DRG2 Deficient Mice Exhibit Impaired Motor Behaviors with Reduced Striatal Dopamine Release


 , , , ,
, , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
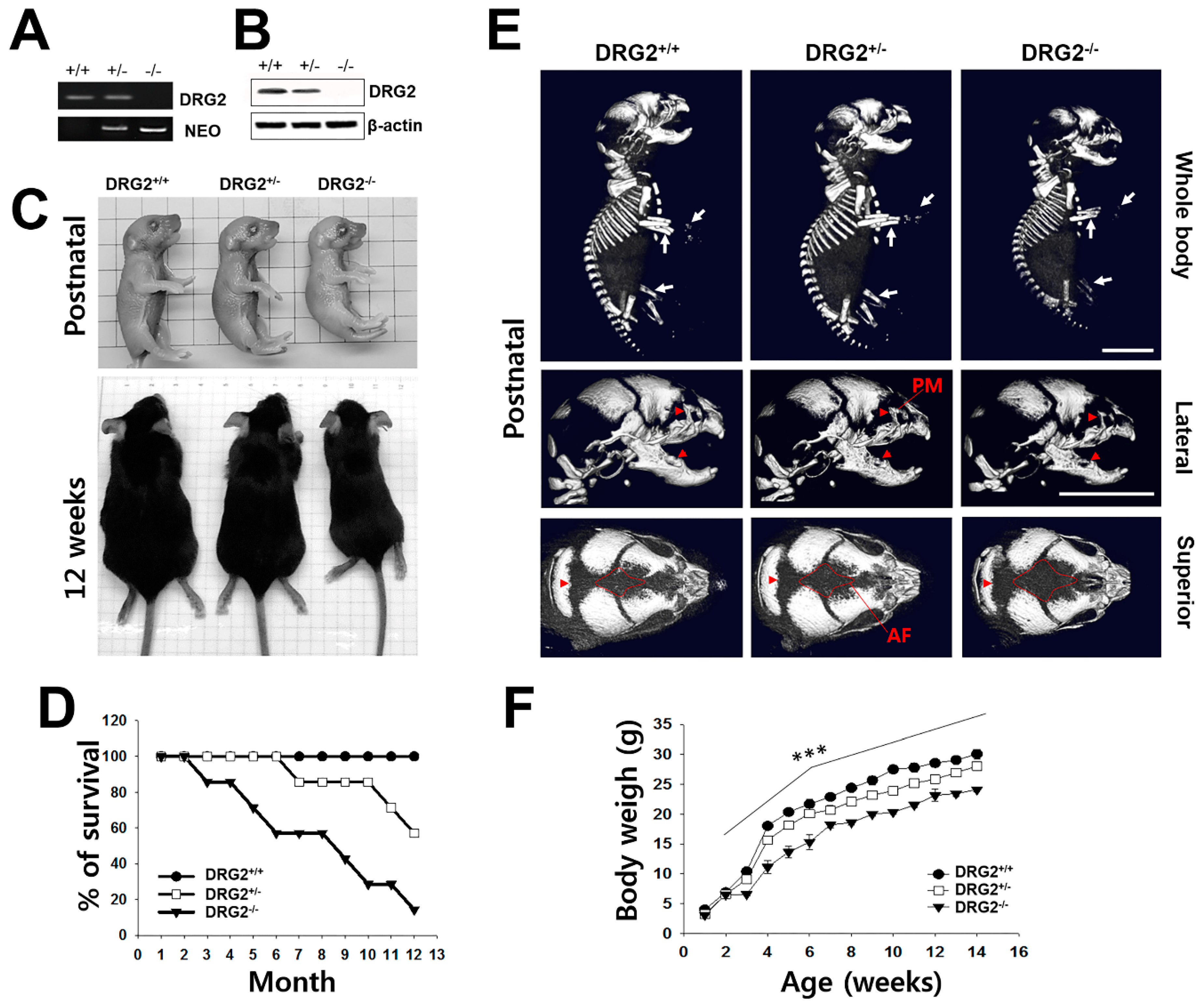
2.1. Growth Retardation and Skeletal Defects in DRG2 KO Mice
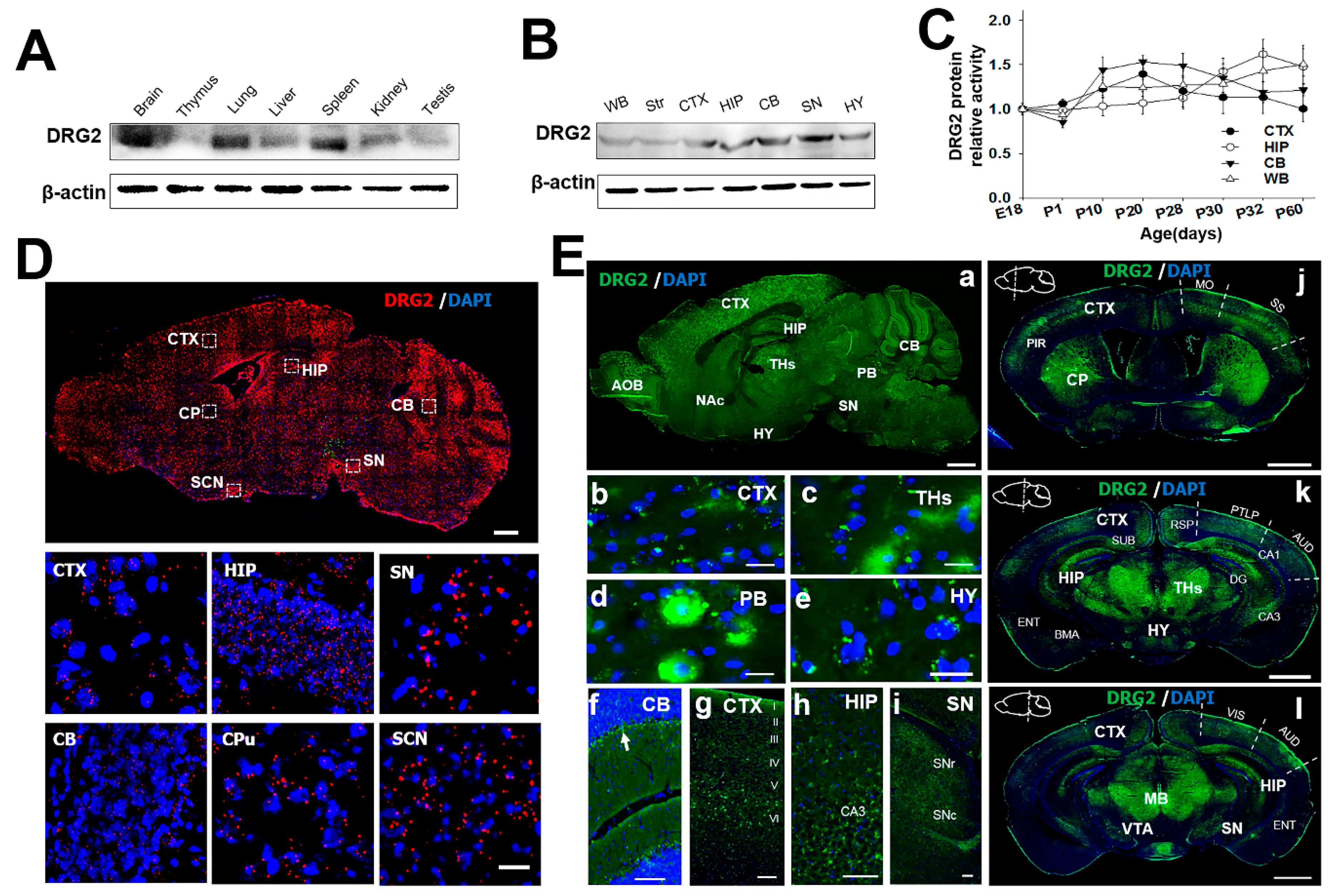
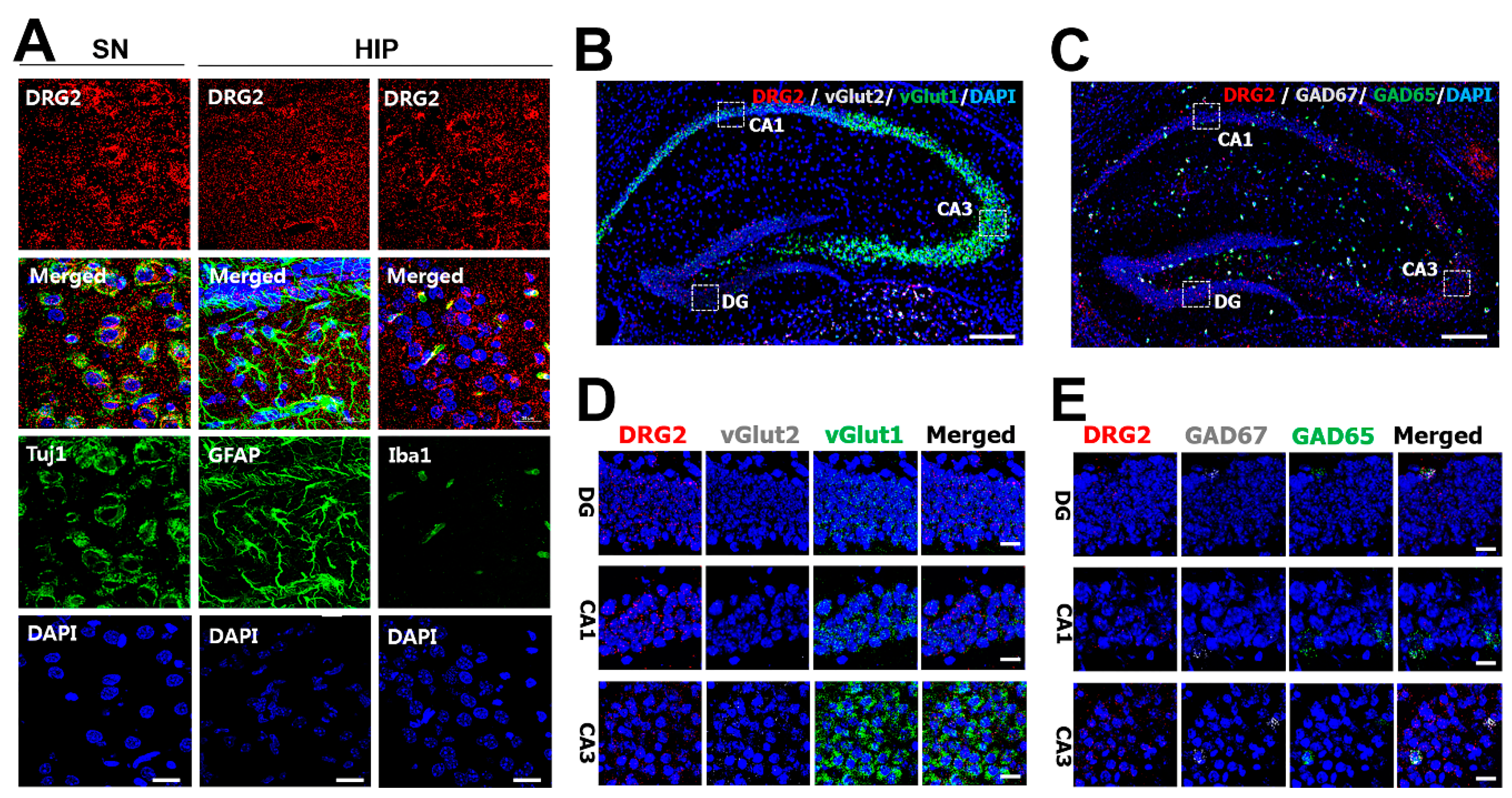
2.2. Regional Distribution of DRG2 in Mouse Brain
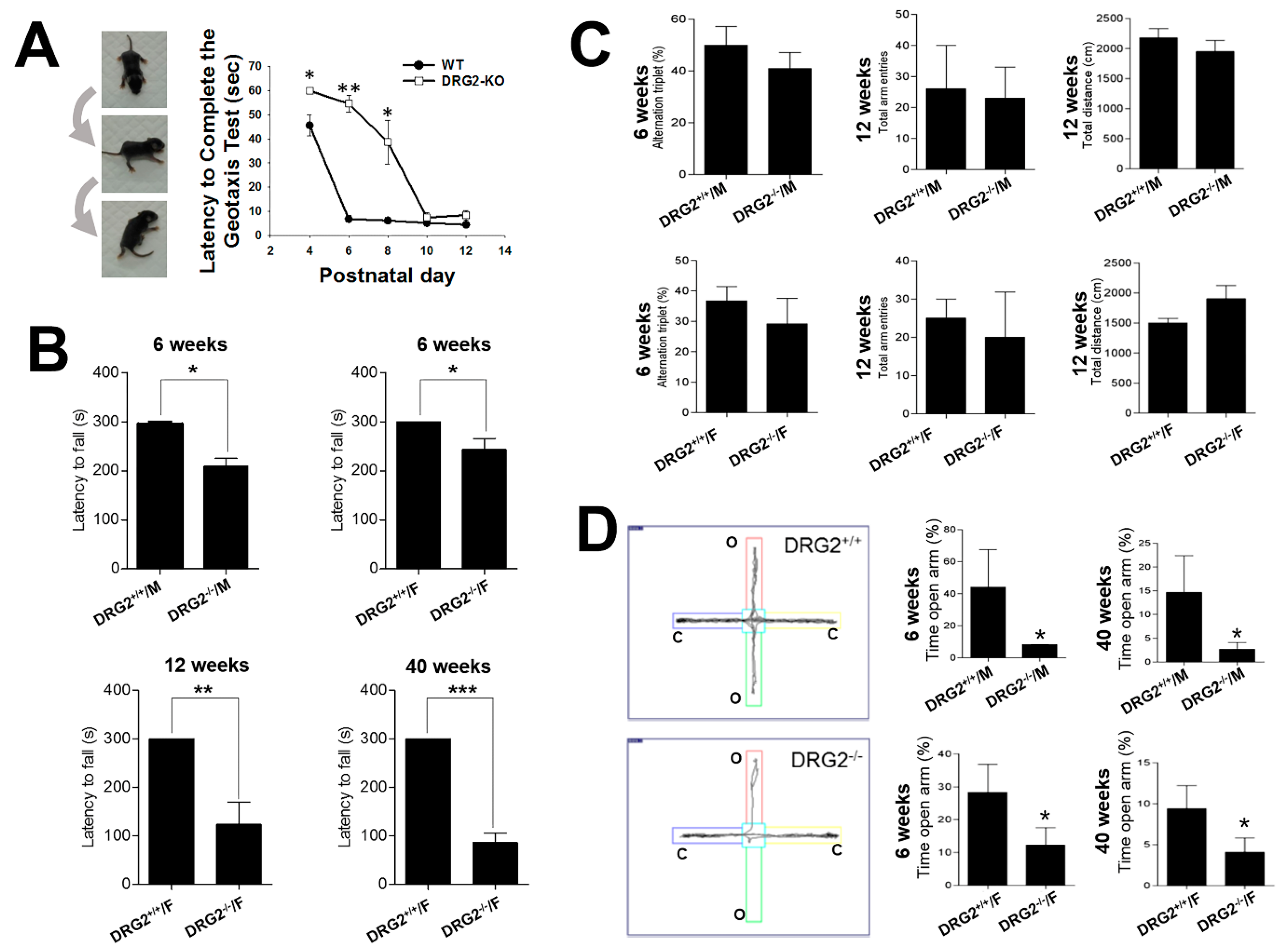
2.3. DRG2 Depletion Causes Motor Deficit and Increases Anxiety in Mice
2.4. Abnormal Behaviors of DRG2-Deficient Mice Are Caused by Dopamine Release Alterations
3. Discussion
4. Materials and Methods
4.1. Mouse Husbandry
4.2. Rotarod Test
4.3. Y-Maze Test
4.4. Elevated Plus-Maze Test
4.5. Negative Geotaxis Test
4.6. In Situ Hybridization
4.7. Immunoblotting and Immunostaining and Quantification of TH Positive Neurons
4.8. Dopamine Release Measurement with High-Performance Liquid Chromatography (HPLC)
4.9. Measurement of Dopamine Release from the Str by Slice Fast-Scan Cyclic Voltammetry (FSCV)
4.10. L-DOPA Administration
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| DRG2 | Developmentally regulated GTP-binding protein 2 |
| Str | Striatum |
| VTA | Ventral tegmental area |
| SN | Substantia nigra |
| SNc | Substantia nigra pars compacta |
| SNr | Substantia pars reticulate |
| HIP | Hippocampu |
| CB | Cerebellum |
| CTX | Cerebral cortex |
| HYP | Hypothalamus |
| SCN | Suprachiasmatic nucleus |
| THs | Thalamus |
| PB | Parabrachial nucleus |
| HY | Hypothalamus |
| AOB | Accessory olfactory bulb |
| AUD | Auditory areas |
| BMA | Basomedial amygdala nuclear |
| CA1 | Cornu ammonis1 |
| CA3 | Cornu ammonis3 |
| DG | Dentate gyrus |
| CP | Caudate putamen |
| ENT | Entorhinal area |
| MB | Mid brain |
| Mo | Momatomotor area |
| NAc | Nucleus accumbens |
| Pir | Piriform cortex |
| PTLP | Posterior parietal association areas |
| RSP | Retrosplenial area |
| SS | Somatosensory area |
| SUB | Subiculum |
