Essential Oils as Antiviral Agents, Potential of Essential Oils to Treat SARS-CoV-2 Infection: An In-Silico Investigation

 ,
,  ,
, 
Abstract
:
1. Introduction
2. Results and Discussion
3. Materials and Methods
3.1. Bibliographic Search Criteria
3.2. Ligand Selection
3.3. Molecular Docking
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| Adeno | Adenovirus |
| AIV | Avian influenza virus |
| BaNadD | Bacillus anthracis nicotinate mononucleotide adenylytransferase |
| BtOBP | Bovine odorant binding protein |
| COVID-19 | Coronavirus disease 2019 |
| Cox B1 | Coxsackie B1 virus |
| DENV-1 | Dengue virus serotype 1 |
| DENV-2 | Dengue virus serotype 2 |
| DENV-3 | Dengue virus serotype 3 |
| DENV-4 | Dengue virus serotype 4 |
| DrPLA2 | Russell’s viper phospholipase A2 |
| DSnorm | Normalized docking score |
| EcAspTA | Esherichia coli L-aspartate aminotransferase |
| ECHO 9 | Echovirus 9 |
| hACE2 | Human angiotensin-converting enzyme |
| HeLa | Human cervical tumor cells |
| HEp-2 | Human epithelial type 2 cells |
| HRSV | Human respiratory syncytial virus |
| HSV-1 | Herpes simplex virus 1 |
| HSV-2 | Herpes simplex virus 2 |
| IC50 | Median inhibitory concentration |
| JUNV | Junin virus |
| MDCK | Madin-Darby canine kidney cells |
| MTT | 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide |
| PCR | Polymerase chain reaction |
| PDB | Protein data bank |
| Polio 1 | Poliomyelitis virus 1 |
| RC-37 | African green monkey kidney cells |
| SARS-CoV-2 | 2019 severe acute respiratory syndrome coronavirus 2 |
| TcAChE | Torpedo acetylcholinesterase |
| Vero | African green monkey kidney cells |
| YFV | Yellow fever virus |
References
- Lai, C.-C.; Shih, T.-P.; Ko, W.-C.; Tang, H.-J.; Hsueh, P.-R. Severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) and coronavirus disease-2019 (COVID-19): The epidemic and the challenges. Int. J. Antimicrob. Agents 2020, 55, 105924. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Yu, Y.; Xu, J.; Shu, H.; Xia, J.; Liu, H.; Wu, Y.; Zhang, L.; Yu, Z.; Fang, M.; et al. Clinical course and outcomes of critically ill patients with SARS-CoV-2 pneumonia in Wuhan, China: A single-centered, retrospective, observational study. Lancet Respir. Med. 2020, 2600, 1–7. [Google Scholar] [CrossRef] [Green Version]
- WHO. Coronavirus disease 2019 (COVID-19) Situation Report—94. Available online: https://www.who.int/emergencies/diseases/novel-coronavirus-2019/situation-reports/ (accessed on 23 April 2020).
- Glezen, W.P.; Couch, R.B. Influenza viruses. In Viral Infections of Humans; Evans, A.S., Kaslow, R.A., Eds.; Springer: New York, NY, USA, 1997; pp. 473–505. ISBN 978-0306448560. [Google Scholar]
- Baigent, S.J.; McCauley, J.W. Influenza type A in humans, mammals and birds: Determinants of virus virulence, host-range and interspecies transmission. BioEssays 2003, 25, 657–671. [Google Scholar] [CrossRef] [PubMed]
- Guan, Y.; Vijaykrishna, D.; Bahl, J.; Zhu, H.; Wang, J.; Smith, G.J.D. The emergence of pandemic influenza viruses. Protein Cell 2010, 1, 9–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, N.P.A.S.; Mueller, J. Updating the accounts: Global mortality of the 1918-1920 “Spanish” influenza pandemic. Bull. Hist. Med. 2002, 76, 105–115. [Google Scholar] [CrossRef]
- Schnitzler, S.U.; Schnitzler, P. An update on swine-origin influenza virus A/H1N1: A review. Virus Genes 2009, 39, 279–292. [Google Scholar] [CrossRef]
- Hsieh, Y.-C.; Wu, T.-Z.; Liu, D.-P.; Shao, P.-L.; Chang, L.-Y.; Lu, C.Y.; Lee, C.Y.; Huang, F.-Y.; Huang, L.-M. Influenza pandemics: Past, present and future. J. Formos. Med. Assoc. 2006, 105, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Gauthier-Clerc, M.; Lebarbenchon, C.; Thomas, F. Recent expansion of highly pathogenic avian influenza H5N1: A critical review. Ibis 2007, 149, 202–214. [Google Scholar] [CrossRef]
- Earn, D.J.D.; Dushoff, J.; Levin, S.A. Ecology and evolution of the flu. Trends Ecol. Evol. 2002, 17, 334–340. [Google Scholar] [CrossRef]
- Vimalanathan, S.; Hudson, J. Anti-influenza virus activity of essential oils and vapors. Am. J. Essent. Oils Nat. Prod. 2014, 2, 47–53. [Google Scholar]
- Mallavarapu, G.R.; Ramesh, S.; Chandrasekhara, R.S.; Rao, B.R.R.; Kaul, P.N.; Bhattacharya, A.K. Investigation of the essential oil of cinnamon leaf grown at Bangalore and Hyderabad. Flavour Fragr. J. 1995, 10, 239–242. [Google Scholar] [CrossRef]
- Raina, V.K.; Srivastava, S.K.; Aggarwal, K.K.; Ramesh, S.; Kumar, S. Essential oil composition of Cinnamomum zeylanicum Blume leaves from Little Andaman, India. Flavour Fragr. J. 2001, 16, 374–376. [Google Scholar] [CrossRef]
- Fichi, G.; Flamini, G.; Zaralli, L.J.; Perrucci, S. Efficacy of an essentifal oil of Cinnamomum zeylanicum against Psoroptes cuniculi. Phytomedicine 2007, 14, 227–231. [Google Scholar] [CrossRef] [PubMed]
- Verzera, A.; Trozzi, A.; Gazea, F.; Cicciarello, G.; Cotroneo, A. Effects of rootstock on the composition of bergamot (Citrus bergamia Risso et Poiteau) essential oil. J. Agric. Food Chem. 2003, 51, 206–210. [Google Scholar] [CrossRef]
- Sawamura, M.; Onishi, Y.; Ikemoto, J.; Tu, N.T.M.; Phi, N.T.L. Characteristic odour components of bergamot (Citrus bergamia Risso) essential oil. Flavour Fragr. J. 2006, 21, 609–615. [Google Scholar] [CrossRef]
- Costa, R.; Dugo, P.; Navarra, M.; Raymo, V.; Dugo, G.; Mondello, L. Study on the chemical composition variability of some processed bergamot (Citrus bergamia) essential oils. Flavour Fragr. J. 2008, 25, 4–12. [Google Scholar] [CrossRef]
- Schipilliti, L.; Dugo, G.; Santi, L.; Dugo, P.; Mondello, L. Authentication of bergamot essential oil by gas chromatography-combustion-isotope ratio mass spectrometer (GC-C-IRMS). J. Essent. Oil Res. 2011, 23, 60–71. [Google Scholar] [CrossRef]
- Tundis, R.; Loizzo, M.R.; Bonesi, M.; Menichini, F.; Mastellone, V.; Colica, C.; Menichini, F. Comparative study on the antioxidant capacity and cholinesterase inhibitory activity of Citrus aurantifolia Swingle, C. aurantium L., and C. bergamia Risso and Poit. peel essential oils. J. Food Sci. 2012, 77, H40–H46. [Google Scholar] [CrossRef]
- Padalia, R.C.; Verma, R.S.; Chanotiya, C.S.; Yadav, A. Chemical fingerprinting of the fragrant volatiles of nineteen Indian cultivars of Cymbopogon Spreng. (Poaceae). Rec. Nat. Prod. 2011, 5, 290–299. [Google Scholar]
- Gupta, A.K.; Muhury, R.; Ganjewala, D. A study on antimicrobial activities of essential oils of different cultivars of lemongrass (Cymbopogon flexuosus). Pharm. Sci. 2016, 22, 164–169. [Google Scholar] [CrossRef]
- Satyal, P.; Murray, B.L.; McFeeters, R.L.; Setzer, W.N. Essential oil characterization of Thymus vulgaris from various geographical locations. Foods 2016, 5, 70. [Google Scholar] [CrossRef] [PubMed]
- Erdoǧan Orhan, I.; Özçelik, B.; Kartal, M.; Kan, Y. Antimicrobial and antiviral effects of essential oils from selected Umbelliferae and Labiatae plants and individual essential oil components. Turkish J. Biol. 2012, 36, 239–246. [Google Scholar]
- Alburn, H.E.; Chester, W.; Greenspan, G. Thymol as an anti-influenza agent. U.S. Patent 3,632,782, 4 January 1972. [Google Scholar]
- Evandri, M.G.; Battinelli, L.; Daniele, C.; Mastrangelo, S.; Bolle, P.; Mazzanti, G. The antimutagenic activity of Lavandula angustifolia (lavender) essential oil in the bacterial reverse mutation assay. Food Chem. Toxicol. 2005, 43, 1381–1387. [Google Scholar] [CrossRef] [PubMed]
- De Martino, L.; De Feo, V.; Nazzaro, F. Chemical composition and in vitro antimicrobial and mutagenic activities of seven Lamiaceae essential oils. Molecules 2009, 14, 4213–4230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lafhal, S.; Vanloot, P.; Bombarda, I.; Kister, J.; Dupuy, N. Chemometric analysis of French lavender and lavandin essential oils by near infrared spectroscopy. Ind. Crops Prod. 2016, 80, 156–164. [Google Scholar] [CrossRef]
- De Rapper, S.; Viljoen, A.; van Vuuren, S. The in vitro antimicrobial effects of Lavandula angustifolia essential oil in combination with conventional antimicrobial agents. Evidence-Based Complement. Altern. Med. 2016, 2016, 2752739. [Google Scholar] [CrossRef] [Green Version]
- Hammer, K.A.; Carson, C.F.; Riley, T.V.; Nielsen, J.B. A review of the toxicity of Melaleuca alternifolia (tea tree) oil. Food Chem. Toxicol. 2006, 44, 616–625. [Google Scholar] [CrossRef]
- Garozzo, A.; Timpanaro, R.; Bisignano, B.; Furneri, P.M.; Bisignano, G.; Castro, A. In vitro antiviral activity of Melaleuca alternifolia essential oil. Lett. Appl. Microbiol. 2009, 49, 806–808. [Google Scholar] [CrossRef]
- Garozzo, A.; Timpanaro, R.; Stivala, A.; Bisignano, G.; Castro, A. Activity of Melaleuca alternifolia (tea tree) oil on influenza virus A/PR/8: Study on the mechanism of action. Antiviral Res. 2011, 89, 83–88. [Google Scholar] [CrossRef]
- Usachev, E.V.; Pyankov, O.V.; Usacheva, O.V.; Agranovski, I.E. Antiviral activity of tea tree and eucalyptus oil aerosol and vapour. J. Aerosol Sci. 2013, 59, 22–30. [Google Scholar] [CrossRef]
- Sonnberg, S.; Webby, R.J.; Webster, R.G. Natural history of highly pathogenic avian influenza H5N1. Virus Res. 2013, 178, 63–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belser, J.A.; Tumpey, T.M. H5N1 pathogenesis studies in mammalian models. Virus Res. 2013, 178, 168–185. [Google Scholar] [CrossRef] [Green Version]
- Nagy, M.M.; Al-Mahdy, D.A.; Abd El Aziz, O.M.; Kandil, A.M.; Tantawy, M.A.; El Alfy, T.S.M. Chemical composition and antiviral activity of essential oils from Citrus reshni Hort. ex Tanaka (Cleopatra mandarin) cultivated in Egypt. J. Essent. Oil-Bearing Plants 2018, 21, 264–272. [Google Scholar] [CrossRef]
- García, C.C.; Talarico, L.; Almeida, N.; Colombres, S.; Duschatzky, C.; Damonte, E.B. Virucidal activity of essential oils from aromatic plants of San Luis, Argentina. Phyther. Res. 2003, 17, 1073–1075. [Google Scholar] [CrossRef]
- Duschatzky, C.B.; Possetto, M.L.; Talarico, L.B.; García, C.C.; Michis, F.; Almeida, N.V.; De Lampasona, M.P.; Schuff, C.; Damonte, E.B. Evaluation of chemical and antiviral properties of essential oils from South American plants. Antivir. Chem. Chemother. 2005, 16, 247–251. [Google Scholar] [CrossRef] [PubMed]
- Sinico, C.; De Logu, A.; Lai, F.; Valenti, D.; Manconi, M.; Loy, G.; Bonsignore, L.; Fadda, A.M. Liposomal incorporation of Artemisia arborescens L. essential oil and in vitro antiviral activity. Eur. J. Pharm. Biopharm. 2005, 59, 161–168. [Google Scholar] [CrossRef]
- Saddi, M.; Sanna, A.; Cottiglia, F.; Chisu, L.; Casu, L.; Bonsignore, L.; De Logu, A. Antiherpevirus activity of Artemisia arborescens essential oil and inhibition of lateral diffusion in Vero cells. Ann. Clin. Microbiol. Antimicrob. 2007, 6, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gavanji, S.; Sayedipour, S.S.; Larki, B.; Bakhtari, A. Antiviral activity of some plant oils against herpes simplex virus type 1 in Vero cell culture. J. Acute Med. 2015, 5, 62–68. [Google Scholar] [CrossRef] [Green Version]
