The Use of PTI-Marker Genes to Identify Novel Compounds that Establish Induced Resistance in Rice

 ,
, 
 and
and
Abstract
:1. Introduction
2. Results
2.1. In Silico Identification of Rice Genes Associated with Biotic Stress through a WGCNA
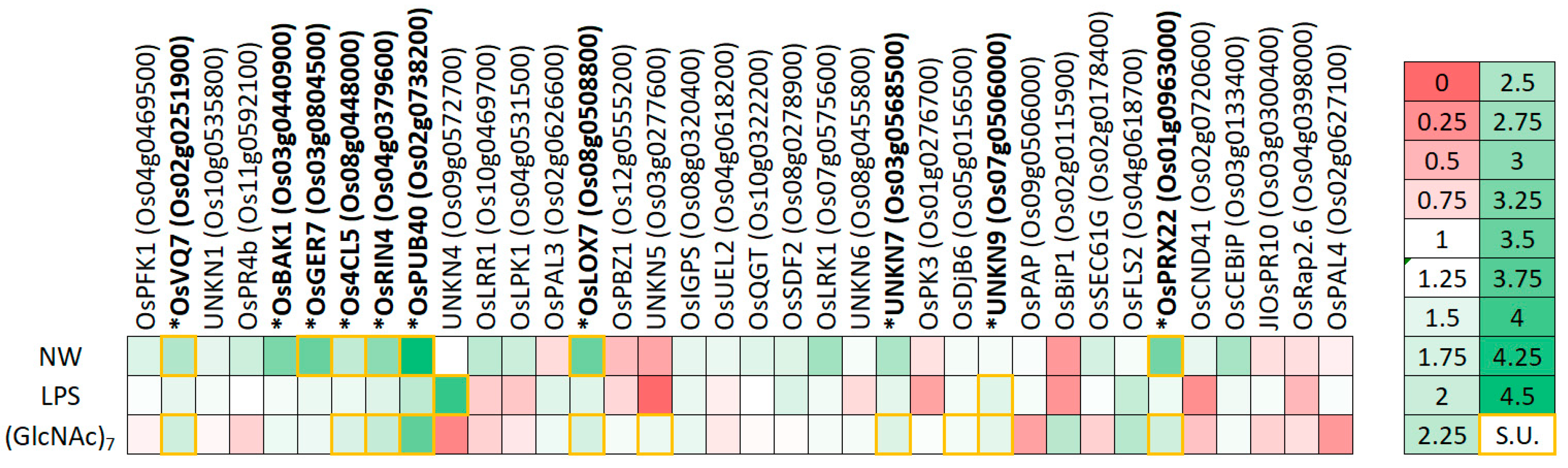
2.2. In Planta Validation of PTI-Association of Candidate Genes
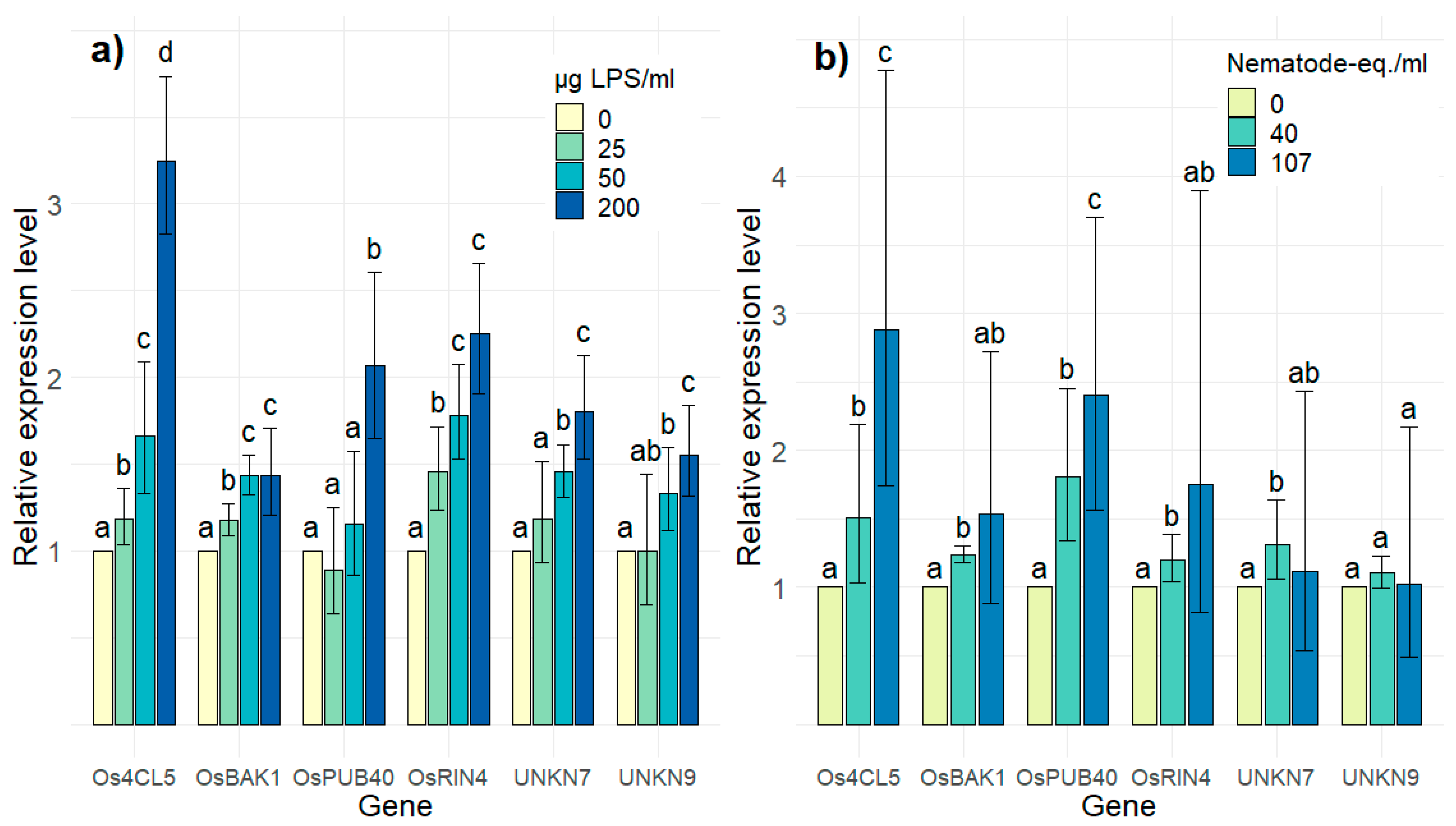
2.3. In Vitro Validation of PTI-Association of Candidate Genes
2.4. Induction of PTI-Marker Genes as Proxy to Screen for IR
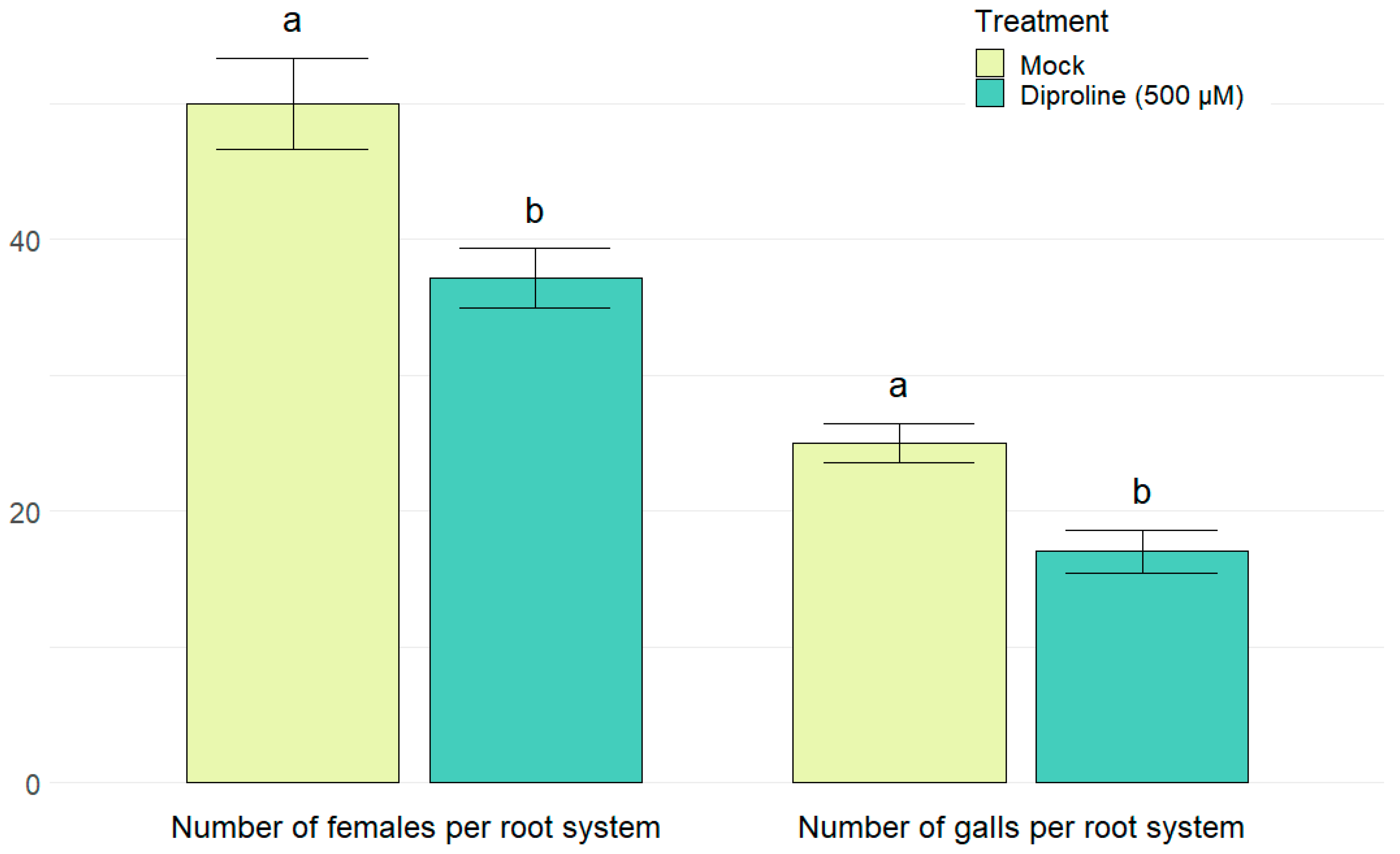
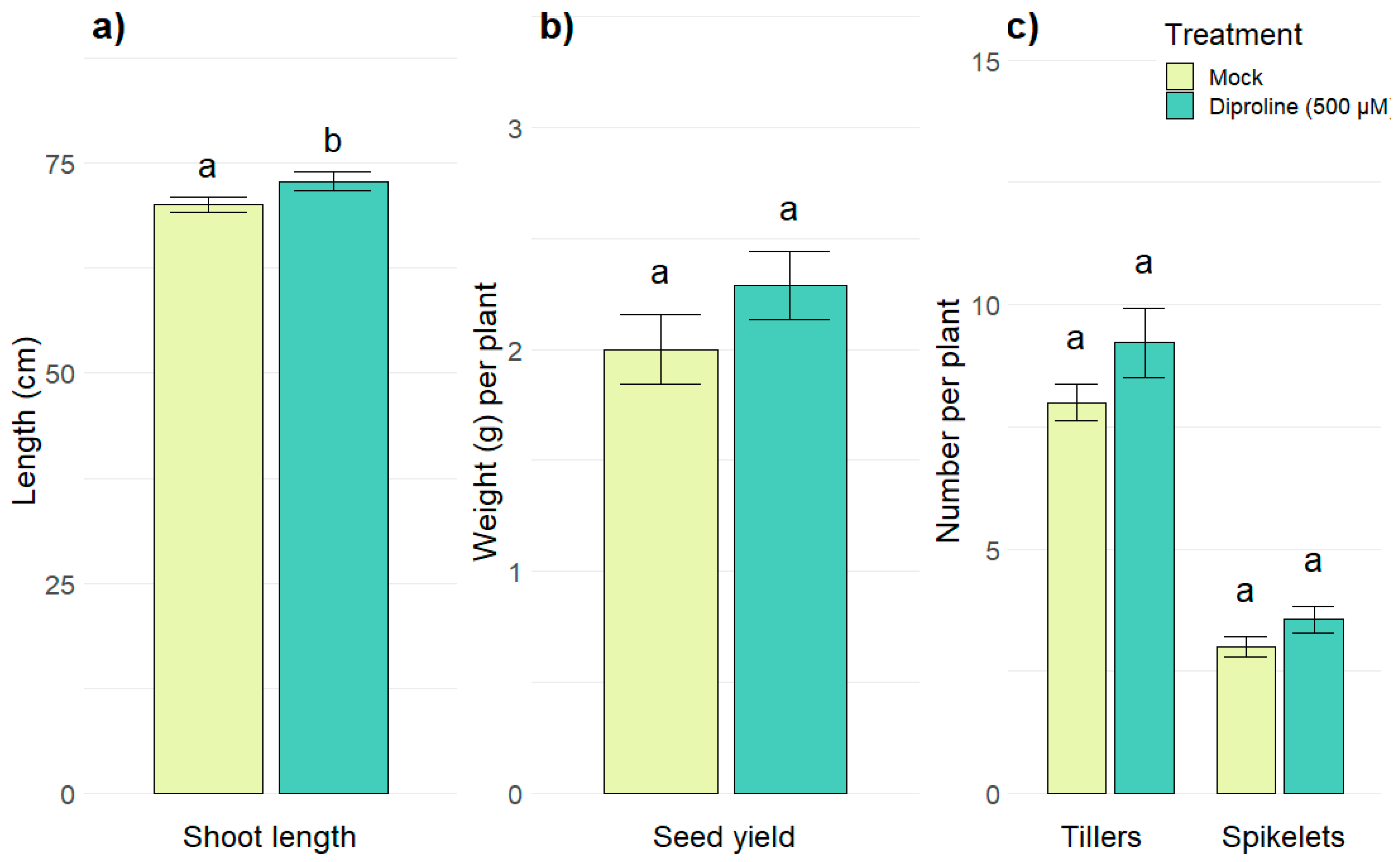
2.5. Diproline as a Novel IR-Inducing Compound for Rice
3. Discussion
4. Materials and Methods
4.1. In Silico Identification of Rice Genes Associated with Biotic Stress through WGCNA
4.1.1. Data Acquisition and Pre-Processing
