The GOLIATH Project: Towards an Internationally Harmonised Approach for Testing Metabolism Disrupting Compounds

 , ,
, ,  , , , , , , , ,
on behalf of the GOLIATH Consortium
, , , , , , , ,
on behalf of the GOLIATH Consortium
Abstract
:1. Introduction
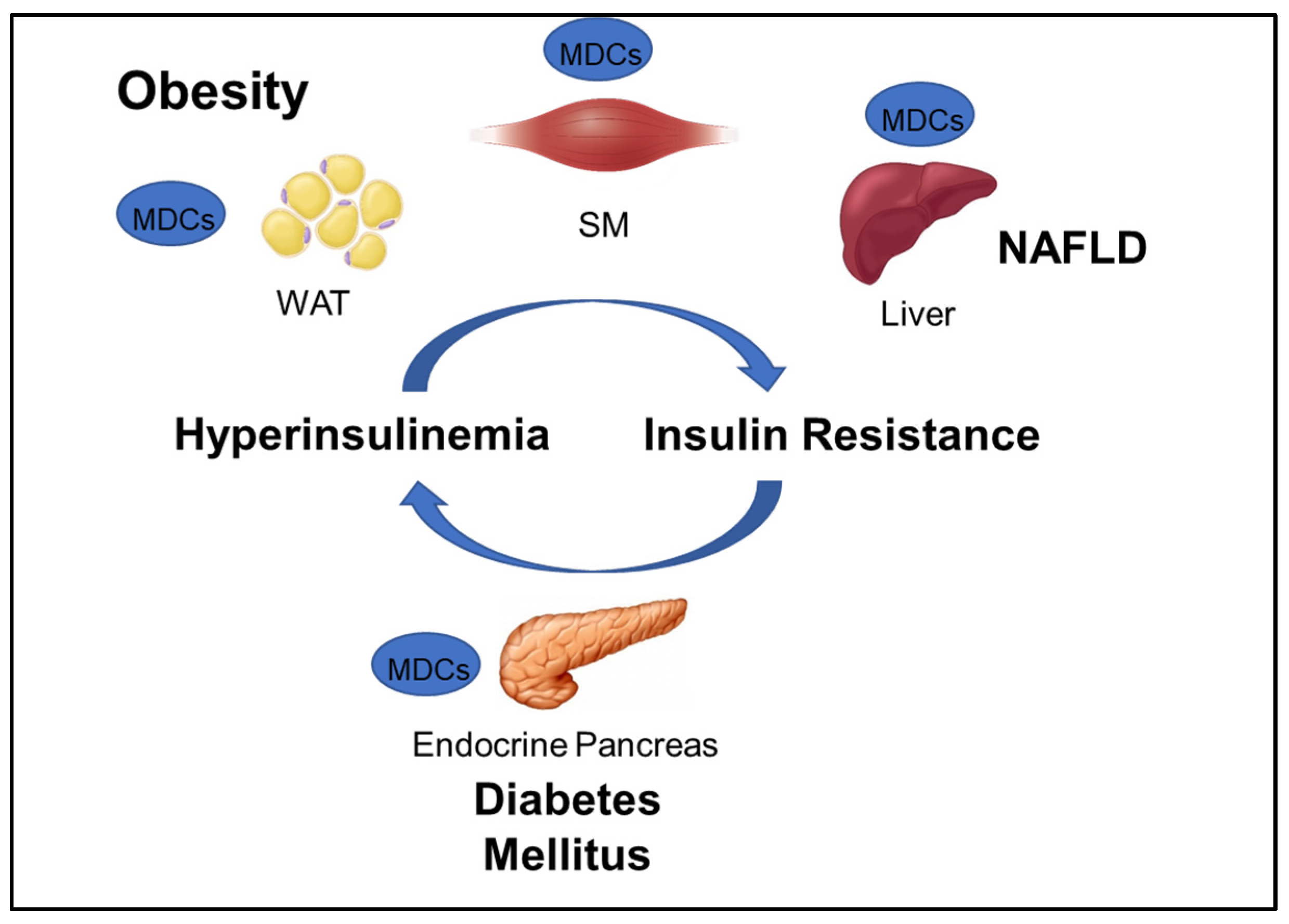
2. Metabolic Disorders
Metabolic Disrupting Chemicals
3. Test Methods in GOLIATH to Determine Metabolism Disrupting Activity
3.1. Test Methods to Determine MIEs
3.1.1. In Silico Predictive Models
3.1.2. NR Screening
3.1.3. Bioactive Thermal Protein Profiling (bTPP)
3.2. Test Methods to Determine Key Events, i.e., In Vitro Models of Metabolic Disruption
3.2.1. Liver
Metabolism in HPR116 Cells
Steatosis
3.2.2. Endocrine Pancreas
3.2.3. Muscle
3.2.4. Adipocytes
Adipocyte Commitment and Differentiation
Insulin Resistance
3.3. Methods and Approaches to Determine Adverse Outcomes
3.3.1. Zebrafish
3.3.2. Rodent Studies
3.3.3. Epidemiological Studies
4. Approaches to Determine Endocrine Mode of Action
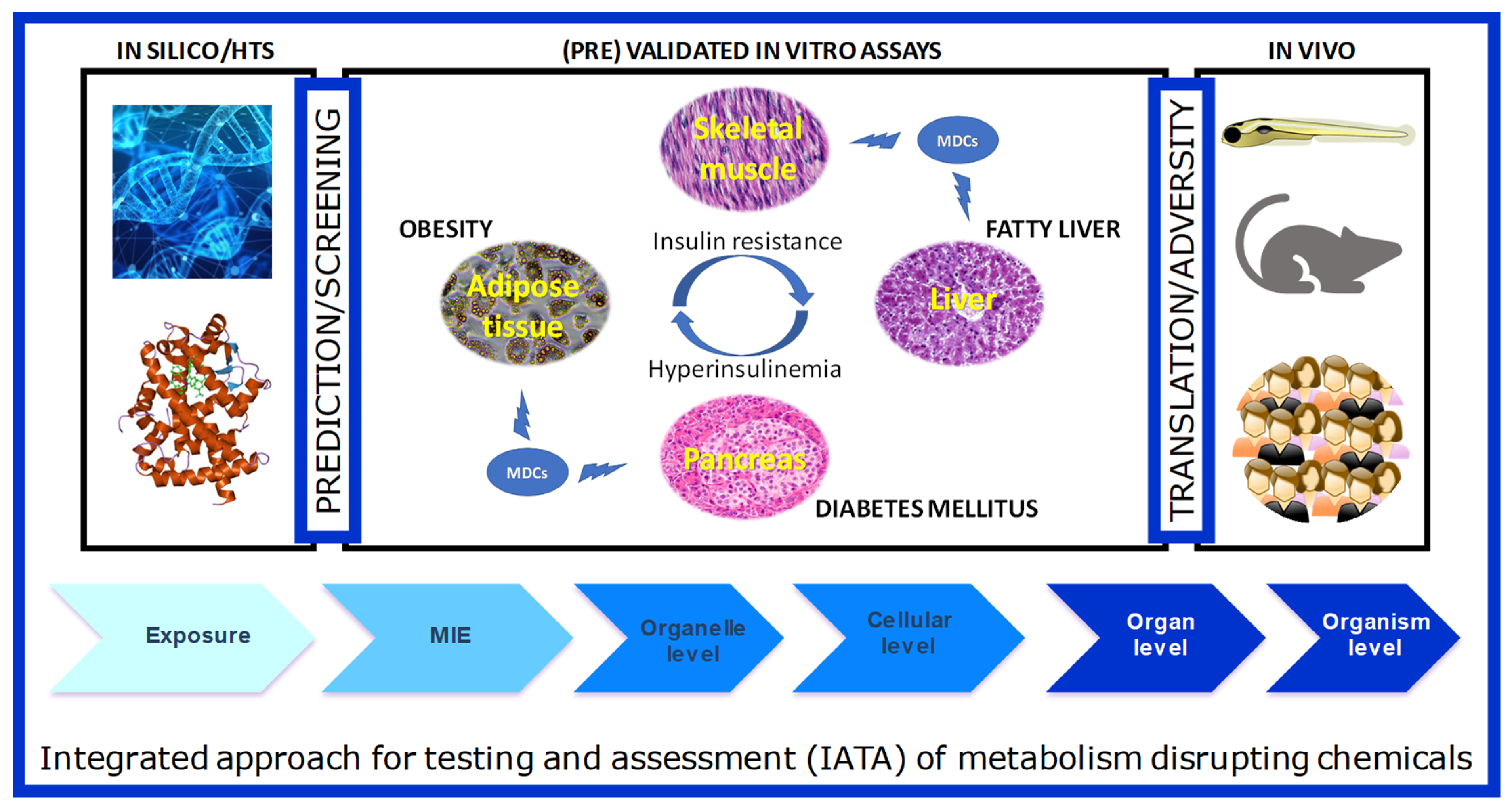
5. From Individual Assays to AOPs and an Integrated Approach to Testing and Assessment of MDCs
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- WHO Diabetes—Data and Statistics. Available online: http://www.euro.who.int/en/health-topics/noncommunicable-diseases/diabetes/data-and-statistics (accessed on 25 February 2020).
- Younossi, Z.M.; Blissett, D.; Blissett, R.; Henry, L.; Stepanova, M.; Younossi, Y.; Racila, A.; Hunt, S.; Beckerman, R. The economic and clinical burden of nonalcoholic fatty liver disease in the United States and Europe. Hepatology 2016, 64, 1577–1586. [Google Scholar] [CrossRef]
