Characterization of a Maltase from an Early-Diverged Non-Conventional Yeast Blastobotrys adeninivorans

 ,
,  and
and
Abstract
:1. Introduction
2. Results
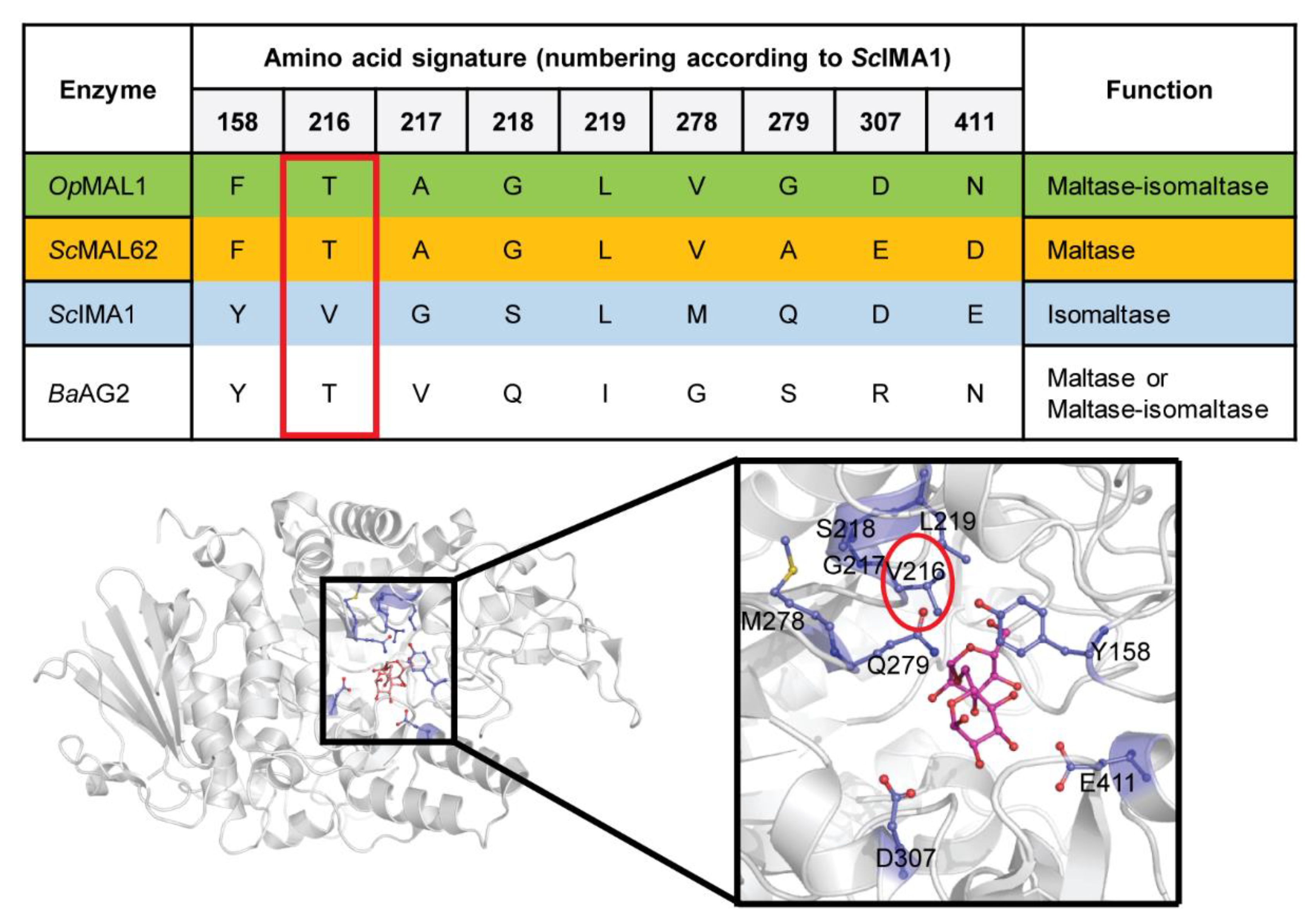
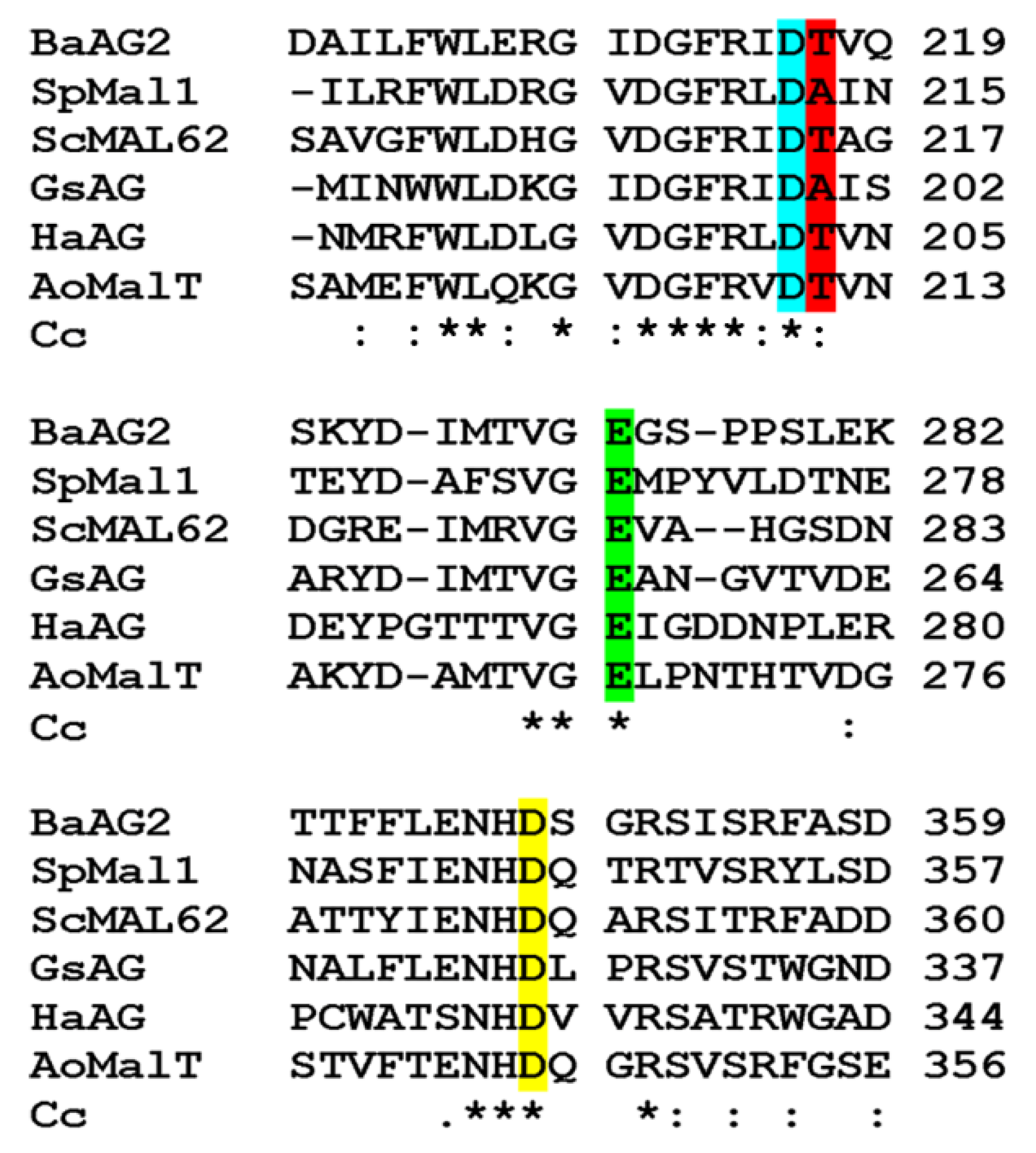
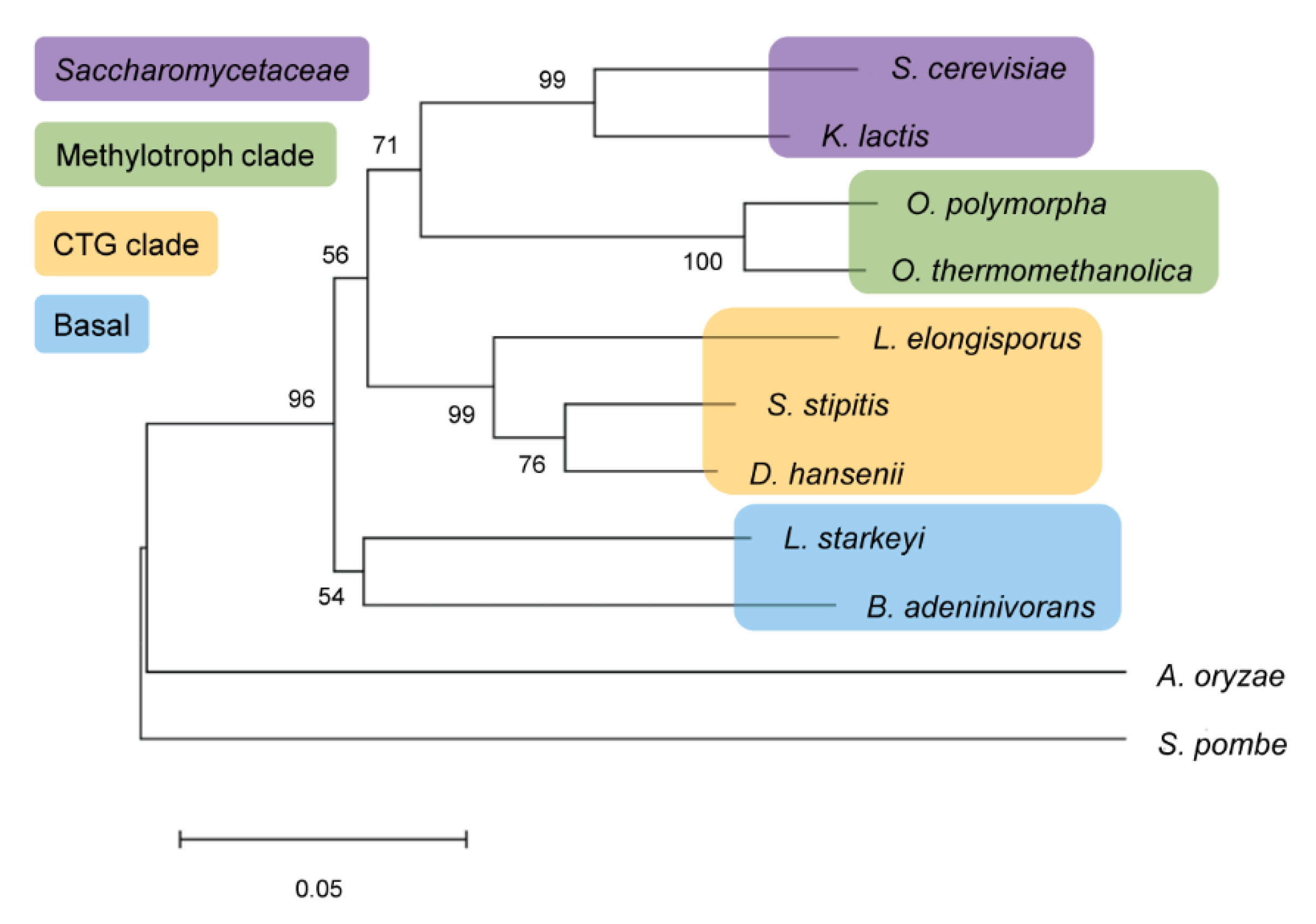
2.1. In Silico Analysis of BaAG2
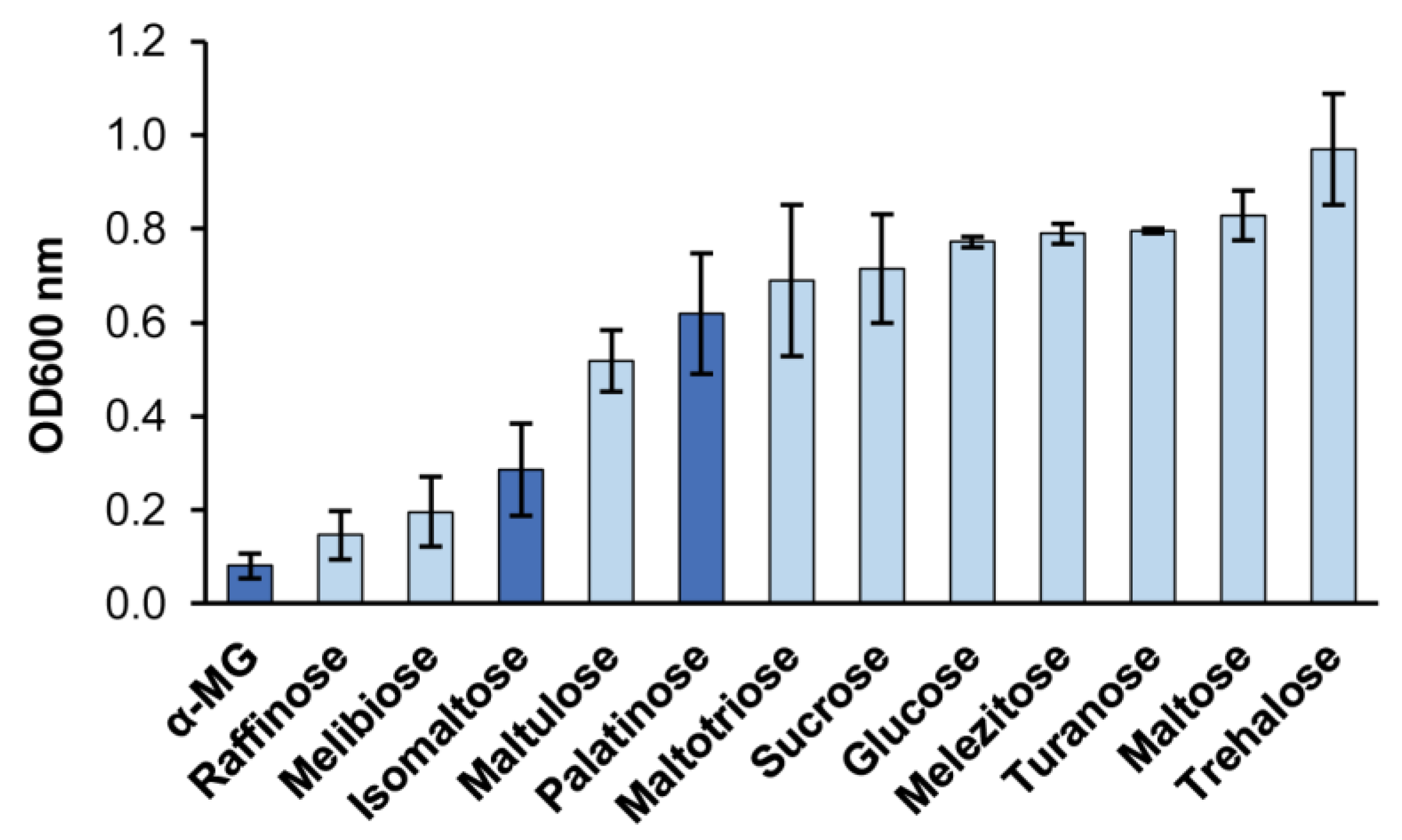
2.2. Maltose-Like and Isomaltose-Like Sugars Are Growth Substrates for B. adeninivorans
2.3. Cloning of the BaAG2 Gene and Heterologous Expression of the BaAG2 Enzyme
2.4. Properties of BaAG2
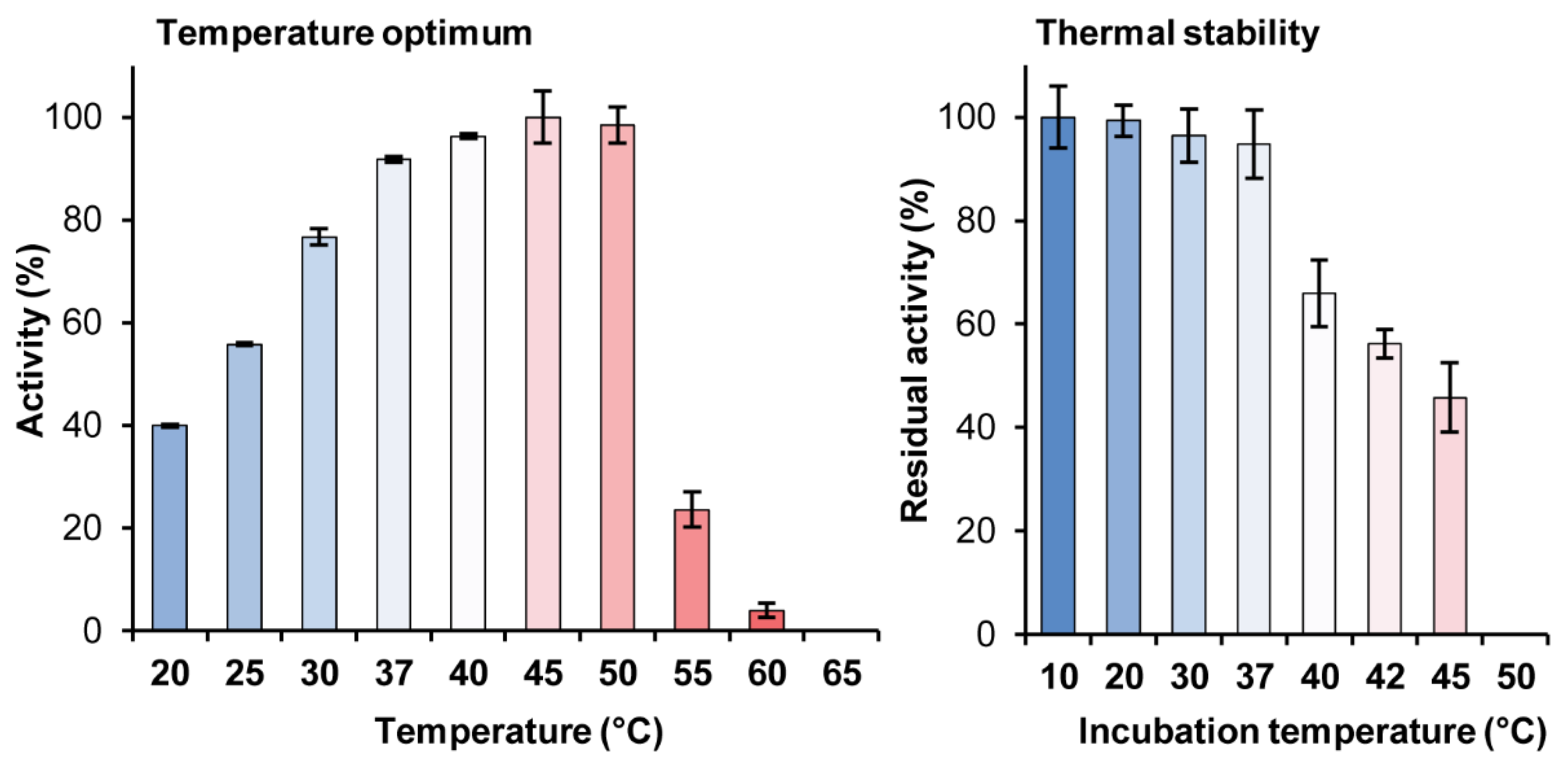
2.4.1. Dependence of the BaAG2 Activity on Temperature and pH. Thermal Stability of BaAG2
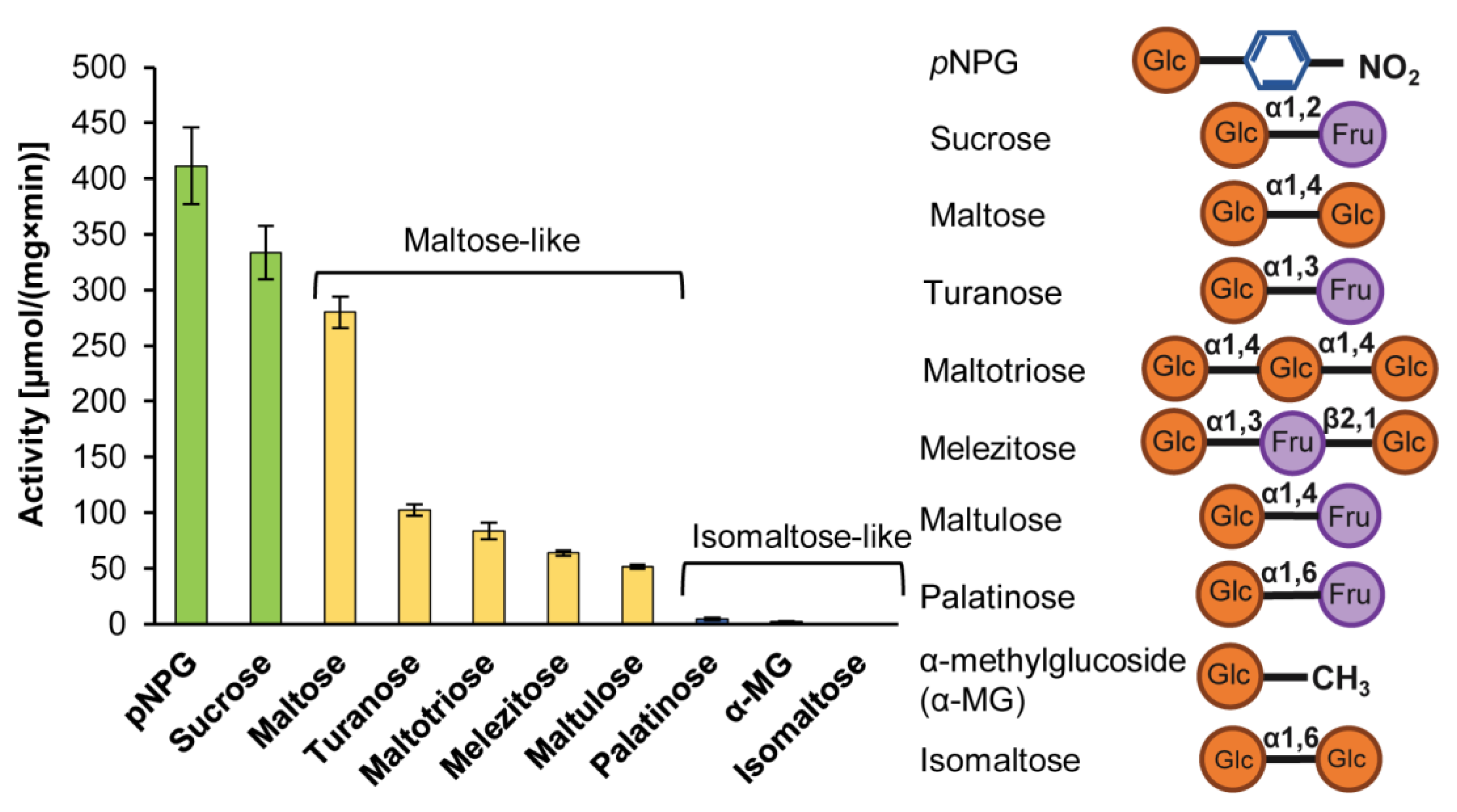
2.4.2. The Hydrolysis of Maltose and Maltose-Like Sugars
2.4.3. The Kinetic Parameters of Hydrolysis of Maltose, Maltose-Like and Universal Substrates
2.4.4. The Inhibition Studies of Acarbose, Tris, Isomaltose-Like Sugars and Glucose
2.4.5. The Hydrolysis of Malto-Oligosaccharides of DP 3 to 7
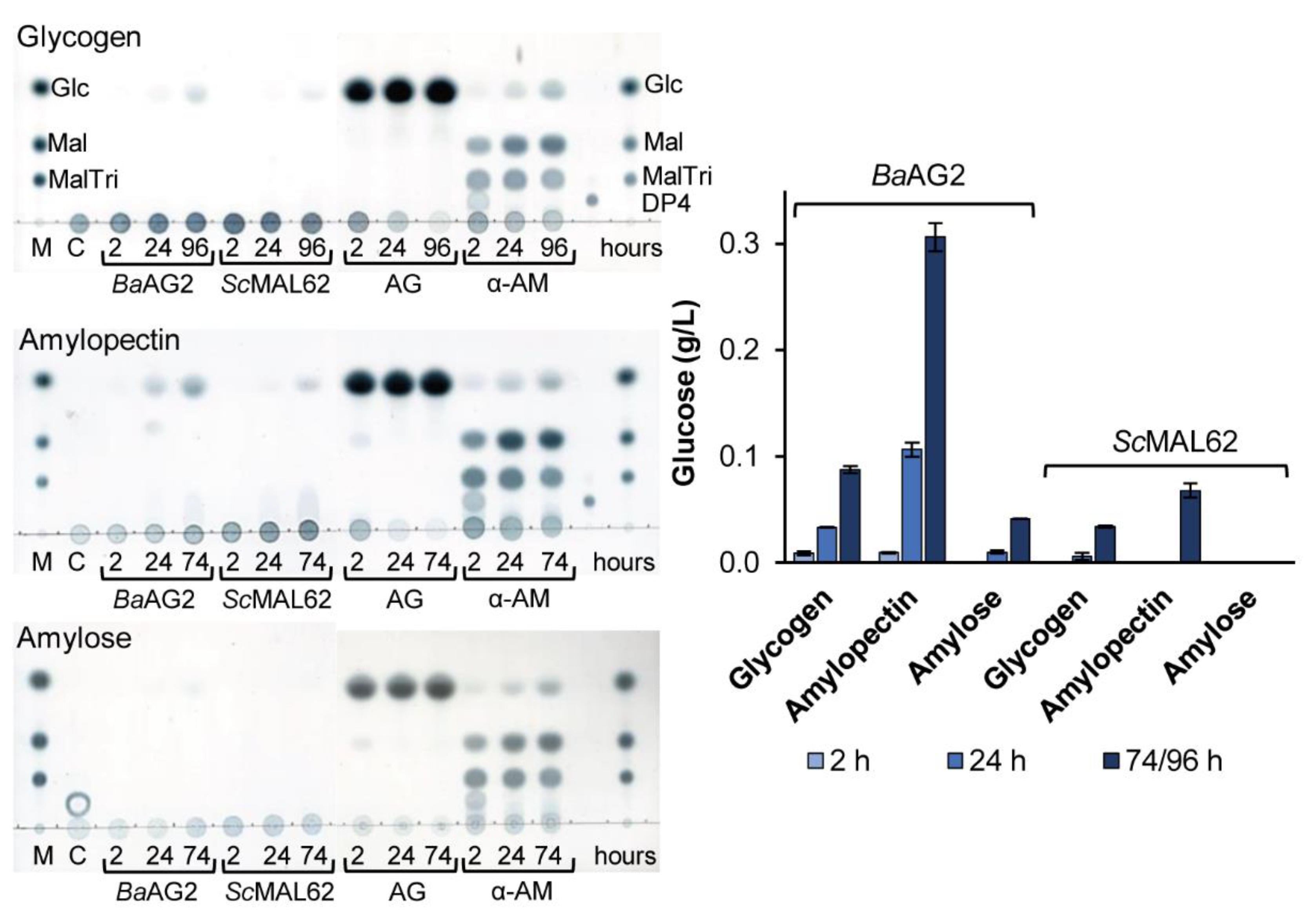
2.4.6. The Hydrolysis of Amylose, Amylopectin and Glycogen
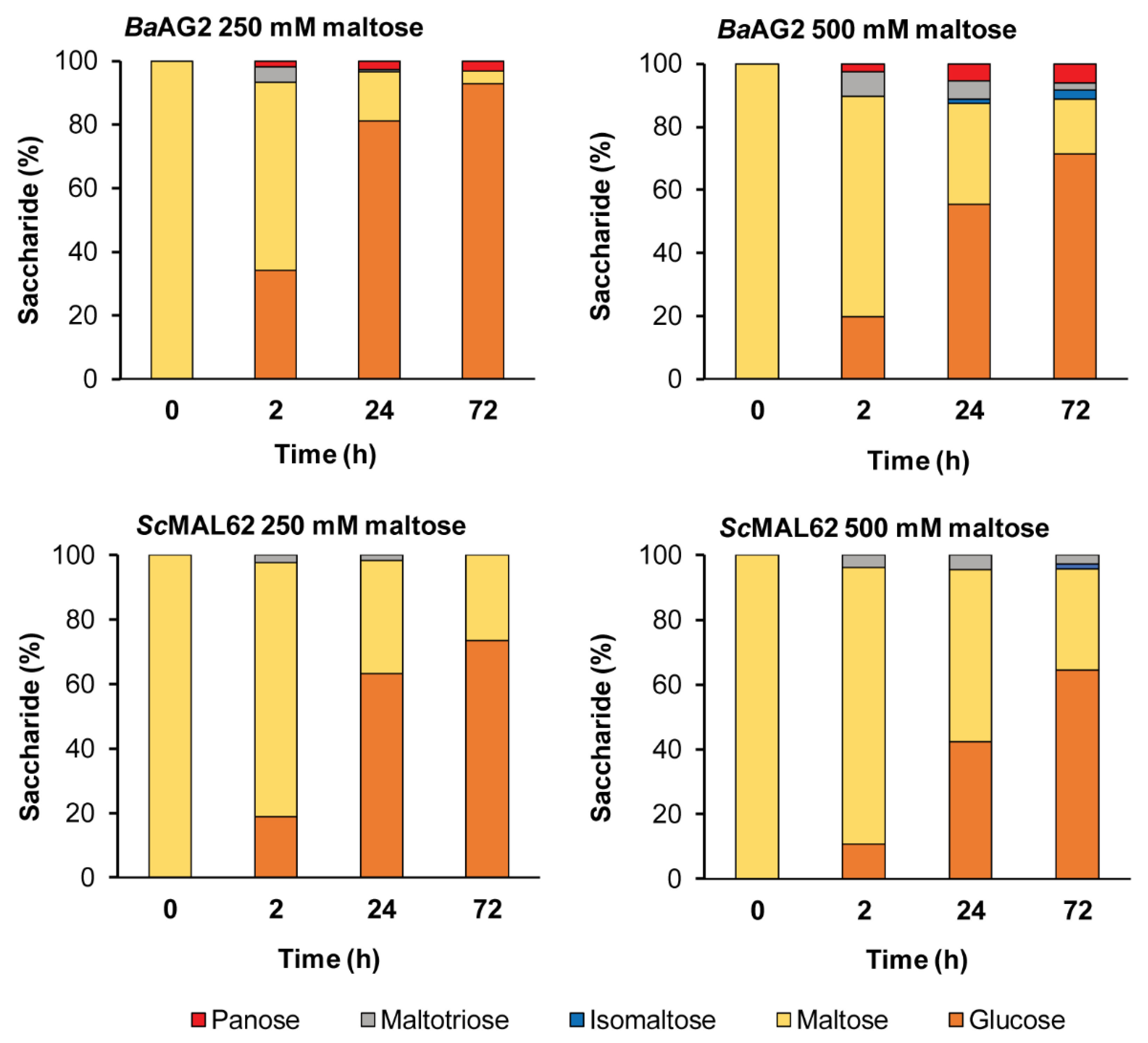
2.4.7. The Transglycosylation of Maltose
3. Discussion
4. Materials and Methods
4.1. Yeast and Bacterial Strains, Cultivation of B. adeninivorans
4.2. Cloning, Heterologous Expression and Protein Purification
4.3. Determination of Substrate Specificity, Kinetic Parameters and Inhibition
4.4. Determination of pH and Temperature Optima, Evaluation of Thermostability
4.5. Study of Transglycosylation
4.6. Chromatography of Substrates and Reaction Products
4.7. Alignment of RNA and Protein Sequences and Construction of the Phylogram
