salt stress. Plant Physiol. Biochem. 2007, 45, 542–550. [Google Scholar] [CrossRef] [PubMed]
- Netondo, G.W.; Onyango, J.C.; Beck, E. Sorghum and salinity: II. Gas exchange and chlorophyll fluorescence of sorghum under salt stress. Crop Sci. 2004, 44, 806. [Google Scholar] [CrossRef]
- Sapre, S.; Gontia-Mishra, I.; Tiwari, S. Klebsiella sp. confers enhanced tolerance to salinity and plant growth promotion in oat seedlings (Avena sativa). Microbiol. Res. 2018, 206, 25–32. [Google Scholar] [CrossRef]
- Rozeff, N. Sugarcane and salinity—A review paper. Sugar Cane 1995. [Google Scholar]
- Cantabella, D.; Piqueras, A.; Acosta-Motos, J.R.; Bernal-Vicente, A.; Hernández, J.A.; Díaz-Vivancos, P. Salt-tolerance mechanisms induced in Stevia rebaudiana Bertoni: Effects on mineral nutrition, antioxidative metabolism and steviol glycoside content. Plant Physiol. Biochem. 2017, 115, 484–496. [Google Scholar] [CrossRef]
- Khan, M.I.R.; Asgher, M.; Khan, N.A. Alleviation of salt-induced photosynthesis and growth inhibition by salicylic acid involves glycinebetaine and ethylene in mungbean (Vigna radiata L.). Plant Physiol. Biochem. 2014, 80, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Munns, R. Genes and salt tolerance: Bringing them together. New Phytol. 2005, 167, 645–663. [Google Scholar] [CrossRef] [PubMed]
- Hussain, S.; Zhang, J.H.; Zhong, C.; Zhu, L.F.; Cao, X.C.; YU, S.M.; Allen Bohr, J.; Hu, J.J.; Jin, Q.Y. Effects of salt stress on rice growth, development characteristics, and the regulating ways: A review. J. Integr. Agric. 2017, 16, 2357–2374. [Google Scholar] [CrossRef] [Green Version]
- Hussain, S.; Zhong, C.; Bai, Z.; Cao, X.; Zhu, L.; Hussain, A.; Zhu, C.; Fahad, S.; James, A.B.; Zhang, J.; et al. Effects of 1-Methylcyclopropene on Rice Growth Characteristics and Superior and Inferior Spikelet Development Under Salt Stress. J. Plant Growth Regul. 2018, 37, 1368–1384. [Google Scholar] [CrossRef]
- Lee, M.H.; Cho, E.J.; Wi, S.G.; Bae, H.; Kim, J.E.; Cho, J.Y.; Lee, S.; Kim, J.H.; Chung, B.Y. Divergences in morphological changes and antioxidant responses in salt-tolerant and salt-sensitive rice seedlings after salt stress. Plant Physiol. Biochem. 2013, 70, 325–335. [Google Scholar] [CrossRef] [PubMed]
- Zahedi, S.M.; Abdelrahman, M.; Hosseini, M.S.; Hoveizeh, N.F.; Tran, L.P. Alleviation of the effect of salinity on growth and yield of strawberry by foliar spray of selenium-nanoparticles. In Environmental Pollution; Elsevier: Amsterdam, The Netherlands, 2019. [Google Scholar]
- Gururani, M.A.; Venkatesh, J.; Tran, L.S.P. Regulation of photosynthesis during abiotic stress-induced photoinhibition. Mol. Plant 2015, 8, 1304–1320. [Google Scholar] [CrossRef] [Green Version]
- James, R.A.; Blake, C.; Byrt, C.S.; Munns, R. Major genes for Na+ exclusion, Nax1 and Nax2 (wheat HKT1;4 and HKT1;5), decrease Na+ accumulation in bread wheat leaves under saline and waterlogged conditions. J. Exp. Bot. 2011, 62, 2939–2947. [Google Scholar] [CrossRef] [Green Version]
- Singam, K.; Juntawong, N.; Cha-Um, S.; Kirdmanee, C. Salt stress induced ion accumulation, ion homeostasis, membrane injury and sugar contents in salt-sensitive rice (Oryza sativa L. spp. indica) roots under isoosmotic conditions. Afr. J. Biotechnol. 2011, 10, 1340–1346. [Google Scholar]
- Hussain, S.; Bai, Z.; Huang, J.; Cao, X.; Zhu, L.; Zhu, C.; Khaskheli, M.A.; Zhong, C.; Jin, Q.; Zhang, J. 1-methylcyclopropene modulates physiological, biochemical, and antioxidant responses of rice to different salt stress levels. Front. Plant Sci. 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Demiral, T.; Türkan, I. Comparative lipid peroxidation, antioxidant defense systems and proline content in roots of two rice cultivars differing in salt tolerance. Environ. Exp. Bot. 2005, 53, 247–257. [Google Scholar] [CrossRef]
- Levitt, J. Responses of Plants to Environmental Stresses. J. Range Manag. 1985, 38, 480. [Google Scholar] [CrossRef]
- Pattanayak, G.K.; Tripathy, B.C. Overexpression of protochlorophyllide oxidoreductase c regulates oxidative stress in arabidopsis. PLoS ONE 2011, 6, e26532. [Google Scholar] [CrossRef] [PubMed]
- Santos, M.G.; Ribeiro, R.V.; Machado, E.C.; Pimentel, C. Photosynthetic parameters and leaf water potential of five common bean genotypes under mild water deficit. Biol. Plant. 2009, 53, 229–236. [Google Scholar] [CrossRef]
- Foyer, C.H.; Noctor, G. Redox sensing and signalling associated with reactive oxygen in chloroplasts, peroxisomes and mitochondria. Physiol. Plant. 2003, 119, 355–364. [Google Scholar] [CrossRef] [Green Version]
- Kamran, M.; Malik, Z.; Parveen, A.; Huang, L.; Riaz, M.; Bashir, S.; Mustafa, A.; Abbasi, G.H.; Xue, B.; Ali, U. Ameliorative Effects of Biochar on Rapeseed (Brassica napus L.) Growth and Heavy Metal Immobilization in Soil Irrigated with Untreated Wastewater. In Journal of Plant Growth Regulation; Springer: Berlin, Germany, 2019; pp. 1–16. [Google Scholar]
- Adly, A.A.M. Oxidative stress and disease: An updated review. Res. J. Immunol. 2010, 3, 129–145. [Google Scholar]
- Houot, V.; Etienne, P.; Petitot, A.S.; Barbier, S.; Blein, J.P.; Suty, L. Hydrogen peroxide induces programmed cell death features in cultured tobacco BY-2 cells, in a dose-dependent manner. J. Exp. Bot. 2001, 52, 1721–1730. [Google Scholar]