- Franks, P.W.; McCarthy, M.I. Exposing the exposures responsible for type 2 diabetes and obesity. Science 2016, 354, 69–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heindel, J.J.; Blumberg, B.; Cave, M.; Machtinger, R.; Mantovani, A.; Mendez, M.A.; Nadal, Á.; Palanza, P.; Panzica, G.; Sargis, R.; et al. Metabolism disrupting chemicals and metabolic disorders. Reprod. Toxicol. 2016, 68, 3–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- European Commission Endocrine Disruptors. Available online: https://ec.europa.eu/environment/chemicals/endocrine/documents/index_en.htm (accessed on 25 February 2020).
- EC. European Commission COUNCIL REGULATION (EC) No 440/2008. Off. J. Eur. Union 2008, OJ L 142, 1–739. [Google Scholar]
- OECD. OECD Work on Endocrine Disrupting Chemicals. Available online: https://www.oecd.org/env/ehs/testing/oecdworkrelatedtoendocrinedisrupters.htm (accessed on 25 February 2020).
- Jacobs, M.; Janssens, W.; Bernauer, U.; Brandon, E.; Coecke, S.; Combes, R.; Edwards, P.; Freidig, A.; Freyberger, A.; Kolanczyk, R.; et al. The Use of Metabolising Systems for In Vitro Testing of Endocrine Disruptors. Curr. Drug Metab. 2008, 9, 796–826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacobs, M. In vitro metabolism and bioavailability tests for endocrine active substances: What is needed next for regulatory purposes? ALTEX 2013, 30, 331–351. [Google Scholar] [CrossRef] [Green Version]
- Bopp, S.; Nepelska, M.; Halder, M.; Munn, S. Expert survey on identification of gaps in available test methods for evaluation of endocrine disruptors. JRC Tech. Rep. 2017. [Google Scholar] [CrossRef]
- OECD. Detailed Review Paper on the State of the Science on Novel In Vitro and In Vivo Screening and Testing Methods and Endpoints for Evaluating Endocrine Disruptors; OECD Series on Testing and Assessment; OECD: Paris, France, 2014; ISBN 9789264221352. [Google Scholar]
- FAIR. Available online: https://www.go-fair.org/ (accessed on 30 March 2020).