4.8. Extraction of Amino Acid Signature from the Alignments and Visualization of Respective Positions on the three-dimensional (3D) Model of S. cerevisiae Isomaltase IMA1
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| BaAG2 | α-glucosidase 2 of Blastobotrys adeninivorans |
| BSA | bovine serum albumin |
| DP | degree of polymerization |
| DSF | differential scanning fluorimetry |
| GH | glycoside hydrolase |
| MOS | malto-oligosaccharides |
| α-MG | α-methylglucoside |
| PDB | RCSB Protein Data Bank |
| pNPG | p-nitrophenyl-α-d-glucopyranoside |
| OD | optical density |
| rRNA | ribosomal RNA |
| ScMAL62 | maltase MAL62 of Saccharomyces cerevisiae |
| SD | standard deviation |
| TLC | thin layer chromatography |
| Tm | melting temperature |
| Tris | tris(hydroxymethyl)aminomethane |
| YNB | Yeast Nitrogen Base |
References
- Kurtzman, C.P.; Robnett, C.J. Multigene phylogenetic analysis of the Trichomonascus, Wickerhamiella and Zygoascus yeast clades, and the proposal of Sugiyamaella gen. nov. and 14 new species combinations. FEMS Yeast Res. 2007, 7, 141–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kunze, G.; Gaillardin, C.; Czernicka, M.; Durrens, P.; Martin, T.; Böer, E.; Gabaldón, T.; Cruz, J.A.; Talla, E.; Marck, C.; et al. The complete genome of Blastobotrys (Arxula) adeninivorans LS3—A yeast of biotechnological interest. Biotechnol. Biofuels 2014, 7, 66. [Google Scholar] [CrossRef]