- Farmer, E.E.; Mueller, M.J. ROS-Mediated Lipid Peroxidation and RES-Activated Signaling. Ann. Rev. Plant Biol. 2013, 64, 429–450. [Google Scholar] [CrossRef]
- Asada, K. Production and Action of Active Oxygen Species in Photosynthetic Tissues. In Causes of Photooxidative Stress and Amelioration of Defense Systems in Plants; CRC press: Boca Raton, FL, USA, 1994; pp. 77–104. ISBN 0-8493-5443-9. [Google Scholar]
- Drobot, L.B.; Samoylenko, A.A.; Vorotnikov, A.V.; Tyurin -Kuzmin, P.A.; Bazalii, A.V.; Kietzmann, T.; Tkachuk, V.A.; Komisarenko, S.V. Reactive oxygen species in signal transduction. Ukrain’skyi Biokhimichnyi Zhurnal 2013, 85, 209–217. [Google Scholar] [CrossRef] [Green Version]
- Ramachandra Reddy, A.; Chaitanya, K.V.; Jutur, P.P.; Sumithra, K. Differential antioxidative responses to water stress among five mulberry (Morus alba L.) cultivars. Environ. Exp. Bot. 2004, 52, 33–42. [Google Scholar] [CrossRef]
- Li, M.; Guo, S.; Xu, Y.; Meng, Q.; Li, G.; Yang, X. Glycine betaine-mediated potentiation of HSP gene expression involves calcium signaling pathways in tobacco exposed to NaCl stress. Physiol. Plant. 2014, 150, 63–75. [Google Scholar] [CrossRef]
- Farhangi-Abriz, S.; Ghassemi-Golezani, K. How can salicylic acid and jasmonic acid mitigate salt toxicity in soybean plants? Ecotoxicol. Environ. Saf. 2018, 154, 1010–1016. [Google Scholar] [CrossRef] [PubMed]
- Kamran, M.; Malik, Z.; Parveen, A.; Zong, Y.; Abbasi, G.H.; Rafiq, M.T.; Shaaban, M.; Mustafa, A.; Bashir, S.; Rafay, M.; et al. Biochar alleviates Cd phytotoxicity by minimizing bioavailability and oxidative stress in pak choi (Brassica chinensis L.) cultivated in Cd-polluted soil. J. Environ. Manag. 2019, 250, 109500. [Google Scholar] [CrossRef] [PubMed]
- Krishnaiah, D.; Sarbatly, R.; Nithyanandam, R. A review of the antioxidant potential of medicinal plant species. Food Bioprod. Process. 2011, 89, 217–233. [Google Scholar] [CrossRef]
- Cheeseman, J.M. The evolution of halophytes, glycophytes and crops, and its implications for food security under saline conditions. New Phytol. 2015, 206, 557–570. [Google Scholar] [CrossRef] [PubMed]
- Mbarki, S.; Sytar, O.; Cerda, A.; Zivcak, M.; Rastogi, A.; He, X.; Zoghlami, A.; Abdelly, C.; Brestic, M. Strategies to mitigate the salt stress effects on photosynthetic apparatus and productivity of crop plants. In Salinity Responses and Tolerance in Plants, Volume 1: Targeting Sensory, Transport and Signaling Mechanisms; Springer: Berlin, Germany, 2018; ISBN 9-7833-1975-6714. [Google Scholar]
- Zhang, H.X.; Blumwald, E. Transgenic salt-tolerant tomato plants accumulate salt in foliage but not in fruit. Nat. Biotechnol. 2001, 19, 765. [Google Scholar] [CrossRef]
- Takahashi, R.; Liu, S.; Takano, T. Isolation and characterization of plasma membrane Na+/H+ antiporter genes from salt-sensitive and salt-tolerant reed plants. J. Plant Physiol. 2009, 166, 301–309. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, F.; Zhang, C.; Zhang, J.; Chen, Y.; Liu, G.; Zhao, Y.; Hao, F.; Zhang, J. A novel VIGS method by agroinoculation of cotton seeds and application for elucidating functions of GhBI-1 in salt-stress response. Plant Cell Rep. 2018, 37, 1091–1100. [Google Scholar] [CrossRef]
- Demidchik, V.; Cuin, T.A.; Svistunenko, D.; Smith, S.J.; Miller, A.J.; Shabala, S.; Sokolik, A.; Yurin, V. Arabidopsis root K+-efflux conductance activated by hydroxyl radicals: Single-channel properties, genetic basis and involvement in stress-induced cell death. J. Cell Sci. 2010, 123, 1468–1479. [Google Scholar] [CrossRef] [Green Version]
- Isla, R.; Aragüés, R. Yield and plant ion concentrations in maize (Zea mays L.) subject to diurnal and nocturnal saline sprinkler irrigations. Field Crops Res. 2010, 123, 1468–1479. [Google Scholar] [CrossRef] [Green Version]
- Wakeel, A.; Farooq, M.; Qadir, M.; Schubert, S. Potassium substitution by sodium in plants. Crit. Rev. Plant Sci. 2011, 30, 401–413. [Google Scholar] [CrossRef]
- Bhosale, R.; Boudolf, V.; Cuevas, F.; Lu, R.; Eekhout, T.; Hu, Z.; Van Isterdael, G.; Lambert, G.M.; Xu, F.; Nowack, M.K.; et al. A spatiotemporal dna endoploidy map of the arabidopsis root reveals roles for the endocycle in root development and stress adaptation. Plant Cell 2018, 30, 2330–2351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lv, S.; Yu, D.; Sun, Q.; Jiang, J. Activation of gibberellin 20-oxidase 2 undermines auxin-dependent root and root hair growth in NaCl-stressed Arabidopsis seedlings. Plant Growth Regul. 2018, 84, 225–236. [Google Scholar] [CrossRef]
- Younis, M.E.; El-Shahaby, O.A.; Nemat Alla, M.M.; El-Bastawisy, Z.M. Kinetin alleviates the influence of waterlogging and salinity on growth and affects the production of plant growth regulators in Vigna sinensis and Zea mays. Agronomie 2003, 23, 277–285. [Google Scholar] [CrossRef] [Green Version]
- Javid, M.G.; Sorooshzadeh, A.; Moradi, F.; Sanavy, S.A.M.M.; Allahdadi, I. The role of phytohormones in alleviating salt stress in crop plants. Aust. J. Crop Sci. 2011, 5, 726. [Google Scholar]
- Nawaz, K.; Ashraf, M. Improvement in salt tolerance of maize by exogenous application of glycinebetaine: Growth and water relations. Pak. J. Bot. 2007, 39, 1647–1653. [Google Scholar]