- European Association for the Study of the Liver (EASL). EASL–EASD–EASO Clinical Practice Guidelines for the management of non-alcoholic fatty liver disease. Diabetologia 2016, 59, 1121–1140. [Google Scholar] [CrossRef] [Green Version]
- Gore, A.C.; Chappell, V.A.; Fenton, S.E.; A Flaws, J.; Nadal, Á.; Prins, G.S.; Toppari, J.; Zoeller, R.T. EDC-2: The Endocrine Society’s Second Scientific Statement on Endocrine-Disrupting Chemicals. Endocr. Rev. 2015, 36, E1–E150. [Google Scholar] [CrossRef]
- Nadal, Á.; Quesada, I.; Tudurí, E.; Nogueiras, R.; Alonso-Magdalena, P. Endocrine-disrupting chemicals and the regulation of energy balance. Nat. Rev. Endocrinol. 2017, 13, 536–546. [Google Scholar] [CrossRef]
- Cano-Sancho, G.; Salmon, A.G.; La Merrill, M.A. Association between Exposure to p,p ′-DDT and Its Metabolite p,p ′-DDE with Obesity: Integrated Systematic Review and Meta-Analysis. Environ. Health Perspect. 2017, 125, 096002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Batista, T.; Alonso-Magdalena, P.; Vieira, E.; Amaral, M.E.C.D.; Cederroth, C.R.; Nef, S.; Quesada, I.; Carneiro, E.M.; Nadal, Á. Short-Term Treatment with Bisphenol-A Leads to Metabolic Abnormalities in Adult Male Mice. PLoS ONE 2012, 7, e33814. [Google Scholar] [CrossRef] [PubMed]
- Bucher, S.; Jalili, P.; Le Guillou, D.; Begriche, K.; Rondel, K.; Martinais, S.; Zalko, D.; Corlu, A.; Robin, M.-A.; Fromenty, B. Bisphenol a induces steatosis in HepaRG cells using a model of perinatal exposure. Environ. Toxicol. 2016, 32, 1024–1036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wassenaar, P.; Trasande, L.; Legler, J. Systematic Review and Meta-Analysis of Early-Life Exposure to Bisphenol A and Obesity-Related Outcomes in Rodents. Environ. Health Perspect. 2017, 125, 106001. [Google Scholar] [CrossRef] [Green Version]
- OECD The QSAR Toolbox. Available online: https://www.oecd.org/chemicalsafety/risk-assessment/oecd-qsar-toolbox.htm (accessed on 10 October 2019).
- OECD. Guidance Document on the Validation of (Quantitative) Structure-Activity Relationship [(Q)SAR] Models; OECD Series on Testing and Assessment; OECD: Paris, France, 2014; ISBN 9789264085442. [Google Scholar]
- Schneider, M.; Pons, J.-L.; Labesse, G.; Bourguet, W. In Silico Predictions of Endocrine Disruptors Properties. Endocrinology 2019, 160, 2709–2716. [Google Scholar] [CrossRef]
- Punt, A.; Aartse, A.; Bovee, T.F.H.; Gerssen, A.; Van Leeuwen, S.P.J.; Hoogenboom, R.; Peijnenburg, A.A.C.M. Quantitative in vitro-to-in vivo extrapolation (QIVIVE) of estrogenic and anti-androgenic potencies of BPA and BADGE analogues. Arch. Toxicol. 2019, 93, 1941–1953. [Google Scholar] [CrossRef] [Green Version]
- Leonard, J.A.; Tan, Y.-M.; Gilbert, M.; Isaacs, K.; El-Masri, H. Estimating Margin of Exposure to Thyroid Peroxidase Inhibitors Using High-Throughput in vitro Data, High-Throughput Exposure Modeling, and Physiologically Based Pharmacokinetic/Pharmacodynamic Modeling. Toxicol. Sci. 2016, 151, 57–70. [Google Scholar] [CrossRef] [Green Version]
- Kato, H.; Yamaotsu, N.; Iwazaki, N.; Okamura, S.; Kume, T.; Hirono, S. Precise prediction of activators for the human constitutive androstane receptor using structure-based three-dimensional quantitative structure–activity relationship methods. Drug Metab. Pharmacokinet. 2017, 32, 179–188. [Google Scholar] [CrossRef]
- Chen, Y.; Yang, H.; Wu, Z.; Liu, G.; Tang, Y.; Li, W. Prediction of Farnesoid X Receptor Disruptors with Machine Learning Methods. Chem. Res. Toxicol. 2018, 31, 1128–1137. [Google Scholar] [CrossRef]
- Al Sharif, M.; Tsakovska, I.; Pajeva, I.; Alov, P.; Fioravanzo, E.; Bassan, A.; Kovarich, S.; Yang, C.; Mostrag-Szlichtyng, A.; Vitcheva, V.; et al. The application of molecular modelling in the safety assessment of chemicals: A case study on ligand-dependent PPARγ dysregulation. Toxicology 2017, 392, 140–154. [Google Scholar] [CrossRef]
- Yin, C.; Yang, X.; Wei, M.; Liu, H. Predictive models for identifying the binding activity of structurally diverse chemicals to human pregnane X receptor. Environ. Sci. Pollut. Res. 2017, 24, 20063–20071. [Google Scholar] [CrossRef] [PubMed]
- Schneider, M.; Pons, J.-L.; Bourguet, W.; Labesse, G. Towards accurate high-throughput ligand affinity prediction by exploiting structural ensembles, docking metrics and ligand similarity. Bioinformatics 2019, 36, 160–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grimaldi, M.; Boulahtouf, A.; Delfosse, V.; Thouënnon, E.; Bourguet, W.; Balaguer, P. Reporter Cell Lines for the Characterization of the Interactions between Human Nuclear Receptors and Endocrine Disruptors. Front. Endocrinol. 2015, 6, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grimaldi, M.; Boulahtouf, A.; Toporova, L.; Balaguer, P. Functional profiling of bisphenols for nuclear receptors. Toxicology 2019, 420, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Riu, A.; Grimaldi, M.; Le Maire, A.; Bey, G.; Phillips, K.; Boulahtouf, A.; Perdu, E.; Zalko, D.; Bourguet, W.; Balaguer, P. Peroxisome Proliferator-Activated Receptor γ Is a Target for Halogenated Analogs of Bisphenol A. Environ. Health Perspect. 2011, 119, 1227–1232. [Google Scholar] [CrossRef] [PubMed]
- Del Amor, A.M.C.; Freitas, S.; Urbatzka, R.; Fresnedo, O.; Cristobal, S. Application of Bioactive Thermal Proteome Profiling to Decipher the Mechanism of Action of the Lipid Lowering 132-Hydroxy-pheophytin Isolated from a Marine Cyanobacteria. Mar. Drugs 2019, 17, 371. [Google Scholar] [CrossRef] [Green Version]