- Chung, M.S. Antiviral activities of Artemisia princeps var. orientalis essential oil and its α-thujone against norovirus surrogates. Food Sci. Biotechnol. 2017, 26, 1457–1461. [Google Scholar] [CrossRef]
- Meneses, R.; Ocazionez, R.E.; Martínez, J.R.; Stashenko, E.E. Inhibitory effect of essential oils obtained from plants grown in Colombia on yellow fever virus replication in vitro. Ann. Clin. Microbiol. Antimicrob. 2009, 8, 8. [Google Scholar]
- Haddad, J.G.; Picard, M.; Bénard, S.; Desvignes, C.; Desprès, P.; Diotel, N.; El Kalamouni, C. Ayapana triplinervis essential oil and its main component thymohydroquinone dimethyl ether inhibit Zika virus at doses devoid of toxicity in zebrafish. Molecules 2019, 24, 3447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loizzo, M.R.; Saab, A.; Tundis, R.; Statti, G.A.; Lampronti, I.; Menichini, F.; Gambari, R.; Cinatl, J.; Doerr, H.W. Phytochemical analysis and in vitro evaluation of the biological activity against herpes simplex virus type 1 (HSV-1) of Cedrus libani A. Rich. Phytomedicine 2008, 15, 79–83. [Google Scholar] [CrossRef] [PubMed]
- Setzer, W.N. Essential oils as complementary and alternative medicines for the treatment of influenza. Am. J. Essent. Oil Nat. Prod. 2016, 4, 16–22. [Google Scholar]
- Bourne, K.Z.; Bourne, N.; Reising, S.F.; Stanberry, L.R. Plant products as topical microbicide candidates: Assessment of in vitro and in vivo activity against herpes simplex virus type 2. Antiviral Res. 1999, 42, 219–226. [Google Scholar] [CrossRef]
- Minami, M.; Kita, M.; Nakaya, T.; Yamamoto, T.; Kuriyama, H.; Imanishi, J. The inhibitory effect of essential oils on herpes simplex virus type-1 replication in vitro. Microbiol. Immunol. 2003, 47, 681–684. [Google Scholar] [CrossRef]
- El Mokni, R.; Youssef, F.S.; Jmii, H.; Khmiri, A.; Bouazzi, S.; Jlassi, I.; Jaidane, H.; Dhaouadi, H.; Ashour, M.L.; Hammami, S. The Essential oil of Tunisian Dysphania ambrosioides and its antimicrobial and antiviral properties. J. Essent. Oil-Bearing Plants 2019, 22, 282–294. [Google Scholar] [CrossRef]
- Elaissi, A.; Rouis, Z.; Salem, N.A.B.; Mabrouk, S.; ben Salem, Y.; Salah, K.B.H.; Aouni, M.; Farhat, F.; Chemli, R.; Harzallah-Skhiri, F.; et al. Chemical composition of 8 Eucalyptus species’ essential oils and the evaluation of their antibacterial, antifungal and antiviral activities. BMC Complement. Altern. Med. 2012, 12, 81. [Google Scholar] [CrossRef] [Green Version]
- El-Baz, F.K.; Mahmoud, K.; El-Senousy, W.M.; Darwesh, O.M.; El Gohary, A.E. Antiviral – antimicrobial and schistosomicidal activities of Eucalyptus camaldulensis essential oils. Int. J. Pharm. Sci. Rev. Res. 2015, 31, 262–268. [Google Scholar]
- Astani, A.; Reichling, J.; Schnitzler, P. Comparative study on the antiviral activity of selected monoterpenes derived from essential oils. Phyther. Res. 2010, 24, 673–679. [Google Scholar] [CrossRef]
- Ibrahim, N.A.; El-Hawary, S.S.; Mohammed, M.M.D.; Farid, M.A.; Abdel-Wahed, N.A.M.; Ali, M.A.; El-Abd, E.A.W. Chemical composition, antiviral against avian influenza (H5N1) virus and antimicrobial activities of the essential oils of the leaves and fruits of Fortunella margarita Lour. Swingle growing in Egypt. J. Appl. Pharm. Sci. 2015, 5, 6–12. [Google Scholar]
- Hayashi, K.; Kamiya, M.; Hayashi, T. Virucidal effects of the steam distillate from Houttuynia cordata and its components on HSV-1, influenza virus, and HIV. Planta Med. 1995, 61, 237–241. [Google Scholar] [CrossRef] [PubMed]
- Brand, Y.M.; Roa-Linares, V.C.; Betancur-Galvis, L.A.; Durán-García, D.C.; Stashenko, E. Antiviral activity of Colombian Labiatae and Verbenaceae family essential oils and monoterpenes on human herpes viruses. J. Essent. Oil Res. 2016, 28, 130–137. [Google Scholar] [CrossRef]
- Schnitzler, P.; Koch, C.; Reichling, J. Susceptibility of drug-resistant clinical herpes simplex virus type 1 strains to essential oils of ginger, thyme, hyssop, and sandalwood. Antimicrob. Agents Chemother. 2007, 51, 1859–1862. [Google Scholar] [CrossRef] [Green Version]
- Koch, C.; Reichling, J.; Schneele, J.; Schnitzler, P. Inhibitory effect of essential oils against herpes simplex virus type 2. Phytomedicine 2008, 15, 71–78. [Google Scholar] [CrossRef]
- Koch, C.; Reichling, J.; Kehm, R.; Sharaf, M.M.; Zentgraf, H.; Schneele, J.; Schnitzler, P. Efficacy of anise oil, dwarf-pine oil and chamomile oil against thymidine-kinase-positive and thymidine-kinase-negative herpesviruses. J. Pharm. Pharmacol. 2008, 60, 1545–1550. [Google Scholar] [CrossRef] [PubMed]
- Astani, A.; Schnitzler, P.; Reichling, J. Screening for antiviral activities of isolated compounds from essential oils. Evidence-Based Complement. Altern. Med. 2011, 2011, 253643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reichling, J.; Koch, C.; Stahl-Biskup, E.; Sojka, C.; Schnitzler, P. Virucidal activity of a β-triketone-rich essential oil of Leptospermum scoparium (Manuka Oil) Against HSV-1 and HSV-2 in cell culture. Planta Med. 2005, 71, 1123–1127. [Google Scholar] [CrossRef]
- Ocazionez, R.E.; Meneses, R.; Torres, F.Á.; Stashenko, E. Virucidal activity of Colombian Lippia essential oils on dengue virus replication in vitro. Mem. Inst. Oswaldo Cruz 2010, 105, 304–309. [Google Scholar] [CrossRef] [Green Version]
- Gómez, L.A.; Stashenko, E.; Ocazionez, R.E. Comparative study on in vitro activities of citral, limonene and essential oils from Lippia citriodora and L. alba on yellow fever virus. Nat. Prod. Commun. 2013, 8, 249–252. [Google Scholar] [CrossRef] [Green Version]
- Pilau, M.R.; Alves, S.H.; Weiblen, R.; Arenhart, S.; Cueto, A.P.; Lovato, L.T. Antiviral activity of the Lippia graveolens (Mexican oregano) essential oil and its main compound carvacrol against human and animal viruses. Brazilian J. Microbiol. 2011, 42, 1616–1624. [Google Scholar] [CrossRef] [Green Version]
- Civitelli, L.; Panella, S.; Marcocci, M.E.; De Petris, A.; Garzoli, S.; Pepi, F.; Vavala, E.; Ragno, R.; Nencioni, L.; Palamara, A.T.; et al. In vitro inhibition of herpes simplex virus type 1 replication by Mentha suaveolens essential oil and its main component piperitenone oxide. Phytomedicine 2014, 21, 857–865. [Google Scholar] [CrossRef] [PubMed]
- Brun, P.; Bernabè, G.; Filippini, R.; Piovan, A. In vitro antimicrobial activities of commercially available tea tree (Melaleuca alternifolia) essential oils. Curr. Microbiol. 2019, 76, 108–116. [Google Scholar] [CrossRef] [PubMed]
- Farag, R.S.; Shalaby, A.S.; El-Baroty, G.A.; Ibrahim, N.A.; Ali, M.A.; Hassan, E.M. Chemical and biological evaluation of the essential oils of different Melaleuca species. Phyther. Res. 2004, 18, 30–35. [Google Scholar] [CrossRef]
- Allahverdiyev, A.; Duran, N.; Ozguven, M.; Koltas, S. Antiviral activity of the volatile oils of Melissa officinalis L. against Herpes simplex virus type-2. Phytomedicine 2004, 11, 657–661. [Google Scholar] [CrossRef] [PubMed]
- Schnitzler, P.; Schuhmacher, A.; Astani, A.; Reichling, J. Melissa officinalis oil affects infectivity of enveloped herpesviruses. Phytomedicine 2008, 15, 734–740. [Google Scholar] [CrossRef]
- Schuhmacher, A.; Reichling, J.; Schnitzler, P. Virucidal effect of peppermint oil on the enveloped viruses herpes simplex virus type 1 and type 2 in vitro. Phytomedicine 2003, 10, 504–510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Moussaoui, N.; Sanchez, G.; Khay, E.O.; Idaomar, M.; Ibn Mansour, A.; Abrini, J.; Aznar, R. Antibacterial and antiviral activities of essential oils of northern Moroccan plants. Br. Biotechnol. J. 2013, 3, 318–331. [Google Scholar] [CrossRef]
- Bouazzi, S.; Jmii, H.; El Mokni, R.; Faidi, K.; Falconieri, D.; Piras, A.; Jaïdane, H.; Porcedda, S.; Hammami, S. Cytotoxic and antiviral activities of the essential oils from Tunisian fern, Osmunda regalis. S. Afr. J. Bot. 2018, 118, 52–57. [Google Scholar] [CrossRef]
- Sivropoulou, A.; Nikolaou, C.; Papanikolaou, E.; Kokkini, S.; Lanaras, T.; Arsenakis, M. Antimicrobial, cytotoxic, and antiviral activities of Salvia fructicosa essential oil. J. Agric. Food Chem. 1997, 45, 3197–3201. [Google Scholar] [CrossRef]
- Benencia, F.; Courrèges, M.C. Antiviral activity of sandalwood oil against herpes simplex viruses-1 and -2. Phytomedicine 1999, 6, 119–123. [Google Scholar] [CrossRef]
- De Logu, A.; Loy, G.; Pellerano, M.L.; Bonsignore, L.; Schivo, M.L. Inactivation of HSV-1 and HSV-2 and prevention of cell-to-cell virus spread by Santolina insularis essential oil. Antiviral Res. 2000, 48, 177–185. [Google Scholar] [CrossRef]
- Zeghib, A.; Calliste, C.-A.; Simon, A.; Charfeddine, R.; Aouni, M.; Duroux, J.-L.; Kabouche, A.; Kabouche, Z. Chemical composition and biological potential of Thymus willdenowii Boiss. & Reut. essential oil. Nat. Prod. Res. 2019, 1–4. [Google Scholar] [CrossRef]
- Roy, S.; Chaurvedi, P.; Chowdhary, A. Evaluation of antiviral activity of essential oil of Trachyspermum ammi against Japanese encephalitis virus. Pharmacognosy Res. 2015, 7, 263–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shayeganmehr, A.; Vasfi Marandi, M.; Karimi, V.; Barin, A.; Ghalyanchilangeroudi, A. Zataria multiflora essential oil reduces replication rate of avian influenza virus (H9N2 subtype) in challenged broiler chicks. Br. Poult. Sci. 2018, 59, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Camero, M.; Lanave, G.; Catella, C.; Capozza, P.; Gentile, A.; Fracchiolla, G.; Britti, D.; Martella, V.; Buonavoglia, C.; Tempesta, M. Virucidal activity of ginger essential oil against caprine alphaherpesvirus-1. Vet. Microbiol. 2019, 230, 150–155. [Google Scholar] [CrossRef]
- Trovati, G.; Chierice, G.O.; Sanches, E.A.; Galhiane, M.S. Essential oil composition of Aloysia gratissima from Brazil. J. Essent. Oil Res. 2009, 21, 325–326. [Google Scholar] [CrossRef]
- Santos, F.M.; Pinto, J.E.B.P.; Bertolucci, S.K.V.; Alvarenga, A.A.; Alves, M.N.; Duarte, M.C.T.; Sartoratto, A. Chemical composition and antimicrobial activity of the essential oil from the leaves and flowers of Aloysia gratissima. Rev. Bras. Plantas Med. 2013, 15, 583–588. [Google Scholar] [CrossRef] [Green Version]
- Militello, M.; Settanni, L.; Aleo, A.; Mammina, C.; Moschetti, G.; Giammanco, G.M.; Blàzquez, M.A.; Carrubba, A. Chemical composition and antibacterial potential of Artemisia arborescens L. essential oil. Curr. Microbiol. 2011, 62, 1274–1281. [Google Scholar] [CrossRef] [Green Version]
