Nobiletin Delays Aging and Enhances Stress Resistance of Caenorhabditis elegans


Abstract
:1. Introduction
2. Results
2.1. Nobiletin (NOB) Increased Lifespan of C. elegans
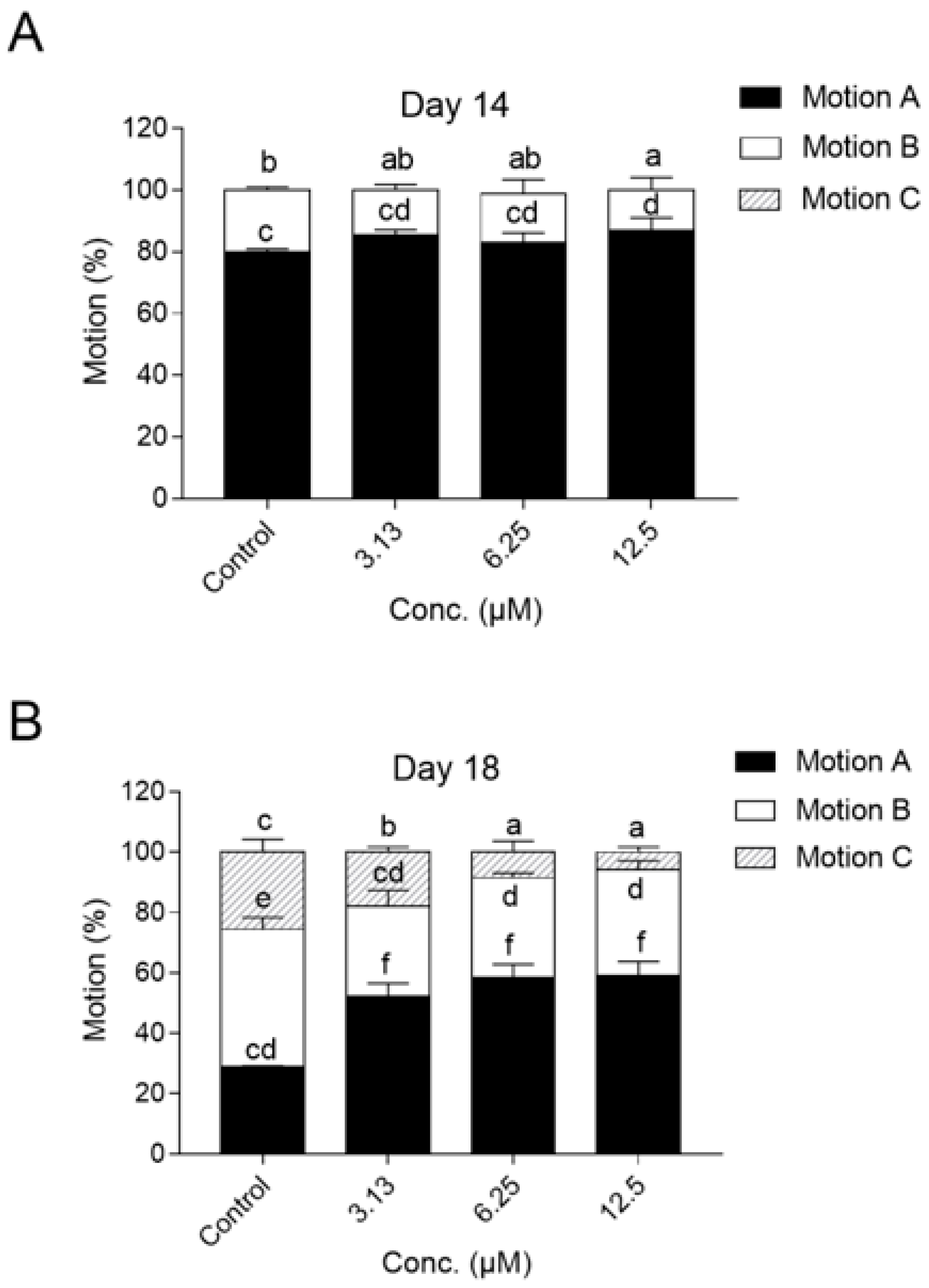
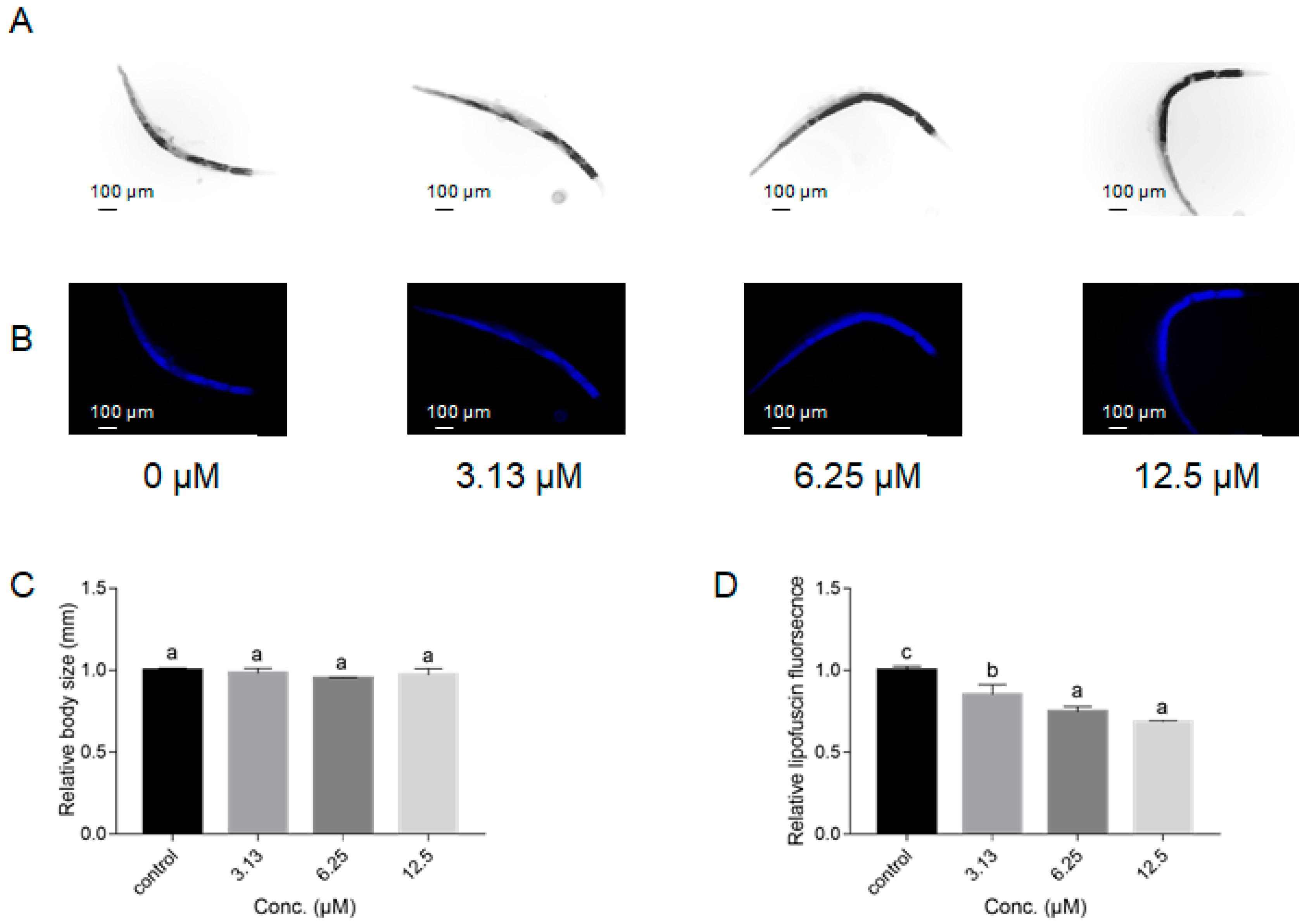
2.2. NOB Delayed Aging-Related Functional Decline of C. elegans
2.3. NOB Had No Significant Effect on Fertility