4.1.2. Network Creation and Clustering
4.1.3. Identification of Biotic Stress-Associated Modules
4.1.4. WGCNA on Transcriptome Datasets of Rice under Abiotic Stress
4.2. In Planta Validation of PTI-Association of Candidate Genes
4.2.1. Gene Selection
4.2.2. In Planta Evaluation of Gene Expression Profiles upon PTI Induction
4.2.3. Data Analysis
4.3. In Vitro Validation of PTI-Association of Candidate Genes
4.3.1. Establishment of Rice Cell Suspension Cultures (RCSCs)
4.3.2. Cell Treatments and RT-qPCR
4.4. Induction of PTI-Marker Genes as Proxy to Screen for IR
4.5. Nematode Infection Experiments
4.6. Long-Term Effects of Diproline Treatments on Plant Growth and Development
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| IR | Induced resistance |
| RCSC | Rice cell suspension cultures |
| WGCNA | Weighted gene co-expression network analysis |
| PTI | Pattern-triggered immunity |
| PAMP | Pathogen-associated molecular pattern |
| NW | NemaWater |
| ETI | Effector triggered immunity |
| DHA | Dehydroascorbic acid |
| BTH | Benzo-1,2,3-thiadiazole-7-carbothioic acid S-methyl ester |
| PCSC | Plant cell suspension culture |
| BABA | β-aminobutyric acid |
| GO | Gene ontology |
| GS | Gene significance |
| MM | Module membership |
| k | Intramodular connectivity |
| ME | Module eigengenes |
| RT-qPCR | Revere transcription-quantitative polymerase chain reaction |
| LPS | Lipopolysaccharides |
| (GlcNAc)7 | N-acetylglucosamine heptamer |
References
- FAO. High Level Expert Forum—How to Feed the World in 2050. 2009. Available online: http://www.fao.org/fileadmin/templates/wsfs/docs/expert_paper/How_to_Feed_the_World_in_2050.pdf (accessed on 4 November 2019).