- Sobahan, M.A.; Akter, N.; Ohno, M.; Okuma, E.; Hirai, Y.; Mori, I.C.; Nakamura, Y.; Murata, Y. Effects of exogenous proline and glycinebetaine on the salt tolerance of rice cultivars. Biosci. Biotechnol. Biochem. 2012, 76, 1568–1570. [Google Scholar] [CrossRef] [PubMed]
- Athar, H.U.R.; Zafar, Z.U.; Ashraf, M. Glycinebetaine Improved Photosynthesis in Canola under Salt Stress: Evaluation of Chlorophyll Fluorescence Parameters as Potential Indicators. J. Agron. Crop Sci. 2015, 201, 428–442. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Fujita, M. Plant response to salt stress and role of exogenous protectants to mitigate salt-induced damages. In Ecophysiology and Responses of Plants under Salt Stress; Springer: Berlin, Germany, 2013; ISBN 9-78146-144-7474. [Google Scholar]
- Xiang, L.; Liu, C.; Luo, J.; He, L.; Deng, Y.; Yuan, J.; Wu, C.; Cai, Y. A tuber mustard AP2/ERF transcription factor gene, BjABR1, functioning in abscisic acid and abiotic stress responses, and evolutionary trajectory of the ABR1 homologous genes in Brassica species. PeerJ 2018, 6, e6071. [Google Scholar] [CrossRef] [Green Version]
- Long, L.; Yang, W.W.; Liao, P.; Guo, Y.W.; Kumar, A.; Gao, W. Transcriptome analysis reveals differentially expressed ERF transcription factors associated with salt response in cotton. Plant Sci. 2019, 281, 72–81. [Google Scholar] [CrossRef]
- Uddin, M.N.; Hanstein, S.; Leubner, R.; Schubert, S. Leaf Cell-Wall Components as Influenced in the First Phase of Salt Stress in Three Maize (Zea mays L.) Hybrids Differing in Salt Resistance. J. Agron. Crop Sci. 2013, 199, 405–415. [Google Scholar] [CrossRef]
- Menezes-Benavente, L.; Kernodle, S.P.; Margis-Pinheiro, M.; Scandalios, J.G. Salt-induced antioxidant metabolism defenses in maize (Zea mays L.) seedlings. Redox Rep. 2004, 9, 29–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez-Kessler, M.; Alpuche-Solís, A.G.; Ruiz, O.A.; Jiménez-Bremont, J.F. Effect of salt stress on the regulation of maize (Zea mays L.) genes involved in polyamine biosynthesis. Plant Growth Regul. 2006, 48, 175–185. [Google Scholar] [CrossRef]
- Hawrylak-Nowak, B. Selenite is more efficient than selenate in alleviation of salt stress in lettuce plants. Acta Biol. Crac. Ser. Bot. 2015, 57, 49–54. [Google Scholar] [CrossRef]
- Mehdi, Y.; Hornick, J.L.; Istasse, L.; Dufrasne, I. Selenium in the environment, metabolism and involvement in body functions. Molecules 2013, 18, 3292–3311. [Google Scholar] [CrossRef] [Green Version]
- Shahid, M.; Niazi, N.K.; Khalid, S.; Murtaza, B.; Bibi, I.; Rashid, M.I. A critical review of selenium biogeochemical behavior in soil-plant system with an inference to human health. Environ. Pollut. 2018, 234, 915–934. [Google Scholar]
- Galeas, M.L.; Zhang, L.H.; Freeman, J.L.; Wegner, M.; Pilon-Smits, E.A.H. Seasonal fluctuations of selenium and sulfur accumulation in selenium hyperaccumulators and related nonaccumulators. New Phytol. 2007, 173, 517–525. [Google Scholar] [CrossRef]
- Hartikainen, H. Biogeochemistry of selenium and its impact on food chain quality and human health. J. Trace Elem. Med. Biol. 2005, 18, 309–318. [Google Scholar] [CrossRef]
- White, P.J. Selenium accumulation by plants. Ann. Bot. 2015, 117, 217–235. [Google Scholar] [CrossRef] [Green Version]
- Cartes, P.; Gianfreda, L.; Paredes, C.; Mora, M.L. Selenium uptake and its antioxidant role in ryegrass cultivars as affected by selenite seed pelletization. J. Soil Sci. Plant Nutr. 2011, 11, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Nakamaru, Y.M.; Altansuvd, J. Speciation and bioavailability of selenium and antimony in non-flooded and wetland soils: A review. Chemosphere 2014, 111, 366–371. [Google Scholar] [CrossRef]
- Kikkert, J.; Berkelaar, E. Plant uptake and translocation of inorganic and organic forms of selenium. Arch. Environ. Contam. Toxicol. 2013, 65, 458–465. [Google Scholar] [CrossRef] [PubMed]
- Bruggeman, C.; Maes, A.; Vancluysen, J. The interaction of dissolved Boom Clay and Gorleben humic substances with selenium oxyanions (selenite and selenate). Appl. Geochem. 2007, 22, 1371–1379. [Google Scholar] [CrossRef]
- Shand, C.A.; Eriksson, J.; Dahlin, A.S.; Lumsdon, D.G. Selenium concentrations in national inventory soils from Scotland and Sweden and their relationship with geochemical factors. J. Geochem. Exp. 2012, 121, 4–14. [Google Scholar] [CrossRef]
- Beckett, G.J.; Arthur, J.R. Selenium and endocrine systems. J. Endocrinol. 2005, 184, 455–465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, J.; Zhu, W.; Wang, W.; Li, R.; Hou, S.; Wang, D.; Yang, L. Selenium in soil and endemic diseases in China. Sci. Total Environ. 2002, 284, 227–235. [Google Scholar] [CrossRef]
- Song, Z.; Shao, H.; Huang, H.; Shen, Y.; Wang, L.; Wu, F.; Han, D.; Song, J.; Jia, H. Overexpression of the phosphate transporter gene OsPT8 improves the Pi and selenium contents in Nicotiana tabacum. Environ. Exp. Bot. 2017, 137, 158–165. [Google Scholar] [CrossRef]
- Wang, P.; Menzies, N.W.; Lombi, E.; McKenna, B.A.; James, S.; Tang, C.; Kopittke, P.M. Synchrotron-based X-ray absorption near-edge spectroscopy imaging for laterally resolved speciation of selenium in fresh roots and leaves of wheat and rice. J. Exp. Bot. 2015, 66, 4795–4806. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Lu, C. Photosynthesis is improved by exogenous glycinebetaine in salt-stressed maize plants. Physiol. Plant. 2005, 124, 343–352. [Google Scholar] [CrossRef]