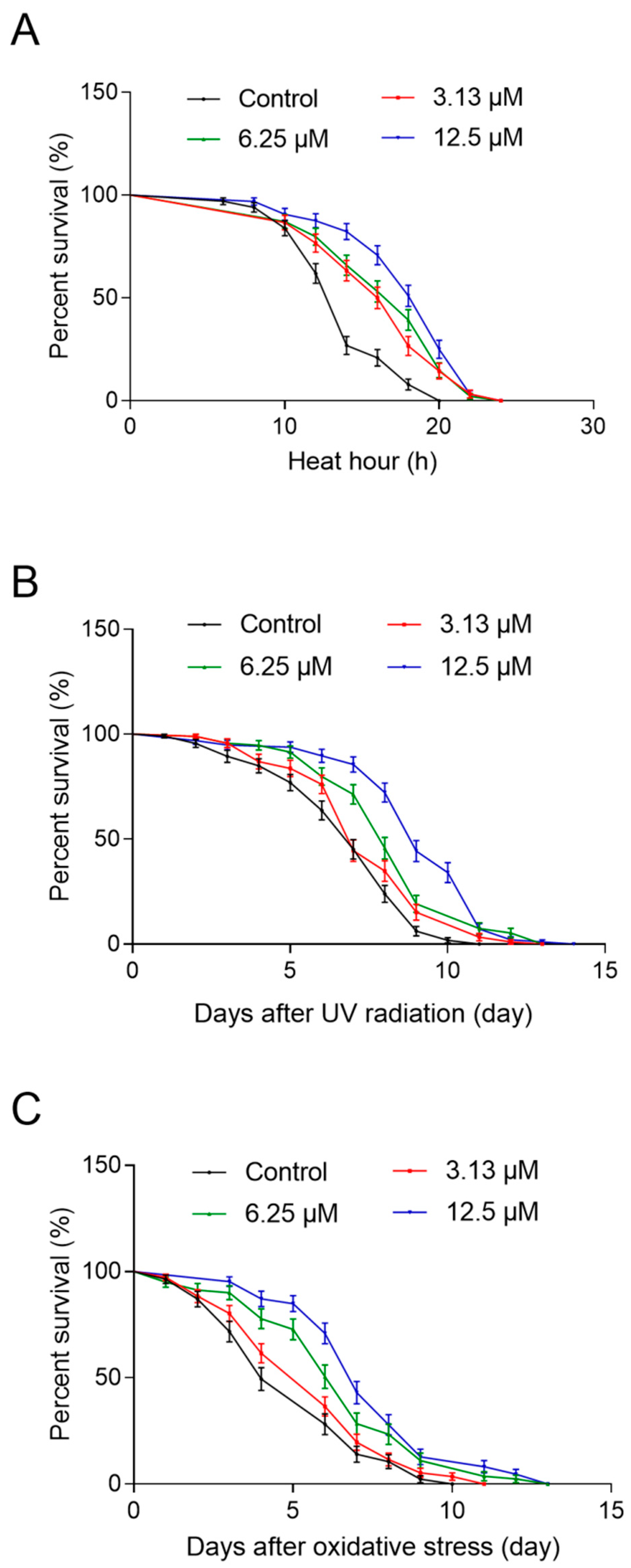
2.4. NOB Increased Resistance to Heat Shock, UV Radiation, and Oxidative Stress
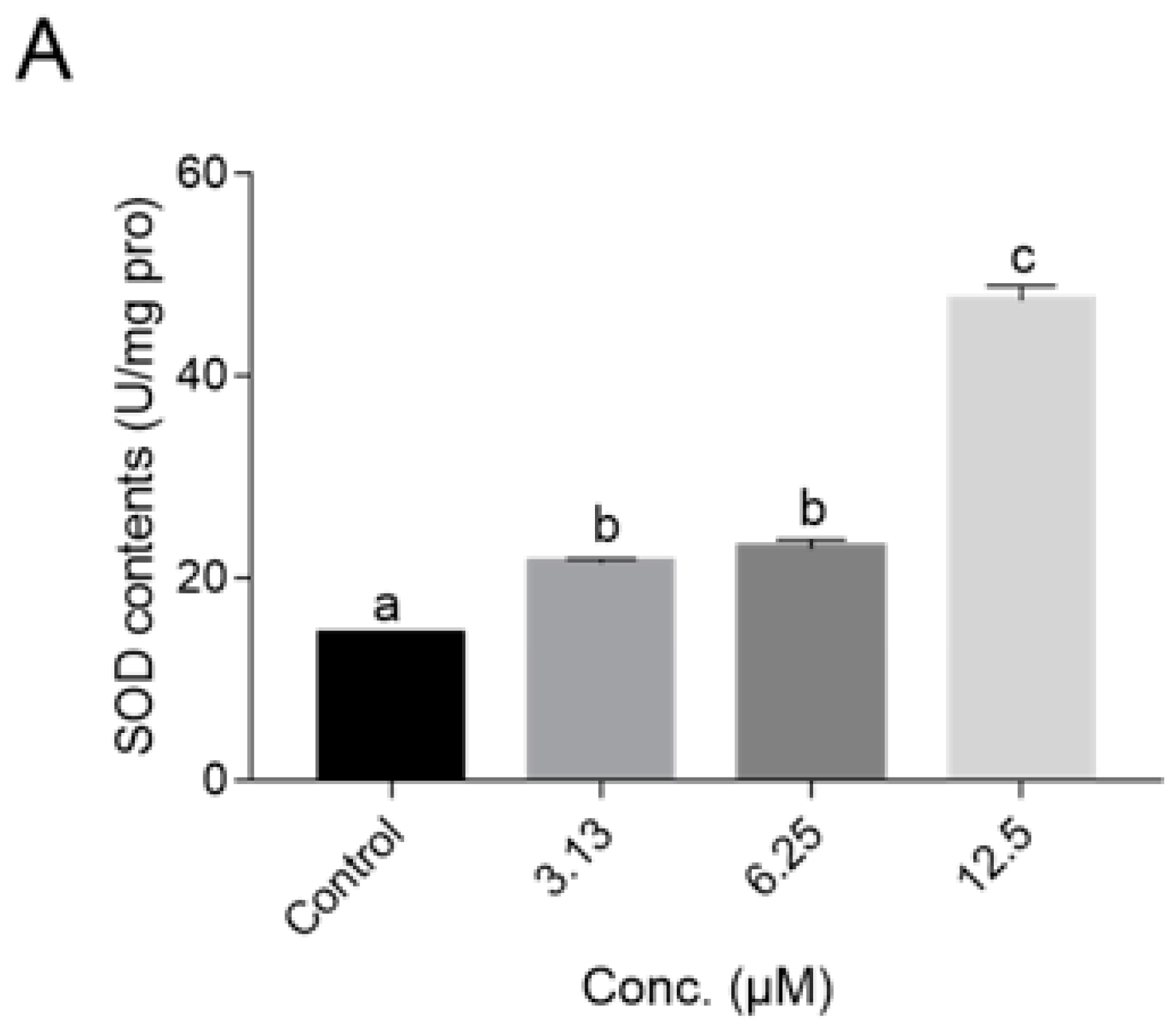
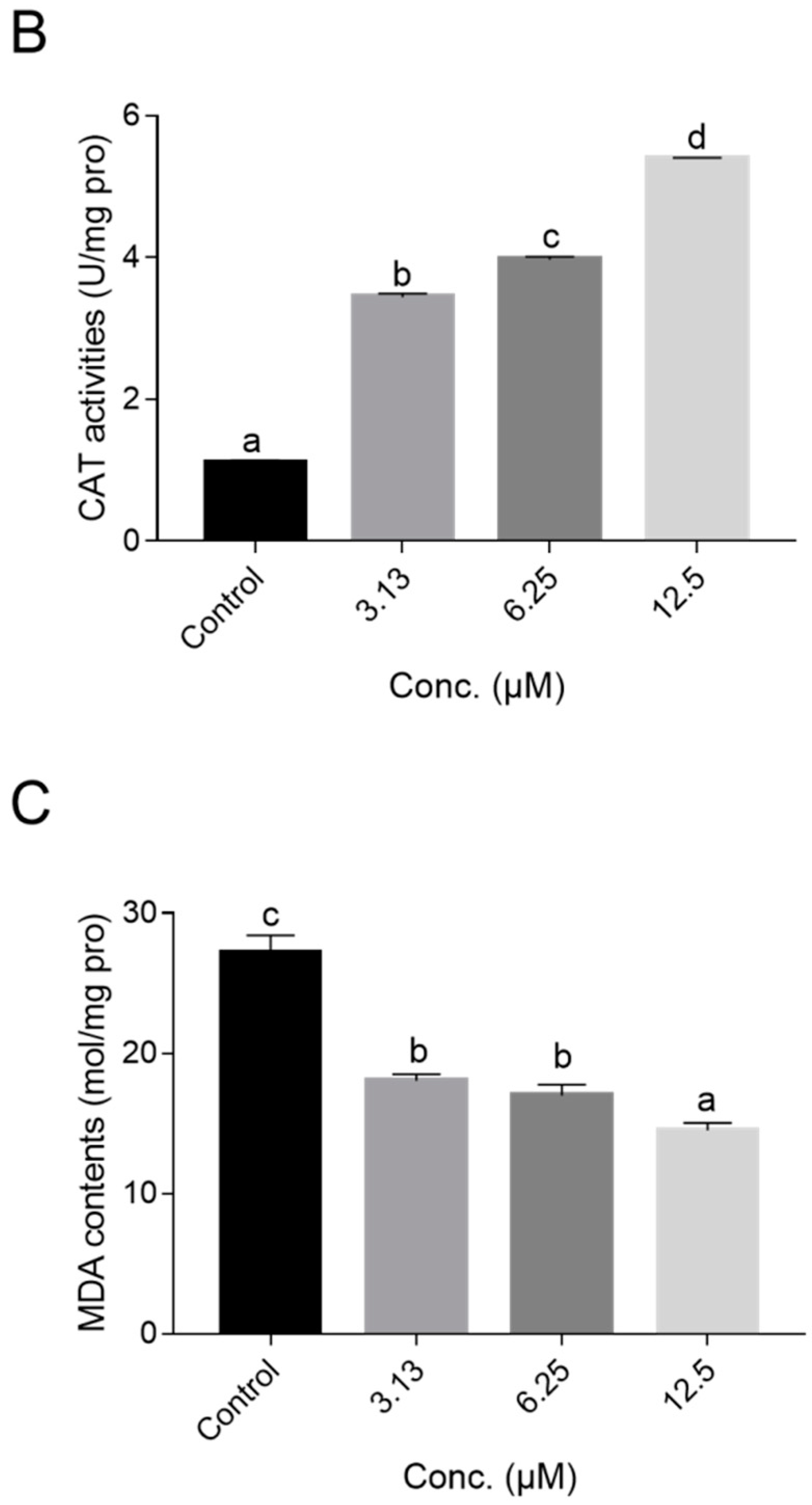
2.5. NOB Decreased ROS Accumulation In Vivo and Increased Antioxidant Enzyme Activities
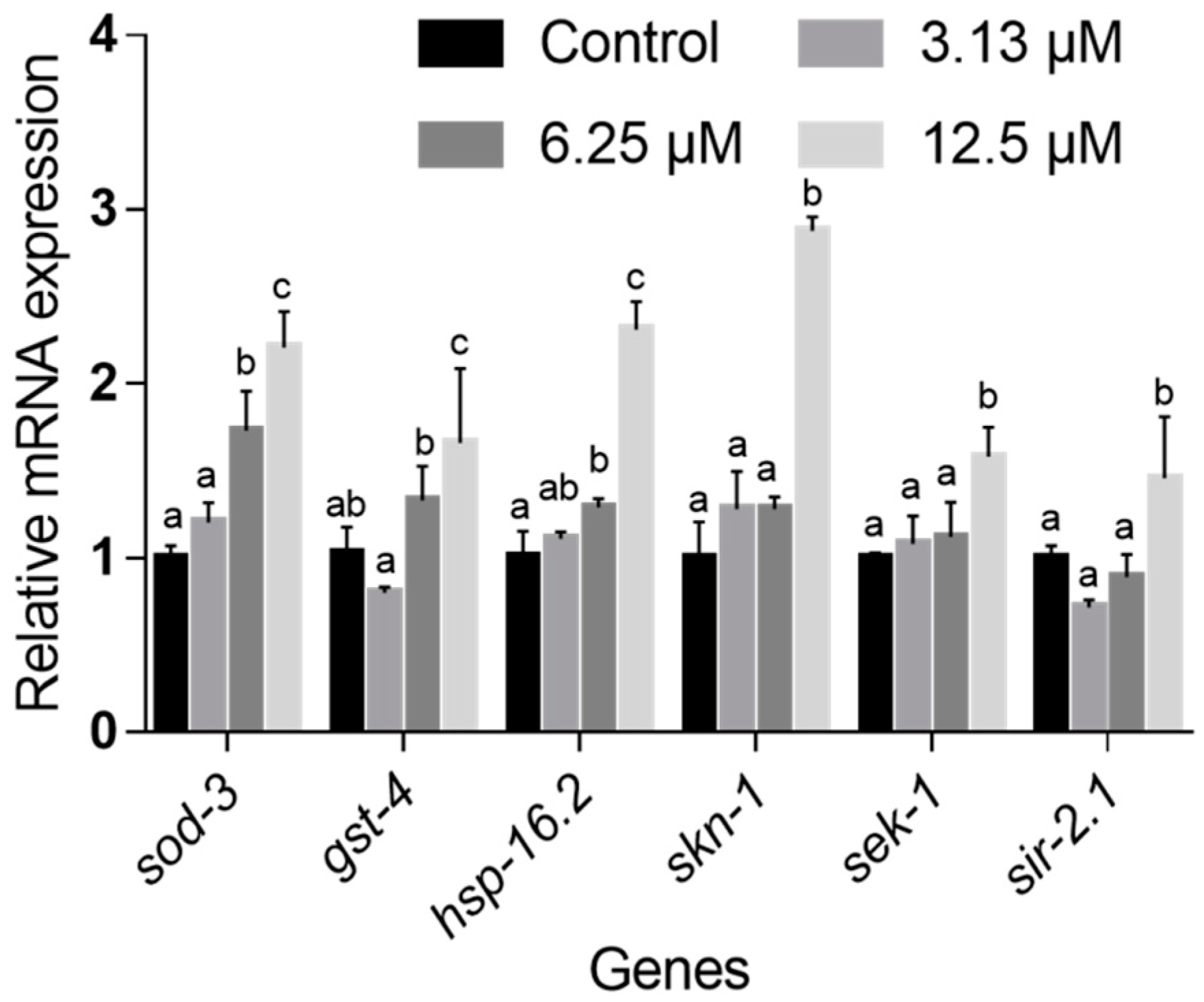