- Ray, D.K.; Mueller, N.D.; West, P.C.; Foley, J.A. Yield trends are insufficient to double global crop production by 2050. PLoS ONE 2013, 8, e66428. [Google Scholar] [CrossRef] [Green Version]
- Linquist, B.A.; Anders, M.M.; Adviento-Borbe, M.A.A.; Chaney, R.L.; Nalley, L.L.; Da Rosa, E.F.; Kessel, C. Reducing greenhouse gas emissions, water use, and grain arsenic levels in rice systems. Glob. Chang. Biol. 2015, 21, 407–417. [Google Scholar] [CrossRef]
- Kreye, C.; Bouman, B.; Reversat, G.; Fernandez, L.; Cruz, C.V.; Elazegui, F.; Faronilo, J.; Llorca, L. Biotic and abiotic causes of yield failure in tropical aerobic rice. Field Crop. Res. 2009, 112, 97–106. [Google Scholar] [CrossRef]
- Nie, L.; Peng, S.; Chen, M.; Shah, F.; Huang, J.; Cui, K.; Xiang, J. Aerobic rice for water-saving agriculture. A review. Agron. Sustain. Dev. 2012, 32, 411–418. [Google Scholar] [CrossRef] [Green Version]
- Jones, J.D.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walling, L.L. Adaptive defense responses to pathogens and insects. Adv. Bot. Res. 2009, 51, 551–612. [Google Scholar]
- Mendy, B.; Wang’ombe, M.W.; Radakovic, Z.S.; Holbein, J.; Ilyas, M.; Chopra, D.; Holton, N.; Zipfel, C.; Grundler, F.M.W.; Siddique, S. Arabidopsis leucine-rich repeat receptor–like kinase NILR1 is required for induction of innate immunity to parasitic nematodes. PLoS Pathog. 2017, 13, e1006284. [Google Scholar] [CrossRef]
- Conrath, U. Molecular aspects of defence priming. Trends Plant Sci. 2011, 16, 524–531. [Google Scholar] [CrossRef]
- Conrath, U.; Beckers, G.J.; Langenbach, C.J.; Jaskiewicz, M.R. Priming for enhanced defense. Annu. Rev. Phytopathol. 2015, 53, 97–119. [Google Scholar] [CrossRef]
- Martinez-Medina, A.; Flors, V.; Heil, M.; Mauch-Mani, B.; Pieterse, C.M.J.; Pozo, M.J.; Ton, J.; van Dam, N.M.; Conrath, U. Recognizing plant defense priming. Trends Plant Sci. 2016, 21, 818–822. [Google Scholar] [CrossRef] [Green Version]
- Pastor, V.; Luna, E.; Mauch-Mani, B.; Ton, J.; Flors, V. Primed plants do not forget. Environ. Exp. Bot. 2013, 94, 46–56. [Google Scholar] [CrossRef]
- Walters, D.R.; Ratsep, J.; Havis, N.D. Controlling crop diseases using induced resistance: Challenges for the future. J. Exp. Bot. 2013, 64, 1263–1280. [Google Scholar] [CrossRef] [PubMed]
- Ji, P.; Yin, J.; Koné, D. Application of acibenzolar-S-methyl and standard fungicides for control of Phytophthora blight on squash. Crop Prot. 2011, 30, 1601–1605. [Google Scholar] [CrossRef]
- Wu, L.; Wu, H.; Chen, L.; Zhang, H.; Gao, X. Induction of systemic disease resistance in Nicotiana benthamiana by the cyclodipeptides cyclo (l-Pro-l-Pro) and cyclo (d-Pro-d-Pro). Mol. Plant Pathol. 2017, 18, 67–74. [Google Scholar] [CrossRef] [Green Version]
- Jung, H.W.; Tschaplinski, T.J.; Wang, L.; Glazebrook, J.; Greenberg, J.T. Priming in systemic plant immunity. Science 2009, 324, 89–91. [Google Scholar] [CrossRef]
- Singh, R.R.; Verstraeten, B.; Siddique, S.; Tegene, A.M.; Tenhaken, R.; Frei, M.; Haeck, A.; Demeestere, K.; Pokhare, S.; Gheysen, G.; et al. Ascorbate oxidation activates systemic defence against root-knot nematode Meloidogyne graminicola in rice. J. Exp. Bot. Article under Review.