- Bertuzzi, G.; Tirillini, B.; Angelini, P.; Venanzoni, R. Antioxidative action of Citrus limonum essential oil on skin. European J. Med. Plants 2013, 3, 1–9. [Google Scholar] [CrossRef] [Green Version]
- El-Leel, O.F.A.; El-Said, N.A.M. Growth, essential oil and molecular genetic identification studies of some Eucalyptus species cultivated under Egyptian conditions. BAOJ Biotechnol. 2016, 2, 20. [Google Scholar]
- Silvestre, A.J.D.; Cavaleiro, J.A.S.; Delmond, B.; Filliatre, C.; Bourgeois, G. Analysis of the variation of the essential oil composition of Eucalyptus globulus Labill. from Portugal using multivariate statistical analysis. Ind. Crops Prod. 1997, 6, 27–33. [Google Scholar] [CrossRef]
- Barazandeh, M.M. Essential oil composition of Lavandula latifolia Medik from Iran. J. Essent. Oil Res. 2002, 14, 103–104. [Google Scholar] [CrossRef]
- Pozzatti, P.; Scheid, L.A.; Spader, T.B.; Atayde, M.L.; Santurio, J.M.; Alves, S.H. In vitro activity of essential oils extracted from plants used as spices against fluconazole-resistant and fluconazole-susceptible Candida spp. Can. J. Microbiol. 2008, 54, 950–956. [Google Scholar] [CrossRef] [PubMed]
- Raal, A.; Orav, A.; Püssa, T.; Valner, C.; Malmiste, B.; Arak, E. Content of essential oil, terpenoids and polyphenols in commercial chamomile (Chamomilla recutita L. Rauschert) teas from different countries. Food Chem. 2012, 131, 632–638. [Google Scholar] [CrossRef]
- Demir, H.; Kalaycı, S. Chemical composition and antimicrobial activity of essential oils of Ocimum basilicum var. album (L.) Benth, Lavandula angustifolia subsp. angustifolia, Melissa officinalis belonging to Lamiaceae family. J. Food Sci. Eng. 2017, 7, 461–471. [Google Scholar]
- Stevanovic, T.; Garneau, F.-X.; Jean, F.-I.; Gagnon, H.; Vilotic, D.; Petrovic, S.; Ruzic, N.; Pichette, A. The essential oil composition of Pinus mugo Turra from Serbia. Flavour Fragr. J. 2005, 20, 96–97. [Google Scholar] [CrossRef]
- Hoim, Y.; Hiltunen, R. Chemical composition of a commercial oil of Ravensara aromatica Sonn. used in aromatherapy. J. Essent. Oil Res. 1999, 11, 677–678. [Google Scholar] [CrossRef]
- Poli, F.; Bonsignore, L.; Loy, G.; Sacchetti, G.; Ballero, M. Comparison between the essential oils of Santolina insularis (Genn. ex Fiori) Arrigoni and Santolina corsica Jord. et Fourr. from the island of Sardinia (Italy). J. Ethnopharmacol. 1997, 56, 201–208. [Google Scholar] [CrossRef]
- Tsai, J.-J.; Liu, W.-L.; Lin, P.-C.; Huang, B.-Y.; Tsai, C.-Y.; Chou, P.-H.; Lee, F.-C.; Ping, C.-F.; Lee, P.-Y.A.; Liu, L.-T.; et al. An RT-PCR panel for rapid serotyping of dengue virus serotypes 1 to 4 in human serum and mosquito on a field-deployable PCR system. PLoS ONE 2019, 14, e0214328. [Google Scholar] [CrossRef] [Green Version]
- Vasconcelos, P.F.C.; Monath, T.P. Yellow fever remains a potential threat to public health. Vector-Borne Zoonotic Dis. 2016, 16, 566–567. [Google Scholar] [CrossRef]
- Douam, F.; Ploss, A. Yellow fever virus: Knowledge gaps impeding the fight against an old foe. Trends Microbiol. 2018, 26, 913–928. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, K.; Imanishi, N.; Kashiwayama, Y.; Kawano, A.; Terasawa, K.; Shimada, Y.; Ochiai, H. Inhibitory effect of cinnamaldehyde, derived from Cinnamomi cortex, on the growth of influenza A/PR/8 virus in vitro and in vivo. Antiviral Res. 2007, 74, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Benencia, F.; Courrèges, M.C. In vitro and in vivo activity of eugenol on human herpesvirus. Phyther. Res. 2000, 14, 495–500. [Google Scholar] [CrossRef]
- Liao, Q.; Qian, Z.; Liu, R.; An, L.; Chen, X. Germacrone inhibits early stages of influenza virus infection. Antiviral Res. 2013, 100, 578–588. [Google Scholar] [CrossRef]
- Armaka, M.; Papanikolaou, E.; Sivropoulou, A.; Arsenakis, M. Antiviral properties of isoborneol, a potent inhibitor of herpes simplex virus type 1. Antiviral Res. 1999, 43, 79–92. [Google Scholar] [CrossRef]
- Ryabchenko, B.; Tulupova, E.; Schmidt, E.; Wlcek, K.; Buchbauer, G.; Jirovetz, L. Investigation of anticancer and antiviral properties of selected aroma samples. Nat. Prod. Commun. 2008, 3, 1085–1088. [Google Scholar] [CrossRef] [Green Version]
- Kiyohara, H.; Ichino, C.; Kawamura, Y.; Nagai, T.; Sato, N.; Yamada, H. Patchouli alcohol: In vitro direct anti-influenza virus sesquiterpene in Pogostemon cablin Benth. J. Nat. Med. 2012, 66, 55–61. [Google Scholar] [CrossRef]
- Wu, H.; Li, B.; Wang, X.; Jin, M.; Wang, G. Inhibitory effect and possible mechanism of action of patchouli alcohol against influenza a (H2N2) virus. Molecules 2011, 16, 6489–6501. [Google Scholar] [CrossRef] [Green Version]
- Gentile, D.; Patamia, V.; Scala, A.; Sciortino, M.T.; Piperno, A.; Rescifina, A. Inhibitors of SARS-CoV-2 main protease from a library of marine natural products: A virtual screening and molecular modeling study. Mar. Drugs 2020, 18, 225. [Google Scholar] [CrossRef] [Green Version]
- Thuy, B.T.P.; My, T.T.A.; Hai, N.T.T.; Hieu, L.T.; Hoa, T.T.; Thi Phuong Loan, H.; Triet, N.T.; Van Anh, T.T.; Quy, P.T.; Van Tat, P.; et al. Investigation into SARS-CoV-2 resistance of compounds in garlic essential oil. ACS Omega 2020. early view. [Google Scholar] [CrossRef]
- Joshi, R.; Jagdale, S.; Bansode, S.; Shankar, S.S.; Tellis, M.; Pandya, V.K.; Giri, A.; Kulkarni, M. Discovery of multi-target-directed ligands by targeting host-specific SARS-CoV-2’s structurally conserved main protease. Preprints 2020, 2020, 2020040068. [Google Scholar]
- Manish, M. Studies on computational molecular interaction between SARS-CoV-2 main protease and natural products. ChemRxiv 2020. preprint. [Google Scholar] [CrossRef]
- Beck, B.R.; Shin, B.; Choi, Y.; Park, S.; Kang, K. Predicting commercially available antiviral drugs that may act on the novel coronavirus (SARS-CoV-2) through a drug-target interaction deep learning model. Comput. Struct. Biotechnol. J. 2020, 18, 784–790. [Google Scholar] [CrossRef] [PubMed]
- Hofmarcher, M.; Mayr, A.; Rumetshofer, E.; Ruch, P.; Renz, P.; Schimunek, J.; Seidl, P.; Vall, A.; Widrich, M.; Hochreiter, S.; et al. Large-scale ligand-based virtual screening for SARS-CoV-2 inhibitors using deep neural networks. SSRN Electron. J. 2020. Available online: https://ssrn.com/abstract=3561442 (accessed on 13 April 2020). [CrossRef] [Green Version]
- Zhang, L.; Lin, D.; Sun, X.; Curth, U.; Drosten, C.; Sauerhering, L.; Becker, S.; Rox, K.; Hilgenfeld, R. Crystal structure of SARS-CoV-2 main protease provides a basis for design of improved α-ketoamide inhibitors. Science 2020, 3405, 409–412. [Google Scholar] [CrossRef] [Green Version]
- Bhardwaj, K.; Sun, J.; Holzenburg, A.; Guarino, L.A.; Kao, C.C. RNA recognition and cleavage by the SARS coronavirus endoribonuclease. J. Mol. Biol. 2006, 361, 243–256. [Google Scholar] [CrossRef]
- Saikatendu, K.S.; Joseph, J.S.; Subramanian, V.; Clayton, T.; Griffith, M.; Moy, K.; Velasquez, J.; Neuman, B.W.; Buchmeier, M.J.; Stevens, R.C.; et al. Structural basis of severe acute respiratory syndrome coronavirus ADP-ribose-1″-phosphate dephosphorylation by a conserved domain of nsP3. Structure 2005, 13, 1665–1675. [Google Scholar] [CrossRef] [Green Version]
- Putics, A.; Filipowicz, W.; Hall, J.; Gorbalenya, A.E.; Ziebuhr, J. ADP-ribose-1"-monophosphatase: A conserved coronavirus enzyme that is dispensable for viral replication in tissue culture. J. Virol. 2005, 79, 12721–12731. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Luo, H.; Chen, L.; Chen, J.; Shen, J.; Zhu, W.; Chen, K.; Shen, X.; Jiang, H. An overall picture of SARS coronavirus (SARS-CoV) genome-encoded major proteins: Structures, functions and drug development. Curr. Pharm. Des. 2006, 12, 4539–4553. [Google Scholar] [CrossRef]
- Yan, R.; Zhang, Y.; Li, Y.; Xia, L.; Guo, Y.; Zhou, Q. Structural basis for the recognition of SARS-CoV-2 by full-length human ACE2. Science 2020, 367, 1444–1448. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Penninger, J.M.; Li, Y.; Zhong, N.; Slutsky, A.S. Angiotensin-converting enzyme 2 (ACE2) as a SARS-CoV-2 receptor: Molecular mechanisms and potential therapeutic target. Intensive Care Med. 2020, 46, 586–590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pourghanbari, G.; Nili, H.; Moattari, A.; Mohammadi, A.; Iraji, A. Antiviral activity of the oseltamivir and Melissa officinalis L. essential oil against avian influenza A virus (H9N2). VirusDisease 2016, 27, 170–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cowan, M.M. Plant products as antimicrobial agents. Clin. Microbiol. Rev. 1999, 12, 564–582. [Google Scholar] [CrossRef] [Green Version]
- Peana, A.T.; D’Aquila, P.S.; Chessa, M.L.; Moretti, M.D.L.; Serra, G.; Pippia, P. (-)-Linalool produces antinociception in two experimental models of pain. Eur. J. Pharmacol. 2003, 460, 37–41. [Google Scholar] [CrossRef]
- Li, X.-J.; Yang, Y.-J.; Li, Y.-S.; Zhang, W.K.; Tang, H.-B. α-Pinene, linalool, and 1-octanol contribute to the topical anti-inflammatory and analgesic activities of frankincense by inhibiting COX-2. J. Ethnopharmacol. 2016, 179, 22–26. [Google Scholar] [CrossRef]
- Bakir, B.; Him, A.; Özbek, H.; Düz, E.; Tütüncü, M. Investigation of the anti-inflammatory and analgesic activities of β-caryophyllene. Int. J. Essent. Oil Ther. 2008, 2, 41–44. [Google Scholar]
- Klauke, A.; Racz, I.; Pradier, B.; Markert, A.; Zimmer, A.M.; Gertsch, J.; Zimmer, A. The cannabinoid CB2 receptor-selective phytocannabinoid beta-caryophyllene exerts analgesic effects in mouse models of inflammatory and neuropathic pain. Eur. Neuropsychopharmacol. 2014, 24, 608–620. [Google Scholar] [CrossRef] [Green Version]
- Santos, F.A.; Rao, V.S.N. Antiinflammatory and antinociceptive effects of 1,8-cineole a terpenoid oxide present in many plant essential oils. Phyther. Res. 2000, 14, 240–244. [Google Scholar] [CrossRef]
- Takaishi, M.; Fujita, F.; Uchida, K.; Yamamoto, S.; Shimizu, M.S.; Uotsu, C.H.; Shimizu, M.; Tominaga, M. 1,8-Cineole, a TRPM8 agonist, is a novel natural antagonist of human TRPA1. Mol. Pain 2012, 8, 86. [Google Scholar] [CrossRef] [Green Version]
- Eccles, R. Menthol and related cooling compounds. J. Pharm. Pharmacol. 1994, 46, 618–630. [Google Scholar] [CrossRef]
- Kamatou, G.P.P.; Vermaak, I.; Viljoen, A.M.; Lawrence, B.M. Menthol: A simple monoterpene with remarkable biological properties. Phytochemistry 2013, 96, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Laude, E.A.; Morice, A.H.; Grattan, T.J. The antitussive effects of menthol, camphor, and cineole in conscious Guinea-pigs. Pulm. Pharmacol. 1994, 7, 179–184. [Google Scholar] [CrossRef] [PubMed]
- Gavliakova, S.; Dolak, T.; Licha, H.; Krizova, S.; Plevkova, J. Cineole, thymol and camphor nasal challenges and their effect on nasal symptoms and cough in an animal model. Acta Medica Martiniana 2013, 13, 5–13. [Google Scholar] [CrossRef] [Green Version]