- Molina, D.M.; Jafari, R.; Chen, D.; Seki, T.; Larsson, A.; Dan, C.; Sreekumar, L.; Cao, Y.; Nordlund, P. Monitoring Drug Target Engagement in Cells and Tissues Using the Cellular Thermal Shift Assay. Science 2013, 341, 84–87. [Google Scholar] [CrossRef]
- Savitski, M.M.; Reinhard, F.B.M.; Franken, H.; Werner, T.; Savitski, M.F.; Eberhard, D.; Molina, D.M.; Jafari, R.; Dovega, R.B.; Klaeger, S.; et al. Tracking cancer drugs in living cells by thermal profiling of the proteome. Science 2014, 346, 1255784. [Google Scholar] [CrossRef] [Green Version]
- Franken, H.; Mathieson, T.; Childs, D.; A Sweetman, G.M.; Werner, T.; Tögel, I.; Doce, C.; Gade, S.; Bantscheff, M.; Drewes, G.; et al. Thermal proteome profiling for unbiased identification of direct and indirect drug targets using multiplexed quantitative mass spectrometry. Nat. Protoc. 2015, 10, 1567–1593. [Google Scholar] [CrossRef]
- Becher, I.; Werner, T.; Doce, C.; Zaal, E.A.; Tögel, I.; Khan, C.; Rueger, A.; Muelbaier, M.; Salzer, E.; Berkers, C.R.; et al. Thermal profiling reveals phenylalanine hydroxylase as an off-target of panobinostat. Nat. Methods 2016, 12, 908–910. [Google Scholar] [CrossRef]
- Azimi, A.; Caramuta, S.; Seashore-Ludlow, B.; Boström, J.; Robinson, J.L.; Edfors, F.; Tuominen, R.; Kemper, K.; Krijgsman, O.; Peeper, D.S.; et al. Targeting CDK 2 overcomes melanoma resistance against BRAF and Hsp90 inhibitors. Mol. Syst. Boil. 2018, 14, e7858. [Google Scholar] [CrossRef] [PubMed]
- Türkowsky, D.; Lohmann, P.; Mühlenbrink, M.; Schubert, T.; Adrian, L.; Goris, T.; Jehmlich, N.; Von Bergen, M. Thermal proteome profiling allows quantitative assessment of interactions between tetrachloroethene reductive dehalogenase and trichloroethene. J. Proteom. 2019, 192, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Gripon, P.; Rumin, S.; Urban, S.; Le Seyec, J.; Glaise, D.; Cannie, I.; Guyomard, C.; Lucas, J.; Trepo, C.; Guguen-Guillouzo, C. Nonlinear partial differential equations and applications: Infection of a human hepatoma cell line by hepatitis B virus. Proc. Natl. Acad. Sci. USA 2002, 99, 15655–15660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guillouzo, A.; Corlu, A.; Aninat, C.; Glaise, D.; Morel, F.; Guguen-Guillouzo, C. The human hepatoma HepaRG cells: A highly differentiated model for studies of liver metabolism and toxicity of xenobiotics. Chem. Interact. 2007, 168, 66–73. [Google Scholar] [CrossRef]
- Andersson, T.B.; Kanebratt, K.P.; Kenna, G. The HepaRG cell line: A unique in vitro tool for understanding drug metabolism and toxicology in human. Expert Opin. Drug Metab. Toxicol. 2012, 8, 909–920. [Google Scholar] [CrossRef]
- Aninat, C.; Piton, A.; Glaise, D.; Le Charpentier, T.; Langouët, S.; Morel, F.; Guguen-Guillouzo, C.; Guillouzo, A. Expression of cytochromes P450, conjugating enzymes and nuclear receptors in human hepatoma HepaRG cells. Drug Metab. Dispos. 2005, 34, 75–83. [Google Scholar] [CrossRef] [Green Version]
- Rogue, A.; Lambert, C.; Josse, R.; Antherieu, S.; Spire, C.; Claude, N.; Guillouzo, A. Comparative Gene Expression Profiles Induced by PPARγ and PPARα/γ Agonists in Human Hepatocytes. PLoS ONE 2011, 6, e18816. [Google Scholar] [CrossRef] [Green Version]
- Peyta, L.; Jarnouen, K.; Pinault, M.; Guimaraes, C.; De Barros, J.-P.P.; Chevalier, S.; Dumas, J.-F.; Maillot, F.; Hatch, G.M.; Loyer, P.; et al. Reduced cardiolipin content decreases respiratory chain capacities and increases ATP synthesis yield in the human HepaRG cells. Biochim. Biophys. Acta (BBA) Gen. Subj. 2016, 1857, 443–453. [Google Scholar] [CrossRef]
- Porceddu, P.; Buron, N.; Rustin, P.; Fromenty, B.; Borgne-Sanchez, A. In Vitro Assessment of Mitochondrial Toxicity to Predict Drug-Induced Liver Injury. In Methods in Molecular Biology; Springer Science and Business Media LLC: New York, NY, USA, 2018; pp. 283–300. [Google Scholar]
- Tascher, G.; Burban, A.; Camus, S.; Plumel, M.; Chanon, S.; Le Guevel, R.; Shevchenko, V.; Van Dorsselaer, A.; Lefai, E.; Guguen-Guillouzo, C.; et al. In-Depth Proteome Analysis Highlights HepaRG Cells as a Versatile Cell System Surrogate for Primary Human Hepatocytes. Cells 2019, 8, 192. [Google Scholar] [CrossRef] [Green Version]
- Waxman, D.J.; Lapenson, D.P.; Aoyama, T.; Gelboin, H.V.; Gonzalez, F.J.; Korzekwa, K. Steroid hormone hydroxylase specificities of eleven cDNA-expressed human cytochrome P450s. Arch. Biochem. Biophys. 1991, 290, 160–166. [Google Scholar] [CrossRef]
- EURL ECVAM. Multi-Study Validation Trial for Cytochrome P450 Induction Providing a Reliable Human Metabolically Competent Standard Model or Method Using the Human Cryopreserved Primary Hepatocytes and the Human Cryopreserved HepaRG® Cell Line; EURL ECVAM: Ispra, Italy, 2014. [Google Scholar]
- Bernasconi, C.; Pelkonen, O.; Andersson, T.B.; Strickland, J.; Wilk-Zasadna, I.; Asturiol, D.; Cole, T.; Liska, R.; Worth, A.; Müller-Vieira, U.; et al. Validation of in vitro methods for human cytochrome P450 enzyme induction: Outcome of a multi-laboratory study. Toxicol. Vitr. 2019, 60, 212–228. [Google Scholar] [CrossRef]
- OECD Determination of Cytochrome 450 (CYP) Enzyme Activity Induction Using Differentiated Human Hepatic Cells. Available online: http://www.oecd.org/chemicalsafety/testing/Draft_TG_CYP_induction_for_2nd_WNT_review.pdf (accessed on 25 February 2020).