- Dong, H.; Beer, S.V. Riboflavin induces disease resistance in plants by activating a novel signal transduction pathway. Phytopathology 2000, 90, 801–811. [Google Scholar] [CrossRef] [Green Version]
- Schurter, R.; Kunz, W.; Nyfeler, R. Process and A Composition for Immunizing Plants against Diseases. U.S. Patent No. 4931581, 5 June 1990. [Google Scholar]
- Knoth, C.; Salus, M.S.; Girke, T.; Eulgem, T. The synthetic elicitor 3, 5-dichloroanthranilic acid induces npr1-dependent and npr1-independent mechanisms of disease resistance in arabidopsis. Plant Physiol. 2009, 150, 333–347. [Google Scholar] [CrossRef] [Green Version]
- Schreiber, K.; Ckurshumova, W.; Peek, J.; Desveaux, D. A high-throughput chemical screen for resistance to pseudomonas syringae in arabidopsis. Plant J. 2008, 54, 522–531. [Google Scholar] [CrossRef]
- Schreiber, K.J.; Nasmith, C.G.; Allard, G.; Singh, J.; Subramaniam, R.; Desveaux, D. Found in translation: High-throughput chemical screening in arabidopsis thaliana identifies small molecules that reduce fusarium head blight disease in wheat. Mol. Plant Microbe Interact. 2011, 24, 640–648. [Google Scholar] [CrossRef]
- De Vleesschauwer, D.; Xu, J.; Höfte, M. Making sense of hormone-mediated defense networking: From rice to Arabidopsis. Front. Plant Sci. 2014, 5, 611. [Google Scholar] [CrossRef] [PubMed]
- Hampp, C.; Richter, A.; Osorio, S.; Zellnig, G.; Sinha, A.K.; Jammer, A.; Fernie, A.R.; Grimm, B.; Roitsch, T. Establishment of a photoautotrophic cell suspension culture of arabidopsis thaliana for photosynthetic, metabolic, and signaling studies. Mol. Plant 2012, 5, 524–527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mustafa, N.R.; De Winter, W.; Van Iren, F.; Verpoorte, R. Initiation, growth and cryopreservation of plant cell suspension cultures. Nat. Protoc. 2011, 6, 715–742. [Google Scholar] [CrossRef] [PubMed]
- Segečová, A.; Cervený, J.; Roitsch, T. Stress response monitoring of photoautotrophic higher plant suspension cultures by fluorescence imaging for high-throughput toxic compound screening. J. Environ. Prot. 2017, 8, 678–692. [Google Scholar] [CrossRef] [Green Version]
- Noutoshi, Y.; Ikeda, M.; Shirasu, K. Diuretics prime plant immunity in arabidopsis thaliana. PLoS ONE 2012, 7, e48443. [Google Scholar] [CrossRef]
- Noutoshi, Y.; Jikumaru, Y.; Kamiya, Y.; Shirasu, K. Imprimatinc1, a novel plant immune-priming compound, functions as a partial agonist of salicylic acid. Sci. Rep. 2012, 2, 705. [Google Scholar] [CrossRef] [Green Version]
- Noutoshi, Y.; Okazaki, M.; Kida, T.; Nishina, Y.; Morishita, Y.; Ogawa, T.; Suzuki, H.; Shibata, D.; Jikumaru, Y.; Hanada, A.; et al. Novel plant immune-priming compounds identified via highthroughput chemical screening target salicylic acid glucosyltransferases in arabidopsis. Plant Cell 2012, 24, 3795–3804. [Google Scholar] [CrossRef] [Green Version]
- Mauch-Mani, B.; Baccelli, I.; Luna, E.; Flors, V. Defense priming: An adaptive part of induced resistance. Annu. Rev. Plant Biol. 2017, 68, 485–512. [Google Scholar] [CrossRef] [Green Version]
- Ji, H.; Kyndt, T.; He, W.; Vanholme, B.; Gheysen, G. β-Aminobutyric acid–induced resistance against root-knot nematodes in rice is based on increased basal defense. Mol. Plant Microbe Interact. 2015, 28, 519–533. [Google Scholar] [CrossRef] [Green Version]
- Singh, P.; Kuo, Y.C.; Mishra, S.; Tsai, C.H.; Chien, C.C.; Chen, C.W.; Desclos-Theveniau, M.; Chu, P.W.; Schulze, B.; Chinchilla, D.; et al. The lectin receptor kinase-VI. 2 is required for priming and positively regulates Arabidopsis pattern-triggered immunity. Plant Cell 2012, 24, 1256–1270. [Google Scholar] [CrossRef] [Green Version]
- Po-Wen, C.; Singh, P.; Zimmerli, L. Priming of the Arabidopsis pattern-triggered immunity response upon infection by necrotrophic Pectobacterium carotovorum bacteria. Mol. Plant Pathol. 2013, 14, 58–70. [Google Scholar] [CrossRef] [PubMed]
- Fuller, T.; Langfelder, P.; Presson, A.; Horvath, S. Review of weighted gene coexpression network analysis. In Handbook of Statistical Bioinformatics; Lu, H.H.-S., Schölkopf, B., Zhao, H., Eds.; Springer: New York City, NY, USA, 2011; pp. 369–388. [Google Scholar]