- Gavliakova, S.; Biringerova, Z.; Buday, T.; Brozmanova, M.; Calkovsky, V.; Poliacek, I.; Plevkova, J. Antitussive effects of nasal thymol challenges in healthy volunteers. Respir. Physiol. Neurobiol. 2013, 187, 104–107. [Google Scholar] [CrossRef] [PubMed]
- Snow Setzer, M.; Sharifi-Rad, J.; Setzer, W.N. The search for herbal antibiotics: An in-silico investigation of antibacterial phytochemicals. Antibiotics 2016, 5, 30. [Google Scholar] [CrossRef] [Green Version]
- Setzer, M.S.; Byler, K.G.; Ogungbe, I.V.; Setzer, W.N. Natural products as new treatment options for trichomoniasis: A molecular docking investigation. Sci. Pharm. 2017, 85, 5. [Google Scholar] [CrossRef] [Green Version]
- Pan, Y.; Huang, N.; Cho, S.; MacKerell, A.D. Consideration of molecular weight during compound selection in virtual target-based database screening. J. Chem. Inf. Comput. Sci. 2003, 43, 267–272. [Google Scholar] [CrossRef]
- Huang, N.; Nagarsekar, A.; Xia, G.; Hayashi, J.; MacKerell, A.D. Identification of non-phosphate-containing small molecular weight inhibitors of the tyrosine kinase p56 Lck SH2 domain via in silico screening against the pY + 3 binding site. J. Med. Chem. 2004, 47, 3502–3511. [Google Scholar] [CrossRef]
- Carta, G.; Knox, A.J.S.; Lloyd, D.G. Unbiasing scoring functions: A new normalization and rescoring strategy. J. Chem. Inf. Model. 2007, 47, 1564–1571. [Google Scholar] [CrossRef]








{kind=link}
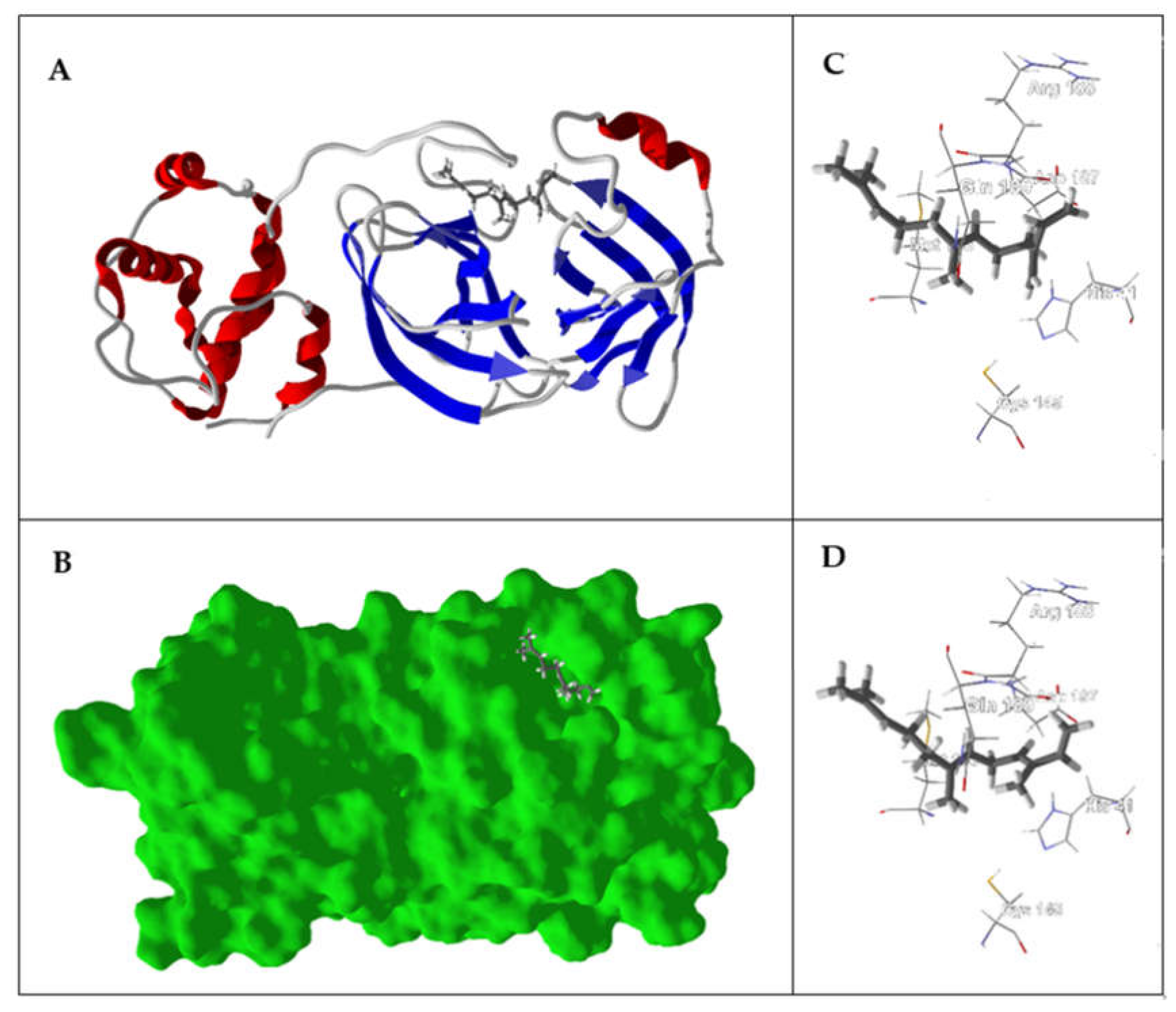{kind=link}
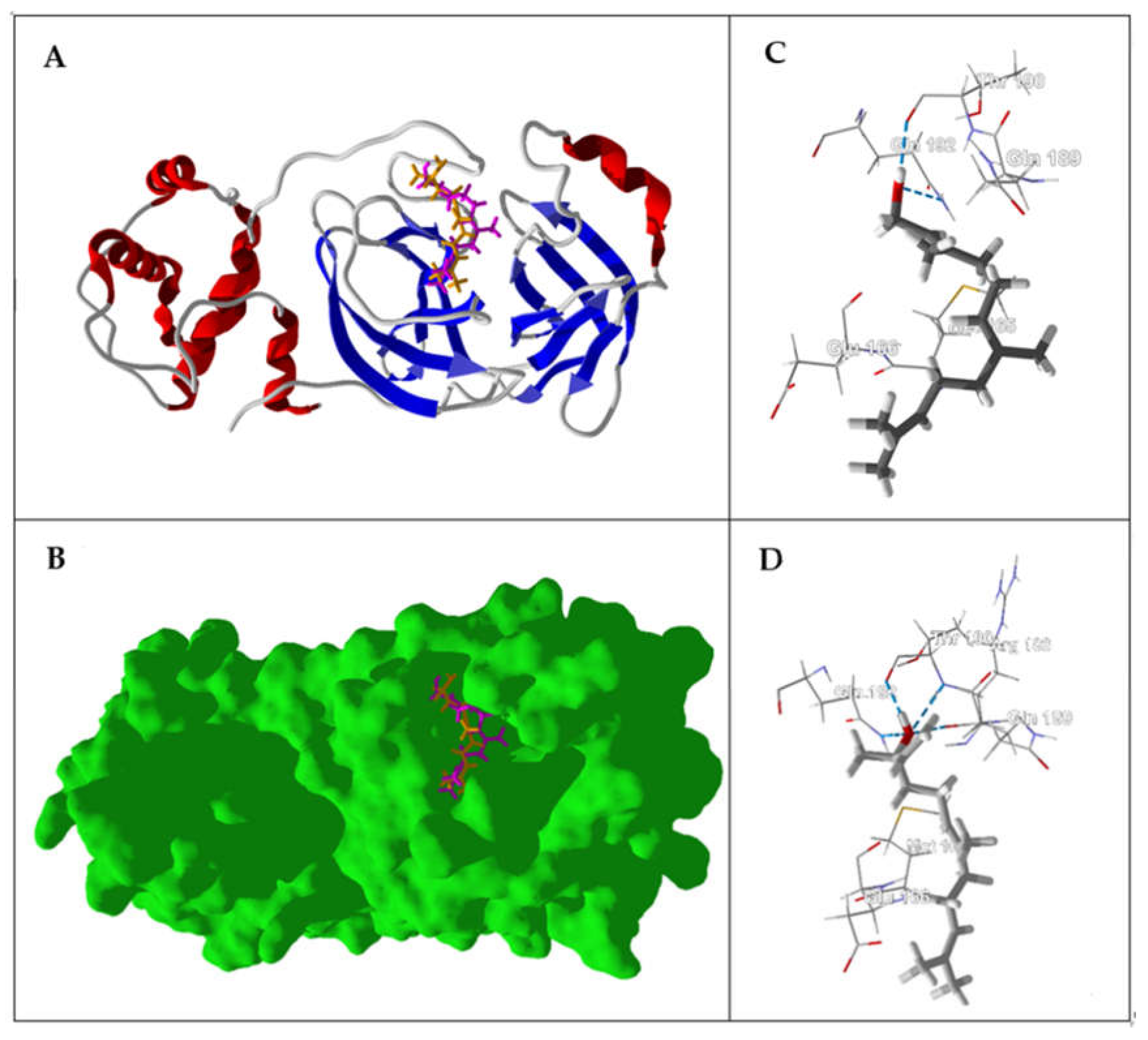{kind=link}
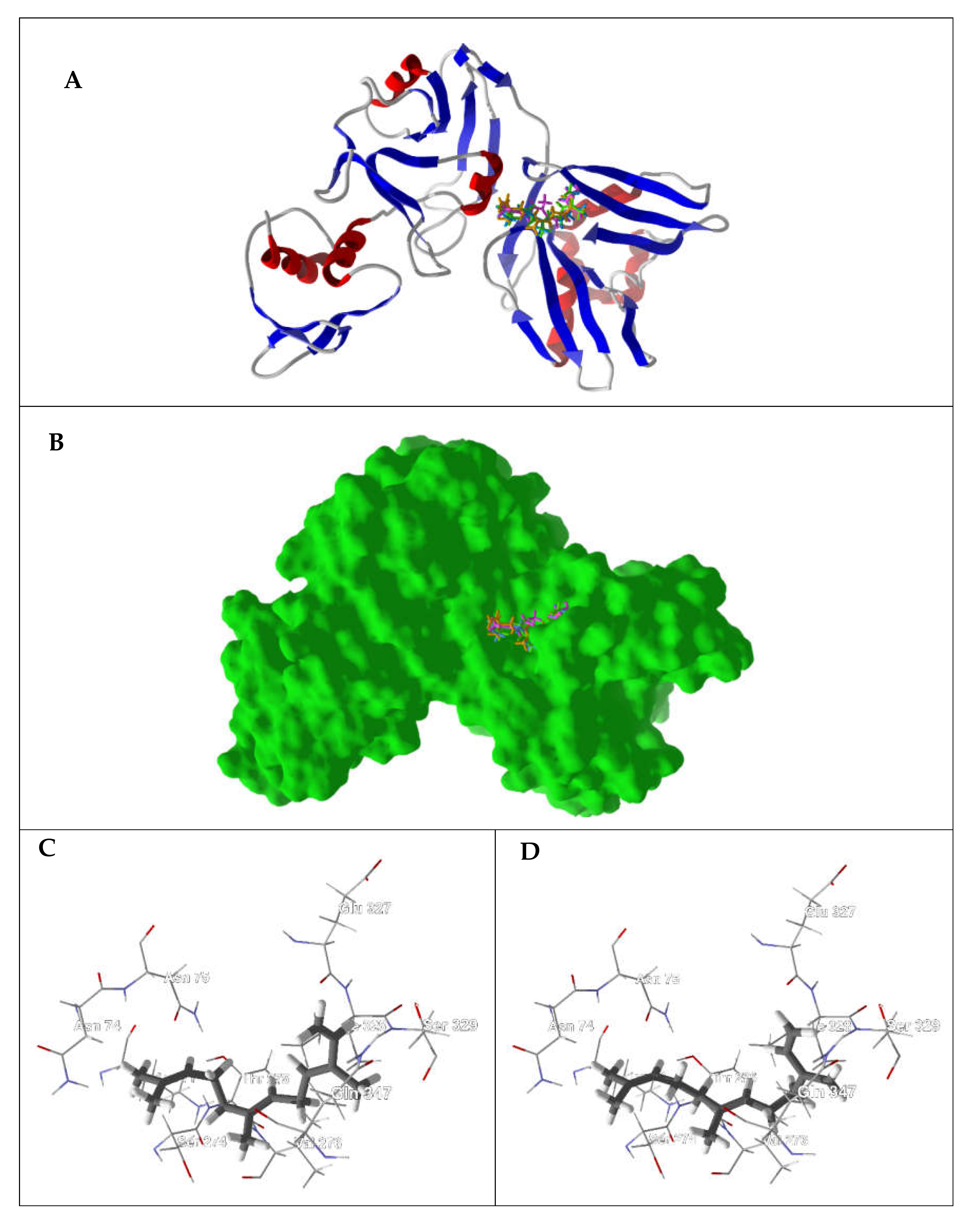{kind=link}
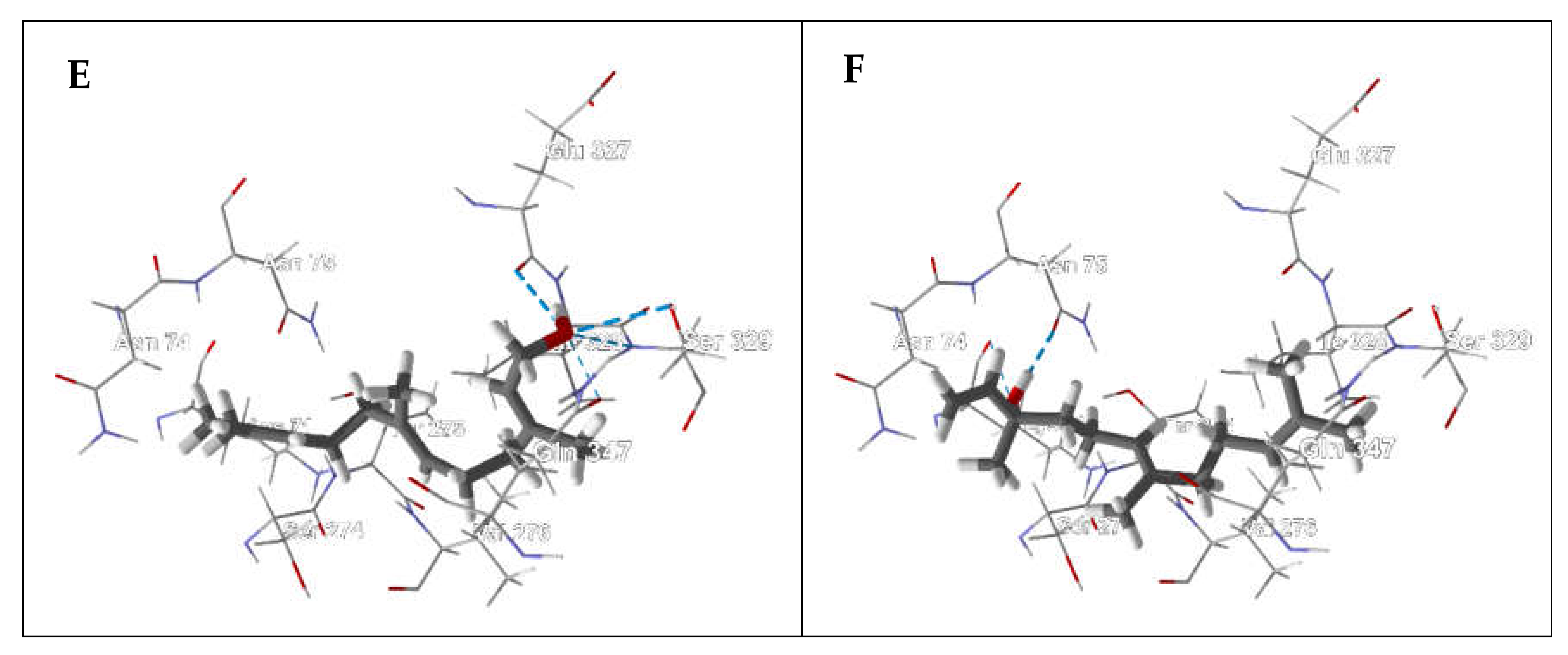{kind=link}
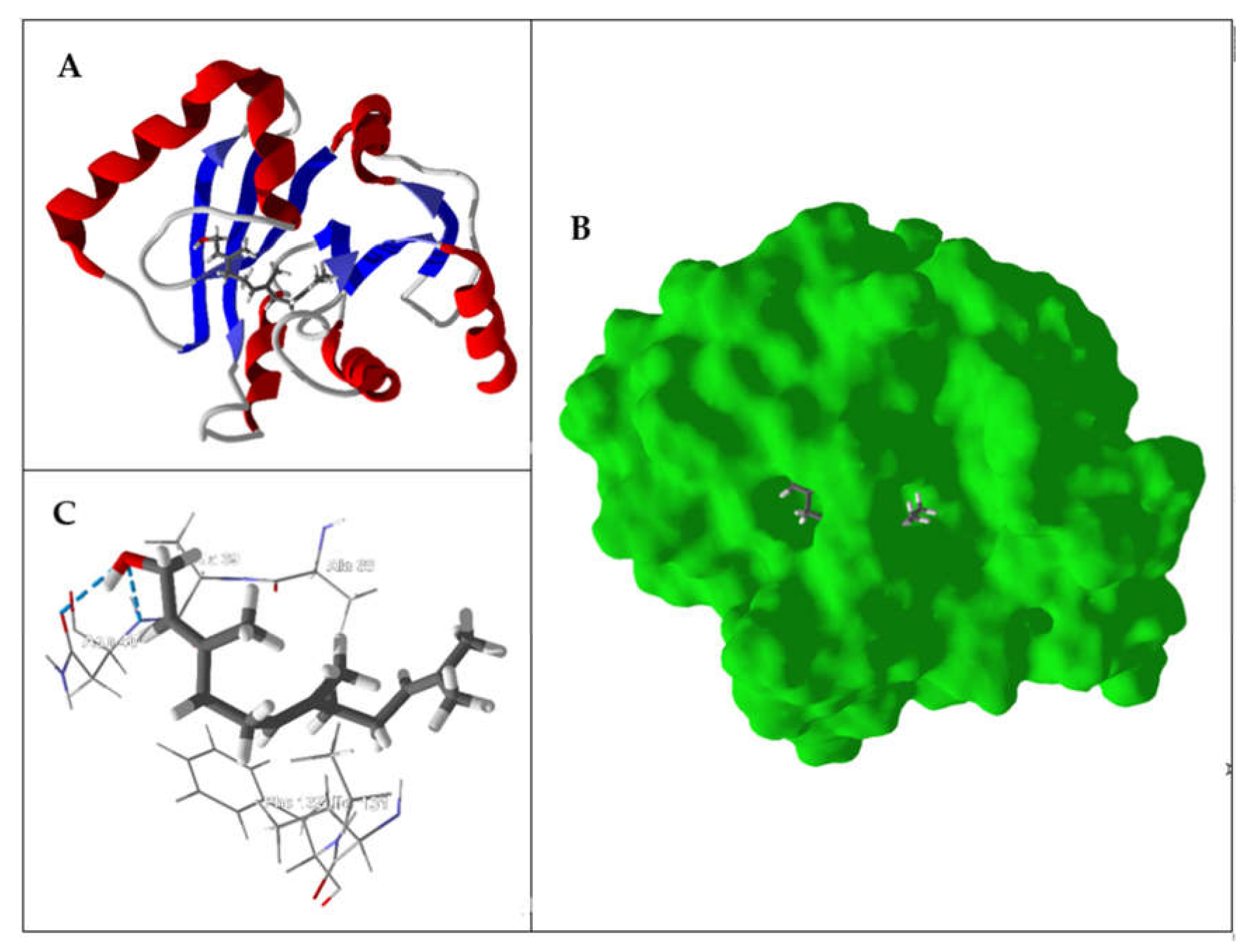{kind=link}
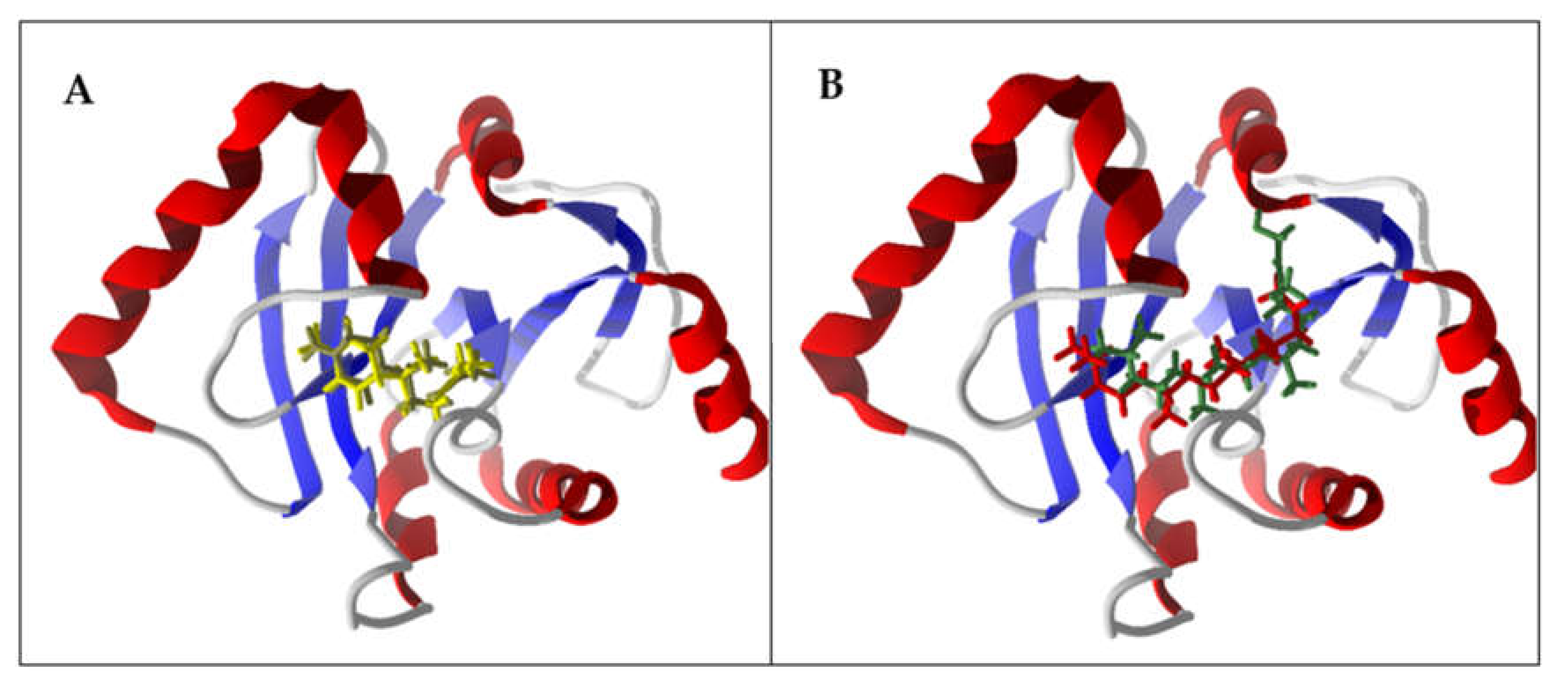{kind=link}
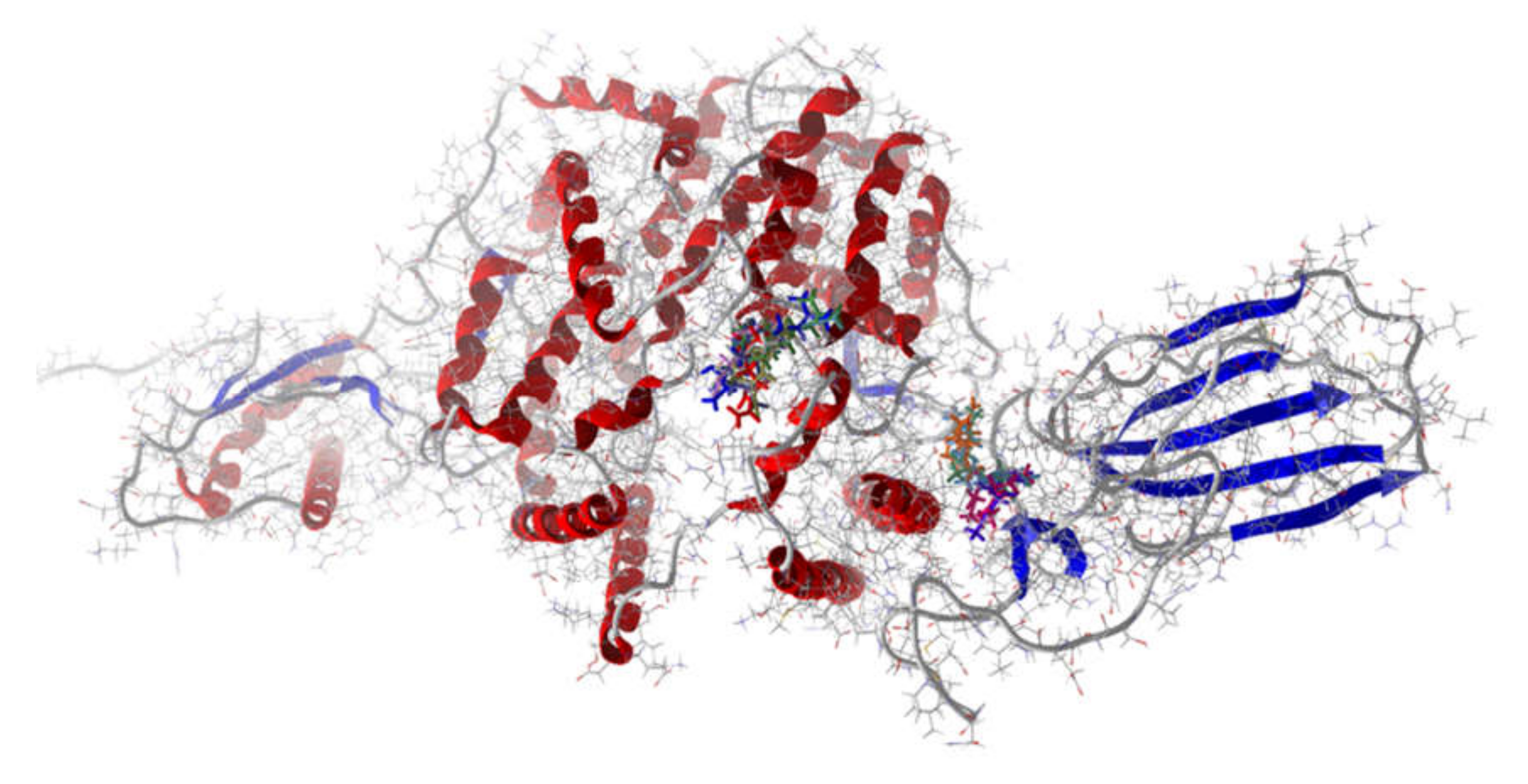{kind=link}
| Essential Oil | Major Components | Assay | IC50 (μg/mL) | Reference | |
|---|---|---|---|---|---|
| Aloysia gratissima (Gillies & Hook.) Tronc. | caryophyllene oxide (15.8%), guaiol (17.4%) a, chrysanthenyl acetate (5.6%), and limonene oxide (5.3%) | Plaque reduction assay (Vero cells), HSV-1 | 65.0 | [37] | |
| Aloysia triphylla Royle | α-thujone (22.9%), cis−carveol (17.5%), carvone (13.2%), and limonene (12.7%) | Plaque reduction assay (Vero cells), HSV-1 | >250 | [38] | |
| Artemisia arborescens L. | camphor (35.7%), β-thujone (24.0%), and chamazulene (7.7%) | Plaque reduction assay (Vero cells), HSV-1 | 25% inhibition at 100 μg/mL | [39] | |
| Artemisia arborescens L. | Artemisia arborescens L. | β-thujone (45.0%), camphor (6.8%), and chamazulene (22.7%) c | Plaque reduction assay (Vero cells), HSV-1, HSV-2 | 2.4, 4.1 | [40] |
| Artemisia douglasiana Besser | α-thujone (68.3%) and β-thujone (7.5%) | Plaque reduction assay (Vero cells), HSV-1 | 83 | [37] | |
| Artemisia kermanensis Podlech (syn. Seriphidium kermanense (Podlech) K. Bremer & Humphries | α-thujone (13.8%), camphor (10.2%), and β-thujone (6.2%) | Plaque reduction assay (Vero cells), HSV-1 | 40 | [41] | |
| Artemisia mendozana DC. | camphor (22.4%), artemiseole (11.7%), artemisia alcohol (10.8%), and borneol (7.2%) | Plaque reduction assay, DENV−2, JUNV and HSV-1 | 129.3, 178.6, and 153.7 | [38] | |
| Artemisia princeps Pamp. | borneol (12.1%), α-thujone (8.7%), τ−cadinol, (6.7%), and 1,8-cineole (6.2%) | Plaque reduction assay, murine norovirus−1 | 64% inhibition at 0.01% | [42] | |
| Plaque reduction assay, feline calicivirus−F9 | 48% inhibition at 0.1% | ||||
| Artemisia vulgaris L. | α-thujone (38.1%), β-thujone (10.6%), and 1,8-cineole (8.8%) | Virus yield assay (Vero cells), yellow fever virus (YFV) | 100 μg/mL (100% inhibition) | [43] | |
| Ayapana triplinervis (Vahl) R.M. King & H. Rob. | thymohydroquinone dimethyl ether | Plaque reduction assay (A549 cells), Zika virus | 38.0 | [44] | |
| Buddleja cordobensis Griseb. | caryophyllene oxide (32.1%), β-caryophyllene (16.5%), and α-copaene (8.5%) | Plaque reduction assay, DENV−2, JUNV and HSV-1 | 86.4, 39.0, and 54.1 | [38] | |
| Cedrus libani A. Rich. | himachalol (22.5%), β-himachalene (21.9%), and α-himachalene (10.5%) | Cytopathic effect (CPE) on Vero cells, HSV-1 | 440 | [45] | |
| Cinnamomum zeylanicum Blume (syn. Cinnamomum verum J. Presl) | eugenol (75–85%), linalool (1.6−8.5%), (E)−cinnamaldehyde (0.6−1.5%), (E)−cinnamyl acetate (0.7–2.6%), β-caryophyllene (0.5–6.7%), eugenyl acetate (0.1–2.9%), and benzyl benzoate (0.1–8.3%) | Influenza type A (H1N1) | [46] | ||
| Cinnamomum zeylanicum Blume (syn. Cinnamomum verum J. Presl) | (E)−cinnamaldehyde (63.9%), eugenol (7.0%), and (E)−cinnamyl acetate (5.1%) c | Plaque reduction assay (HeLa cells), HSV-2 | 82 | [47] | |
| Citrus × bergamia Risso & Poit. | limonene (23–55%), linalool (2–37%), linalyl acetate (12–41%), β-pinene (up to 10%), and γ-terpinene (up to 10%) | Influenza virus type A H1N1 | 100% inhibition at 0.3% | [46] | |
| Citrus limonum Risso | limonene (54.6%), γ-terpinene (19.1%), and β-pinene (14.5%) d | Plaque reduction assay (Vero cells), HSV-1 | 2500 | [48] | |
| Citrus reshni Hort ex Tan. (leaf EO) | sabinene (40.5%), linalool (23.3%), and terpinen-4-ol (8.3%) | Plaque reduction assay, influenza A virus H5N1 | 19.4% inhibition at 0.1 μL/mL | [36] | |
| Citrus reshni Hort ex Tan. (unripe fruit peel EO) | limonene (82.4%) and linalool (7.2%) | Plaque reduction assay, influenza A virus H5N1 | 61.5% inhibition at 1.5 μL/mL | [36] | |
| Citrus reshni Hort ex Tan. (ripe fruit peel EO) | limonene (91.6%) | Plaque reduction assay, influenza A virus H5N1 | 50% inhibition at 1.5 μL/mL | [36] | |
| Cupressus sempervirens L. | α-pinene (51.9%) and δ-3-carene (24.9%) c | Plaque reduction assay (Vero cells), HSV-1 | 6600 | [48] | |
| Cymbopogon citratus (DC.) Stapf | geranial (40.2%), neral (30.6%), and geraniol (6.7%) c | Plaque reduction assay (Vero cells), HSV-1 | 91 | [48] | |
| Cymbopogon flexuosus (Nees) Will. Watson | geranial (48–54%) and neral (29–33%) | Influenza virus type A (H1N1) | [46] | ||
| Cynanchum stauntonii (Decne.) Schltr. ex H. Lév. | (2E,4E)-decadienal (23.0%), γ-nonalactone (4.2%), 5-pentyl-2(3H)−furanone (3.8%), and 3-isopropyl-1-pentanol (3.5%) | Influenza type A (H1N1) | 64 | [46] | |
| Dysphania ambrosioides (L.) Mosyakin & Clemants | cis-ascaridole (60.7%) and m-cymene (22.2%) | Plaque reduction assay, Coxsackie virus B4 | 21.75 | [49] | |
| Eucalyptus astringens (Maiden) Maiden | 1,8-cineole (42.0%), α-pinene (22.0%), and trans−pinocarveol (7.0%) | Vero cells, Coxsakie virus B3 | 13.0 | [50] | |
| Eucalyptus bicostata Maiden, Blakely & Simmonds | 1,8-cineole (68.0%), globulol (5.4%), and trans−pinocarveol (4.6%) | Vero cells, Coxsakie virus B3 | 13.6 | [50] | |
| Eucalyptus caesia Benth. | 1,8-cineole (40.2%), p-cymene (14.1%), γ-terpinene (12.4%), α-pinene (7.7%), and terpinen-4-ol (5.6%) | Plaque reduction assay (Vero cells), HSV-1 | 70 | [41] | |
| Eucalyptus camaldulensis Dehnh. | α-terpinene (26.3%), α-terpineol (9.1%), and camphene (8.5%) e | Plaque reduction assay, Rotavirus Wa strain | 50% inhibition at 10% EO concentration | [51] | |
| Eucalyptus camaldulensis Dehnh. | α-terpinene (26.3%), α-terpineol (9.1%), and camphene (8.5%) e | Plaque reduction assay, Coxsackie virus B4 | 53.3% at 10% EO concentration | [51] | |
| Eucalyptus camaldulensis Dehnh. | α-terpinene (26.3%), α-terpineol (9.1%), and camphene (8.5%) e | Plaque reduction assay, HSV-1 | 90% at 10% EO concentration | [51] | |
| Eucalyptus camaldulensis Dehnh. | α-terpinene (26.3%), α-terpineol (9.1%), and camphene (8.5%) e | Plaque reduction assay, adenovirus type 7 | 0% at 10% EO concentration | [51] | |
| Eucalyptus cinereal F. Muell. ex Benth. | 1,8-cineole (70.4%), α-pinene (4.5%), and limonene (3.7%) | Vero cells, Coxsakie virus B3 | 13.0 | [50] | |
| Eucalyptus globulus Labill. | 1,8-cineole (63.8%) and α-pinene (14.0%) f | Plaque reduction assay (Vero cells), HSV-1 | 1700 | [48] | |
| Eucalyptus globulus Labill. | 1,8-cineole (63.8%) and α-pinene (14.0%) f | Plaque reduction assay (RC-37 cells), HSV-1 | 55.0 | [52] | |
| Eucalyptus lehmannii (Schauer) Benth. | 1,8-cineole (59.6%), α-pinene (17.6%), and α-terpineol (8.7%) | Vero cells, Coxsakie virus B3 | 11.5 | [50] | |
| Eucalyptus leucoxylon F. Muell. | 1,8-cineole (59.2%), α-pinene (7.8%), and α-terpineol (4.3%) | Vero cells, Coxsakie virus B3 | 8.1 | [50] | |
| Eucalyptus maidenii F. Muell. | 1,8-cineole (57.8%), p-cymene (7.4%), and α-pinene (7.3%) | Vero cells, Coxsakie virus B3 | 14.5 | [50] | |
| Eucalyptus odorata Behr | cryptone (20.9%), p-cymene (16.7%), phellandral (6.6%), and cuminal (6.6%) | Vero cells, Coxsakie virus B3 | 19.2 | [50] | |
| Eucalyptus sideroxylon A. Cunn. ex Woolls | 1,8-cineole (69.2%), α-pinene (6.9%), and α-terpineol (5.4%) | Vero cells, Coxsakie virus B3 | 12.3 | [50] | |
| Eupatorium patens D. Don ex Hook. & Arn. | germacrene D (36.2%), β-caryophyllene (14.1%), and bicyclogermacrene (7.0%) | Plaque reduction assay (Vero cells), HSV-1 | 125 | [37] | |
| Fortunella margarita (Lour.) Swingle (leaf EO) | α-terpineol (55.5%), carvone (5.7%), carveol (5.5%), γ-muurolene (5.5%), and citronellal (5.0%) | MTT assay, reasserted avian influenza A virus, H5N1 | 6.77 | [53] | |
| Fortunella margarita (Lour.) Swingle (fruit EO) | β-eudesmol (28.3%), α-muurolene (10.3%), β-gurjunene (10.0%), γ-eudesmol (8.4%), and γ-muurolene (6.6%) | MTT assay, reasserted avian influenza A virus, H5N1 | 38.89 | [53] | |
| Gaillardia megapotamica (Spreng.) Baker | β-pinene (24.2%), (Z)-β-ocimene (16.5%), α-pinene (7.7%), limonene (7.5%), and β-caryophyllene (6.7%) | Plaque reduction assay, DENV−2, JUNV and HSV-1 | 140.6, 49.8, and 99.1 | [38] | |
| Heterothalamus alienus (Spreng.) Kuntze | β-pinene (35.5%), spathulenol (10.7%), and germacrene D (6.8%) | Plaque reduction assay DENV−2, JUNV and HSV-1 | 122.3, 44.2 and 148.4 | [38] | |
| Heterotheca latifolia Buckley | borneol (40.0%), camphor (24.3%), and limonene (5.1%) | Plaque reduction assay (Vero cells), HSV-1 | >150 | [37] | |
| Houttuynia cordata Thunb. | decanal (3.4−8.9%), decanol (up to 7.0%), 2−undecanone (23.0−36.1%), decanoic acid (1.4−6.3%), dodecanal (up to 7.3%), and 2−tridecanone (2.6−5.6%) | Influenza type A (H1N1) | 48 | [54] | |
| Hyptis mutabilis (Rich.) Briq. | fenchone (17.1%), 1,8-cineole (12.6%), β-caryophyllene (10.9%), bicyclogermacrene (8.7%), and germacrene D (6.2%) | Plaque reduction assay (Vero cells), HSV-2 (Human Herpesvirus type 1) | 79.01 | [55] | |
| Hyptis mutabilis (Rich.) Briq. | germacrene D (15.1%), β-caryophyllene (13.3%), curzerene (13.3%), and bicyclogermacrene (13.2%) | Plaque reduction assay (Vero cells) | >150 | [37] | |