- Shen, X.-X.; Zhou, X.; Kominek, J.; Kurtzman, C.P.; Hittinger, C.T.; Rokas, A. Reconstructing the backbone of the Saccharomycotina yeast phylogeny using genome-scale data. G3 Genes Genomes Genet. 2016, 6, 3927–3939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Correia, K.; Yu, S.M.; Mahadevan, R. AYbRAH: A curated ortholog database for yeasts and fungi spanning 600 million years of evolution. Database 2019, 2019, baz022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hedges, S.B.; Marin, J.; Suleski, M.; Paymer, M.; Kumar, S. Tree of life reveals clock-like speciation and diversification. Mol. Biol. Evol. 2015, 32, 835–845. [Google Scholar] [CrossRef] [PubMed]
- Thomas, S.; Sanya, D.R.A.; Fouchard, F.; Nguyen, H.V.; Kunze, G.; Neuvéglise, C.; Crutz-Le Coq, A.M. Blastobotrys adeninivorans and B. raffinosifermentans, two sibling yeast species which accumulate lipids at elevated temperatures and from diverse sugars. Biotechnol. Biofuels 2019, 12, 154. [Google Scholar] [CrossRef] [PubMed]
- Malak, A.; Baronian, K.; Kunze, G. Blastobotrys (Arxula) adeninivorans: A promising alternative yeast for biotechnology and basic research. Yeast 2016, 33, 535–547. [Google Scholar] [CrossRef] [Green Version]
- Kasprzak, J.; Rauter, M.; Riechen, J.; Worch, S.; Baronian, K.; Bode, R.; Schauer, F.; Kunze, G. Characterization of an Arxula adeninivorans alcohol dehydrogenase involved in the metabolism of ethanol and 1-butanol. FEMS Yeast Res. 2016, 16, fow018. [Google Scholar] [CrossRef] [Green Version]
- Bui, D.M.; Kunze, I.; Förster, S.; Wartmann, T.; Horstmann, C.; Manteuffel, R.; Kunze, G. Cloning and expression of an Arxula adeninivorans glucoamylase gene in Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 1996, 44, 610–619. [Google Scholar] [CrossRef]