| VIS | Visual area |
| WB | Whole brain |
| PD | Parkinson’s disease |
| MT | Microtubule |
| Micro-CT | Micro-computerized tomography |
| AF | Aanterior fontanel |
| PM | Premaxillary-maxillary |
| DOPAC | 3,4-dihydroxyphenylaceti |
| 3-MT | 3-methoxytyramine |
| HVA | Homovanillic acid |
| EPI | Epinephrine |
| 5-HT | Serotonin |
| TH | Tyrosine hydroxylase |
| DA | Dopamine |
| L-DOPA | l-3,4-dihydroxyphenylalanine |
| KO | Knockout |
| SEM | Standard error of mean |
References
- Grace, A.A. Dysregulation of the dopamine system in the pathophysiology of schizophrenia and depression. Nat. Rev. Neurosci. 2016, 17, 524–532. [Google Scholar] [CrossRef] [PubMed]
- Birtwistle, J.; Baldwin, D. Role of dopamine in schizophrenia and Parkinson’s disease. Br. J. Nurs. 1998, 7, 832–841. [Google Scholar] [CrossRef] [PubMed]
- Dunlop, B.W.; Nemeroff, C.B. The role of dopamine in the pathophysiology of depression. Arch. Gen. Psychiatry 2007, 64, 327–337. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Li, S.; Wen, C.; Zhang, Y.; Guo, Q.; Wang, H.; Su, B. Intrastriatal injection of colchicine induces striatonigral degeneration in mice. J. Neurochem. 2008, 106, 1815–1827. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Zhang, K.; Zhong, J.; Wang, J.; Yu, Z.; Lei, X.; Chen, X.; Quan, Y.; Xian, J.; Chen, Y.; et al. Stably maintained microtubules protect dopamine neurons and alleviate depression-like behavior after intracerebral hemorrhage. Sci. Rep. 2018, 8, 12647. [Google Scholar] [CrossRef] [Green Version]
- Janke, C.; Kneussel, M. Tubulin post-translational modifications: Encoding functions on the neuronal microtubule cytoskeleton. Trends Neurosci. 2010, 33, 362–372. [Google Scholar] [CrossRef] [PubMed]
- Hastings, T.G.; Lewis, D.A.; Zigmond, M.J. Role of oxidation in the neurotoxic effects of intrastriatal dopamine injections. Proc. Natl. Acad. Sci. USA 1996, 93, 1956–1961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Best, A.; Ahmed, S.; Kozma, R.; Lim, L. The Ras-related GTPase Rac1 binds tubulin. J. Biol. Chem. 1996, 271, 3756–3762. [Google Scholar] [CrossRef] [Green Version]