| Hyssopus officinalis L. | cis−pinocamphone (40.1%), trans−pinocamphone (13.3%), β-pinene (10.7%), and β-phellandrene (5.3%) c | Plaque reduction assay (RC-37 cells), HSV-1 | 1 | [56] | |
| Hyssopus officinalis L. | cis−pinocamphone (40.1%), trans−pinocamphone (13.3%), β-pinene (10.7%), and β-phellandrene (5.3%) c | Plaque reduction assay (RC-37 cells), HSV-2 | 6 | [57] | |
| Illicium verum Hook. f. | (E)−anethole (90.5%) c | Plaque reduction assay (RC-37 cells), HSV-1 | 40 | [58] | |
| Illicium verum Hook. f. | (E)−anethole (90.5%) c | Plaque reduction assay (RC-37 cells), HSV-1 | 1 | [59] | |
| Illicium verum Hook. f. | (E)−anethole (90.5%) c | Plaque reduction assay (RC-37 cells), HSV-2 | 30 | [57] | |
| Jungia polita Griseb. | caryophyllene oxide (9.2%) and β-caryophyllene (8.1%) | Plaque reduction assay DENV−2, JUNV and HSV-1 | 39.8, 134.2 and 136.4 | [38] | |
| Juniperus communis L. | α-pinene (46.7%), myrcene (15.0%), sabinene (13.2%), and limonene (7.0%) c | Plaque reduction assay (Vero cells), HSV-1 | >10000 | [48] | |
| Lavandula angustifolia Mill. | linalyl acetate (37.0−43.6%), linalool (19.7−39.1%), geraniol (up to 9.3%), β-caryophyllene (up to 5.1%), terpinene−4−ol (up to 14.9%), lavandulol (up to 1.5%), lavandulyl acetate (up to 5.5%), 1,8-cineole (up to 4.1%), and borneol (up to 6.4%) | 85% in vitro inhibition of influenza type A (H1N1) | [46] | ||
| Lavandula latifolia Medik. | linalool (31.9%), 1,8-cineole (18.8%), and borneol (10.1%) g | Plaque reduction assay (Vero cells), HSV-1 | 2200 | [48] | |
| Lepechinia salviifolia (Kunth) Epling | camphor (10.3%), limonene (9.7%), p−mentha−1(7),8−diene (7.4%), α-pinene (6.9%), and γ-terpinene (6.7%) | Plaque reduction assay (Vero cells), HSV-1, HSV-2 | 68.8, 81.9 | [55] | |
| Lepechinia vulcanicola J.R.I. Wood | limonene (18.9%), germacrene D (10.4%), 1−octen−3−ol (8.8%), β-caryophyllene (8.7%), and α-pinene (8.2%) | Plaque reduction assay (Vero cells), HSV-1, HSV-2 | 112, 68.9 | [55] | |
| Leptospermum scoparium J.R. Forst. & G. Forst. | calamene (16.0%), leptospermone (14.5%), δ−cadinene (6.1%), flavesone (4.5%), viridiflorene (4.4%), and isoleptospermone (3.9%) | Plaque reduction assay (RC-37 cells), HSV-1, HSV-2 | 0.96, 0.58 | [60] | |
| Lippia alba (Mill.) N.E. Br. ex Britton & P. Wilson | carvone (51.0%), Limonene (33%), and bicyclosesquiphellandrene (7.0%) | Virus yield assay (Vero cells), yellow fever virus (YFV) | 100 μg/mL (100% inhibition) | [43] | |
| Lippia alba (Mill.) N.E. Br. ex Britton & P. Wilson | carvone (39.7%), limonene (30.6%), and bicyclosesquiphellandrene (8.9%) | Plaque reduction assay (Vero cells), DENV−1, DENV−2, DENV−3, DENV−4, YFV 17 DD | 10.1, 0.4, 32.6, 21.1, 4.9 | [61] | |
| Lippia alba (Mill.) N.E. Br. ex Britton & P. Wilson | carvone (39.7%), limonene (30.6%), and bicyclosesquiphellandrene (8.9%) | Plaque reduction assay (Vero cells), Yellow fever (YFV) | 4.3 | [62] | |
| Lippia citriodora Kunth (syn. Aloysia citriodora Palau) | geranial (18.9%), neral (15.6%), limonene (10.7%), and 1,8-cineole (5.0) | Plaque reduction assay (Vero cells), DENV−1, DENV−2, DENV−3, DENV−4, YFV 17 DD | 1.9, 2.9, 2.6, 33.7, 5.7 | [61] | |
| Lippia citriodora Kunth (syn. Aloysia citriodora Palau) | geranial (18.9%), neral (15.6%), limonene (10.7%), and 1,8-cineole (5.0) | Plaque reduction assay (Vero cells), Yellow fever (YFV) | 19.4 | [62] | |
| Lippia graveolens Kunth | carvacrol (56.8%), o−cymene (32.1%), and γ-terpinene (3.7%) h | MTT assay (Mardin−Darby bovine kidney cells), HSV-1, ACVR−HSV-1 (acyclovir−resistant HSV-1), HRSV (human respiratory syncytial virus) | 99.6, 55.9, 68.0 | [63] | |
| Lippia junelliana (Moldenke) Tronc. | piperitenone oxide (= rotundifolone) (36.5%), limonene (23.1%), camphor (7.9%), and spathulenol (6.5%) | Plaque reduction assay (Vero cells), HSV-1 | >150 | [37] | |
| Lippia origanoides | carvacrol (44.0%), thymol (15.0%), and γ-terpinene (10.0%) | Virus yield assay (Vero cells), yellow fever virus (YFV) | 11.1 μg/mL (100% inhibition) | [43] | |
| Lippia turbinata Griseb. | limonene (60.6%), piperitenone oxide (17.4%), and β-caryophyllene (6.4%) | Plaque reduction assay (Vero cells), HSV-1 | > 150 | [37] | |
| Matricaria recutita L. | α-bisabolol oxide A (13.4–55.9%), α-bisabolol oxide B (8.4–25.1%), bisabolone oxide A (2.9–11.4%), cis−bicycloether (= (Z)−spiroether) (3.6–17.7%), and (E)-β-farnesene (1.9–10.4%) i | Plaque reduction assay (RC-37 cells), HSV-1 | 0.3 | [58] | |
| Matricaria recutita L. | α-bisabolol oxide A (13.4–55.9%), α-bisabolol oxide B (8.4–25.1%), bisabolone oxide A (2.9−11.4%), cis−bicycloether (= (Z)−spiroether) (3.6–17.7%), and (E)-β-farnesene (1.9–10.4%) i | Plaque reduction assay (RC-37 cells), HSV-2 | 1.5 | [57] | |
| Melaleuca alternifolia Cheel | terpinen-4-ol (36.71%), γ-terpinene (22.20%), and α-terpinene (10.10%) | Plaque reduction assay, influenza A⁄PR ⁄ 8 virus subtype H1N1 | 60 | [31] | |
| Melaleuca alternifolia Cheel | terpinen-4-ol (30–48%), γ-terpinene (10–28%), α-terpinene (5–13%), 1,8-cineole (up to 15%), terpinolene (1.5−5%), p-cymene (0.5–12%), α-pinene (1–6%), and α-terpineol (1.5–8%) | 100% inhibition of influenza type A (H1N1) virus at 0.01%; type A (H11N9) virus to tea tree oil vapor caused 100% inhibition | [46] | ||
| Melaleuca alternifolia Cheel | terpinen-4-ol (47.5%), γ-terpinene (20.2%), and α-terpinene (8.6%) c | Plaque reduction assay (Vero cells), HSV-1 | 2700 | [48] | |
| Melaleuca alternifolia Cheel | terpinen-4-ol (47.5%), γ-terpinene (20.2%), and α-terpinene (8.6%) c | Plaque reduction assay (Vero cells), HSV-1 | 13.2 | [64] | |
| Melaleuca alternifolia Cheel | terpinen-4-ol (47.5%), γ-terpinene (20.2%), and α-terpinene (8.6%) c | Plaque reduction assay (RC-37 cells), HSV-1 | 2 | [52] | |
| Melaleuca alternifolia Cheel | terpinen-4-ol (47.5%), γ-terpinene (20.2%), and α-terpinene (8.6%) c | Plaque reduction assay (HeLa cells), HSV-2 | 2700 | [47] | |
| Melaleuca alternifolia Cheel | terpinen-4-ol (47.5%), γ-terpinene (20.2%), and α-terpinene (8.6%) c | Plaque reduction assay (Vero cells), HSV-1 | significant plaque reduction at 10 and 5% v/v | [65] | |
| Melaleuca armillaris (Sol. ex Gaertn.) Sm. | 1,8-cineole (33.9%), terpinen-4-ol (18.8%), and γ-terpinene (10.4%) | Plaque reduction assay (Vero cells), HSV-1 | 99% plaque reduction (concentration not given) | [66] | |
| Melaleuca ericifolia Sm. | methyl eugenol (96.84%) | Plaque reduction assay (Vero cells), HSV-1 | 91.5% plaque reduction (concentration not given) | [66] | |
| Melaleuca leucadendra (L.) L. | 1,8-cineole (64.3%), α-terpineol (11.0%), and valencene (3.91%) | Plaque reduction assay (Vero cells), HSV-1 | 92% plaque reduction (concentration not given) | [66] | |
| Melissa officinalis L. | neral (17−32%), geranial (23–43%), linalool (up to 9.0%), citronellal (0.7–20.3%), geraniol (up to 23.2%), β-caryophyllene (up to 11.3%), and caryophyllene oxide (0.4−31.7%) | influenza A virus (H9N2) | Significant reduction at 5 μg/mL | [46] | |
| Melissa officinalis L. | β-cubebene (15.4%), β-caryophyllene (14.2%), α-cadinol (7.2%), geranial (6.6%), and neral (5.8%) | Plaque reduction assay (HEp−2 cells), HSV-2 | 21 | [67] | |
| Melissa officinalis L. | geranial (20.1%), β-caryophyllene (17.3%), and neral (13.6%) | Plaque reduction assay (RC-37 cells), HSV-1, HSV-2 | 4, 0.8 | [68] | |
| Mentha × piperita L. | menthol (42.8%), menthone (14.6%), and isomenthone (5.9%) | Plaque reduction assay (RC-37 cells), HSV-1, HSV-2 | 20, 8 | [69] | |
| Mentha × piperita L. | menthol (43.8%), menthone (19.7%), menthyl acetate (6.5%), and 1,8-cineole (5.0%) c | Plaque reduction assay (Vero cells), HSV-1 | 2200 | [48] | |
| Mentha suaveolens Ehrh. | limonene (7.4%), isopulegol (12.0%), and piperitenone oxide (41.8%) | Plaque reduction assay, cytopathogenic murine norovirus | 0.87 | [70] | |
| Mentha suaveolens Ehrh. | piperitenone oxide (86.9%) | Plaque reduction assay (Vero cells), HSV-1 | 5.1 | [64] | |
| Minthostachys mollis Griseb. | cis−piperitone epoxide (29.9%), piperitenone oxide (25.6%), menthone (7.4%), germacrene D (5.8%), and pulegone (5.5%) | Plaque reduction assay (Vero cells), HSV-1, HSV-2 | 70.7, 68.0 | [55] | |
| Ocimum basilicum var. album (L.) Benth. | linalool (53.8%) and eugenol (12.6%) j | Plaque reduction assay (Vero cells), HSV-1 | >10000 | [48] | |
| Ocimum campechianum Mill. | methyl eugenol (53.9%), β-caryophyllene (13.0%), α-bulnesene (5.4%), germacrene D (3.4%), and α-humulene (3.3%) | Plaque reduction assay (Vero cells), HSV-2 | 74.33 | [55] | |
| Origanum elongatum Emb. (leaf EO) | p-cymene (16.2%), γ-terpinene (13.5%), thymol (14.2%), and carvacrol (19.2%) | Plaque reduction assay, cytopathogenic murine norovirus | 0.37 | [70] | |
| Origanum elongatum Emb. (inflorescence EO) | p-cymene (16.1%), γ-terpinene (7.3%), and carvacrol (40.1%), | Plaque reduction assay, cytopathogenic murine norovirus | 0.75 | [70] | |
| Origanum majorana L. | terpinen-4-ol (28.9%), γ-terpinene (14.9%), trans−sabinene hydrate (9.5%), α-terpinene (8.7%), and sabinene (7.2%) c | Plaque reduction assay (Vero cells), HSV-1 | 2800 | [48] | |
| Origanum majorana L. | terpinen-4-ol (28.9%), γ-terpinene (14.9%), trans−sabinene hydrate (9.5%), α-terpinene (8.7%), and sabinene (7.2%) c | Plaque reduction assay (HeLa cells), HSV-2 | 520 | [47] | |
| Origanum vulgare L. | trans−sabinene hydrate (21.0%), thymol (11.0%), and carvacrol methyl ether (11.0%) | Virus yield assay (Vero cells), yellow fever virus (YFV) | 100 μg/mL (100% inhibition) | [43] | |
| Osmunda regalis L. | hexahydrofarnesyl acetone ( = phytone) (11.8%), 2,4−di−t−butylphenol (6.8%), phytol (6.5%), hexadecene (4.1%), and octadecene (4.4%) | Plaque reduction assay, Coxsackie virus B4 type 2 | 2.24 | [71] | |
| Pectis odorata Griseb. | limonene (50.2%), neral (27.2%), and geranial (23.6%) | Plaque reduction assay DENV−2, JUNV and HSV-1 | 39.6, 36.6, and 71.5 | [38] | |
| Pelargonium graveolens L’Hér. ex Aiton | citronellol (21.9–37.5%), citronellyl formate (9.8–20.6%), geraniol (6.0–16.5%), geranyl formate (1.5–6.5%), menthone (up to 13%), isomenthone (up to 9.9%), and linalool (0.8−14.9%) | influenza type A (H1N1) | 95% inhibition at 0.3% | [46] | |
| Pinus mugo Turra | δ-3-carene (23.9%), α-pinene (17.9%), β-pinene (7.8%), and β-phellandrene (7.2%) k | Plaque reduction assay (RC-37 cells), HSV-1 | 7 | [58] | |
| Ravensara aromatica Sonn. (syn. Cryptocarya agathophylla van der Werff) | 1,8-cineole (52.6%), α-terpineol (12.4%), and sabinene (11.0%) l | Plaque reduction assay (Vero cells), HSV-1 | 2800 | [48] | |
| Rosmarinus officinalis L. | α-pinene (23.9%), verbenone (15.4%), camphor (11.0%), camphene (8.7%), p-cymene (7.5%), and 3−octanone (5.6%) | Plaque reduction assay (Vero cells), HSV-1 | 60 | [41] | |