- Böer, E.; Wartmann, T.; Luther, B.; Manteuffel, R.; Bode, R.; Gellissen, G.; Kunze, G. Characterization of the AINV gene and the encoded invertase from the dimorphic yeast Arxula adeninivorans. Antonie Van Leeuwenhoek 2004, 86, 121–134. [Google Scholar] [CrossRef]
- Bao, M.; Niu, C.; Xu, X.; Zheng, F.; Liu, C.; Wang, J.; Li, Q. Identification, soluble expression, and characterization of a novel endo-inulinase from Lipomyces starkeyi NRRL Y-11557. Int. J. Biol. Macromol. 2019, 137, 537–544. [Google Scholar] [CrossRef] [PubMed]
- Viigand, K.; Põšnograjeva, K.; Visnapuu, T.; Alamäe, T. Genome mining of non-conventional yeasts: Search and analysis of MAL clusters and proteins. Genes 2018, 9, 354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gabriško, M. Evolutionary history of eukaryotic α-glucosidases from the α-amylase family. J. Mol. Evol. 2013, 76, 129–145. [Google Scholar] [CrossRef] [PubMed]
- Janeček, Š.; Gabriško, M. Remarkable evolutionary relatedness among the enzymes and proteins from the α-amylase family. Cell. Mol. Life Sci. 2016, 73, 2707–2725. [Google Scholar] [CrossRef]
- Voordeckers, K.; Brown, C.A.; Vanneste, K.; van der Zande, E.; Voet, A.; Maere, S.; Verstrepen, K.J. Reconstruction of ancestral metabolic enzymes reveals molecular mechanisms underlying evolutionary innovation through gene duplication. PLoS Biol. 2012, 10, e1001446. [Google Scholar] [CrossRef] [Green Version]
- Viigand, K.; Visnapuu, T.; Mardo, K.; Aasamets, A.; Alamäe, T. Maltase protein of Ogataea (Hansenula) polymorpha is a counterpart to the resurrected ancestor protein ancMALS of yeast maltases and isomaltases. Yeast 2016, 33, 415–432. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Arrojo, L.; Marín, D.; Gómez De Segura, A.; Linde, D.; Alcalde, M.; Gutiérrez-Alonso, P.; Ghazi, I.; Plou, F.J.J.; Fernández-Lobato, M.; Ballesteros, A. Transformation of maltose into prebiotic isomaltooligosaccharides by a novel α-glucosidase from Xantophyllomyces dendrorhous. Process Biochem. 2007, 42, 1530–1536. [Google Scholar] [CrossRef] [Green Version]
- Mangas-Sánchez, J.; Adlercreutz, P. Enzymatic preparation of oligosaccharides by transglycosylation: A comparative study of glucosidases. J. Mol. Catal. B Enzym. 2015, 122, 51–55. [Google Scholar] [CrossRef] [Green Version]
- Casa-Villegas, M.; Marín-Navarro, J.; Polaina, J. Amylases and related glycoside hydrolases with transglycosylation activity used for the production of isomaltooligosaccharides. Amylase 2018, 2, 17–29. [Google Scholar] [CrossRef]
- Gutiérrez-Alonso, P.; Gimeno-Pérez, M.; Ramírez-Escudero, M.; Plou, F.J.; Sanz-Aparicio, J.; Fernández-Lobato, M. Molecular characterization and heterologous expression of a Xanthophyllomyces dendrorhous α-glucosidase with potential for prebiotics production. Appl. Microbiol. Biotechnol. 2016, 100, 3125–3135. [Google Scholar] [CrossRef] [Green Version]
- Chiba, S.; Murata, M.; Matsusaka, K.; Shimomura, T. A new trisaccharide, 6F-α-D-glucosyl-sucrose, synthesized by transglucosylation reaction of brewer’s yeast α-glucosidase. Agric. Biol. Chem. 1979, 43, 775–779. [Google Scholar] [CrossRef] [Green Version]
- Stewart, G. Saccharomyces species in the production of beer. Beverages 2016, 2, 34. [Google Scholar] [CrossRef]
- Needleman, R.B.; Marmur, J.; Federoff, H.J.; Eccleshall, T.R.; Buchferer, B. Purification and characterization of an α-glucosidase from Saccharomyces carlsbergensis. Biochemistry 1978, 17, 4657–4661. [Google Scholar] [CrossRef] [PubMed]