- Huang, Q.Q.; Wang, Q.; Wan, Y.N.; Yu, Y.; Jiang, R.F.; Li, H.F. Application of X-ray absorption near edge spectroscopy to the study of the effect of sulphur on selenium uptake and assimilation in wheat seedlings. Biol. Plant. 2017, 61, 726–732. [Google Scholar] [CrossRef]
- Malcheska, F.; Ahmad, A.; Batool, S.; Müller, H.M.; Ludwig-Müller, J.; Kreuzwieser, J.; Randewig, D.; Hänsch, R.; Mendel, R.R.; Hell, R.; et al. Drought-enhanced xylem sap sulfate closes stomata by affecting ALMT12 and guard cell ABA synthesis. Plant Physiol. 2017, 174, 798–814. [Google Scholar] [CrossRef] [Green Version]
- Gigolashvili, T.; Kopriva, S. Transporters in plant sulfur metabolism. Front. Plant Sci. 2014, 5, 442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winkel, L.H.E.; Vriens, B.; Jones, G.D.; Schneider, L.S.; Pilon-Smits, E.; Bañuelos, G.S. Selenium cycling across soil-plant-atmosphere interfaces: A critical review. Nutrients 2015, 7, 4199–4239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Cappa, J.J.; Harris, J.P.; Edger, P.P.; Zhou, W.; Pires, J.C.; Adair, M.; Unruh, S.A.; Simmons, M.P.; Schiavon, M.; et al. Transcriptome-wide comparison of selenium hyperaccumulator and nonaccumulator Stanleya species provides new insight into key processes mediating the hyperaccumulation syndrome. Plant Biotechnol. J. 2018, 16, 1582–1594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carey, A.M.; Scheckel, K.G.; Lombi, E.; Newville, M.; Choi, Y.; Norton, G.J.; Price, A.H.; Meharg, A.A. Grain accumulation of selenium species in rice (Oryza sativa L.). Environ. Sci. Technol. 2012, 46, 5557–5564. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, M.A.; Akbar, A.; Parveen, A.; Rasheed, R.; Hussain, I.; Iqbal, M. Phenological application of selenium differentially improves growth, oxidative defense and ion homeostasis in maize under salinity stress. Plant Physiol. Biochem. 2018, 123, 268–280. [Google Scholar] [CrossRef]
- Butt, M.; Ayyub, C.M.; Amjad, M.; Ahmad, R. Proline application enhances growth of chilli by improving physiological and biochemical attributes under salt stress. Pak. J. Agric. Sci. 2016, 53, 43–49. [Google Scholar]
- Hawrylak-Nowak, B. Beneficial effects of exogenous selenium in cucumber seedlings subjected to salt stress. Biol. Trace Elem. Res. 2009, 132, 259–269. [Google Scholar] [CrossRef]
- Habibi, G.; Sarvary, S. The Roles of Selenium in Protecting Lemon Balm against Salt Stress. Iran. J. Plant Physiol. 2015, 5, 1425–1433. [Google Scholar]
- Sattar, A.; Cheema, M.A.; Abbas, T.; Sher, A.; Ijaz, M.; Hussain, M. Separate and combined effects of silicon and selenium on salt tolerance of wheat plants. Russ. J. Plant Physiol. 2017, 64, 341–348. [Google Scholar] [CrossRef]
- Astaneh, R.K.; Bolandnazar, S.; Nahandi, F.Z.; Oustan, S. Effects of selenium on enzymatic changes and productivity of garlic under salinity stress. S. Afr. J. Bot. 2019, 121, 447–455. [Google Scholar] [CrossRef]
- Mozafariyan, M.; Kamelmanesh, M.M.; Hawrylak-Nowak, B. Ameliorative effect of selenium on tomato plants grown under salinity stress. Arch. Agron. Soil Sci. 2016, 62, 1368–1380. [Google Scholar] [CrossRef]
- Astaneh, R.K.; Bolandnazar, S.; Nahandi, F.Z.; Oustan, S. The effects of selenium on some physiological traits and K, Na concentration of garlic (Allium sativum L.) under NaCl stress. Inf. Process. Agric. 2018, 5, 156–161. [Google Scholar] [CrossRef]
- Atarodi, B.; Fotovat, A.; Khorassani, R.; Keshavarz, P.; Hawrylak-Nowak, B. Selenium improves physiological responses and nutrient absorption in wheat (Triticum aestivum L.) grown under salinity. Toxicol. Environ. Chem. 2018, 100, 440–451. [Google Scholar] [CrossRef]
- Bybordi, A.; Saadat, S.; Zargaripour, P. The effect of zeolite, selenium and silicon on qualitative and quantitative traits of onion grown under salinity conditions. Arch. Agron. Soil Sci. 2018, 64, 520–530. [Google Scholar] [CrossRef]
- Moussa, H.R.; Hassen, A.M. Selenium Affects Physiological Responses of Phaseolus vulgaris in Response to Salt Level. Int. J. Veg. Sci. 2018, 24, 236–253. [Google Scholar] [CrossRef]
- Habibi, G. Selenium ameliorates salinity stress in Petroselinum crispum by modulation of photosynthesis and by reducing shoot Na accumulation. Russ. J. Plant Physiol. 2017, 64, 368–374. [Google Scholar] [CrossRef]
- Khalifa, G.S.; Abdelrassoul, M.; Hegazi, A.; Elsherif, M.H. Mitigation of Saline Stress Adverse Effects in Lettuce Plant Using Selenium and Silicon. Middle East J. Agric. Res. 2016, 347–361. [Google Scholar]
- Abul-Soud, M.A.; Abd-Elrahman, S.H. Foliar selenium application to improve the tolerance of eggplant grown under salt stress conditions. Int. J. Plant Soil Sci. 2016, 9, 1–10. [Google Scholar] [CrossRef]
- Hashem, H.A.; Hassanein, R.A.; Bekheta, M.A.; El-Kady, F.A. Protective role of selenium in canola (Brassica napus L.) plant subjected to salt stress. Egypt. J. Exp. Biol. 2013, 9, 199–211. [Google Scholar]
- Selenium Induces Antioxidant Defensive Enzymes and Promotes Tolerance Against Salinity Stress in Cucumber Seedlings (Cucumis sativus). Arab. Univ. J. Agric. Sci. 2010, 18, 65–76.