2.6. NOB Regulated the Expression Levels of Genes Involved in Life Extension and Stress Resistance
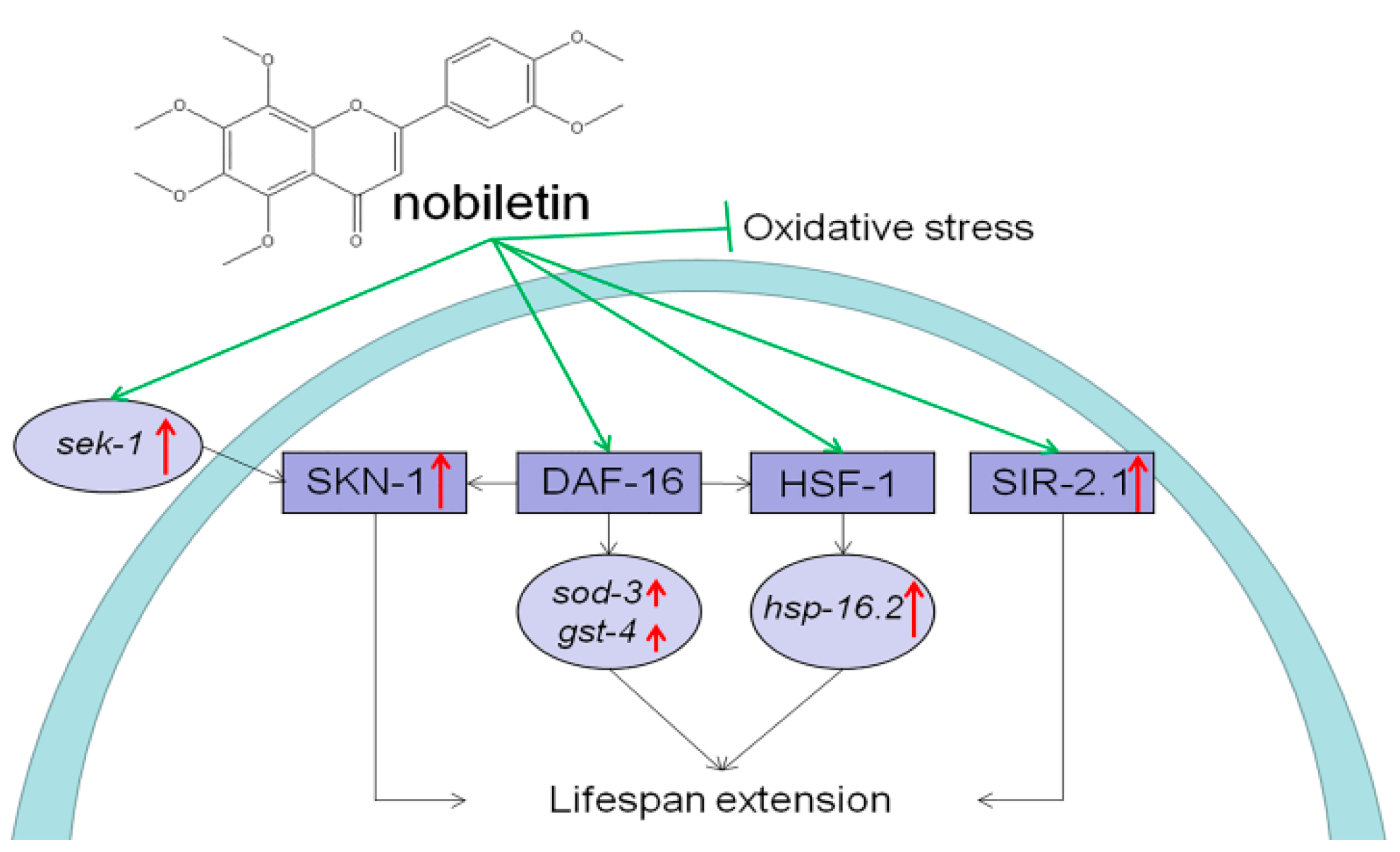
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Nematode Strains and Culture Conditions
4.3. Lifespan Assay
4.4. Aging-Related Phenotypic Analysis
4.4.1. Motility Analysis
4.4.2. Length and Lipofuscin Levels Alterations
4.5. Oviposition Measurement
4.6. Stress Resistance Assays
4.6.1. Heat Shock
4.6.2. UV Radiation
4.6.3. Oxidative Stress
4.7. Intracellular ROS Measurement and Detection of Antioxidant Enzyme Activity
4.8. Reverse Transcription-Quantitative PCR (RT-qPCR)
4.9. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- World Population Prospects 2019. Available online: http://population.un.org/wpp/ (accessed on 28 August 2019).