| Rosmarinus officinalis L. | 1,8-cineole (45.9%), α-pinene (12.0%), camphor (10.9%), and β-pinene (6.3%) b | Plaque reduction assay (Vero cells), HSV-1 | 2700 | [48] | |
| Salvia fruticosa Mill. | 1,8-cineole (47.5%), camphor (9.0%), β-thujone (7.6%), and α-thujone (4.3%) | Plaque reduction assay (Vero cells), HSV-1 and HSV-2 | 1300 | [72] | |
| Santalum album L. | (Z)−α-santalol (45.2%), (Z)-β-santalol (25.4%), and (Z)−trans−α-bergamotol (7.8%) c | Plaque reduction assay (Vero cells), HSV-1, HSV-2 | 22.7, 45.3 | [73] | |
| Santalum album L. | (Z)−α-santalol (45.2%), (Z)-β-santalol (25.4%), and (Z)−trans−α-bergamotol (7.8%) c | Plaque reduction assay (RC-37 cells), HSV-1 | 2 | [56] | |
| Santalum album L. | (Z)−α-santalol (45.2%), (Z)-β-santalol (25.4%), and (Z)−trans−α-bergamotol (7.8%) c | Plaque reduction assay (RC-37 cells), HSV-2 | 5 | [57] | |
| Santolina insularis (Gennari ex Fiori) Arrigoni | artemisia ketone (21.2%), allo−aromadendrene (12.7%), 1,8-cineole (9.0%), and camphene (8.5%) m | Plaque reduction assay (Vero cells), HSV-1 and HSV-2 | 0.88, 0.7 | [74] | |
| Satureja hortensis L. | carvacrol (32.4%), γ-terpinene (32.0%), thymol (10.0%), and p-cymene (6.6%) | Plaque reduction assay (Vero cells), HSV-1 | 80 | [41] | |
| Tessaria absinthioides (Hook. & Arn.) DC. | caryophyllene oxide (12.2%), (E)-β-damascenone, γ-eudesmol (8.5%), α-gurjunene (5.8%), and terpinen-4-ol (5.4%) | Plaque reduction assay (Vero cells), HSV-1 | 105 | [37] | |
| Thymus capitatus (L.) Hoffmanns. & Link (unripe fruit EO) | carvacrol (68.6%), p-cymene (4.8%), γ-terpinene (3.0%), and β-caryophyllene (2.9%) | Plaque reduction assay, cytopathogenic murine norovirus | 0.49 | [70] | |
| Thymus capitatus (L.) Hoffmanns. & Link (ripe fruit EO) | carvacrol (58.8%), p-cymene (5.6%), γ-terpinene (2.8%), and β-caryophyllene (2.6%) | Plaque reduction assay, cytopathogenic murine norovirus | 0.50 | [70] | |
| Thymus vulgaris L. | thymol (31–50%), p-cymene (0.1–26.6%), and γ-terpinene (up to 22.8%) | 100% inhibition of type A (H1N1) virus at 0.3% concentration | [46] | ||
| Thymus vulgaris L. | thymol (43.9%), carvacrol (14.4%), p-cymene (10.5%), β-caryophyllene (7.0%), and γ-terpinene (5.1%) c | Plaque reduction assay (RC-37 cells), HSV-1 | 10 | [56] | |
| Thymus vulgaris L. | thymol (43.9%), carvacrol (14.4%), p-cymene (10.5%), β-caryophyllene (7.0%), and γ-terpinene (5.1%) c | Plaque reduction assay (RC-37 cells), HSV-1 | 11 | [52] | |
| Thymus vulgaris L. | thymol (43.9%), carvacrol (14.4%), p-cymene (10.5%), β-caryophyllene (7.0%), and γ-terpinene (5.1%) c | Plaque reduction assay (RC-37 cells), HSV-2 | 7 | [57] | |
| Thymus willdenowii Boiss. | 1,8-cineole (34.62%), camphor (18.55%), α-pinene (9.46%), and camphene (5.38%) | Plaque reduction assay, Coxsackie virus | Inactive | [75] | |
| Trachyspermum ammi (L.) Sprague | thymol (35–60%), α-pinene, p-cymene, and limonene | Plaque reduction assay, Japanese encephalitis virus | 80% reduction at 500 μg/mL | [76] | |
| Zataria multiflora Boiss. | thymol (47.3%), carvacrol (21.9%), p-cymene (8.6%), γ-terpinene (4.2%), and β-caryophyllene (3.0%) | Real time PCR (H9N2 subtype of AIV) | Reduced viral replication in trachea of broiler chickens | [77] | |
| Zataria multiflora Boiss. | thymol (33.1%), carvacrol (25.9%), and p-cymene (11.3%) | Plaque reduction assay (Vero cells), HSV-1 | 30 | [41] | |
| Zingiber officinale Roscoe | α-zingiberene (32.1%), ar−curcumene (15.2%), β-sesquiphellandrene (10.9%), α-farnesene (7.2%), and α-phellandrene (4.4%) | Plaque reduction assay, Caprine alphaherpesvirus type I | not determined | [78] | |
| Zingiber officinale Roscoe | α-zingiberene (26.4%), camphene (12.6%), β-sesquiphellandrene (9.2%), ar−curcumene (6.5%), β-phellandrene (6.2%), and β-bisabolene (5.1%) c | Plaque reduction assay (RC-37 cells), HSV-1 | 2 | [56] | |
| Zingiber officinale Roscoe | α-zingiberene (26.4%), camphene (12.6%), β-sesquiphellandrene (9.2%), ar−curcumene (6.5%), β-phellandrene (6.2%), and β-bisabolene (5.1%) c | Plaque reduction assay (RC-37 cells), HSV-2 | 1 | [57] |
| Essential Oil Component | Assay | IC50 (μg/mL) | Reference |
|---|---|---|---|
| (E)−Anethole | Plaque reduction assay (RC-37 cells), HSV-1 | 20 | [59] |
| Camphor | Plaque reduction assay (Vero cells), HSV-1 | 2600 | [72] |
| Carvacrol | MTT assay (Mardin−Darby bovine kidney (MDBK) cells, HSV-1 | 48.6 | [63] |
| β-Caryophyllene | Plaque reduction assay (RC-37 cells), HSV-1 | 0.25 | [59] |
| Caryophyllene oxide | Plaque reduction assay (RC-37 cells), HSV-1 | 0.7 | [59] |
| 1,8-Cineole | Plaque reduction assay (Vero cells), HSV-1 | 1800 | [72] |
| 1,8-Cineole | Plaque reduction assay (RC-37 cells), HSV-1 | 1200 | [52] |
| (E)-Cinnamaldehyde | Influenza type A (H1N1) virus | 70% at a concentration of 0.53% after 3 h | [95] |
| (E)-Cinnamaldehyde | H1N1 in-vivo by inhalation in a mouse model | 89% inhibition | [95] |
| Citral (Geranial + Neral) | Yellow fever (YFV), Vero cells | 17.6 | [62] |
| Citral (Geranial + Neral) | Plaque reduction assay (RC-37 cells), HSV-1 | 3.50 | [52] |
| p-Cymene | Influenza, HSV-1, HSV-2, ECHO 9, Cox B1, Polio 1, Adeno 6 | >500 | [31] |
| p-Cymene | Plaque reduction assay (RC-37 cells), HSV-1 | 16 | [52] |
| p-Cymene | Influenza type A (H1N1) virus | Inactive | [25] |
| Dodecanal | Influenza type A (H1N1) virus | 51 | [54] |
| β-Eudesmol | Plaque reduction assay (Vero cells), HSV-1 | 6 | [59] |
| Eugenol | Plaque reduction assay (RC-37 cells), HSV-1 | 35 | [59] |
| Eugenol | Plaque reduction assay (RC-37 cells), HSV-1 | 25.6 | [96] |
| Farnesol | Plaque reduction assay (RC-37 cells), HSV-1 | 3.5 | [59] |
| Germacrone | Influenza type A (H1N1) virus | 1.22–1.55 | [97] |
| Germacrone | Influenza type A (H3N2) virus | 0.34 | [97] |
| Germacrone | Influenza type B virus | 1.38 | [97] |
| Isoborneol | Plaque reduction assay (Vero cells), HSV-1 | <1000 | [98] |
| Nerolidol (natural) | Plaque reduction assay (RC-37 cells), HSV-1 | 4.2 | [99] |
| Nerolidol (synthetic) | Plaque reduction assay (RC-37 cells), HSV-1 | 1.5 | [99] |
| Octanal | Influenza type A (H1N1) virus | 15 | [54] |
| Patchouli alcohol | Influenza type A (H1N1) | 89% inhibition at 10 μg/mL | [100] |
| Patchouli alcohol | Influenza type A (H2N2) | 0.89 | [101] |
| Patchouli alcohol | Influenza type A (H2N2), in-vivo test using a mouse model | 70% survival rate at a dose of 5 mg/(kg day) | [101] |
| α-Pinene | Plaque reduction assay (RC-37 cells), HSV-1 | 4.5 | [52] |
| Piperitenone oxide | Plaque reduction assay (Vero cells) | 1.4 | [64] |
| α-Terpinene | Influenza type A (H1N1) virus | Inactive | [25] |
| α-Terpinene | Influenza, HSV-1, HSV-2, ECHO 9, Cox B1, Polio 1, Adeno 4 | >12 | [31] |
| α-Terpinene | Plaque reduction assay (RC-37 cells), HSV-1 | 8.5 | [52] |
| γ-Terpinene | Plaque reduction assay (RC-37 cells), HSV-1 | 7 | [52] |
| γ-Terpinene | Influenza, HSV-1, HSV-2, ECHO 9, Cox B1, Polio 1, Adeno 5 | >120 | [31] |
| γ-Terpinene | influenza type A (H1N1) virus | inactive | [25] |
| Terpinen-4-ol | Influenza, HSV-1, HSV-2, ECHO 9, Cox B1, Polio 1, Adeno 2 | 25 (influenza) >50 (others) | [31] |
| Terpinen-4-ol | Plaque reduction assay (Vero cells), HSV-1 | 60 | [52] |
| α-Terpineol | Plaque reduction assay (RC-37 cells), HSV-1 | 22 | [52] |
| Terpinolene | Influenza, HSV-1, HSV-2, ECHO 9, Cox B1, Polio 1, Adeno 3 | 12 (influenza) >12 (others) | [31] |
| Thujones (α & β) | Plaque reduction assay (RC-37 cells), HSV-1 | 400 | [72] |
| Thymol | Influenza type A (H1N1) virus | Active | [25] |
| Thymol | Plaque reduction assay (RC-37 cells), HSV-1 | 30 | [52] |
| 2-Undecanone | influenza type A (H1N1) virus | 62 | [54] |
| Compound | Main Protease | Endoribo- Nuclease | ADP Ribose Phosphatase | RNA-Dependent RNA Polymerase | Spike Protein Binding Domain | Angiotensin- Converting Enzyme a |
|---|---|---|---|---|---|---|
| (E)-Anethole | −84.9 | −83.0 | −97.5 | −74.0 | −65.2 | −83.8 |
| allo-Aromadendrene | −86.6 | −86.9 | −95.8 | −74.9 | −66.1 | −85.3 |
| Artemiseole | −89.2 | −83.4 | −96.6 | −73.0 | −67.8 | −78.0 |
| (R)-Artemisia alcohol | −84.3 | −78.3 | −88.2 | −66.8 | −61.1 | −74.7 |
| (S)-Artemisia alcohol | −86.1 | −85.4 | −98.5 | −69.5 | −67.1 | −77.4 |
| Artemisia ketone | −91.0 | −85.7 | −97.9 | −71.9 | −66.9 | −77.3 |
| Ascaridole | −74.8 | −68.2 | −86.0 | −65.2 | −62.6 | −64.4 |
| Benzyl benzoate | −104.9 | −96.5 | −110.3 | −82.5 | −74.9 | −96.4 |
| (Z)-trans-α-Bergamotol | −98.0 | −91.2 | −105.9 | −71.9 | −63.6 | −83.6 |
| Bicyclogermacrene | −85.9 | −88.0 | −92.5 | −75.6 | −68.2 | −86.1 |
| Bicyclosesquiphellandrene | −79.4 | −89.6 | −86.0 | −61.8 | −62.5 | −78.2 |
| α-Bisabolol oxide A | −88.5 | −87.6 | −89.1 | −71.5 | −63.5 | −89.7 |
| α-Bisabolol oxide B | −96.6 | −92.2 | −101.0 | −79.7 | −75.2 | −95.6 |
| Bisabolone oxide A | −93.7 | −87.9 | −98.2 | −73.3 | −71.0 | −78.7 |
| (+)-Borneol | −77.4 | −91.8 | −100.1 | −74.1 | −70.9 | −73.3 |
| (−)-Borneol | −71.4 | −71.5 | −78.4 | −56.7 | −50.1 | −73.7 |
| (+)-iso-Borneol | −75.6 | −75.3 | −75.9 | −60.8 | −53.2 | −71.8 |
| (−)-iso-Borneol | −73.8 | −69.2 | −75.6 | −54.8 | −49.3 | −72.4 |
| α-Bulnesene | −95.2 | −72.4 | −78.4 | −57.0 | −57.8 | −101.7 |
| δ-Cadinene | −91.6 | −90.2 | −112.6 | −75.2 | −60.0 | −93.4 |
| τ-Cadinol | −92.3 | −82.8 | −82.5 | −69.9 | −69.8 | −87.8 |
| (R)-Calamene | −92.3 | −83.5 | −97.4 | −69.0 | −65.6 | −87.3 |
| (S)-Calamene | −88.4 | −86.8 | −95.9 | −70.2 | −64.7 | −87.5 |
| (+)-Camphene | −85.6 | −78.9 | −83.2 | −64.4 | −57.9 | −72.2 |
| (−)-Camphene | −77.2 | −74.1 | −87.1 | −66.0 | −59.8 | −67.1 |
| (+)-Camphor | −75.2 | −73.1 | −77.9 | −63.9 | −53.8 | −69.4 |
| (−)-Camphor | −72.5 | −70.0 | −75.6 | −64.3 | −52.8 | −70.8 |
| (+)-δ-3-Carene | −87.9 | −78.6 | −87.0 | −65.0 | −72.4 | −75.8 |
| (−)-δ-3-Carene | −83.9 | −88.0 | −90.4 | −66.8 | −67.3 | −73.4 |
| Carvacrol | −84.5 | −86.6 | −94.8 | −74.1 | −71.0 | −81.2 |
| Carvacrol methyl ether | −85.5 | −82.5 | −103.1 | −74.6 | −65.8 | −83.7 |
| (+)-cis−Carveol | −87.0 | −81.8 | −97.7 | −76.5 | −69.7 | −80.4 |
| (−)-cis−Carveol | −85.6 | −85.0 | −95.6 | −76.8 | −74.3 | −81.2 |
| (R)-Carvone | −87.7 | −82.4 | −98.2 | −74.7 | −69.2 | −83.4 |
| (S)-Carvone | −86.2 | −83.2 | −98.9 | −73.2 | −66.3 | −82.2 |
| (E)-Caryophyllene | −81.2 | −82.2 | −93.9 | −73.4 | −59.2 | −75.1 |
| Caryophyllene oxide | −80.6 | −86.7 | −97.0 | −74.1 | −66.5 | −83.3 |
| Cedrol | −82.3 | −84.4 | −80.1 | −69.7 | −58.3 | −69.9 |
| epi-Cedrol | −76.4 | −88.4 | −92.2 | −68.5 | −55.5 | −75.9 |
| Chamazulene | −97.6 | −96.4 | −110.9 | −76.9 | −73.7 | −95.6 |
| (−)-cis-Chrystanthyl acetate | −83.2 | −77.5 | −87.1 | −71.5 | −60.2 | −80.9 |
| (−)-trans−Chrysanthenyl acetate | −77.0 | −81.4 | −85.3 | −65.9 | −68.9 | −72.4 |
| 1,8-Cineole | −72.7 | −67.6 | −71.5 | −58.1 | −58.6 | −61.0 |