- Langfelder, P.; Horvath, S. WGCNA: An R package for weighted correlation network analysis. BMC Bioinform. 2008, 9, 559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, B.; Horvath, S. A general framework for weighted gene co-expression network analysis. Stat. Appl. Genet. Mol. Biol. 2005, 4. [Google Scholar] [CrossRef] [PubMed]
- Amrine, K.C.; Blanco-Ulate, B.; Cantu, D. Discovery of core biotic stress responsive genes in arabidopsis by weighted gene co-expression network analysis. PLoS ONE 2015, 10, e0118731. [Google Scholar] [CrossRef] [Green Version]
- Geer, L.Y.; Marchler-Bauer, A.; Geer, R.C.; Han, L.; He, J.; He, S.; Liu, C.; Shi, W.; Bryant, S.H. The NCBI biosystems database. Nucleic Acids Res. 2009, 38 (Suppl. S1), D492–D496. [Google Scholar] [CrossRef] [Green Version]
- Sakai, H.; Lee, S.S.; Tanaka, T.; Numa, H.; Kim, J.; Kawahara, Y.; Wakimoto, H.; Yang, C.C.; Iwamoto, M.; Abe, T.; et al. Rice annotation project database (rap-db): An integrative and interactive database for rice genomics. Plant Cell Physiol. 2013, 54, e6. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT–PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Li, B.; Meng, X.; Shan, L.; He, P. Transcriptional regulation of pattern-triggered immunity in plants. Cell Host Microbe 2016, 19, 641–650. [Google Scholar] [CrossRef] [Green Version]
- Sun, H.; Li, Y.; Feng, S.; Zou, W.; Guo, K.; Fan, C.; Si, S.; Peng, L. Analysis of five rice 4-coumarate: Coenzyme A ligase enzyme activity and stress response for potential roles in lignin and flavonoid biosynthesis in rice. Biochem. Biophys. Res. Commun. 2013, 430, 1151–1156. [Google Scholar] [CrossRef]
- Liao, H.; Xiao, X.; Li, X.; Chen, Y.; Fu, X.; Lin, D.; Niu, X.; He, C. OsBAK1 is involved in rice resistance to Xanthomonas oryzae pv. oryzae PXO99. Plant Biotechnol. Rep. 2016, 10, 75–82. [Google Scholar] [CrossRef]
- Mackey, D.; Belkhadir, Y.; Alonso, J.M.; Ecker, J.R.; Dangl, J.L. Arabidopsis RIN4 is a target of the type III virulence effector AvrRpt2 and modulates RPS2-mediated resistance. Cell 2003, 112, 379–389. [Google Scholar] [CrossRef] [Green Version]
- Nahar, K.; Kyndt, T.; De Vleesschauwer, D.; Höfte, M.; Gheysen, G. The jasmonate pathway is a key player in systemically induced defense against root knot nematodes in rice. Plant Physiol. 2011, 157, 305–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Šašek, V.; Nováková, M.; Dobrev, P.I.; Valentová, O.; Burketová, L. β-aminobutyric acid protects Brassica napus plants from infection by Leptosphaeria maculans. Resistance induction or a direct antifungal effect? Eur. J. Plant Pathol. 2012, 133, 279–289. [Google Scholar] [CrossRef]
- Van Hulten, M.; Pelser, M.; Van Loon, L.C.; Pieterse, C.M.; Ton, J. Costs and benefits of priming for defense in Arabidopsis. Proc. Natl. Acad. Sci. USA 2006, 103, 5602–5607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hillocks, R.J. Farming with fewer pesticides: Eu pesticide review and resulting challenges for uk agriculture. Crop Prot. 2012, 31, 85–93. [Google Scholar] [CrossRef]
- Eijssen, L.M.; Jaillard, M.; Adriaens, M.E.; Gaj, S.; de Groot, P.J.; Müller, M.; Evelo, C.T. User-friendly solutions for microarray quality control and pre-processing on arrayanalysis.org. Nucleic Acids Res. 2013, 41, W71–W76. [Google Scholar] [CrossRef] [Green Version]
- Pepper, S.D.; Saunders, E.K.; Edwards, L.E.; Wilson, C.L.; Miller, C.J. The utility of mas5 expression summary and detection call algorithms. BMC Bioinform. 2007, 8, 273. [Google Scholar] [CrossRef] [Green Version]
- Irizarry, R.A.; Bolstad, B.M.; Collin, F.; Cope, L.M.; Hobbs, B.; Speed, T.P. Summaries of affymetrix genechip probe level data. Nucleic Acids Res. 2003, 31, e15. [Google Scholar] [CrossRef]
- Leek, J.T.; Johnson, W.E.; Parker, H.S.; Jaffe, A.E.; Storey, J.D. The sva package for removing batch effects and other unwanted variation in high-throughput experiments. Bioinformatics 2012, 28, 882–883. [Google Scholar] [CrossRef]
- Gentleman, R.; Carey, V.; Huber, W.; Hahne, F. Genefilter: Methods for Filtering Genes from High-Throughput Experiments. R Package Version 1.62.0; Available online: https://bioconductor.org/packages/release/bioc/html/genefilter.html (accessed on 28 December 2019).