- Krakenaĭte, R.P.; Glemzha, A.A. Some properties of two forms of alpha-glucosidase from Saccharomyces cerevisiae-II. Biokhimiia 1983, 48, 62–68. [Google Scholar]
- Alamäe, T.; Viigand, K.; Põšnograjeva, K. Utilization of α-glucosidic disaccharides by Ogataea (Hansenula) polymorpha: Genes, proteins, and regulation. In Non-Conventional Yeasts: From Basic Research to Application; Sibirny, A., Ed.; Springer International Publishing: Cham, Switzerland, 2019; pp. 1–22. [Google Scholar]
- Grigoriev, I.V.; Nikitin, R.; Haridas, S.; Kuo, A.; Ohm, R.; Otillar, R.; Riley, R.; Salamov, A.; Zhao, X.; Korzeniewski, F.; et al. MycoCosm portal: Gearing up for 1000 fungal genomes. Nucleic Acids Res. 2014, 42, D699–D704. [Google Scholar] [CrossRef]
- Yamamoto, K.; Miyake, H.; Kusunoki, M.; Osaki, S. Crystal structures of isomaltase from Saccharomyces cerevisiae and in complex with its competitive inhibitor maltose. FEBS J. 2010, 277, 4205–4214. [Google Scholar] [CrossRef]
- Yamamoto, K.; Nakayama, A.; Yamamoto, Y.; Tabata, S. Val216 decides the substrate specificity of α-glucosidase in Saccharomyces cerevisiae. Eur. J. Biochem. 2004, 271, 3414–3420. [Google Scholar] [CrossRef]
- Yamamoto, K.; Miyake, H.; Kusunoki, M.; Osaki, S. Steric hindrance by 2 amino acid residues determines the substrate specificity of isomaltase from Saccharomyces cerevisiae. J. Biosci. Bioeng. 2011, 112, 545–550. [Google Scholar] [CrossRef]
- Schrödinger, L.L.C. The {PyMOL} Molecular Graphics System, Version~1.8.6.0. 2015. Available online: https://pymol.org/2/ (accessed on 19 May 2017).