- Kong, L.; Wang, M.; Bi, D. Selenium modulates the activities of antioxidant enzymes, osmotic homeostasis and promotes the growth of sorrel seedlings under salt stress. Plant Growth Regul. 2005, 45, 155–163. [Google Scholar] [CrossRef]
- Rietz, D.N.; Haynes, R.J. Effects of irrigation-induced salinity and sodicity on soil microbial activity. Soil Biol. Biochem. 2003, 35, 845–854. [Google Scholar] [CrossRef]
- Carillo, P.; Mastrolonardo, G.; Nacca, F.; Fuggi, A. Nitrate reductase in durum wheat seedlings as affected by nitrate nutrition and salinity. Funct. Plant Biol. 2005, 32, 209–219. [Google Scholar] [CrossRef]
- Szabados, L.; Savouré, A. Proline: A multifunctional amino acid. Trends Plant Sci. 2010, 15, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Shahzadi, I.; Iqbal, M.; Rasheed, R.; Arslan Ashraf, M.; Perveen, S.; Hussain, M. Foliar application of selenium increases fertility and grain yield in bread wheat under contrasting water availability regimes. Acta Physiol. Plant. 2017, 39, 173. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, L.; Liu, Y.; Zhang, Q.; Wei, Q.; Zhang, W. Nitric oxide enhances salt tolerance in maize seedlings through increasing activities of proton-pump and Na+/H+ antiport in the tonoplast. Planta 2006, 224, 545–555. [Google Scholar] [CrossRef]
- Ahmad, P.; Sarwat, M.; Bhat, N.A.; Wani, M.R.; Kazi, A.G.; Tran, L.S.P. Alleviation of cadmium toxicity in Brassica juncea L. (Czern. & Coss.) by calcium application involves various physiological and biochemical strategies. PLoS ONE 2015, 10, e0114571. [Google Scholar]
- Shabala, S.; Pottosin, I. Regulation of potassium transport in plants under hostile conditions: Implications for abiotic and biotic stress tolerance. Physiol. Plant. 2014, 151, 257–279. [Google Scholar] [CrossRef]
- Çakir, Ö.; Turgut-Kara, N.; Ari, Ş. Selenium induced selenocysteine methyltransferase gene expression and antioxidant enzyme activities in Astragalus chrysochlorus. Acta Bot. Croat. 2016, 75, 11–16. [Google Scholar] [CrossRef] [Green Version]
- Shekari, F.; Abbasi, A.; Mustafavi, S.H. Effect of silicon and selenium on enzymatic changes and productivity of dill in saline condition. J. Saudi Soc. Agric. Sci. 2017, 16, 367–374. [Google Scholar] [CrossRef] [Green Version]
- Gul, H.; Kinza, S.; Shinwari, Z.K.; Hamayun, M. Effect of selenium on the biochemistry of Zea mays under salt stress. Pak. J. Bot. 2017, 49, 25–32. [Google Scholar]
- Alyemeni, M.N.; Ahanger, M.A.; Wijaya, L.; Alam, P.; Bhardwaj, R.; Ahmad, P. Selenium mitigates cadmium-induced oxidative stress in tomato (Solanum lycopersicum L.) plants by modulating chlorophyll fluorescence, osmolyte accumulation, and antioxidant system. Protoplasma 2018, 255, 459–469. [Google Scholar] [CrossRef] [PubMed]
- Padmaja, K.; Prasad, D.D.K.; Prasad, A.R.K. Effect of selenium on chlorophyll biosynthesis in mung bean seedlings. Phytochemistry 1989, 28, 3321–3324. [Google Scholar] [CrossRef]
- Moulick, D.; Santra, S.C.; Ghosh, D. Seed priming with Se mitigates As-induced phytotoxicity in rice seedlings by enhancing essential micronutrient uptake and translocation and reducing As translocation. Environ. Sci. Pollut. Res. 2018, 25, 26978–26991. [Google Scholar] [CrossRef]
- Carvalho, E.R.; Oliveira, J.A.; Von Pinho, É.V.D.R.; Costa Neto, J. Enzyme activity in soybean seeds produced under foliar application of manganese. Ciência e Agrotecnologia 2014, 38, 317–327. [Google Scholar] [CrossRef] [Green Version]
- Asgher, M.; Per, T.S.; Verma, S.; Pandith, S.A.; Masood, A.; Khan, N.A. Ethylene Supplementation Increases PSII Efficiency and Alleviates Chromium-Inhibited Photosynthesis Through Increased Nitrogen and Sulfur Assimilation in Mustard. J. Plant Growth Regul. 2018, 37, 1300–1317. [Google Scholar] [CrossRef]
- Guo, C.Y.; Wang, X.Z.; Chen, L.; Ma, L.N.; Wang, R.Z. Physiological and biochemical responses to saline-alkaline stress in two halophytic grass species with different photosynthetic pathways. Photosynthetica 2015, 53, 128–135. [Google Scholar] [CrossRef]
- Zong, X.J.; Li, D.P.; Gu, L.K.; Li, D.Q.; Liu, L.X.; Hu, X.L. Abscisic acid and hydrogen peroxide induce a novel maize group C MAP kinase gene, ZmMPK7, which is responsible for the removal of reactive oxygen species. Planta 2009, 229, 485–495. [Google Scholar] [CrossRef]
- Shalaby, T.; Bayoumi, Y.; Alshaal, T.; Elhawat, N.; Sztrik, A.; El-Ramady, H. Selenium fortification induces growth, antioxidant activity, yield and nutritional quality of lettuce in salt-affected soil using foliar and soil applications. Plant Soil 2017, 421, 245–258. [Google Scholar] [CrossRef]