- Horvath, S. Correlation and Gene Co-Expression Networks. In Weighted Network Analysis—Applications in Genomics and Systems Biology, 1st ed.; Springer: New York, NY, USA, 2011; pp. 91–121. [Google Scholar]
- Reimand, J.; Arak, T.; Adler, P.; Kolberg, L.; Reisberg, S.; Peterson, H.; Vilo, J. g: Profiler—A web server for functional interpretation of gene lists. Nucleic Acids Res. 2016, 39, W307–W315. [Google Scholar] [CrossRef] [Green Version]
- Kawahara, Y.; de la Bastide, M.; Hamilton, J.P.; Kanamori, H.; McCombie, W.R.; Ouyang, S.; Schwartz, D.C.; Tanaka, T.; Wu, J.; Zhou, S.; et al. Improvement of the oryza sativa nipponbare reference genome using next generation sequence and optical map data. Rice 2013, 6, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, F.; Breitling, R.; McEntee, C.W.; Wittner, B.S.; Nemhauser, J.L.; Chory, J. Rankprod: A bioconductor package for detecting differentially expressed genes in meta-analysis. Bioinformatics 2006, 22, 2825–2827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kersey, P.J.; Allen, J.E.; Armean, I.; Boddu, S.; Bolt, B.J.; CarvalhoSilva, D.; Christensen, M.; Davis, P.; Falin, L.J.; Grabmueller, C.; et al. Ensembl genomes 2016: More genomes, more complexity. Nucleic Acids Res. 2015, 44, D574–D580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sander, C.; Schneider, R. Database of homology-derived protein structures and the structural meaning of sequence alignment. Proteins Struct. Funct. Bioinform. 1991, 9, 56–68. [Google Scholar] [CrossRef]
- Reversat, G.; Boyer, J.; Sannier, C.; Pando-Bahuon, A. Use of a mixture of sand and water-absorbent synthetic polymer as substrate for the xenic culturing of plant-parasitic nematodes in the laboratory. Nematology 1999, 1, 209–212. [Google Scholar]
- Hellemans, J.; Mortier, G.; De Paepe, A.; Speleman, F.; Vandesompele, J. qbase relative quantification framework and software for management and automated analysis of real-time quantitative pcr data. Genome Biol. 2007, 8, R19. [Google Scholar] [CrossRef] [Green Version]
- Campbell, J.; Lin, Q.; Geske, G.D.; Blackwell, H.E. New and unexpected insights into the modulation of LuxR-type quorum sensing by cyclic dipeptides. ACS Chem. Biol. 2009, 4, 1051–1059. [Google Scholar] [CrossRef]


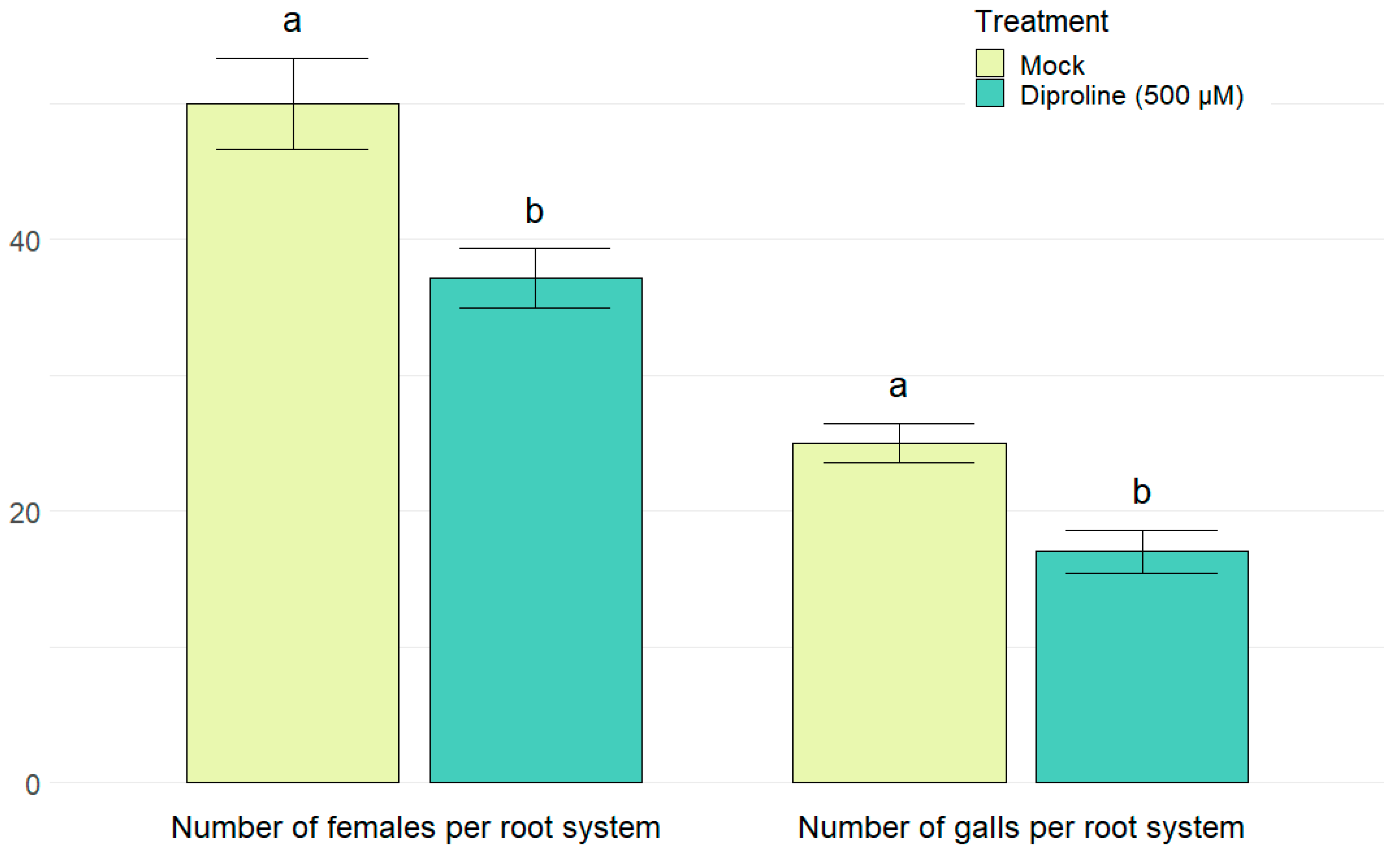


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Name | Annotation |
|---|---|---|
| Os08g0448000 | Os4CL5 | 4-coumarate–CoA ligase 5 |