- Janeček, Š.; Svensson, B.; MacGregor, E.A. A remote but significant sequence homology between glycoside hydrolase clan GH-H and family GH31. FEBS Lett. 2007, 581, 1261–1268. [Google Scholar] [CrossRef] [Green Version]
- Uitdehaag, J.C.M.; Mosi, R.; Kalk, K.H.; Van der Veen, B.A.; Dijkhuizen, L.; Withers, S.G.; Dijkstra, B.W. X-ray structures along the reaction pathway of cyclodextrin glycosyltransferase elucidate catalysis in the α-amylase family. Nat. Struct. Biol. 1999, 6, 432–436. [Google Scholar] [CrossRef] [Green Version]
- Chi, Z.; Ni, X.; Yao, S. Cloning and overexpression of a maltase gene from Schizosaccharomyces pombe in Escherichia coli and characterization of the recombinant maltase. Mycol. Res. 2008, 112, 983–989. [Google Scholar] [CrossRef] [PubMed]
- Tsujimoto, Y.; Tanaka, H.; Takemura, R.; Yokogawa, T.; Shimonaka, A.; Matsui, H.; Kashiwabara, S.I.; Watanabe, K.; Suzuki, Y. Molecular determinants of substrate recognition in thermostable α-glucosidases belonging to glycoside hydrolase family 13. J. Biochem. 2007, 142, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Ojima, T.; Saburi, W.; Yamamoto, T.; Kudo, T. Characterization of Halomonas sp. strain H11 α-glucosidase activated by monovalent cations and its application for efficient synthesis of α-D-glucosylglycerol. Appl. Environ. Microbiol. 2012, 78, 1836–1845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasegawa, S.; Takizawa, M.; Suyama, H.; Shintani, T.; Gomi, K. Characterization and expression analysis of a maltose-utilizing (MAL) cluster in Aspergillus oryzae. Fungal Genet. Biol. 2010, 47, 1–9. [Google Scholar] [CrossRef] [PubMed]
- CBS Database. Available online: http://www.wi.knaw.nl/Collections/DefaultInfo.aspx?Page=Home (accessed on 27 November 2019).
- Teste, M.A.; Marie François, J.; Parrou, J.L. Characterization of a new multigene family encoding isomaltases in the yeast Saccharomyces cerevisiae, the IMA family. J. Biol. Chem. 2010, 285, 26815–26824. [Google Scholar] [CrossRef] [Green Version]
- Almagro Armenteros, J.J.; Tsirigos, K.D.; Sønderby, C.K.; Petersen, T.N.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 5.0 improves signal peptide predictions using deep neural networks. Nat. Biotechnol. 2019, 37, 420–423. [Google Scholar] [CrossRef]
- Liiv, L.; Pärn, P.; Alamäe, T. Cloning of maltase gene from a methylotrophic yeast, Hansenula polymorpha. Gene 2001, 265, 77–85. [Google Scholar] [CrossRef]
- Geber, A.; Williamson, P.R.; Rex, J.H.; Sweeney, E.C.; Bennett, J.E. Cloning and characterization of a Candida albicans maltase gene involved in sucrose utilization. J. Bacteriol. 1992, 174, 6992–6996. [Google Scholar] [CrossRef] [Green Version]
- Alamäe, T.; Liiv, L. Glucose repression of maltase and methanol-oxidizing enzymes in the methylotrophic yeast Hansenula polymorpha: Isolation and study of regulatory mutants. Folia Microbiol. 1998, 43, 443–452. [Google Scholar] [CrossRef]
- Marín, D.; Linde, D.; Lobato, M.F. Purification and biochemical characterization of an α-glucosidase from Xanthophyllomyces dendrorhous. Yeast 2006, 23, 117–125. [Google Scholar] [CrossRef] [Green Version]
- da Silva, T.M.; Michelin, M.; de Lima Damásio, A.R.; Maller, A.; Almeida, F.B.D.R.; Ruller, R.; Ward, R.J.; Rosa, J.C.; Jorge, J.A.; Terenzi, H.F.; et al. Purification and biochemical characterization of a novel α-glucosidase from Aspergillus niveus. Antonie Van Leeuwenhoek 2009, 96, 569–578. [Google Scholar] [CrossRef] [PubMed]
- Kita, A.; Matsui, H.; Somoto, A.; Kimura, A.; Takata, M.; Chiba, S. Substrate specificity and subsite affinities of crystalline α-glucosidase from Aspergillus niger. Agric. Biol. Chem. 1991, 55, 2327–2335. [Google Scholar] [CrossRef]
- Okuyama, M.; Tanimoto, Y.; Ito, T.; Anzai, A.; Mori, H.; Kimura, A.; Matsui, H.; Chiba, S. Purification and characterization of the hyper-glycosylated extracellular α-glucosidase from Schizosaccharomyces pombe. Enzym. Microb. Technol. 2005, 37, 472–480. [Google Scholar] [CrossRef]
- Ernits, K.; Viigand, K.; Visnapuu, T.; Põšnograjeva, K.; Alamäe, T. Thermostability measurement of an α-glucosidase using a classical activity-based assay and a novel Thermofluor method. Bio-Protocol 2017, 7. [Google Scholar] [CrossRef]
- Naumov, G.I.; Naumova, E.S.; Michels, C.A. Genetic variation of the repeated MAL loci in natural populations of Saccharomyces cerevisiae and Saccharomyces paradoxus. Genetics 1994, 136, 803–812. [Google Scholar]
- Reinders, A.; Ward, J.M. Functional characterization of the α-glucoside transporter Sut1p from Schizosaccharomyces pombe, the first fungal homologue of plant sucrose transporters. Mol. Microbiol. 2001, 39, 445–454. [Google Scholar] [CrossRef] [Green Version]
- Dujon, B.A.; Louis, E.J. Genome diversity and evolution in the budding yeasts (Saccharomycotina). Genetics 2017, 206, 717–750. [Google Scholar] [CrossRef] [Green Version]
- Kelly, C.T.; Moriarty, M.E.; Fogarty, W.M. Thermostable extracellular α-amylase and α-glucosidase of Lipomyces starkeyi. Appl. Microbiol. Biotechnol. 1985, 22, 352–358. [Google Scholar] [CrossRef]
- Egeter, O.; Bruckner, R. Characterization of a genetic locus essential for maltose-maltotriose utilization in Staphylococcus xylosus. J. Bacteriol. 1995, 177, 2408–2415. [Google Scholar] [CrossRef] [Green Version]
- Schönert, S.; Buder, T.; Dahl, M.K. Identification and enzymatic characterization of the maltose-inducible α-glucosidase MalL (sucrase-isomaltase-maltase) of Bacillus subtilis. J. Bacteriol. 1998, 180, 2574–2578. [Google Scholar] [CrossRef] [Green Version]