- Filek, M.; Keskinen, R.; Hartikainen, H.; Szarejko, I.; Janiak, A.; Miszalski, Z.; Golda, A. The protective role of selenium in rape seedlings subjected to cadmium stress. J. Plant Physiol. 2008, 165, 833–844. [Google Scholar] [CrossRef]
- Hawrylak-Nowak, B. Comparative effects of selenite and selenate on growth and selenium accumulation in lettuce plants under hydroponic conditions. Plant Growth Regul. 2013, 70, 149–157. [Google Scholar] [CrossRef] [Green Version]
- Garg, N.; Manchanda, G. ROS generation in plants: Boon or bane? Plant Biosyst. 2009, 143, 81–96. [Google Scholar] [CrossRef]
- Ahanger, M.A.; Agarwal, R.M.; Tomar, N.S.; Shrivastava, M. Potassium induces positive changes in nitrogen metabolism and antioxidant system of oat (Avena sativa L. cultivar Kent). J. Plant Interact. 2015, 10, 211–223. [Google Scholar] [CrossRef]
- Saleem, M.H.; Kamran, M.; Zhou, Y.; Parveen, A.; Rehman, M.; Ahmar, S.; Malik, Z.; Mustafa, A.; Anjum, R.M.A.; Wang, B.; et al. Appraising growth, oxidative stress and copper phytoextraction potential of flax (Linum usitatissimum L.) grown in soil differentially spiked with copper. J. Environ. Manag. 2020, 257, 109994. [Google Scholar] [CrossRef] [PubMed]
- Nouet, C.; Motte, P.; Hanikenne, M. Chloroplastic and mitochondrial metal homeostasis. Trends Plant Sci. 2011, 16, 395–404. [Google Scholar] [CrossRef] [PubMed]
- Gao, W.; Xu, F.C.; Guo, D.D.; Zhao, J.R.; Liu, J.; Guo, Y.W.; Singh, P.K.; Ma, X.N.; Long, L.; Botella, J.R.; et al. Calcium-dependent protein kinases in cotton: Insights into early plant responses to salt stress. BMC Plant Biol. 2018, 18, 15. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Wang, J.; Lee, S.; Wen, R. Copper-caused oxidative stress triggers the activation of antioxidant enzymes via ZmMPK3 in maize leaves. PLoS ONE 2018, 13, e0203612. [Google Scholar] [CrossRef]
- Sripinyowanich, S.; Klomsakul, P.; Boonburapong, B.; Bangyeekhun, T.; Asami, T.; Gu, H.; Buaboocha, T.; Chadchawan, S. Exogenous ABA induces salt tolerance in indica rice (Oryza sativa L.): The role of OsP5CS1 and OsP5CR gene expression during salt stress. Environ. Exp. Bot. 2013, 86, 94–105. [Google Scholar] [CrossRef]
- Zhang, A.; Jiang, M.; Zhang, J.; Tan, M.; Hu, X. Mitogen-activated protein kinase is involved in abscisic acid-induced antioxidant defense and acts downstream of reactive oxygen species production in leaves of maize plants. Plant Physiol. 2006, 141, 475–487. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Li, Y.; Miao, W.; Piao, T.; Hao, Y.; Hao, F.S. NADK2 positively modulates abscisic acid-induced stomatal closure by affecting accumulation of H2O2, Ca2+ and nitric oxide in Arabidopsis guard cells. Plant Sci. 2017, 262, 81–90. [Google Scholar] [CrossRef]
- Sun, L.R.; Wang, Y.B.; He, S.B.; Hao, F.S. Mechanisms for Abscisic Acid Inhibition of Primary Root Growth. Plant Signal. Behav. 2018, 13, e1500069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manaf, H.H. Beneficial effects of exogenous selenium, glycine betaine and seaweed extract on salt stressed cowpea plant. Ann. Agric. Sci. 2016, 61, 41–48. [Google Scholar] [CrossRef] [Green Version]
- Kaur, S.; Nayyar, H. Selenium fertilization to salt-stressed mungbean (Vigna radiata L. Wilczek) plants reduces sodium uptake, improves reproductive function, pod set and seed yield. Sci. Hortic. 2015, 197, 304–317. [Google Scholar] [CrossRef]
- Ardebili, N.O.; Saadatmand, S.; Niknam, V.; Khavari-Nejad, R.A. The alleviating effects of selenium and salicylic acid in salinity exposed soybean. Acta Physiol. Plant. 2014, 36, 3199–3205. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Fujita, M. Selenium pretreatment upregulates the antioxidant defense and methylglyoxal detoxification system and confers enhanced tolerance to drought stress in rapeseed seedlings. Biol. Trace Elem. Res. 2011, 143, 1758–1776. [Google Scholar] [CrossRef] [PubMed]
- Feng, R.; Wei, C.; Tu, S. The roles of selenium in protecting plants against abiotic stresses. Environ. Exp. Bot. 2013, 87, 58–68. [Google Scholar] [CrossRef]
- Zhang, A.; Zhang, J.; Ye, N.; Cao, J.; Tan, M.; Zhang, J.; Jiang, M. ZmMPK5 is required for the NADPH oxidase-mediated self-propagation of apoplastic H2O2 in brassinosteroid-induced antioxidant defence in leaves of maize. J. Exp. Bot. 2010, 61, 4399–4411. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Cao, J.; Ni, L.; Zhu, Y.; Zhang, A.; Tan, M.; Jiang, M. ZmCPK11 is involved in abscisic acid-induced antioxidant defence and functions upstream of ZmMPK5 in abscisic acid signalling in maize. J. Exp. Bot. 2013, 64, 871–884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.X.; Hodson, J.N.; Williams, J.P.; Blumwald, E. Engineering salt-tolerant Brassica plants: Characterization of yield and seed oil quality in transgenic plants with increased vacuolar sodium accumulation. Proc. Natl. Acad. Sci. USA 2001, 98, 12832–12836. [Google Scholar] [CrossRef] [Green Version]