- Tivodar, S.; Kalemaki, K.; Kounoupa, Z.; Vidaki, M.; Theodorakis, K.; Denaxa, M.; Kessaris, N.; de Curtis, I.; Pachnis, V.; Karagogeos, D. Rac-GTPases Regulate Microtubule Stability and Axon Growth of Cortical GABAergic Interneurons. Cereb. Cortex 2015, 25, 2370–2382. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Billault, C.; Munoz-Llancao, P.; Henriquez, D.R.; Wojnacki, J.; Conde, C.; Caceres, A. The role of small GTPases in neuronal morphogenesis and polarity. Cytoskelet. Hoboken 2012, 69, 464–485. [Google Scholar] [CrossRef]
- Kim, H.; Calatayud, C.; Guha, S.; Fernandez-Carasa, I.; Berkowitz, L.; Carballo-Carbajal, I.; Ezquerra, M.; Fernandez-Santiago, R.; Kapahi, P.; Raya, A.; et al. The Small GTPase RAC1/CED-10 Is Essential in Maintaining Dopaminergic Neuron Function and Survival Against alpha-Synuclein-Induced Toxicity. Mol. Neurobiol. 2018, 55, 7533–7552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tahirovic, S.; Hellal, F.; Neukirchen, D.; Hindges, R.; Garvalov, B.K.; Flynn, K.C.; Stradal, T.E.; Chrostek-Grashoff, A.; Brakebusch, C.; Bradke, F. Rac1 regulates neuronal polarization through the WAVE complex. J. Neurosci. 2010, 30, 6930–6943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schenker, T.; Lach, C.; Kessler, B.; Calderara, S.; Trueb, B. A novel GTP-binding protein which is selectively repressed in SV40 transformed fibroblasts. J. Biol. Chem. 1994, 269, 25447–25453. [Google Scholar] [PubMed]
- Li, B.; Trueb, B. DRG represents a family of two closely related GTP-binding proteins. Biochim. Biophys. Acta 2000, 1491, 196–204. [Google Scholar] [CrossRef]
- Ishikawa, K.; Akiyama, T.; Ito, K.; Semba, K.; Inoue, J. Independent stabilizations of polysomal Drg1/Dfrp1 complex and non-polysomal Drg2/Dfrp2 complex in mammalian cells. Biochem. Biophys. Res. Commun. 2009, 390, 552–556. [Google Scholar] [CrossRef]