- Kris-Etherton, P.M.; Hecker, K.D.; Bonanome, A.; Coval, S.M.; Binkoski, A.E.; Hilpert, K.F.; Griel, A.E.; Etherton, T.D. Bioactive compounds in foods: Their role in the prevention of cardiovascular disease and cancer. Am. J. Med. 2002, 113, 71–88. [Google Scholar] [CrossRef]
- Joseph, J.A.; Shukitt-Hale, B.; Casadesus, G. Reversing the deleterious effects of aging on neuronal communication and behavior: Beneficial properties of fruit polyphenolic compounds. Am. J. Clin. Nutr. 2005, 81, 313s–316s. [Google Scholar] [CrossRef] [PubMed]
- Brownson, D.M.; Azios, N.G.; Fuqua, B.K.; Dharmawardhane, S.F.; Mabry, T.J. Flavonoid effects relevant to cancer. J. Nutr. 2002, 132, 3482s–3489s. [Google Scholar] [CrossRef] [PubMed]
- Nurk, E.; Refsum, H.; Drevon, C.A.; Tell, G.S.; Nygaard, H.A.; Engedal, K.; Smith, A.D. Intake of flavonoid-rich wine, tea, and chocolate by elderly men and women is associated with better cognitive test performance. J. Nutr. 2009, 139, 120–127. [Google Scholar] [CrossRef]
- Herndon, L.A.; Schmeissner, P.J.; Dudaronek, J.M.; Brown, P.A.; Listner, K.M.; Sakano, Y.; Paupard, M.C.; Hall, D.H.; Driscoll, M. Stochastic and genetic factors influence tissue-specific decline in ageing C-elegans. Nature 2002, 419, 808–814. [Google Scholar] [CrossRef]
- Kenyon, C.J. The genetics of ageing. Nature 2010, 467, 622. [Google Scholar] [CrossRef] [Green Version]
- Dillin, A.; Hsu, A.L.; Arantes-Oliveira, N.A.; Lehrer-Graiwer, J.; Hsin, H.; Fraser, A.G.; Kamath, R.S.; Ahringer, J.; Kenyon, C. Rates of behavior and aging specified by mitochondrial function during development. Science 2002, 298, 2398–2401. [Google Scholar] [CrossRef]
- Honda, Y.; Honda, S. The daf-2 gene network for longevity regulates oxidative stress resistance and Mn-superoxide dismutase gene expression in Caenorhabditis elegans. FASEB J. 1999, 13, 1385–1393. [Google Scholar] [CrossRef] [Green Version]
- Tatar, M.; Bartke, A.; Antebi, A. The endocrine regulation of aging by insulin-like signals. Science 2003, 299, 1346–1351. [Google Scholar] [CrossRef]
- Murphy, C.T.; McCarroll, S.A.; Bargmann, C.I.; Fraser, A.; Kamath, R.S.; Ahringer, J.; Li, H.; Kenyon, C. Genes that act downstream of DAF-16 to influence the lifespan of Caenorhabditis elegans. Nature 2003, 424, 277–284. [Google Scholar] [CrossRef]
- Hsu, A.L. Regulation of aging and age-related disease by DAF-16 and heat-shock factor. Science 2003, 300, 2033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tullet, J.M.A.; Hertweck, M.; An, J.H.; Baker, J.; Hwang, J.Y.; Liu, S.; Oliveira, R.P.; Baumeister, R.; Blackwell, T.K. Direct inhibition of the longevity-promoting factor SKN-1 by insulin-like signaling in C. elegans. Cell 2008, 132, 1025–1038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kampkotter, A.; Timpel, C.; Zurawski, R.F.; Ruhl, S.; Chovolou, Y.; Proksch, P.; Watjen, W. Increase of stress resistance and lifespan of Caenorhabditis elegans by quercetin. Comp. Biochem. Physiol. 2008, 149, 314–323. [Google Scholar] [CrossRef] [PubMed]
- Saul, N.; Pietsch, K.; Menzel, R.; Steinberg, C.E.W. Quercetin-mediated longevity in Caenorhabditis elegans: Is DAF-16 involved? Mech. Ageing Dev. 2008, 129, 611–613. [Google Scholar] [CrossRef] [PubMed]
- Buchter, C.; Ackermann, D.; Havermann, S.; Honnen, S.; Chovolou, Y.; Fritz, G.; Kampkotter, A.; Watjen, W. Myricetin-mediated lifespan extension in Caenorhabditis elegans is modulated by DAF-16. Int. J. Mol. Sci. 2013, 14, 11895–11914. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.L.; Liu, J.; Li, T.; Liu, R.H. Blueberry extract promotes longevity and stress tolerance via DAF-16 in Caenorhabditis elegans. Food Funct. 2018, 9, 5273–5282. [Google Scholar] [CrossRef]
- Caland, R.B.D.; Cadavid, C.O.M.; Carmona, L.; Pena, L.; Oliveira, R.D. Pasteurized orange juice rich in carotenoids protects Caenorhabditis elegans against oxidative stress and beta-amyloid toxicity through direct and indirect mechanisms. Oxidative Med. Cell Longev. 2019, 2019, 5046280. [Google Scholar] [CrossRef] [Green Version]
- Tripoli, E.; La Guardia, M.; Giammanco, S.; Di Majo, D.; Giammanco, M. Citrus flavonoids: Molecular structure, biological activity and nutritional properties: A review. Food Chem. 2007, 104, 466–479. [Google Scholar] [CrossRef]
- Di Donna, L.; Taverna, D.; Mazzotti, F.; Benabdelkamel, H.; Attya, M.; Napoli, A.; Sindona, G. Comprehensive assay of flavanones in citrus juices and beverages by UHPLC-ESI-MS/MS and derivatization chemistry. Food Chem. 2013, 141, 2328–2333. [Google Scholar] [CrossRef]
- Zhao, Z.Y.; He, S.S.; Hu, Y.; Yang, Y.; Jiao, B.N.; Fang, Q.; Zhou, Z.Q. Fruit flavonoid variation between and within four cultivated Citrus species evaluated by UPLC-PDA system. Sci. Hortic. 2017, 224, 93–101. [Google Scholar] [CrossRef]