- Le Guillou, D.; Bucher, S.; Begriche, K.; Hoët, D.; Lombes, A.; Labbe, G.; Fromenty, B. Drug-Induced Alterations of Mitochondrial DNA Homeostasis in Steatotic and Nonsteatotic HepaRG Cells. J. Pharmacol. Exp. Ther. 2018, 365, 711–726. [Google Scholar] [CrossRef] [PubMed]
- Mueller, S.O.; Guillouzo, A.; Hewitt, P.G.; Richert, L. Drug biokinetic and toxicity assessments in rat and human primary hepatocytes and HepaRG cells within the EU-funded Predict-IV project. Toxicol. Vitr. 2015, 30, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Bucher, S.; Le Guillou, D.; Allard, J.; Pinon, G.; Begriche, K.; Tête, A.; Sergent, O.; Lagadic-Gossmann, M.; Fromenty, B. Possible Involvement of Mitochondrial Dysfunction and Oxidative Stress in a Cellular Model of NAFLD Progression Induced by Benzo[a]pyrene/Ethanol CoExposure. Oxidative Med. Cell. Longev. 2018, 2018, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antherieu, S.; Rogue, A.; Fromenty, B.; Guillouzo, A.; Robin, M.-A. Induction of vesicular steatosis by amiodarone and tetracycline is associated with up-regulation of lipogenic genes in heparg cells. Hepatology 2011, 53, 1895–1905. [Google Scholar] [CrossRef]
- Tolosa, L.; Gómez-Lechón, M.J.; Jimenez, N.; Hervás, D.; Jover, R.; Donato, M.T. Advantageous use of HepaRG cells for the screening and mechanistic study of drug-induced steatosis. Toxicol. Appl. Pharmacol. 2016, 302, 1–9. [Google Scholar] [CrossRef]
- Grünig, D.; Felser, A.; Duthaler, U.; Bouitbir, J.; Krähenbühl, S. Effect of the Catechol-O-Methyltransferase Inhibitors Tolcapone and Entacapone on Fatty Acid Metabolism in HepaRG Cells. Toxicol. Sci. 2018, 164, 477–488. [Google Scholar] [CrossRef]
- Allard, J.; Bucher, S.; Ferron, P.-J.; Begriche, K.; Loyer, P.; Fromenty, B. Importance of non-mitochondrial pathways in drug-induced hepatic steatosis: Investigations with 12 steatotic drugs in HepaRG cells. Toxicol. Lett. 2019, 314, S60. [Google Scholar]
- Stossi, F.; Dandekar, R.D.; Johnson, H.; LaVere, P.; Foulds, C.E.; Mancini, M.G.; Mancini, M.A. Tributyltin chloride (TBT) induces RXRA down-regulation and lipid accumulation in human liver cells. PLoS ONE 2019, 14, e0224405. [Google Scholar] [CrossRef]
- Alonso-Magdalena, P.; Quesada, I.; Nadal, Á. Endocrine disruptors in the etiology of type 2 diabetes mellitus. Nat. Rev. Endocrinol. 2011, 7, 346–353. [Google Scholar] [CrossRef]
- Mimoto, M.S.; Nadal, Á.; Sargis, R.M. Polluted Pathways: Mechanisms of Metabolic Disruption by Endocrine Disrupting Chemicals. Curr. Environ. Health Rep. 2017, 4, 208–222. [Google Scholar] [CrossRef] [PubMed]
- Alonso-Magdalena, P.; Laribi, O.; Ropero, A.B.; Fuentes, E.; Ripoll, C.; Soria, B.; Nadal, Á. Low Doses of Bisphenol A and Diethylstilbestrol Impair Ca2+ Signals in Pancreatic α-Cells through a Nonclassical Membrane Estrogen Receptor within Intact Islets of Langerhans. Environ. Health Perspect. 2005, 113, 969–977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Esterik, J.; Dollé, M.; Lamoree, M.; Van Leeuwen, S.P.J.; Hamers, T.; Legler, J.; Van Der Ven, L.T. Programming of metabolic effects in C57BL/6JxFVB mice by exposure to bisphenol A during gestation and lactation. Toxicology 2014, 321, 40–52. [Google Scholar] [CrossRef] [PubMed]
- Davis, A.F.; Thomas, A.A.; Shorter, K.S.; Brown, S.L.; Baumgarner, B.L. Cellular fatty acid level regulates the effect of tolylfluanid on mitochondrial dysfunction and insulin sensitivity in C2C12 skeletal myotubes. Biochem. Biophys. Res. Commun. 2018, 505, 392–398. [Google Scholar] [CrossRef] [PubMed]
- Drobna, Z.; Talarovicova, A.; Schrader, H.E.; Fennell, T.R.; Snyder, R.W.; Rissman, E. Bisphenol F has different effects on preadipocytes differentiation and weight gain in adult mice as compared with Bisphenol A and S. Toxicology 2019, 420, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Pomatto, V.; Cottone, E.; Cocci, P.; Mozzicafreddo, M.; Mosconi, G.; Nelson, E.R.; Palermo, F.A.; Bovolin, P. Plasticizers used in food-contact materials affect adipogenesis in 3T3-L1 cells. J. Steroid Biochem. Mol. Boil. 2018, 178, 322–332. [Google Scholar] [CrossRef]
- Sargis, R.M.; Johnson, D.N.; Choudhury, R.A.; Brady, M.J. Environmental endocrine disruptors promote adipogenesis in the 3T3-L1 cell line through glucocorticoid receptor activation. Obesity 2009, 18, 1283–1288. [Google Scholar] [CrossRef] [Green Version]
- Wada, K.; Sakamoto, H.; Nishikawa, K.; Sakuma, S.; Nakajima, A.; Fujimoto, Y.; Kamisaki, Y. Life style-related diseases of the digestive system: Endocrine disruptors stimulate lipid accumulation in target cells related to metabolic syndrome. J. Pharmacol. Sci. 2007, 105, 133–137. [Google Scholar] [CrossRef] [Green Version]
- Taxvig, C.; Dreisig, K.; Boberg, J.; Nellemann, C.; Schelde, A.B.; Pedersen, D.; Boergesen, M.; Mandrup, S.; Vinggaard, A.M. Differential effects of environmental chemicals and food contaminants on adipogenesis, biomarker release and PPARγ activation. Mol. Cell. Endocrinol. 2012, 361, 106–115. [Google Scholar] [CrossRef]