- Schellhaus, A.K.; Moreno-Andres, D.; Chugh, M.; Yokoyama, H.; Moschopoulou, A.; De, S.; Bono, F.; Hipp, K.; Schaffer, E.; Antonin, W. Developmentally Regulated GTP binding protein 1 (DRG1) controls microtubule dynamics. Sci. Rep. 2017, 7, 9996. [Google Scholar] [CrossRef] [Green Version]
- Dang, T.; Jang, S.H.; Back, S.H.; Park, J.W.; Han, I.S. DRG2 Deficiency Causes Impaired Microtubule Dynamics in HeLa Cells. Mol. Cells 2018, 41, 1045–1051. [Google Scholar]
- Mani, M.; Lee, U.H.; Yoon, N.A.; Kim, H.J.; Ko, M.S.; Seol, W.; Joe, Y.; Chung, H.T.; Lee, B.J.; Moon, C.H.; et al. Developmentally regulated GTP-binding protein 2 coordinates Rab5 activity and transferrin recycling. Mol. Biol. Cell 2016, 27, 334–348. [Google Scholar] [CrossRef]
- Mani, M.; Thao, D.T.; Kim, B.C.; Lee, U.H.; Kim, D.J.; Jang, S.H.; Back, S.H.; Lee, B.J.; Cho, W.J.; Han, I.S.; et al. DRG2 knockdown induces Golgi fragmentation via GSK3beta phosphorylation and microtubule stabilization. Biochim. Biophys. Acta Mol. Cell Res. 2019, 1866, 1463–1474. [Google Scholar] [CrossRef]
- Zhou, L.; McInnes, J.; Wierda, K.; Holt, M.; Herrmann, A.G.; Jackson, R.J.; Wang, Y.C.; Swerts, J.; Beyens, J.; Miskiewicz, K.; et al. Tau association with synaptic vesicles causes presynaptic dysfunction. Nat. Commun. 2017, 8, 15295. [Google Scholar] [CrossRef]
- Kumar, S.; Tomooka, Y.; Noda, M. Identification of a set of genes with developmentally down-regulated expression in the mouse brain. Biochem. Biophys. Res. Commun. 1992, 185, 1155–1161. [Google Scholar] [CrossRef]
- Fremeau, R.T., Jr.; Troyer, M.D.; Pahner, I.; Nygaard, G.O.; Tran, C.H.; Reimer, R.J.; Bellocchio, E.E.; Fortin, D.; Storm-Mathisen, J.; Edwards, R.H. The expression of vesicular glutamate transporters defines two classes of excitatory synapse. Neuron 2001, 31, 247–260. [Google Scholar] [CrossRef] [Green Version]