- Yasuda, T.; Yoshimura, Y.; Yabuki, H.; Nakazawa, T.; Ohsawa, K.; Mimaki, Y.; Sashida, Y. Urinary metabolites of nobiletin orally administered to rats. Chem. Pharm. Bull. 2003, 51, 1426–1428. [Google Scholar] [CrossRef] [Green Version]
- Zheng, J.K.; Song, M.Y.; Dong, P.; Qiu, P.J.; Guo, S.S.; Zhong, Z.M.; Li, S.M.; Ho, C.T.; Xiao, H. Identification of novel bioactive metabolites of 5-demethylnobiletin in mice. Mol. Nutr. Food Res. 2013, 57, 1999–2007. [Google Scholar] [CrossRef] [PubMed]
- Li, S.M.; Wang, Z.Y.; Sang, S.M.; Huang, M.T.; Ho, C.T. Identification of nobiletin metabolites in mouse urine. Mol. Nutr. Food Res. 2006, 50, 291–299. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, A.; Ohizumi, Y. Potential benefits of nobiletin, a citrus flavonoid, against Alzheimer’s Disease and Parkinson’s Disease. Int. J. Mol. Sci. 2019, 20, 3380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, G.Y.; Mi, Y.S.; Fan, R.; Li, R.N.; Liu, Z.G.; Liu, X.B. Nobiletin protects against systemic inflammation-stimulated memory impairment via MAPK and NF-kappa B signaling pathway. J. Agric. Food Chem. 2019, 67, 5122–5134. [Google Scholar] [CrossRef] [PubMed]
- Nohara, K.; Mallampalli, V.; Nemkov, T.; Wirianto, M.; Yang, J.; Ye, Y.Q.; Sun, Y.X.; Han, L.; Esser, K.A.; Mileykovskaya, E.; et al. Nobiletin fortifies mitochondrial respiration in skeletal muscle to promote healthy aging against metabolic challenge. Nat. Commun. 2019, 10, 3923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldstein, B. Sydney brenner on the genetics of Caenorhabditis elegans. Genetics 2016, 204, 1–2. [Google Scholar] [CrossRef]
- Wilson, M.A.; Shukitt-Hale, B.; Kalt, W.; Ingram, D.K.; Joseph, J.A.; Wolkow, C.A. Blueberry polyphenols increase lifespan and thermotolerance in Caenorhabditis elegans. Aging Cell 2006, 5, 59–68. [Google Scholar] [CrossRef] [Green Version]
- Zhou, L.; Wang, L.; Zhang, J.L.; Li, J.H.; Bai, S.J.; Ma, J.F.; Fu, X.Q. Didymin improves UV irradiation resistance in C. elegans. PeerJ 2019, 6, e6218. [Google Scholar] [CrossRef]
- Pereira, M.D.; Herdeiro, R.S.; Fernandes, P.N.; Eleutherio, E.C.A.; Panek, A.D. Targets of oxidative stress in yeast sod mutants. BBA-Gen. Subj. 2003, 1620, 245–251. [Google Scholar] [CrossRef]
- Wolfe, K.L.; Liu, R.H. Cellular antioxidant activity (CAA) assay for assessing antioxidants, foods, and dietary supplements. J. Agric. Food Chem. 2007, 55, 8896–8907. [Google Scholar] [CrossRef] [PubMed]
- Melov, S.; Ravenscroft, J.; Malik, S.; Gill, M.S.; Walker, D.W.; Clayton, P.E.; Wallace, D.C.; Malfroy, B.; Doctrow, S.R.; Lithgow, G.J. Extension of life-span with superoxide dismutase/catalase mimetics. Science 2000, 289, 1567–1569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rea, S.L.; Wu, D.Q.; Cypser, J.R.; Vaupel, J.W.; Johnson, T.E. A stress-sensitive reporter predicts longevity in isogenic populations of Caenorhabditis elegans. Nat. Genet. 2005, 37, 894–898. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, A.; Ohizumi, Y.; Yamada, K. Anti-dementia activity of nobiletin, a citrus flavonoid: A review of animal studies. Clin. Psychopharmacol. Neurosci. 2014, 12, 75–82. [Google Scholar] [CrossRef] [Green Version]
- Kimura, J.; Shimizu, K.; Kajima, K.; Yokosuka, A.; Mimaki, Y.; Oku, N.; Ohizumi, Y. Nobiletin reduces intracellular and extracellular beta-amyloid in iPS cell-derived Alzheimer’s Disease model neurons. Biol. Pharm. Bull. 2018, 41, 451–457. [Google Scholar] [CrossRef] [Green Version]
- Yao, X.L.; Zhu, X.R.; Pan, S.Y.; Fang, Y.P.; Jiang, F.T.; Phillips, G.O.; Xu, X.Y. Antimicrobial activity of nobiletin and tangeretin against Pseudomonas. Food Chem. 2012, 132, 1883–1890. [Google Scholar] [CrossRef]
- Morley, K.L.; Ferguson, P.J.; Koropatnick, J. Tangeretin and nobiletin induce G1 cell cycle arrest but not apoptosis in human breast and colon cancer cells. Cancer Lett. 2007, 251, 168–178. [Google Scholar] [CrossRef]
- Yasuda, N.; Ishii, T.; Oyama, D.; Fukuta, T.; Agato, Y.; Sato, A.; Shimizu, K.; Asai, T.; Asakawa, T.; Kan, T.; et al. Neuroprotective effect of nobiletin on cerebral ischemia-reperfusion injury in transient middle cerebral artery-occluded rats. Brain Res. 2014, 1559, 46–54. [Google Scholar] [CrossRef]
- Liu, E.H.; Zhao, P.; Duan, L.; Zheng, G.D.; Guo, L.; Yang, H.; Li, P. Simultaneous determination of six bioactive flavonoids in citri reticulatae pericarpium by rapid resolution liquid chromatography coupled with triple quadrupole electrospray tandem mass spectrometry. Food Chem. 2013, 141, 3977–3983. [Google Scholar] [CrossRef]