- Kassotis, C.; Masse, L.; Kim, S.; Schlezinger, J.; Webster, T.F.; Stapleton, H.M. Characterization of Adipogenic Chemicals in Three Different Cell Culture Systems: Implications for Reproducibility Based on Cell Source and Handling. Sci. Rep. 2017, 7, 42104. [Google Scholar] [CrossRef] [Green Version]
- Sales, L.B.; Kamstra, J.H.; Cenijn, P.; Van Rijt, L.; Hamers, T.; Legler, J. Effects of endocrine disrupting chemicals on in vitro global DNA methylation and adipocyte differentiation. Toxicol. Vitr. 2013, 27, 1634–1643. [Google Scholar] [CrossRef] [PubMed]
- Lima, L.C.F.; Merlo, E.; Zicker, M.C.; Navia-Pelaez, J.M.; De Oliveira, M.; Capettini, L.D.S.A.; Nogueira, C.R.; Ferreira, A.V.M.; Santos, S.H.S.; Graceli, J.B. Tributyltin impacts in metabolic syndrome development through disruption of angiotensin II receptor signaling pathways in white adipose tissue from adult female rats. Toxicol. Lett. 2018, 299, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Bertuloso, B.D.; Podratz, P.L.; Merlo, E.; De Araújo, J.F.; Lima, L.C.F.; De Miguel, E.C.; De Souza, L.N.; Gava, A.L.; De Oliveira, M.; Miranda-Alves, L.; et al. Tributyltin chloride leads to adiposity and impairs metabolic functions in the rat liver and pancreas. Toxicol. Lett. 2015, 235, 45–59. [Google Scholar] [CrossRef] [PubMed]
- Grün, F.; Watanabe, H.; Zamanian, Z.; Maeda, L.; Arima, K.; Cubacha, R.; Gardiner, D.M.; Kanno, J.; Iguchi, T.; Blumberg, B. Endocrine-disrupting organotin compounds are potent inducers of adipogenesis in vertebrates. Mol. Endocrinol. 2006, 20, 2141–2155. [Google Scholar] [CrossRef]
- Pereira-Fernandes, A.; Demaegdt, H.; Vandermeiren, K.; Hectors, T.L.M.; Jorens, P.G.; Blust, R.; Vanparys, C. Evaluation of a Screening System for Obesogenic Compounds: Screening of Endocrine Disrupting Compounds and Evaluation of the PPAR Dependency of the Effect. PLoS ONE 2013, 8, e77481. [Google Scholar] [CrossRef] [Green Version]
- Watkins, A.M.; Wood, C.R.; Lin, M.T.; Abbott, B. The effects of perfluorinated chemicals on adipocyte differentiation in vitro. Mol. Cell. Endocrinol. 2015, 400, 90–101. [Google Scholar] [CrossRef]
- Mangum, L.H.; Howell, G.E.; Chambers, J.E. Exposure to p,p’-DDE enhances differentiation of 3T3-L1 preadipocytes in a model of sub-optimal differentiation. Toxicol. Lett. 2015, 238, 65–71. [Google Scholar] [CrossRef]
- Kim, J.; Sun, Q.; Yue, Y.; Yoon, K.; Whang, K.-Y.; Clark, J.; Park, Y. 4,4′-Dichlorodiphenyltrichloroethane (DDT) and 4,4′-dichlorodiphenyldichloroethylene (DDE) promote adipogenesis in 3T3-L1 adipocyte cell culture. Pestic. Biochem. Physiol. 2016, 131, 40–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cano-Sancho, G.; Smith, A.; La Merrill, M.A. Triphenyl phosphate enhances adipogenic differentiation, glucose uptake and lipolysis via endocrine and noradrenergic mechanisms. Toxicol. Vitr. 2017, 40, 280–288. [Google Scholar] [CrossRef]
- Shoucri, B.M.; Martinez, E.S.; Abreo, T.J.; Hung, V.; Moosova, Z.; Shioda, T.; Blumberg, B. Retinoid X Receptor Activation Alters the Chromatin Landscape To Commit Mesenchymal Stem Cells to the Adipose Lineage. Endocrinology 2017, 158, 3109–3125. [Google Scholar] [CrossRef] [Green Version]
- Shoucri, B.M.; Hung, V.; Chamorro-García, R.; Shioda, T.; Blumberg, B. Retinoid X Receptor Activation During Adipogenesis of Female Mesenchymal Stem Cells Programs a Dysfunctional Adipocyte. Endocrinology 2018, 159, 2863–2883. [Google Scholar] [CrossRef] [PubMed]
- Kirchner, S.; Kieu, T.; Chow, C.; Casey, S.; Blumberg, B. Prenatal exposure to the environmental obesogen tributyltin predisposes multipotent stem cells to become adipocytes. Mol. Endocrinol. 2010, 24, 526–539. [Google Scholar] [CrossRef]
- Liu, S.; Yang, R.; Yin, N.; Wang, Y.-L.; Faiola, F. Environmental and human relevant PFOS and PFOA doses alter human mesenchymal stem cell self-renewal, adipogenesis and osteogenesis. Ecotoxicol. Environ. Saf. 2019, 169, 564–572. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.-W.; Wu, Q.; Green, B.; Nolen, G.; Shi, L.; Losurdo, J.; Deng, H.; Bauer, S.; Fang, J.-L.; Ning, B. Cytotoxicity and inhibitory effects of low-concentration triclosan on adipogenic differentiation of human mesenchymal stem cells. Toxicol. Appl. Pharmacol. 2012, 262, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Chamorro-García, R.; Kirchner, S.; Li, X.; Janesick, A.; Casey, S.C.; Chow, C.; Blumberg, B. Bisphenol A Diglycidyl Ether Induces Adipogenic Differentiation of Multipotent Stromal Stem Cells through a Peroxisome Proliferator–Activated Receptor Gamma-Independent Mechanism. Environ. Health Perspect. 2012, 120, 984–989. [Google Scholar] [CrossRef]
- Lo, K.A.; Labadorf, A.; Kennedy, N.J.; Han, M.S.; Yap, Y.S.; Matthews, B.; Xin, X.; Sun, L.; Davis, R.J.; Lodish, H.F.; et al. Analysis of in vitro insulin-resistance models and their physiological relevance to in vivo diet-induced adipose insulin resistance. Cell Rep. 2013, 5, 259–270. [Google Scholar] [CrossRef] [Green Version]