- Almeida, D.M.; Gregorio, G.B.; Oliveira, M.M.; Saibo, N.J.M. Five novel transcription factors as potential regulators of OsNHX1 gene expression in a salt tolerant rice genotype. Plant Mol. Biol. 2017, 93, 61–77. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Salinity Stress | Plant Species | Se Dosages | Se Speciation | Experimental Details | Various Protective Effects and Mechanisms of Se in Salinity Stressed Plants | References |
|---|---|---|---|---|---|---|
| 150 mM | Oryza sativa L. | 2, 4, 6, 8, 10, 12 mg·L−1 | Se (VI) | Sand culture | Enhances plant biomass, K+/Na+ ratio, and Se accumulation; reduces malondialdehyde contents (MDA) and H2O2 contents; increases chlorophyll and water contents; causes upregulation of OsNHX1 gene transcript levels | [27] |
| 0, 30, 60, 90 mM | Allium sativum L. | 0, 4, 8, 16 mg·L−1 | Se (VI) | Hydroponic culture | Increases root biomass, bulb diameter, bulb height, and photosynthetic pigments; reduces ion leakage and lipid peroxidation; improves K+ and Na+ contents, chlorophyll index, carotenoids, and water balance | [127,129] |
| 100 mM | Triticum aestivum L. | 5, 10 µM | Se (VI) | Reconstituted soil culture (Peat, compost, sand) | Improves wheat growth; promotes the synthesis of photosynthetic pigments, proline, and sugars; reduces H2O2 contents, Na+ uptake, and Na+/K+ ratio | [28] |
| 10, 30, 60, 90 mM | Stevia rebaudiana Bertoni | 20 g/ha (2 ppt) | Se (IV) | Field experiment | Increases leaf and plant biological yields; enhances rebaudioside-A and stevioside of stevia leaves; improved the accumulation of sweet steviol glycosides contents | [21] |
| 0.12, 0.30, 0.60 S m−1 | Triticum aestivum L. | 0, 0.5, 1, 4 mg·kg−1 | Se (IV) | Pot soil culture | Dramatic decrease in shoot dry biomass; chlorophyll a, chlorophyll b, and carotenoid contents increase at lower Se, while they decrease at higher Se; enhances free proline and Se contents in shoots; | [130] |
| 8 dS m−1 | Allium cepa L. | 0, 0.5, 1 kg·ha−1 | Se (IV) | Field experiment | Increases bulb yield and dry matter; improves water and chlorophyll contents; causes bulb Se and K enrichment; causes a decrease in Na | [131] |
| 0, 100 mM | Phaseolus vulgaris L. | 0, 5, 10 μM | Se (IV) | Pot soil culture | Enhances plant growth and seed yield; promotes membrane stability index, photosynthetic capacity, and RuBPCase activity; reduces (MDA) and electrolyte leakage | [132] |
| 0, 100 mM | Zea mays L. | 0, 1, 5, 25 μM | Se (IV) | Pot vermiculite culture | Enhances growth and biomass; improves gas exchange attributes and the shape of thylakoids by alleviation of damage in the ultrastructure of chloroplasts; upregulates ZmMPK5, ZmMPK7, ZmCPK11, and ZmNHX1 genes transcript levels in roots | [29] |
| 0, 80 mM | Petroselinum crispum L. | 1 mg·L−1 | Se (VI) | Hydroponic culture | Decreases root to shoot transport of Na+; improves photochemical efficiency of photosystem II (PSII) and chlorophyll contents; protects photosynthetic apparatus by upregulation of non-photochemical quenching (NPQ); decreases cell sap Na+ | [133] |
| 3.22 dS m−1 | Lactuca sativa L. | 16, 32 µM | Se (VI) | Field experiment | Improves growth characteristics, yield, and relative water contents; decreases cell membrane permeability and malondialdehyde; enhances chlorophyll, carotenoids, K+/Na+, and total soluble sugars | [134] |
| 0, 25, 50 mM | Lycopersicon esculentum-Mill. | 0, 5, 10 µM | Se (IV) | Hydroponic culture | Enhances growth by improving water balance and cell membrane integrity; increases photosynthetic pigments; decreases proline and phenolics | [128] |
| 0, 30, 60, 120 mM | Solanum melongena L. cv. Baladi | 0, 5, 10, 20, 30 µM | Se (IV) | Bedding sand culture | Increases vegetative growth, yield, nitrogen, phosphorous and potassium NPK contents in leaves and fruits; improves chlorophyll contents (SPAD value) and proline contents; Enhances K+/Na+ ratio | [135] |
| 0, 40 mM | Lactuca sativa L. var. capitate | 0, 2, 6 µM | Se (IV, VI) | Hydroponic culture | Enhances fresh biomass, leaf area, chlorophyll, proline, and carotenoid contents; reduces H2O2 and TBARS; improves shoot ionic concentrations | [100] |
| 0, 40 mM | Melissa officinalis L. | 10 mM | - | Hydroponic culture | Improves growth rate; increases photosynthetic pigments, protein, and total amino acid contents; reduces lipid peroxidation to alleviate membrane damage | [125] |
| 0, 2000, 4000, 6000 mg L−1 | Brassica napus L. | 0, 2.5, 5, 10 mg·L−1 | Se (VI) | Pot clay soil culture | Enhances growth, photosynthetic pigments, canola oil quality; increases soluble sugar, polysaccharides, and total carbohydrates; significantly improves saturated and unsaturated fatty acids composition | [136] |