- Nohara, K.; Nemkov, T.; D’Alessandro, A.; Yoo, S.H.; Chen, Z. Coordinate Regulation of Cholesterol and Bile Acid Metabolism by the Clock Modifier Nobiletin in Metabolically Challenged Old Mice. Int. J. Mol. Sci. 2019, 20, 4821. [Google Scholar] [CrossRef] [Green Version]
- Grunz, G.; Haas, K.; Soukup, S.; Klingenspor, M.; Kulling, S.E.; Daniel, H.; Spanier, B. Structural features and bioavailability of four flavonoids and their implications for lifespan-extending and antioxidant actions in C. elegans. Mech. Ageing Dev. 2012, 133, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Goh, J.X.H.; Tan, L.T.H.; Goh, J.K.; Chan, K.G.; Pusparajah, P.; Lee, L.H.; Goh, B.H. Nobiletin and Derivatives: Functional Compounds from Citrus Fruit Peel for Colon Cancer Chemoprevention. Cancers 2019, 11, 867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delaney, B.; Phillips, K.; Vasquez, C.; Wilson, A.; Cox, D.; Wang, H.B.; Manthey, J. Genetic toxicity of a standardized mixture of citrus polymethoxylated flavones. Food Chem. Toxicol. 2002, 40, 617–624. [Google Scholar] [CrossRef]
- Kirkwood, T.B.L. Understanding the odd science of aging. Cell 2005, 120, 437–447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pietsch, K.; Saul, N.; Menzel, R.; Sturzenbaum, S.R.; Steinberg, C.E.W. Quercetin mediated lifespan extension in Caenorhabditis elegans is modulated by age-1, daf-2, sek-1 and unc-43. Biogerontology 2009, 10, 565–578. [Google Scholar] [CrossRef] [PubMed]
- Vayndorf, E.M.; Lee, S.S.; Liu, R.H. Whole apple extracts increase lifespan, healthspan and resistance to stress in Caenorhabditis elegans. J. Funct. Foods 2013, 5, 1235–1243. [Google Scholar] [CrossRef] [Green Version]
- Finkel, T.; Holbrook, N.J. Oxidants, oxidative stress and the biology of ageing. Nature 2000, 408, 239–247. [Google Scholar] [CrossRef]
- Forstermann, U. Oxidative stress in vascular disease: Causes, defense mechanisms and potential therapies. Nat. Rev. Cardiol. 2008, 5, 338–349. [Google Scholar] [CrossRef]
- Lin, M.T.; Beal, M.F. Mitochondrial dysfunction and oxidative stress in neurodegenerative diseases. Nature 2006, 443, 787–795. [Google Scholar] [CrossRef]
- Reuter, S.; Gupta, S.C.; Chaturvedi, M.M.; Aggarwal, B.B. Oxidative stress, inflammation, and cancer How are they linked? Free Radic. Biol. Med. 2010, 49, 1603–1616. [Google Scholar] [CrossRef] [Green Version]
- Guan, S.Y.; Tang, Q.Z.; Liu, W.W.; Zhu, R.; Li, B. Nobiletin inhibits PDGF-BB-Induced vascular smooth muscle cell proliferation and migration and attenuates neointimal hyperplasia in a rat carotid artery injury model. Drug Develop. Res. 2014, 75, 489–496. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Amarsanaa, K.; Wu, J.; Jeon, S.C.; Cui, Y.; Jung, S.C.; Park, D.B.; Kim, S.J.; Han, S.H.; Kim, H.W.; et al. Nobiletin attenuates neurotoxic mitochondrial calcium overload through K+ influx and Delta psi(m) across mitochondrial inner membrane. Korean J. Physiol. Pharmacol. 2018, 22, 311–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baumeister, R.; Schaffitzel, E.; Hertweck, M. Endocrine signaling in Caenorhabditis elegans controls stress response and longevity. J. Endocrinol. 2006, 190, 191–202. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.Z.; Jie, G.L.; Zhang, J.J.; Zhao, B.L. Significant longevity-extending effects of EGCG on Caenorhabditis elegans under stress. Free Radic. Biol. Med. 2009, 46, 414–421. [Google Scholar] [CrossRef] [PubMed]
- Kawasaki, I.; Jeong, M.H.; Oh, B.K.; Shim, Y.H. Apigenin inhibits larval growth of Caenorhabditis elegans through DAF-16 activation. FEBS Lett. 2010, 584, 3587–3591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendenhall, A.R.; Tedesco, P.M.; Taylor, L.D.; Lowe, A.; Cypser, J.R.; Johnson, T.E. Expression of a single-copy hsp-16.2 reporter predicts life span. J. Gerontol. Ser. Biomed. Sci. Med. Sci. 2012, 67, 726–733. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.H.; Feinbaum, R.; Alloing, G.; Emerson, F.E.; Garsin, D.A.; Inoue, H.; Tanaka-Hino, M.; Hisamoto, N.; Matsumoto, K.; Tan, M.W.; et al. A conserved p38 MAP kinase pathway in Caenorhabditis elegans innate immunity. Science 2002, 297, 623–626. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, M.; Yamamoto, M. Nrf2-Keap1 regulation of cellular defense mechanisms against electrophiles and reactive oxygen species. Adv. Enzyme. Regul. 2006, 46, 113–140. [Google Scholar] [CrossRef]
- Oh, S.W.; Mukhopadhyay, A.; Svrzikapa, N.; Jiang, F.; Davis, R.J.; Tissenbaum, H.A. JNK regulates lifespan in Caenorhabditis elegans by modulating nuclear translocation of forkhead transcription factor/DAF-16. Proc. Natl. Acad. Sci. USA 2005, 102, 4494–4499. [Google Scholar] [CrossRef] [Green Version]