- Valentino, R.; D’Esposito, V.; Passaretti, F.; Liotti, A.; Cabaro, S.; Longo, M.; Perruolo, G.; Oriente, F.; Beguinot, F.; Formisano, P. Bisphenol-A Impairs Insulin Action and Up-Regulates Inflammatory Pathways in Human Subcutaneous Adipocytes and 3T3-L1 Cells. PLoS ONE 2013, 8, e82099. [Google Scholar] [CrossRef] [Green Version]
- Ariemma, F.; D’Esposito, V.; Liguoro, D.; Oriente, F.; Cabaro, S.; Liotti, A.; Cimmino, I.; Longo, M.; Beguinot, F.; Formisano, P.; et al. Low-Dose Bisphenol-A Impairs Adipogenesis and Generates Dysfunctional 3T3-L1 Adipocytes. PLoS ONE 2016, 11, e0150762. [Google Scholar] [CrossRef] [Green Version]
- Sakurai, K.; Kawazuma, M.; Adachi, T.; Harigaya, T.; Saito, Y.; Hashimoto, N.; Mori, C. Bisphenol A affects glucose transport in mouse 3T3-F442A adipocytes. Br. J. Pharmacol. 2004, 141, 209–214. [Google Scholar] [CrossRef] [Green Version]
- Regnier, S.M.; El-Hashani, E.; Kamau, W.S.; Zhang, X.; Massad, N.L.; Sargis, R.M. Tributyltin differentially promotes development of a phenotypically distinct adipocyte. Obesity 2015, 23, 1864–1871. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, J.; Yamane, T.; Oishi, Y.; Kobayashi-Hattori, K. Perfluorooctanoic acid binds to peroxisome proliferator-activated receptor? And promotes adipocyte differentiation in 3T3-L1 adipocytes. Biosci. Biotechnol. Biochem. 2014, 79, 636–639. [Google Scholar] [CrossRef] [PubMed]
- Howell, G.E.; Meek, E.; Kilic, J.; Mohns, M.; Mulligan, C.; Chambers, J.E. Exposure to p,p’-dichlorodiphenyldichloroethylene (DDE) induces fasting hyperglycemia without insulin resistance in male C57BL/6H mice. Toxicology 2014, 320, 6–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howell, G.; Mangum, L. Corrigendum to “Exposure to bioaccumulative organochlorine compounds alters adipogenesis, fatty acid uptake, and adipokine production in NIH3T3-L1 cells” [Toxicol. In Vitro 25(1) 394–402]. Toxicol. Vitr. 2011, 25, 1507. [Google Scholar] [CrossRef]
- Broeder, M.D.; Moester, M.J.B.; Kamstra, J.H.; Cenijn, P.; Davidoiu, V.; Kamminga, L.M.; Ariese, F.; De Boer, J.F.; Legler, J. Altered Adipogenesis in Zebrafish Larvae Following High Fat Diet and Chemical Exposure Is Visualised by Stimulated Raman Scattering Microscopy. Int. J. Mol. Sci. 2017, 18, 894. [Google Scholar] [CrossRef]
- Schoeters, G.; Govarts, E.; Bruckers, L.; Den Hond, E.; Nelen, V.; De Henauw, S.; Sioen, I.; Nawrot, T.S.; Plusquin, M.; Vriens, A.; et al. Three cycles of human biomonitoring in Flanders − Time trends observed in the Flemish Environment and Health Study. Int. J. Hyg. Environ. Health 2017, 220, 36–45. [Google Scholar] [CrossRef]
- De Craemer, S.; Croes, K.; Van Larebeke, N.; De Henauw, S.; Schoeters, G.; Govarts, E.; Loots, I.; Nawrot, T.; Nelen, V.; Hond, E.D.; et al. Metals, hormones and sexual maturation in Flemish adolescents in three cross-sectional studies (2002–2015). Environ. Int. 2017, 102, 190–199. [Google Scholar] [CrossRef]
- Baeyens, W.; Vrijens, J.; Gao, Y.; Croes, K.; Schoeters, G.; Den Hond, E.; Sioen, I.; Bruckers, L.; Nawrot, T.; Nelen, V.; et al. Trace metals in blood and urine of newborn/mother pairs, adolescents and adults of the Flemish population (2007–2011). Int. J. Hyg. Environ. Health 2014, 217, 878–890. [Google Scholar] [CrossRef]
- Croes, K.; Hond, E.D.; Bruckers, L.; Loots, I.; Morrens, B.; Nelen, V.; Colles, A.; Schoeters, G.; Sioen, I.; Covaci, A.; et al. Monitoring chlorinated persistent organic pollutants in adolescents in Flanders (Belgium): Concentrations, trends and dose–effect relationships (FLEHS II). Environ. Int. 2014, 71, 20–28. [Google Scholar] [CrossRef]
- Hond, E.D.; Paulussen, M.; Geens, T.; Bruckers, L.; Baeyens, W.; David, F.; Dumont, E.; Loots, I.; Morrens, B.; De Bellevaux, B.N.; et al. Biomarkers of human exposure to personal care products: Results from the Flemish Environment and Health Study (FLEHS 2007–2011). Sci. Total Environ. 2013, 463, 102–110. [Google Scholar] [CrossRef]
- Vrijens, J.; Leermakers, M.; Stalpaert, M.; Schoeters, G.; Hond, E.D.; Bruckers, L.; Colles, A.; Nelen, V.; Mieroop, E.V.D.; Van Larebeke, N.; et al. Trace metal concentrations measured in blood and urine of adolescents in Flanders, Belgium: Reference population and case studies Genk-Zuid and Menen. Int. J. Hyg. Environ. Health 2014, 217, 515–527. [Google Scholar] [CrossRef] [PubMed]
- Koppen, G.; Hond, E.D.; Nelen, V.; Van De Mieroop, E.; Bruckers, L.; Bilau, M.; Keune, H.; Van Larebeke, N.; Covaci, A.; Van De Weghe, H.; et al. Organochlorine and heavy metals in newborns: Results from the Flemish Environment and Health Survey (FLEHS 2002–2006). Environ. Int. 2009, 35, 1015–1022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koppen, G.; Covaci, A.; Van Cleuvenbergen, R.; Schepens, P.; Winneke, G.; Nelen, V.; van Larebeke, N.; Vlietinck, R.; Schoeters, G.; Cleuvenbergen, R. Van Persistent organochlorine pollutants in human serum of 50–65 years old women in the Flanders Environmental and Health Study ( FLEHS ). Part 1: Concentrations and regional differences. Chemosphere 2002, 48, 811–825. [Google Scholar] [CrossRef]