- Kreider, J.C.; Blumberg, M.S. Geotaxis and beyond: Commentary on Motz and Alberts (2005). Neurotoxicol. Teratol. 2005, 27, 535–538. [Google Scholar] [CrossRef] [PubMed]
- Kravitz, A.V.; Freeze, B.S.; Parker, P.R.; Kay, K.; Thwin, M.T.; Deisseroth, K.; Kreitzer, A.C. Regulation of parkinsonian motor behaviours by optogenetic control of basal ganglia circuitry. Nature 2010, 466, 622–626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dawson, T.M.; Dawson, V.L. Molecular pathways of neurodegeneration in Parkinson’s disease. Science 2003, 302, 819–822. [Google Scholar] [CrossRef]
- Cachope, R.; Cheer, J.F. Local control of striatal dopamine release. Front. Behav. Neurosci. 2014, 8, 188. [Google Scholar] [CrossRef] [PubMed]
- Sulzer, D.; Cragg, S.J.; Rice, M.E. Striatal dopamine neurotransmission: Regulation of release and uptake. Basal Ganglia 2016, 6, 123–148. [Google Scholar] [CrossRef] [Green Version]
- Song, H.; Kim, S.I.; Ko, M.S.; Kim, H.J.; Heo, J.C.; Lee, H.J.; Lee, H.S.; Han, I.S.; Kwack, K.; Park, J.W. Overexpression of DRG2 increases G2/M phase cells and decreases sensitivity to nocodazole-induced apoptosis. J. Biochem. 2004, 135, 331–335. [Google Scholar] [CrossRef]
- Jang, S.H.; Kim, A.R.; Park, N.H.; Park, J.W.; Han, I.S. DRG2 Regulates G2/M Progression via the Cyclin B1-Cdk1 Complex. Mol. Cells. 2016, 39, 699–704. [Google Scholar] [CrossRef] [Green Version]
- Vo, M.T.; Ko, M.S.; Lee, U.H.; Yoon, E.H.; Lee, B.J.; Cho, W.J.; Ha, C.M.; Kim, K.; Park, J.W. Developmentally regulated GTP-binding protein 2 depletion leads to mitochondrial dysfunction through downregulation of dynamin-related protein 1. Biochem. Biophys. Res. Commun. 2017, 486, 1014–1020. [Google Scholar] [CrossRef]
- Ke, K.; Sul, O.J.; Kim, W.K.; Lee, M.H.; Ko, M.S.; Suh, J.H.; Kim, H.J.; Kim, S.Y.; Park, J.W.; Choi, H.S. Overexpression of developmentally regulated GTP-binding protein-2 increases bone loss. Am. J. Physiol. Endocrinol. Metab. 2013, 304, E703–E710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giraldo, G.; Brooks, M.; Giasson, B.I.; Janus, C. Locomotor differences in mice expressing wild-type human alpha-synuclein. Neurobiol. Aging 2018, 65, 140–148. [Google Scholar] [CrossRef] [PubMed]