| (E)-Cinnamaldehyde | −93.1 | −85.8 | −100.0 | −76.4 | −73.1 | −81.0 |
| (E)-Cinnamyl acetate | −99.4 | −88.0 | −108.9 | −76.9 | −80.6 | −89.5 |
| (R)-Citronellal | −99.9 | −91.6 | −105.0 | −75.1 | −73.3 | −88.0 |
| (S)-Citronellal | −98.3 | −88.9 | −107.4 | −72.1 | −71.4 | −86.2 |
| (R)-Citronellol | −99.9 | −90.2 | −104.9 | −75.7 | −72.3 | −84.9 |
| (S)-Citronellol | −99.2 | −92.2 | −107.6 | −74.2 | −77.4 | −85.1 |
| (R)-Citronellyl formate | −105.5 | −92.6 | −109.9 | −72.8 | −76.2 | −90.5 |
| (S)-Citronellyl formate | −101.5 | −96.4 | −114.7 | −81.9 | −78.3 | −91.0 |
| α-Copaene | −84.1 | −78.9 | −85.9 | −60.2 | −59.6 | −77.1 |
| Costunolide | −94.5 | −99.2 | −110.2 | −78.2 | −74.9 | −92.6 |
| (R)-Cryptone | −86.6 | −82.4 | −93.5 | −68.2 | −70.4 | −74.8 |
| (S)-Cryptone | −81.5 | −82.9 | −94.9 | −69.8 | −70.6 | −76.6 |
| β-Cubebene | −90.8 | −92.0 | −86.7 | −76.0 | −71.2 | −89.8 |
| Cuminaldehyde | −86.2 | −80.5 | −103.3 | −74.9 | −67.5 | −81.7 |
| ar-Curcumene | −105.3 | −94.0 | −108.6 | −82.8 | −75.8 | −96.4 |
| Curcumol | −83.0 | −93.4 | −91.2 | −73.5 | −69.6 | −86.0 |
| Curdione | −86.2 | −98.1 | −99.0 | −77.3 | −71.7 | −92.0 |
| Curzerene | −91.5 | −86.0 | −104.2 | −76.8 | −67.6 | −90.5 |
| m-Cymene | −83.6 | −81.3 | −92.8 | −72.0 | −67.6 | −79.0 |
| o-Cymene | −83.2 | −78.3 | −87.7 | −63.9 | −63.3 | −72.9 |
| p-Cymene | −79.7 | −78.7 | −91.0 | −70.3 | −63.9 | −75.4 |
| (E)-β-Damascenone | −85.2 | −87.6 | −110.2 | −73.8 | −73.8 | −84.8 |
| (2E,4E)-Decadienal | −105.7 | −97.7 | −112.5 | −81.8 | −78.7 | −90.0 |
| Dodecanal | −102.8 | −93.9 | −101.9 | −77.9 | −73.3 | −94.9 |
| Eremanthin | −97.7 | −97.7 | −98.0 | −83.7 | −81.3 | −100.9 |
| β-Eudesmol | −97.6 | −87.4 | −106.5 | −75.8 | −64.5 | −74.7 |
| γ-Eudesmol | −93.6 | −89.9 | −94.7 | −69.4 | −66.1 | −84.3 |
| Eugenol | −93.2 | −91.7 | −105.2 | −80.0 | −79.1 | −88.4 |
| Eugenol methyl ether | −88.5 | −85.3 | −111.1 | −71.7 | −68.6 | −89.3 |
| Eugenyl acetate | −96.3 | −89.3 | −115.1 | −75.6 | −70.6 | −94.9 |
| (E,E)-α-Farnesene | −115.0 | −107.5 | −112.8 | −86.8 | −85.3 | −100.3 |
| (E)-β-Farnesene | −115.4 | −105.0 | −116.3 | −87.1 | −82.9 | −100.7 |
| (E,E)-Farnesol | −112.4 | −104.6 | −121.4 | −89.6 | −80.8 | −100.9 |
| (+)-Fenchone | −80.0 | −80.9 | −87.1 | −66.1 | −60.8 | −66.9 |
| (-)-Fenchone | −83.3 | −67.5 | −86.2 | −66.5 | −60.9 | −68.8 |
| Flavesone | −82.3 | −84.0 | −95.7 | −68.9 | −64.4 | −78.9 |
| Geranial | −101.7 | −90.8 | −113.0 | −76.1 | −74.5 | −92.0 |
| Geraniol | −103.5 | −98.5 | −110.2 | −77.4 | −76.4 | −93.8 |
| Geranyl formate | −105.9 | −93.9 | −111.4 | −80.8 | −80.3 | −96.2 |
| Germacrene D | −92.1 | −96.7 | −110.5 | −77.1 | −73.0 | −87.3 |
| Germacrone | −85.1 | −97.1 | −94.9 | −71.6 | −67.1 | −80.9 |
| Guiaol | −94.1 | −92.5 | −113.4 | −79.2 | −79.4 | −91.9 |
| α-Gurjunene | −89.9 | −83.2 | −81.3 | −71.5 | −61.3 | −79.8 |
| β-Gurjunene | −77.4 | −64.1 | −81.1 | −64.1 | −58.0 | −75.1 |
| α-Himachalene | −80.4 | −89.7 | −86.5 | −63.7 | −67.9 | −74.3 |
| β-Himachalene | −85.5 | −81.3 | −88.6 | −68.1 | −62.9 | −77.2 |
| Himachalol | −77.2 | −88.3 | −100.8 | −63.6 | −63.7 | −75.6 |
| α-Humulene | −88.9 | −90.9 | −89.1 | −74.9 | −70.6 | −86.7 |
| Leptospermone | −85.5 | −83.2 | −92.9 | −71.2 | −64.1 | −80.0 |
| iso-Leptospermone | −86.1 | −82.7 | −93.7 | −69.1 | −62.6 | −83.5 |
| (R)-Limonene | −82.2 | −76.2 | −92.0 | −72.6 | −67.3 | −79.0 |
| (S)-Limonene | −82.2 | −73.8 | −92.2 | −72.4 | −66.1 | −77.5 |
| (R,S,R)-Limonene oxide | −86.3 | −77.7 | −100.8 | −76.7 | −68.1 | −85.1 |
| (S,R,R)-Limonene oxide | −83.5 | −87.4 | −89.4 | −66.7 | −63.4 | −76.4 |
| (S,R,S)-Limonene oxide | −84.7 | −80.7 | −95.8 | −74.9 | −66.7 | −83.8 |
| (R)-Linalool | −96.0 | −89.3 | −101.4 | −70.9 | −79.0 | −87.0 |
| (S)-Linalool | −100.7 | −87.5 | −102.1 | −71.4 | −70.4 | −87.8 |
| (R)-Linalyl acetate | −101.4 | −88.9 | −105.1 | −73.0 | −74.3 | −82.5 |
| (S)-Linalyl acetate | −102.8 | −90.8 | −106.0 | −68.5 | −74.5 | −82.3 |
| p-Mentha-1(7),8-diene | −82.3 | −82.2 | −94.3 | −69.5 | −65.9 | −79.3 |
| (+)-Menthol | −82.9 | −86.2 | −95.0 | −71.8 | −66.6 | −77.7 |
| (-)-Menthol | −82.3 | −84.5 | −95.1 | −71.1 | −69.2 | −79.4 |
| Menthone | −83.2 | −77.8 | −94.7 | −69.3 | −70.0 | −74.4 |
| iso-Menthone | −81.0 | −79.8 | −97.6 | −64.4 | −63.5 | −80.1 |
| α-Muurolene | −86.9 | −81.9 | −96.9 | −72.0 | −66.0 | −83.7 |
| (+)-γ-Muurolene | −82.3 | −82.9 | −83.1 | −72.2 | −66.6 | −84.4 |
| (-)-γ-Muurolene | −88.4 | −86.2 | −106.7 | −74.5 | −68.5 | −86.8 |
| Myrcene | −98.7 | −90.1 | −102.2 | −74.9 | −71.4 | −84.3 |
| Neral | −102.6 | −91.8 | −110.5 | −81.5 | −72.4 | −91.2 |
| (E)-Nerolidol | −110.7 | −101.4 | −113.8 | −83.5 | −76.1 | −100.6 |
| (E)-β-Ocimene | −97.0 | −90.2 | −103.6 | −76.3 | −75.6 | −86.7 |
| (Z)-β-Ocimene | −98.3 | −88.7 | −103.2 | −78.6 | −73.9 | −85.1 |
| Octanal | −89.7 | −83.8 | −99.5 | −71.3 | −72.4 | −80.5 |
| (R)-2-Octen-3-ol | −88.1 | −88.4 | −96.6 | −71.9 | −70.7 | −81.9 |
| (S)-1-Octen-3-ol | −94.2 | −90.7 | −93.4 | −77.1 | −74.7 | −80.0 |
| 3-Octanone | −88.9 | −85.3 | −94.7 | −71.7 | −72.9 | −80.3 |
| Patchouli alcohol | −63.5 | −57.1 | −66.7 | −51.8 | −43.0 | −67.9 |
| (R)-Phellandral | −87.6 | −85.9 | −102.5 | −74.4 | −66.2 | −81.1 |
| (S)-Phellandral | −87.7 | −83.7 | −101.6 | −74.4 | −66.5 | −79.2 |
| (R)-α-Phellandrene | −81.1 | −82.9 | −92.5 | −70.1 | −65.3 | −76.4 |
| (S)-α-Phellandrene | −81.6 | −82.5 | −92.3 | −69.7 | −65.6 | −76.6 |
| (R)-β-Phellandrene | −84.4 | −83.7 | −94.0 | −69.8 | −65.5 | −75.2 |
| (S)-β-Phellandrene | −82.9 | −84.5 | −94.0 | −71.2 | −67.5 | −77.7 |
| Phytol | −106.3 | −94.2 | −118.9 | −74.9 | −75.6 | −93.3 |
| Phytone | −106.4 | −94.3 | −116.9 | −79.8 | −72.3 | −90.4 |
| (+)-α-Pinene | −79.0 | −70.7 | −79.7 | −59.4 | −54.6 | −63.7 |
| (-)-α-Pinene | −77.3 | −70.4 | −72.9 | −61.3 | −54.7 | −63.6 |
| (+)-β-Pinene | −76.8 | −71.8 | −79.4 | −63.7 | −53.0 | −65.2 |
| (-)-β-Pinene | −78.8 | −73.6 | −73.9 | −61.9 | −56.1 | −64.7 |
| (+)-Pinocamphone | −74.9 | −72.6 | −79.4 | −63.2 | −62.4 | −66.4 |
| (-)-Pinocamphone | −73.2 | −79.3 | −81.8 | −62.0 | −61.4 | −67.2 |
| (+)-iso-Pinocamphone | −74.9 | −75.0 | −74.9 | −66.6 | −55.1 | −66.5 |
| (-)-iso-Pinocamphone | −77.1 | −80.7 | −77.5 | −64.0 | −55.6 | −68.3 |
| (+)-cis-Pinocarveol | −73.9 | −78.1 | −79.5 | −67.7 | −57.4 | −70.2 |
| (-)-cis-Pinocarveol | −74.1 | −76.5 | −78.8 | −67.0 | −58.6 | −66.3 |
| (+)-trans-Pinocarveol | −74.7 | −80.5 | −78.7 | −65.8 | −57.2 | −70.6 |
| (-)-trans-Pinocarveol | −77.6 | −80.0 | −82.6 | −60.5 | −55.5 | −70.8 |
| (+)-Piperitone oxide | −82.0 | −81.5 | −98.5 | −66.2 | −65.2 | −77.8 |
| (-)-Piperitone oxide | −81.6 | −83.9 | −94.9 | −68.6 | −60.8 | −82.8 |
| (-)-iso-Pulegol | −82.5 | −85.1 | −98.8 | −75.2 | −66.7 | −79.3 |
| (R)-Pulegone | −84.0 | −83.1 | −96.1 | −66.4 | −65.5 | −78.3 |
| (S)-Pulegone | −83.6 | −78.6 | −93.3 | −67.2 | −66.2 | −78.9 |
| Rotundifolone | −83.1 | −79.0 | −96.6 | −66.4 | −62.1 | −80.7 |
| (+)-Sabinene | −86.4 | −84.0 | −92.2 | −70.5 | −68.6 | −77.4 |
| (-)-Sabinene | −87.8 | −85.9 | −94.7 | −71.5 | −69.6 | −77.0 |
| cis-Sabinene hydrate | −82.4 | −81.1 | −92.4 | −68.0 | −60.8 | −78.2 |
| (Z)-α-Santalol | −105.7 | −95.3 | −108.8 | −84.7 | −70.7 | −95.8 |
| (E)-β-Santalol | −104.8 | −95.6 | −106.2 | −77.0 | −70.9 | −86.6 |
| (Z)-β-Santalol | −104.4 | −94.4 | −106.2 | −79.8 | −73.2 | −92.2 |
| β-Sesquiphellandrene | −103.8 | −99.0 | −115.7 | −84.8 | −75.3 | −101.1 |
| Spathulenol | −90.7 | −88.0 | −98.4 | −77.8 | −67.9 | −90.4 |
| (Z)-Spiroether | −111.8 | −84.9 | −103.1 | −87.1 | −79.3 | −102.0 |
| γ-Terpinene | −81.3 | −79.3 | −93.2 | −71.2 | −65.3 | −76.6 |
| (R)-Terpinen-4-ol | −80.4 | −81.9 | −89.5 | −69.0 | −64.9 | −75.3 |
| (S)-Terpinen-4-ol | −82.1 | −81.8 | −88.0 | −70.4 | −64.4 | −76.4 |
| (R)-α-Terpineol | −82.6 | −88.3 | −91.6 | −65.6 | −66.0 | −72.6 |
| (S)-α-Terpineol | −88.7 | −86.8 | −94.1 | −71.6 | −63.5 | −82.0 |
| Terpinolene | −80.7 | −80.1 | −94.7 | −68.8 | −63.9 | −77.4 |
| (-)-α-Thujone | −87.2 | −89.3 | −94.5 | −66.9 | −69.2 | −78.1 |
| (+)-β-Thujone | −86.9 | −80.5 | −94.2 | −73.7 | −69.7 | −79.2 |
| Thymohydroquinone dimethyl ether | −89.5 | −82.8 | −104.6 | −73.1 | −67.0 | −85.0 |
| Thymol | −84.4 | −87.5 | −94.6 | −72.9 | −70.8 | −78.4 |
| 2-Undecanone | −101.3 | −94.0 | −108.3 | −78.4 | −72.8 | −90.2 |
| Valencene | −84.7 | −81.7 | −92.6 | −74.4 | −68.3 | −81.7 |
| (+)-Verbenone | −84.7 | −73.2 | −82.5 | −68.4 | −54.8 | −71.2 |
| (-)-Verbenone | −83.8 | −77.7 | −77.4 | −65.4 | −57.8 | −68.0 |
| Viridiflorene | −86.8 | −90.2 | −91.0 | −76.3 | −69.4 | −88.8 |
| α-Zingiberene | −106.4 | −100.5 | −115.4 | −82.7 | −71.4 | −98.6 |
| Compound | BtOBP | Cruzain | TcAChE | BaNadD | DrPLA2 | EcAspTA |
|---|---|---|---|---|---|---|
| 1GT3 | 1ME3 | 6G1U | 3HFJ | 1FV0 | 2Q7W | |
| (E)-Anethole | −90.2 | −73.3 | −96.8 | −112.9 | −85.3 | −88.1 |
| allo-Aromadendrene | −85.0 | −70.6 | −92.8 | −105.8 | −87.8 | −79.6 |
| Artemiseole | −84.8 | −74.2 | −84.2 | −90.6 | −68.0 | −71.7 |
| (R)-Artemisia alcohol | −88.4 | −68.3 | −89.5 | −94.8 | −80.7 | −81.5 |
| (S)-Artemisia alcohol | −86.1 | −74.0 | −94.8 | −101.4 | −82.5 | −81.4 |
| Artemisia ketone | −93.1 | −77.3 | −91.5 | −107.9 | −87.8 | −80.6 |
| Ascaridole | −79.7 | −54.4 | −72.1 | −79.7 | −75.0 | −65.3 |
| Benzyl benzoate | −105.0 | −82.8 | −115.8 | −128.1 | −97.3 | −89.9 |
| (Z)-trans-α-Bergamotol | −1 00.7 | −73.6 | −107.0 | −94.2 | −92.6 | −85.8 |
| Bicyclogermacrene | −97.5 | −82.4 | −95.6 | −105.5 | −89.7 | −89.0 |
| Bicyclosesquiphellandrene | −92.2 | −52.5 | −91.2 | −110.7 | −88.4 | −75.7 |
| α-Bisabolol oxide A | −94.7 | −66.1 | −102.8 | −102.6 | −77.1 | −82.7 |
| α-Bisabolol oxide B | −104.6 | −90.4 | −107.0 | −121.9 | −94.2 | −89.8 |
| Bisabolone oxide A | −100.1 | −71.9 | −96.8 | −84.7 | −81.2 | −86.7 |
| (+)-Borneol | −72.5 | −51.0 | −72.0 | −47.8 | −61.4 | −62.5 |
| (-)-Borneol | −78.3 | −53.1 | −70.8 | −49.0 | −63.3 | −56.2 |
| (+)-iso-Borneol | −73.7 | −55.6 | −74.6 | −31.0 | −60.9 | −59.7 |
| (-)-iso-Borneol | −73.2 | −55.3 | −77.7 | −52.6 | −51.0 | −57.4 |
| α-Bulnesene | −93.4 | −89.3 | −109.3 | −115.7 | −94.4 | −95.3 |
| δ-Cadinene | −79.5 | −86.8 | −105.1 | −116.8 | −94.8 | −82.3 |
| Τ-Cadinol | −87.8 | −71.8 | −117.7 | −121.7 | −86.0 | −87.0 |
| (R)-Calamene | −80.3 | −84.4 | −116.5 | −120.4 | −91.3 | −87.7 |
| (S)-Calamene | −80.1 | −82.5 | −116.2 | −121.8 | −93.0 | −88.3 |
| (+)-Camphene | −79.0 | −62.1 | −77.0 | −75.9 | −69.3 | −67.5 |
| (-)-Camphene | −77.7 | −62.8 | −75.2 | −77.4 | −71.3 | −70.6 |
| (+)-Camphor | −76.0 | −57.0 | −75.0 | −49.0 | −53.5 | −54.1 |
| (-)-Camphor | −80.0 | −54.1 | −74.4 | −52.9 | −61.2 | −56.4 |
| (+)-δ-3-Carene | −80.4 | −64.2 | −80.6 | −88.0 | −71.8 | −72.5 |
| (-)-δ-3-Carene | −82.0 | −68.9 | −84.4 | −90.2 | −72.8 | −72.9 |