| 0, 2000 ppm | Cucumis sativus L. cv Zena | 0, 1 ppm | Se (IV) | Pot soil culture | Improves plant biomass; increases reduction of oxygen radicals and osmotic regulation by synthesis of osmoregulatory compound such as proline; reduces malondialdehyde concentration and electrolyte leakage | [137] |
| 0, 50 mM | Cucumis sativus L. | 0, 5, 10, 20 µM | Se (VI) | Hydroponic culture | Induces salt tolerance by protection of cell membranes against lipid peroxidation; improves growth rate, photosynthesis, and proline contents; reduces Cl− contents, while showing no effect on Na+ ions and K+/Na+ ratio | [124] |
| 100 mM | Rumex patientia × R. tianshanicus | 0, 1, 3, 5, 10, 30 µM | Se (IV) | Sand culture | Increases seedling growth; lower Se supply improves total water-soluble sugars, K+, and Na+ concentrations; alleviates integrity of cytoplasmic organelles, plasma and nuclear membranes, root tip cells; makes more legible and increases mitochondrial cristae in leaf mesophyll | [138] |
| Salinity Stress | Plant Species | Se Dosages | Se Speciation | Experimental Details | ↑↓ Antioxidant Activity | % Increase or Decrease | Reference |
|---|---|---|---|---|---|---|---|
| 150 mM | Oryza sativa L. | 2, 4, 6, 8, 10, 12 mg·L−1 | Se (VI) | Sand culture | ↑SOD ↑APX ↑CAT ↑GR ↑GSH-Px | 40.7% 92.7% 82.9% 77.2% 66.1% | [27] |
| 0, 25, 50, 75 mM | Fragaria × ananassa Duch | 0, 10, 20 mg·L−1 | Se-NPs | Reconstituted pot culture (perlite, peat, sand) | ↑SOD ↑POD | 35.9% 63.1% | [58] |
| 100 mM | Triticum aestivum L. | 5, 10 µM | Se (VI) | Reconstituted pot culture (Peat, compost, sand) | ↑SOD ↑CAT ↑GST ↑APX ↑GR | 16.2% 10.1% 16.2% 10.6% 22.1% | [28] |
| 0, 30, 60, 90 mM | Allium sativum L. | 0, 4, 8, 16 mg·L−1 | Se (VI) | Hydroponic culture | ↑SOD ↑CAT ↓POX ↑PAL | 81.0% minute minute ~15.0% | [127,129] |
| 12 dS m−1 | Zea mays L. | 0, 20, 40 mg·L−1 | Se (VI) | Sand culture | ↑CAT ↑POD ↑SOD | ~56.0% ~63.0% minute | [122] |
| 0, 100 mM | Phaseolus vulgaris L. | 0, 5, 10 μM | Se (IV) | Pot soil culture | ↑SOD ↑POD ↑CAT | 15.8% 313.3% 56.3% | [132] |
| 8 dS m−1 | Allium cepa L. | 0, 0.5, 1 kg·ha−1 | Se (IV) | Field experiment | ↓CAT ↓POD | 26.6% 10.0% | [131] |
| 0, 25, 50 mM | Lycopersicon esculentum-Mill. | 0, 5, 10 µM | Se (IV) | Hydroponic culture | ↓POD ↑CAT | 60.0% ~240.0% | [128] |
| 0, 50 mM | Vigna unguiculata L. | 5, 10 µM | Se (VI) | Sand-soil culture | ↑SOD ↑POD ↑PAL | 63.4% 238.1% 73.5% | [169] |
| 0, 100 mM | Vigna radiata L. Wilczek | 1, 2.5, 5 ppm | Se (VI) | Reconstituted pot culture(Soil, sand, farmyard manure) | ↑SOD ↑CAT ↑APX ↑GR ↑GPX | 14.2% 37.0% 34.8% 24.6% 41.0% | [170] |
| 0, 10 dS m−1 | Anethum graveolens L. | 0, 5 µM | Se (VI) | Hydroponic culture | ↑CAT ↑SOD ↓APX | ~40.0% ~19.0% minute | [147] |
| 0, 100 mM | Lycopersicon esculentum-Mill. Shuangfeng 87-5 | 0.05 mM | Se (IV) | Hydroponic culture | ↑GR ↓APX ↑DHAR ↑MDAR | ~23.0% ~14.0% ~50.0% ~16.0% | [30] |
| 0, 100 mM | Glycine max var. L17 | 0, 25, 50 mg·L−1 | Pot soil culture | ↑CAT ↑POD ↑SOD | 221.6% 85.0% 40.0% | [171] | |
| 0, 100 mM | Cucumis melo L. | 0, 2, 4, 8, 16 μM | Se (IV) | Hydroponic culture | ↑POD CAT ↑SOD | ~29.0% unchanged ~106.0% | [172] |
| 0, 100, 200 mM | Brassica napus L. | 25 µM | Se (VI) | Semi-hydroponic culture | ↑GSH ↑GSH/GSSG ↑DHAR ↑MDHAR ↑GST ↑GR | 33.0% 86.0% 43.0% 45.0% 18.0% 40.0% | [24] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kamran, M.; Parveen, A.; Ahmar, S.; Malik, Z.; Hussain, S.; Chattha, M.S.; Saleem, M.H.; Adil, M.; Heidari, P.; Chen, J.-T. An Overview of Hazardous Impacts of Soil Salinity in Crops, Tolerance Mechanisms, and Amelioration through Selenium Supplementation. Int. J. Mol. Sci. 2020, 21, 148. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21010148
Kamran M, Parveen A, Ahmar S, Malik Z, Hussain S, Chattha MS, Saleem MH, Adil M, Heidari P, Chen J-T. An Overview of Hazardous Impacts of Soil Salinity in Crops, Tolerance Mechanisms, and Amelioration through Selenium Supplementation. International Journal of Molecular Sciences. 2020; 21(1):148. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21010148
Chicago/Turabian StyleKamran, Muhammad, Aasma Parveen, Sunny Ahmar, Zaffar Malik, Sajid Hussain, Muhammad Sohaib Chattha, Muhammad Hamzah Saleem, Muhammad Adil, Parviz Heidari, and Jen-Tsung Chen. 2020. "An Overview of Hazardous Impacts of Soil Salinity in Crops, Tolerance Mechanisms, and Amelioration through Selenium Supplementation" International Journal of Molecular Sciences 21, no. 1: 148. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21010148