- Guerreiro, P.S.; Coelho, J.E.; Sousa-Lima, I.; Macedo, P.; Lopes, L.V.; Outeiro, T.F.; Pais, T.F. Mutant A53T alpha-Synuclein Improves Rotarod Performance Before Motor Deficits and Affects Metabolic Pathways. Neuromol. Med. 2017, 19, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Coizet, V.; Heilbronner, S.R.; Carcenac, C.; Mailly, P.; Lehman, J.F.; Savasta, M.; David, O.; Deniau, J.M.; Groenewegen, H.J.; Haber, S.N. Organization of the Anterior Limb of the Internal Capsule in the Rat. J. Neurosci. 2017, 37, 2539–2554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikemoto, S. Brain reward circuitry beyond the mesolimbic dopamine system: A neurobiological theory. Neurosci. Biobehav. Rev. 2010, 35, 129–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerfen, C.R.; Surmeier, D.J. Modulation of striatal projection systems by dopamine. Annu. Rev. Neurosci. 2011, 34, 441–466. [Google Scholar] [CrossRef] [Green Version]
- Shippenberg, T.S.; Bals-Kubik, R.; Herz, A. Examination of the neurochemical substrates mediating the motivational effects of opioids: Role of the mesolimbic dopamine system and D-1 vs. D-2 dopamine receptors. J. Pharmacol. Exp. Ther. 1993, 265, 53–59. [Google Scholar]
- Nakamura, T.; Sato, A.; Kitsukawa, T.; Momiyama, T.; Yamamori, T.; Sasaoka, T. Distinct motor impairments of dopamine D1 and D2 receptor knockout mice revealed by three types of motor behavior. Front. Integr. Neurosci. 2014, 8, 56. [Google Scholar] [CrossRef] [Green Version]
- Tsang, J.; Fullard, J.F.; Giakoumaki, S.G.; Katsel, P.; Katsel, P.; Karagiorga, V.E.; Greenwood, T.A.; Braff, D.L.; Siever, L.J.; Bitsios, P.; et al. The relationship between dopamine receptor D1 and cognitive performance. NPJ Schizophr. 2015, 1, 14002. [Google Scholar] [CrossRef] [Green Version]