| Os03g0440900 | OsBAK1 | BRASSINOSTEROID INSENSITIVE 1-associated receptor kinase 1 |
| Os03g0804500 | OsGER7 | Germin-like protein 3-7 |
| Os08g0508800 | OsLOX7 | Lipoxygenase 7, chloroplastic |
| Os01g0963000 | OsPRX22 | Similar to Peroxidase BP 1 precursor |
| Os02g0738200 | OsPUB40 | Zinc finger, RING/FYVE/PHD-type domain containing protein |
| Os04g0379600 | OsRIN4 | Rin4 domain containing protein |
| Os02g0251900 | OsVQ7 | Similar to Tobacco rattle virus-induced protein variant 2 |
| Os03g0568500 | UNKN7 | Uncharacterized protein family UPF0136 |
| Os07g0506000 | UNKN9 | Protein of unknown function DUF300 family protein |
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 |
|---|---|---|---|---|---|---|---|---|---|---|---|
| In Vitro | In Planta | In Silico | |||||||||
| Dipro-line | Acti-gard® | Ribo-flavin | Azelaic acid | DHA | LPS | NW | LPS | NW | (Glc-NAc)7 | ||
| Os4CL5 | 1.860 | 1.664 | 1.809 | 1.075 | 2.166 | 3.249 | 2.878 | 1.359 | 2.172 | 1.720 | 1.774 |
| OsBAK1 | 1.329 | 1.434 | 1.454 | 1.288 | 1.688 | 1.434 | 1.532 | 1.200 | 3.270 | 1.334 | 1.208 |
| OsPUB40 | 1.479 | 1.157 | 1.338 | 1.102 | 5.918 | 2.071 | 2.403 | 2.203 | 4.539 | 3.687 | 1.853 |
| OsRIN4 | 1.602 | 1.778 | 1.516 | 1.053 | 0.34 | 2.250 | 1.753 | 1.627 | 3.051 | 2.098 | 1.401 |
| UNKN7 | 1.288 | 1.454 | 1.094 | 1.106 | 1.821 | 1.803 | 1.117 | 1.525 | 2.507 | 1.671 | 1.244 |
| UNKN9 | 1.253 | 1.333 | 1.261 | 1.121 | 0.809 | 1.553 | 1.021 | 1.579 | 1.183 | 1.520 | 1.129 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Kesel, J.; Gómez-Rodríguez, R.; Bonneure, E.; Mangelinckx, S.; Kyndt, T. The Use of PTI-Marker Genes to Identify Novel Compounds that Establish Induced Resistance in Rice. Int. J. Mol. Sci. 2020, 21, 317. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21010317
De Kesel J, Gómez-Rodríguez R, Bonneure E, Mangelinckx S, Kyndt T. The Use of PTI-Marker Genes to Identify Novel Compounds that Establish Induced Resistance in Rice. International Journal of Molecular Sciences. 2020; 21(1):317. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21010317
Chicago/Turabian StyleDe Kesel, Jonas, Ramsés Gómez-Rodríguez, Eli Bonneure, Sven Mangelinckx, and Tina Kyndt. 2020. "The Use of PTI-Marker Genes to Identify Novel Compounds that Establish Induced Resistance in Rice" International Journal of Molecular Sciences 21, no. 1: 317. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21010317