- Rolfsmeier, M.; Blum, P. Purification and characterization of a maltase from the extremely thermophilic crenarchaeote Sulfolobus solfataricus. J. Bacteriol. 1995, 177, 482–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, X.; Petitjean, M.; Teste, M.A.; Kooli, W.; Tranier, S.; François, J.M.; Parrou, J.L. Similarities and differences in the biochemical and enzymological properties of the four isomaltases from Saccharomyces cerevisiae. FEBS Open Bio. 2014, 4, 200–212. [Google Scholar] [CrossRef] [PubMed]
- McWethy, S.J.; Hartman, P.A. Extracellular maltase of Bacillus brevis. Appl. Environ. Microbiol. 1979, 37, 1096–1102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumara, H.M.; De Cort, S.; Verachtert, H. Localization and characterization of alpha-glucosidase activity in Brettanomyces lambicus. Appl. Environ. Microbiol. 1993, 59, 2352–2358. [Google Scholar] [CrossRef] [Green Version]
- Wilson, W.A.; Roach, P.J.; Montero, M.; Baroja-Fernández, E.; Muñoz, F.J.; Eydallin, G.; Viale, A.M.; Pozueta-Romero, J. Regulation of glycogen metabolism in yeast and bacteria. FEMS Microbiol. Rev. 2010, 34, 952–985. [Google Scholar] [CrossRef] [Green Version]
- Casa-Villegas, M.; Marín-Navarro, J.; Polaina, J. Synthesis of isomaltooligosaccharides by Saccharomyces cerevisiae cells expressing Aspergillus niger α-glucosidase. ACS Omega 2017, 2, 8062–8068. [Google Scholar] [CrossRef] [Green Version]
- Kato, N.; Suyama, S.; Shirokane, M.; Kato, M.; Kobayashi, T.; Tsukagoshi, N. Novel α-glucosidase from Aspergillus nidulans with strong transglycosylation activity. Appl. Environ. Microbiol. 2002, 68, 1250–1256. [Google Scholar] [CrossRef] [Green Version]
- Studier, F.W.; Moffatt, B.A. Use of bacteriophage T7 RNA polymerase to direct selective high-level expression of cloned genes. J. Mol. Biol. 1986, 189, 113–130. [Google Scholar] [CrossRef]
- Curiel, J.A.; de Las Rivas, B.; Mancheño, J.M.; Muñoz, R. The pURI family of expression vectors: A versatile set of ligation independent cloning plasmids for producing recombinant His-fusion proteins. Protein Expr. Purif. 2011, 76, 44–53. [Google Scholar] [CrossRef] [Green Version]
- Visnapuu, T.; Mardo, K.; Mosoarca, C.; Zamfir, A.D.; Vigants, A.; Alamäe, T. Levansucrases from Pseudomonas syringae pv. tomato and P. chlororaphis subsp. aurantiaca: Substrate specificity, polymerizing properties and usage of different acceptors for fructosylation. J. Biotechnol. 2011, 155, 338–349. [Google Scholar] [CrossRef]
- Ernits, K.; Eek, P.; Lukk, T.; Visnapuu, T.; Alamäe, T. First crystal structure of an endo-levanase—The BT1760 from a human gut commensal Bacteroides thetaiotaomicron. Sci. Rep. 2019, 9, 8443. [Google Scholar] [CrossRef] [PubMed]
- Mardo, K.; Visnapuu, T.; Gromkova, M.; Aasamets, A.; Viigand, K.; Vija, H.; Alamäe, T. High-throughput assay of levansucrase variants in search of feasible catalysts for the synthesis of fructooligosaccharides and levan. Molecules 2014, 19, 8434–8455. [Google Scholar] [CrossRef] [PubMed]
- Mcilvaine, T.C. A buffer solution for colorimetric comparison. J. Biol. Chem. 1921, 49, 183–186. [Google Scholar]
- Stingele, F.; Newell, J.W.; Neeser, J.R. Unraveling the function of glycosyltransferases in Streptococcus thermophilus Sfi6. J. Bacteriol. 1999, 181, 6354–6360. [Google Scholar] [CrossRef] [Green Version]
- Jork, H.; Funk, W.; Fischer, W.; Wimmer, H. Thin-Layer Chromatography: Reagents and Detection Methods; Ebel, H.F., Ed.; VCH Verlagsgesellschaft mbH: Weinheim, Germany, 1990; Volume 1a. [Google Scholar]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Tamura, K.; Nei, M.; Kumar, S. Prospects for inferring very large phylogenies by using the neighbor-joining method. Proc. Natl. Acad. Sci. USA 2004, 101, 11030–11035. [Google Scholar] [CrossRef] [Green Version]
- Sievers, F.; Higgins, D.G. Clustal Omega for making accurate alignments of many protein sequences. Protein Sci. 2018, 27, 135–145. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substrate * | Km ± SD (mM) | Vmax ± SD (μmol/(mg × min)) | kcat ± SD (1/s) | kcat/Km (1/(mM × s)) |
|---|---|---|---|---|
| pNPG | 0.76 ± 0.03 | 751.3 ± 14.5 | 850.2 ± 16.4 | 1106.1 |
| Maltose | 25.8 ± 1.6 | 336.4 ± 5.8 | 380.7 ± 6.5 | 14.8 |
| Maltotriose | 32.5 ± 3.3 | 117.9 ± 4.4 | 133.5 ± 5.0 | 4.1 |
| Sucrose | 35.9 ± 2.7 | 412.4 ± 8.9 | 466.8 ± 10.1 | 13.0 |
| Turanose | 45.2 ± 6.0 | 190.1 ± 10.4 | 215.1 ± 11.9 | 4.8 |
| Maltulose | 7.8 ± 1.0 | 52.2 ± 1.8 | 59.1 ± 2.1 | 7.6 |
| Melezitose | 238.3 ± 51.8 | 231.0 ± 31.8 | 261.4 ± 36.1 | 1.1 |
| Inhibitor | Ki ± SD (mM) | Inhibition Mode |
|---|---|---|
| Palatinose | 1.4 ± 0.1 | Competitive |
| Isomaltose | 22.7 ± 3.0 | Competitive |
| α-MG | 21.8 ± 1.4 | Competitive |
| Acarbose | 0.83 ± 0.01* | Competitive |
| Glucose | 0.86 ± 0.05 | Competitive |
| Fructose | 36.9 ± 2.4 | Competitive |
| Tris | 70.5 ± 4.3* | Competitive |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Visnapuu, T.; Meldre, A.; Põšnograjeva, K.; Viigand, K.; Ernits, K.; Alamäe, T. Characterization of a Maltase from an Early-Diverged Non-Conventional Yeast Blastobotrys adeninivorans. Int. J. Mol. Sci. 2020, 21, 297. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21010297
Visnapuu T, Meldre A, Põšnograjeva K, Viigand K, Ernits K, Alamäe T. Characterization of a Maltase from an Early-Diverged Non-Conventional Yeast Blastobotrys adeninivorans. International Journal of Molecular Sciences. 2020; 21(1):297. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21010297
Chicago/Turabian StyleVisnapuu, Triinu, Aivar Meldre, Kristina Põšnograjeva, Katrin Viigand, Karin Ernits, and Tiina Alamäe. 2020. "Characterization of a Maltase from an Early-Diverged Non-Conventional Yeast Blastobotrys adeninivorans" International Journal of Molecular Sciences 21, no. 1: 297. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21010297