- Williams, G.V.; Castner, S.A. Under the curve: Critical issues for elucidating D1 receptor function in working memory. Neuroscience 2006, 139, 263–276. [Google Scholar] [CrossRef]
- Rommelfanger, K.S.; Edwards, G.L.; Freeman, K.G.; Liles, L.C.; Miller, G.W.; Weinshenker, D. Norepinephrine loss produces more profound motor deficits than MPTP treatment in mice. Proc. Natl. Acad. Sci. USA 2007, 104, 13804–13809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korner, G.; Noain, D.; Ying, M.; Hole, M.; Flydal, M.I.; Scherer, T.; Allegri, G.; Rassi, A.; Fingerhut, R.; Becu-Villalobos, D.; et al. Brain catecholamine depletion and motor impairment in a Th knock-in mouse with type B tyrosine hydroxylase deficiency. Brain 2015, 138 Pt 10, 2948–2963. [Google Scholar] [CrossRef] [Green Version]
- Virk, M.S.; Sagi, Y.; Medrihan, L.; Leung, J.; Kaplitt, M.G.; Greengard, P. Opposing roles for serotonin in cholinergic neurons of the ventral and dorsal striatum. Proc. Natl. Acad. Sci. USA 2016, 113, 734–739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoon, T.; Okada, J.; Jung, M.W.; Kim, J.J. Prefrontal cortex and hippocampus subserve different components of working memory in rats. Learn. Mem. 2008, 15, 97–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Potocki, L.; Bi, W.; Treadwell-Deering, D.; Carvalho, C.M.; Eifert, A.; Friedman, E.M.; Glaze, D.; Krull, K.; Lee, J.A.; Lewis, R.A.; et al. Characterization of Potocki-Lupski syndrome (dup(17)(p11.2p11.2)) and delineation of a dosage-sensitive critical interval that can convey an autism phenotype. Am. J. Hum. Genet. 2007, 80, 633–649. [Google Scholar] [CrossRef] [Green Version]
- Bi, W.; Yan, J.; Stankiewicz, P.; Park, S.S.; Walz, K.; Boerkoel, C.F.; Potocki, L.; Shaffer, L.G.; Devriendt, K.; Nowaczyk, M.J.; et al. Genes in a refined Smith-Magenis syndrome critical deletion interval on chromosome 17p11.2 and the syntenic region of the mouse. Genome Res. 2002, 12, 713–728. [Google Scholar] [CrossRef] [Green Version]