- Seo, H.W.; Cheon, S.M.; Lee, M.H.; Kim, H.J.; Jeon, H.; Cha, D.S. Catalpol modulates lifespan via DAF-16/FOXO and SKN-1/Nrf2 activation in Caenorhabditis elegans. Evid. Based Complement. Altern. Med. 2015, 2015, 524878. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.X.; Zhang, X.Y.; Xiao, J.; Zhong, Q.Q.; Kuang, Y.; Cao, Y.; Chen, Y.J. Effects on longevity extension and mechanism of action of carnosic acid in Caenorhabditis elegans. Food Funct. 2019, 10, 1398–1410. [Google Scholar] [CrossRef] [PubMed]
- Gruber, J.; Tang, S.Y.; Halliwell, B. Evidence for a trade-off between survival and fitness caused by resveratrol treatment of Caenorhabditis elegans. Ann. N. Y. Acad. Sci. 2007, 1100, 530–542. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.J.; Cui, J.F.; Tian, G.F.; DiMarco-Crook, C.; Gao, W.; Zhao, C.Y.; Li, G.Y.; Lian, Y.H.; Xiao, H.; Zheng, J.K. Efficiency of four different dietary preparation methods in extracting functional compounds from dried tangerine peel. Food Chem. 2019, 289, 340–350. [Google Scholar] [CrossRef] [PubMed]
- Zhou, D.Y.; Zhang, X.L.; Xu, Q.; Xue, X.Y.; Zhang, F.F.; Liang, X.M. UPLC/Q-TOFMS/MS as a powerful technique for rapid identification of polymethoxylated flavones in Fructus aurantii. J. Pharm. Biomed. 2009, 50, 2–8. [Google Scholar] [CrossRef]
- Yuen, H.Q.; Hwang, Q.H.; Zhang, X.Y.; Zhou, Z.X. Cellular Antioxidant Activity and Pharmacokinetic Study of Polymethoxylated Flavonoids in Extract of Citrus reticulata ‘Chachi’ Peel. Food Sci. Technol. Res. 2014, 20, 629–637. [Google Scholar] [CrossRef] [Green Version]
- Nagata, E.; Ichi, I.; Kataoka, R.; Matsushima, M.; Adachi, N.; Kitamura, Y.; Sasaki, T.; Kojo, S. Effect of Nobiletin on Lipid Metabolism in Rats. J. Health Sci. 2010, 56, 705–711. [Google Scholar] [CrossRef] [Green Version]
- Yao, J.; Zhou, J.P.; Ping, Q.N.; Lu, Y.; Chen, L. Distribution of nobiletin chitosan-based microemulsions in brain following i.v. injection in mice. Int. J. Pharm. 2008, 352, 256–262. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Taxon Names | Variety | Concentration (mg/kg FW *) |
|---|---|---|---|
| mandarin oranges | Citrus reticulata | Nian ju | 123.83–176.54 |
| tangerines | Citrus tangerine | Dahongpao tangerine | 1159.52–1580.75 |
| oranges | Citrus sinensis | Newhall sweet orange | 171.53–264.62 |
| Blood orange | 20.56–29.02 | ||
| grapefruits | Citrus paradisi | Changshanhuyou grapefruit | 4.22–43.71 |
| Group | Number | Mean Lifespan (days) * | % of Control * | Median Survival (days) | Maximum Lifespan (days) |
|---|---|---|---|---|---|
| Control | 110 | 24.90 ± 0.37 a | 100.0 ± 1.47 a | 25.5 | 31 |
| NOB (3.13 μM) | 110 | 27.06 ± 2.06 b | 108.7 ± 6.03 b | 29 | 33 |
| NOB (6.25 μM) | 109 | 28.61 ± 0.07 bc | 114.9 ± 4.34 b | 29 | 34 |
| NOB (12.5 μM) | 116 | 30.14 ± 0.28 c | 121.0 ± 1.14 c | 30 | 36 |
| Group | Number | Mean Lifespan (h) * | % of Control * | Median Survival (h) | Maximum Lifespan (h) |
|---|---|---|---|---|---|
| Control | 100 | 13.86 ± 0.40 a | 100.0 ± 2.90 a | 14 | 20 |
| NOB (3.13 μM) | 90 | 16.40 ± 0.70 b | 118.2 ± 5.06 b | 17 | 22 |
| NOB (6.25 μM) | 94 | 16.80 ± 0.26 b | 121.1 ± 1.90 b | 18 | 24 |
| NOB (12.5 μM) | 96 | 18.10 ± 0.60 c | 130.6 ± 4.36 c | 20 | 24 |
| Group | Number | Mean Lifespan (days) * | % of Control * | Median Survival (days) | Maximum Lifespan (days) |
|---|---|---|---|---|---|
| Control | 113 | 6.88 ± 0.52 a | 100.0 ± 7.52 a | 7 | 11 |
| NOB (3.13 μM) | 92 | 7.48 ± 0.70 ab | 108.7 ± 10.2 ab | 7 | 13 |
| NOB (6.25 μM) | 94 | 8.29 ± 0.11 b | 120.5 ± 1.67 b | 8 | 13 |
| NOB (12.5 μM) | 97 | 9.17 ± 0.23 c | 133.3 ± 3.32 c | 9 | 14 |
| Group | Number | Mean Lifespan (days) * | % of Control * | Median Survival (days) | Maximum Lifespan (days) |
|---|---|---|---|---|---|
| Control | 85 | 5.06 ± 0.44 a | 100.0 ± 8.71 a | 4 | 10 |
| NOB (3.13 μM) | 112 | 5.65 ± 0.17 b | 111.6 ± 3.45 b | 6 | 11 |
| NOB (6.25 μM) | 81 | 6.60 ± 0.19 c | 130.5 ± 3.76 c | 7 | 13 |
| NOB (12.5 μM) | 86 | 7.48 ± 0.18 d | 147.8 ± 3.62 d | 7 | 13 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, X.; Wang, H.; Li, T.; Chen, L.; Zheng, B.; Liu, R.H. Nobiletin Delays Aging and Enhances Stress Resistance of Caenorhabditis elegans. Int. J. Mol. Sci. 2020, 21, 341. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21010341
Yang X, Wang H, Li T, Chen L, Zheng B, Liu RH. Nobiletin Delays Aging and Enhances Stress Resistance of Caenorhabditis elegans. International Journal of Molecular Sciences. 2020; 21(1):341. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21010341
Chicago/Turabian StyleYang, Xueyan, Hong Wang, Tong Li, Ling Chen, Bisheng Zheng, and Rui Hai Liu. 2020. "Nobiletin Delays Aging and Enhances Stress Resistance of Caenorhabditis elegans" International Journal of Molecular Sciences 21, no. 1: 341. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21010341