| Carvacrol | −82.4 | −77.7 | −94.8 | −106.7 | −81.6 | −87.5 |
| Carvacrol methyl ether | −85.6 | −79.3 | −96.6 | −111.5 | −84.2 | −93.6 |
| (+)-cis-Carveol | −89.1 | −77.3 | −96.9 | −109.4 | −81.3 | −87.6 |
| (-)-cis-Carveol | −89.0 | −82.1 | −95.5 | −109.0 | −83.2 | −89.4 |
| (R)-Carvone | −86.0 | −80.7 | −94.6 | −109.9 | −83.5 | −91.5 |
| (S)-Carvone | −88.0 | −77.2 | −94.1 | −105.9 | −82.8 | −87.2 |
| (E)-Caryophyllene | −87.3 | −60.4 | −88.1 | −87.3 | −87.4 | −78.6 |
| Caryophyllene oxide | −95.1 | −73.1 | −90.1 | −97.0 | −83.8 | −77.8 |
| Cedrol | −87.7 | −63.1 | −96.4 | −84.4 | −86.3 | −70.9 |
| epi-Cedrol | −84.0 | −59.7 | −83.3 | −86.7 | −80.0 | −76.2 |
| Chamazulene | −95.2 | −89.2 | −123.0 | −132.1 | −98.9 | −101.6 |
| (-)-cis-Chrystanthyl acetate | −85.8 | −63.7 | −89.8 | −73.8 | −76.5 | −71.5 |
| (-)-trans-Chrysanthenyl acetate | −80.9 | −69.7 | −82.4 | −78.5 | −76.5 | −68.6 |
| 1,8-Cineole | −69.3 | −51.1 | −73.2 | −54.4 | −49.4 | −47.3 |
| (E)-Cinnamaldehyde | −87.7 | −77.6 | −98.2 | −110.1 | −85.7 | −89.5 |
| (E)-Cinnamyl acetate | −95.9 | −88.2 | −107.4 | −132.5 | −94.6 | −103.1 |
| (R)-Citronellal | −98.1 | −90.5 | −109.3 | −120.7 | −88.3 | −105.7 |
| (S)-Citronellal | −95.0 | −89.7 | −108.5 | −123.3 | −86.9 | −104.1 |
| (R)-Citronellol | −91.0 | −90.0 | −108.2 | −122.4 | −88.6 | −107.7 |
| (S)-Citronellol | −92.5 | −91.9 | −108.5 | −122.0 | −90.3 | −100.3 |
| (R)-Citronellyl formate | −98.0 | −93.5 | −118.7 | −129.5 | −87.5 | −108.5 |
| (S)-Citronellyl formate | −99.7 | −95.1 | −111.2 | −135.6 | −91.5 | −94.8 |
| α-Copaene | −88.9 | −69.9 | −84.6 | −78.7 | −81.7 | −61.9 |
| Costunolide | −106.8 | −87.5 | −116.7 | −120.1 | −100.3 | −89.1 |
| (R)-Cryptone | −74.9 | −68.1 | −92.2 | −100.0 | −78.5 | −80.8 |
| (S)-Cryptone | −79.0 | −68.5 | −91.6 | −100.7 | −82.7 | −79.6 |
| β-Cubebene | −99.8 | −67.2 | −106.9 | −112.2 | −93.9 | −94.8 |
| Cuminaldehyde | −84.9 | −79.1 | −95.3 | −109.1 | −90.1 | −89.5 |
| ar-Curcumene | −98.1 | −87.5 | −116.0 | −125.7 | −94.1 | −95.3 |
| Curcumol | −90.1 | −67.6 | −100.4 | −88.0 | −87.6 | −81.5 |
| Curdione | −96.4 | −75.1 | −99.3 | −116.3 | −91.0 | −78.7 |
| Curzerene | −96.4 | −80.3 | −109.4 | −89.7 | −91.1 | −86.4 |
| m-Cymene | −78.9 | −73.1 | −96.1 | −99.1 | −76.6 | −88.0 |
| o-Cymene | −77.0 | −61.9 | −89.8 | −97.8 | −76.2 | −77.2 |
| p-Cymene | −82.8 | −73.2 | −91.3 | −96.8 | −80.9 | −84.3 |
| (E)-β-Damascenone | −90.1 | −80.5 | −100.7 | −105.4 | −88.9 | −80.0 |
| (2E,4E)-Decadienal | −96.4 | −87.0 | −110.4 | −129.7 | −95.8 | −114.8 |
| Dodecanal | −95.8 | −83.7 | −110.3 | −127.3 | −89.2 | −108.9 |
| Eremanthin | −80.1 | −87.9 | −121.3 | −132.4 | −98.7 | −91.8 |
| β-Eudesmol | −95.2 | −76.1 | −98.6 | −114.1 | −85.3 | −87.1 |
| γ-Eudesmol | −94.9 | −78.2 | −106.1 | −105.2 | −89.7 | −89.1 |
| Eugenol | −90.4 | −82.4 | −103.9 | −119.3 | −87.3 | −98.8 |
| Eugenol methyl ether | −93.6 | −78.1 | −106.3 | −116.8 | −89.3 | −94.4 |
| Eugenyl acetate | −94.9 | −83.4 | −109.2 | −126.1 | −100.1 | −104.6 |
| (E,E)-α-Farnesene | −115.5 | −103.7 | −129.8 | −131.8 | −101.2 | −111.4 |
| (E)-β-Farnesene | −112.1 | −103.2 | −122.7 | −131.8 | −105.3 | −108.6 |
| (E,E)-Farnesol | −116.8 | −96.2 | −133.0 | −135.6 | −100.5 | −109.0 |
| (+)-Fenchone | −78.4 | −60.5 | −79.5 | −79.8 | −62.4 | −69.9 |
| (-)-Fenchone | −80.3 | −61.5 | −81.0 | −84.2 | −65.5 | −60.7 |
| Flavesone | −87.1 | −56.0 | −88.6 | −92.1 | −85.2 | −80.3 |
| Geranial | −96.5 | −94.9 | −111.7 | −119.3 | −92.1 | −101.5 |
| Geraniol | −95.5 | −93.6 | −109.7 | −118.0 | −94.3 | −107.3 |
| Geranyl formate | −100.2 | −89.7 | −115.4 | −128.3 | −94.8 | −114.1 |
| Germacrene D | −102.4 | −88.8 | −109.9 | −116.5 | −93.7 | −90.0 |
| Germacrone | −92.4 | −68.3 | −94.5 | −102.9 | −88.3 | −80.0 |
| Guiaol | −100.8 | −88.5 | −113.3 | −107.5 | −92.6 | −94.0 |
| α-Gurjunene | −80.6 | −61.6 | −84.6 | −100.0 | −83.6 | −78.1 |
| β-Gurjunene | −89.3 | −38.8 | −80.3 | 11.3 | −77.6 | −71.1 |
| α-Himachalene | −89.2 | −67.9 | −83.9 | −96.3 | −87.6 | −73.4 |
| β-Himachalene | −81.9 | −65.4 | −96.1 | −110.8 | −91.4 | −76.8 |
| Himachalol | −91.5 | −64.1 | −81.2 | −24.7 | −81.7 | −66.3 |
| α-Humulene | −94.0 | −80.0 | −89.9 | −113.6 | −89.2 | −82.5 |
| Leptospermone | −89.6 | −64.0 | −92.2 | −102.7 | −88.8 | −80.7 |
| iso-Leptospermone | −92.8 | −63.0 | −90.1 | −99.4 | −86.2 | −78.2 |
| (R)-Limonene | −86.2 | −75.7 | −92.4 | −99.6 | −81.7 | −86.8 |
| (S)-Limonene | −84.9 | −77.6 | −93.4 | −99.8 | −83.3 | −87.3 |
| (R,S,R)-Limonene oxide | −87.1 | −76.0 | −93.2 | −110.7 | −84.9 | −86.3 |
| (S,R,R)-Limonene oxide | −84.6 | −68.8 | −85.0 | −84.3 | −76.2 | −69.8 |
| (S,R,S)-Limonene oxide | −91.2 | −73.8 | −97.4 | −93.6 | −85.1 | −87.9 |
| (R)-Linalool | −95.5 | −89.9 | −116.6 | −112.4 | −90.3 | −89.9 |
| (S)-Linalool | −93.0 | −88.9 | −110.1 | −111.9 | −91.0 | −95.7 |
| (R)-Linalyl acetate | −96.9 | −87.1 | −107.1 | −112.5 | −87.1 | −95.2 |
| (S)-Linalyl acetate | −100.0 | −90.5 | −110.2 | −114.3 | −86.6 | −87.1 |
| p-Mentha-1(7),8-diene | −83.3 | −74.4 | −93.6 | −100.0 | −82.9 | −84.9 |
| (+)-Menthol | −86.1 | −66.4 | −98.6 | −106.1 | −79.2 | −90.8 |
| (-)-Menthol | −81.0 | −68.9 | −97.5 | −106.3 | −80.5 | −89.7 |
| Menthone | −82.4 | −72.0 | −92.6 | −98.2 | −82.8 | −83.5 |
| iso-Menthone | −79.9 | −64.8 | −88.2 | −102.4 | −75.1 | −82.0 |
| α-Muurolene | −91.2 | −79.9 | −95.6 | −113.7 | −90.0 | −82.8 |
| (+)-γ-Muurolene | −85.2 | −77.0 | −98.7 | −113.9 | −93.2 | −78.7 |
| (-)-γ-Muurolene | −83.1 | −83.4 | −96.9 | −106.3 | −82.9 | −80.5 |
| Myrcene | −90.4 | −88.8 | −109.0 | −112.7 | −90.3 | −101.1 |
| Neral | −95.1 | −94.1 | −111.1 | −122.8 | −90.9 | −105.4 |
| (E)-Nerolidol | −111.5 | −101.8 | −131.9 | −131.3 | −99.6 | −110.5 |
| (E)-β-Ocimene | −91.8 | −89.4 | −104.6 | −116.4 | −89.0 | −97.0 |
| (Z)-β-Ocimene | −89.7 | −90.6 | −106.8 | −112.1 | −90.3 | −94.4 |
| Octanal | −85.8 | −79.8 | −97.6 | −112.0 | −85.7 | −102.7 |
| (R)-2-Octen-3-ol | −84.3 | −81.5 | −106.9 | −107.1 | −83.3 | −101.1 |
| (S)-1-Octen-3-ol | −84.5 | −82.4 | −108.4 | −105.9 | −85.0 | −99.5 |
| 3-Octanone | −84.2 | −82.5 | −102.6 | −111.6 | −82.9 | −101.8 |
| Patchouli alcohol | −75.1 | −41.4 | −71.1 | 100.1 | −10.1 | −49.2 |
| (R)-Phellandral | −92.3 | −79.3 | −97.4 | −108.1 | −88.9 | −89.5 |
| (S)-Phellandral | −92.9 | −79.9 | −96.1 | −109.2 | −88.9 | −88.8 |
| (R)-α-Phellandrene | −84.0 | −69.7 | −94.7 | −100.8 | −82.4 | −83.8 |
| (S)-α-Phellandrene | −84.3 | −69.6 | −93.5 | −99.4 | −82.6 | −80.6 |
| (R)-β-Phellandrene | −82.2 | −69.9 | −93.5 | −102.0 | −80.6 | −84.1 |
| (S)-β-Phellandrene | −83.8 | −69.0 | −93.2 | −100.5 | −80.2 | −81.2 |
| Phytol | −112.8 | −93.8 | −132.4 | −134.7 | −102.4 | −112.9 |
| Phytone | −117.8 | −92.1 | −124.5 | −131.1 | −99.7 | −107.2 |
| (+)-α-Pinene | −74.2 | −58.0 | −73.4 | −62.4 | −62.5 | −54.4 |
| (-)-α-Pinene | −72.8 | −58.3 | −70.4 | −64.6 | −61.7 | −57.2 |
| (+)-β-Pinene | −70.9 | −57.8 | −74.0 | −64.1 | −60.9 | −55.5 |
| (-)-β-Pinene | −73.5 | −58.4 | −73.1 | −65.0 | −63.7 | −57.0 |
| (+)-Pinocamphone | −76.5 | −58.2 | −73.6 | −53.1 | −55.4 | −63.6 |
| (-)-Pinocamphone | −76.6 | −66.1 | −76.5 | −65.9 | −72.3 | −62.7 |
| (+)-iso-Pinocamphone | −75.3 | −59.0 | −76.6 | −54.0 | −59.1 | −58.1 |
| (-)-iso-Pinocamphone | −76.2 | −59.6 | −76.7 | −56.4 | −65.8 | −56.2 |
| (+)-cis-Pinocarveol | −75.7 | −58.2 | −75.2 | −51.7 | −57.5 | −58.1 |
| (-)-cis-Pinocarveol | −75.7 | −63.9 | −73.5 | −52.5 | −66.3 | −54.5 |
| (+)-trans-Pinocarveol | −77.5 | −60.6 | −76.5 | −71.1 | −63.9 | −61.4 |
| (-)-trans-Pinocarveol | −72.4 | −58.6 | −77.2 | −71.4 | −56.3 | −62.6 |
| (+)-Piperitone oxide | −88.5 | −70.5 | −100.5 | −104.2 | −80.4 | −78.0 |
| (-)-Piperitone oxide | −86.0 | −73.6 | −99.6 | −104.5 | −80.7 | −83.8 |
| (-)-iso-Pulegol | −85.1 | −72.0 | −93.5 | −106.7 | −83.5 | −93.8 |
| (R)-Pulegone | −81.8 | −69.9 | −90.3 | −103.9 | −78.4 | −74.4 |
| (S)-Pulegone | −79.7 | −69.6 | −89.3 | −99.9 | −81.5 | −76.4 |
| Rotundifolone | −86.1 | −72.1 | −96.1 | −100.0 | −82.4 | −72.1 |
| (+)-Sabinene | −87.9 | −66.6 | −84.5 | −92.1 | −80.5 | −78.2 |
| (-)-Sabinene | −79.7 | −68.4 | −91.4 | −96.0 | −80.8 | −75.8 |
| cis-Sabinene hydrate | −88.1 | −70.2 | −84.5 | −91.1 | −72.3 | −68.1 |
| (Z)-α-Santalol | −108.5 | −85.6 | −113.4 | −106.8 | −96.8 | −90.7 |
| (E)-β-Santalol | −105.0 | −90.2 | −110.6 | −113.3 | −94.3 | −94.0 |
| (Z)-β-Santalol | −108.0 | −85.3 | −110.4 | −110.9 | −96.5 | −92.6 |
| β-Sesquiphellandrene | −103.9 | −92.2 | −116.6 | −127.5 | −99.4 | −98.3 |
| Spathulenol | −95.9 | −94.8 | −97.7 | −102.8 | −84.3 | −102.2 |
| (Z)-Spiroether | −112.8 | −85.5 | −124.3 | −136.1 | −101.5 | −102.9 |
| γ-Terpinene | −84.3 | −74.9 | −93.4 | −98.7 | −82.8 | −85.9 |
| (R)-Terpinen-4-ol | −87.3 | −66.7 | −81.5 | −94.0 | −80.2 | −72.2 |
| (S)-Terpinen-4-ol | −87.1 | −67.2 | −78.9 | −92.3 | −80.2 | −71.7 |
| (R)-α-Terpineol | −82.0 | −68.0 | −80.3 | −82.2 | −77.6 | −71.3 |
| (S)-α-Terpineol | −76.9 | −75.5 | −103.6 | −101.7 | −86.6 | −84.9 |
| Terpinolene | −83.1 | −69.8 | −89.0 | −101.8 | −84.8 | −72.9 |
| (-)-α-Thujone | −81.4 | −66.2 | −84.9 | −93.4 | −72.6 | −74.1 |
| (+)-β-Thujone | −87.6 | −66.4 | −89.9 | −102.9 | −71.7 | −86.9 |
| Thymohydroquinone dimethyl ether | −86.3 | −75.8 | −103.7 | −116.0 | −85.3 | −95.5 |
| Thymol | −84.4 | −70.8 | −96.4 | −107.1 | −79.7 | −91.8 |
| 2-Undecanone | −94.5 | −90.2 | −110.0 | −129.7 | −91.9 | −107.6 |
| Valencene | −96.7 | −75.1 | −103.3 | −114.7 | −87.8 | −76.6 |
| (+)-Verbenone | −74.3 | −64.5 | −79.3 | −71.6 | −63.9 | −72.3 |
| (-)-Verbenone | −73.5 | −63.1 | −78.0 | −65.3 | −64.8 | −59.3 |
| Viridiflorene | −81.9 | −68.6 | −102.1 | −106.9 | −73.1 | −81.9 |
| α-Zingiberene | −108.3 | −90.9 | −113.8 | −123.4 | −97.7 | −99.9 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
da Silva, J.K.R.; Figueiredo, P.L.B.; Byler, K.G.; Setzer, W.N. Essential Oils as Antiviral Agents, Potential of Essential Oils to Treat SARS-CoV-2 Infection: An In-Silico Investigation. Int. J. Mol. Sci. 2020, 21, 3426. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21103426
da Silva JKR, Figueiredo PLB, Byler KG, Setzer WN. Essential Oils as Antiviral Agents, Potential of Essential Oils to Treat SARS-CoV-2 Infection: An In-Silico Investigation. International Journal of Molecular Sciences. 2020; 21(10):3426. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21103426
Chicago/Turabian Styleda Silva, Joyce Kelly R., Pablo Luis Baia Figueiredo, Kendall G. Byler, and William N. Setzer. 2020. "Essential Oils as Antiviral Agents, Potential of Essential Oils to Treat SARS-CoV-2 Infection: An In-Silico Investigation" International Journal of Molecular Sciences 21, no. 10: 3426. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21103426