- Smith, A.C.M.; Boyd, K.E.; Brennan, C.; Charles, J.; Elsea, S.H.; Finucane, B.M.; Foster, R.; Gropman, A.; Girirajan, S.; Haas-Givler, B. Smith-Magenis Syndrome. In GeneReviews ((R)); Adam, M.P., Ardinger, H.H., Pagon, R.A., Wallace, S.E., Bean, L.J.H., Stephens, K., Amemiya, A., Eds.; University of Washington: Seattle, WA, USA, 1993. [Google Scholar]
- Poisson, A.; Nicolas, A.; Cochat, P.; Sanlaville, D.; Rigard, C.; de Leersnyder, H.; Franco, P.; Des Portes, V.; Edery, P.; Demily, C. Behavioral disturbance and treatment strategies in Smith-Magenis syndrome. Orphanet J. Rare Dis. 2015, 10, 111. [Google Scholar] [CrossRef] [Green Version]
- Martel, P.; Leo, D.; Fulton, S.; Berard, M.; Trudeau, L.E. Role of Kv1 potassium channels in regulating dopamine release and presynaptic D2 receptor function. PLoS ONE 2011, 6, e20402. [Google Scholar] [CrossRef] [Green Version]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lim, H.R.; Vo, M.-T.; Kim, D.J.; Lee, U.H.; Yoon, J.H.; Kim, H.-J.; Kim, J.; Kim, S.R.; Lee, J.Y.; Yang, C.H.; et al. DRG2 Deficient Mice Exhibit Impaired Motor Behaviors with Reduced Striatal Dopamine Release. Int. J. Mol. Sci. 2020, 21, 60. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21010060
Lim HR, Vo M-T, Kim DJ, Lee UH, Yoon JH, Kim H-J, Kim J, Kim SR, Lee JY, Yang CH, et al. DRG2 Deficient Mice Exhibit Impaired Motor Behaviors with Reduced Striatal Dopamine Release. International Journal of Molecular Sciences. 2020; 21(1):60. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21010060
Chicago/Turabian StyleLim, Hye Ryeong, Mai-Tram Vo, Dong Jun Kim, Unn Hwa Lee, Jong Hyuk Yoon, Hyung-Jun Kim, Jeongah Kim, Sang Ryong Kim, Jun Yeon Lee, Chae Ha Yang, and et al. 2020. "DRG2 Deficient Mice Exhibit Impaired Motor Behaviors with Reduced Striatal Dopamine Release" International Journal of Molecular Sciences 21, no. 1: 60. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21010060