- Brunekreef, B.; Smit, J.; De Jongste, J.; Neijens, H.; Gerritsen, J.; Postma, D.; Aalberse, R.; Koopman, L.; Kerkhof, M.; Wijga, A.; et al. The Prevention and Incidence of Asthma and Mite Allergy (PIAMA) birth cohort study: Design and first results. Pediatr. Allergy Immunol. 2002, 13, 55–60. [Google Scholar] [CrossRef]
- Wijga, A.; Kerkhof, M.; Gehring, U.; De Jongste, J.C.; Postma, D.S.; Aalberse, R.C.; Wolse, A.P.; Koppelman, G.H.; Van Rossem, L.; Oldenwening, M.; et al. Cohort profile: The Prevention and Incidence of Asthma and Mite Allergy (PIAMA) birth cohort. Int. J. Epidemiol. 2013, 43, 527–535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganzleben, C.; Antignac, J.-P.; Barouki, R.; Castano, A.; Fiddicke, U.; Klánová, J.; Lebret, E.; Olea, N.; Sarigiannis, D.; Schoeters, G.R.; et al. Human biomonitoring as a tool to support chemicals regulation in the European Union. Int. J. Hyg. Environ. Health 2017, 220, 94–97. [Google Scholar] [CrossRef] [PubMed]
- Vermeulen, R.; Schymanski, E.L.; Barabási, A.-L.; Miller, G. The exposome and health: Where chemistry meets biology. Science 2020, 367, 392–396. [Google Scholar] [CrossRef] [PubMed]
- Brunk, E.; Sahoo, S.; Zielinski, D.C.; Altunkaya, A.; Dräger, A.; Mih, N.; Gatto, F.; Nilsson, A.; Gonzalez, G.A.P.; Aurich, M.K.; et al. Recon3D enables a three-dimensional view of gene variation in human metabolism. Nat. Biotechnol. 2018, 36, 272–281. [Google Scholar] [CrossRef]
- Poupin, N.; Corlu, A.; Cabaton, N.J.; Dubois-Pot-Schneider, H.; Canlet, C.; Person, E.; Bruel, S.; Frainay, C.; Vinson, F.; Maurier, F.; et al. Large-Scale Modeling Approach Reveals Functional Metabolic Shifts during Hepatic Differentiation. J. Proteome Res. 2018, 18, 204–216. [Google Scholar] [CrossRef]
- Frainay, C.; Jourdan, F. Computational methods to identify metabolic sub-networks based on metabolomic profiles. Briefings Bioinform. 2016, 18, 43–56. [Google Scholar] [CrossRef]
- Haug, K.; Salek, R.M.; Conesa, P.; Hastings, J.; De Matos, P.; Rijnbeek, M.; Mahendraker, T.; Williams, M.; Neumann, S.; Rocca-Serra, P.; et al. MetaboLights—An open-access general-purpose repository for metabolomics studies and associated meta-data. Nucleic Acids Res. 2012, 41, D781–D786. [Google Scholar] [CrossRef]
- Athar, A.; Füllgrabe, A.; George, N.; Iqbal, H.; Huerta, L.; Ali, A.; Snow, C.; A Fonseca, N.; Petryszak, R.; Papatheodorou, I.; et al. ArrayExpress update - from bulk to single-cell expression data. Nucleic Acids Res. 2019, 47, D711–D715. [Google Scholar] [CrossRef] [PubMed]
- Cottret, L.; Frainay, C.; Chazalviel, M.; Cabanettes, F.; Gloaguen, Y.; Camenen, E.; Merlet, B.; Heux, S.; Portais, J.-C.; Poupin, N.; et al. MetExplore: Collaborative edition and exploration of metabolic networks. Nucleic Acids Res. 2018, 46, W495–W502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knapen, D.; Angrish, M.M.; Fortin, M.C.; Katsiadaki, I.; Leonard, M.; Margiotta-Casaluci, L.; Munn, S.; O’Brien, J.M.; Pollesch, N.; Smith, L.; et al. Adverse outcome pathway networks I: Development and applications. Environ. Toxicol. Chem. 2018, 37, 1723–1733. [Google Scholar] [CrossRef] [PubMed]
- OECD. Report on Considerations from Case Studies on Integrated Approaches for Testing and Assessment (IATA) - First Review Cycle (2015); OECD Series on Testing and Assessment; OECD: Paris, France, 2017; ISBN 9789264274815. [Google Scholar]
- Jacobs, M. International regulatory needs for development ofan IATA for non-genotoxic carcinogenic chemical substances. ALTEX 2016, 33, 359–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- OECD Integrated Approaches to Testing and Assessment (IATA). Available online: http://www.oecd.org/chemicalsafety/risk-assessment/iata-integrated-approaches-to-testing-and-assessment.htm (accessed on 25 February 2020).
- European Chemical Agency (ECHA) and European Food Safety Authority (EFSA) with the technical support of the Joint Research Centre (JRC); Andersson, N.; Arena, M.; Auteri, D.; Barmaz, S.; Grignard, E.; Kienzler, A.; Lepper, P.; Lostia, A.M.; Munn, S.; et al. Guidance for the identification of endocrine disruptors in the context of Regulations (EU) No 528/2012 and (EC) No 1107/2009. EFSA J. 2018, 16. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Chemical | Identifiers (CAS; ChEBI; Inchikey) | Structure | MoA | Animal | Human | Biomon. Data |
|---|---|---|---|---|---|---|
| Bisphenol A (BPA) | 80-05-7; 33216; IISBACLAFKSPIT-UHFFFAOYSA-N |  | hERα hERβ hPXR hCAR | ++ | + | + |
| Tributyltin Chloride (TBT) | 1461-22-9; 79734; GCTFWCDSFPMHHS-UHFFFAOYSA-M |  | hRXR hPPARγ | +++ | - | - |
| Perfluorooctanoic acid (PFOA) | 3825-26-1; 35549; SNGREZUHAYWORS-UHFFFAOYSA-N |  | hPXR hPPARγ | ++ | + | ++ |
| Triphenylphosphate (TPP) | 115-86-6; 35033; XZZNDPSIHUTMOC-UHFFFAOYSA-N |  | hERα hERβ hPXR hCAR hPPARγ | ++ | ± | + |
| Dichlorodiphenyldichloroethylene (pp’-DDE) | 72-55-9; 16598; UCNVFOCBFJOQAL-UHFFFAOYSA-N |  | hERα hERβ hPXR hCAR | ++ | ++ | +++ |
| Triclosan (TCS) | 3380-34-5; 164200; XEFQLINVKFYRCS-UHFFFAOYSA-N |  | hPXR hPPARγ | + | ± | ++ |
| Chemical | Cell Type | Effects | Concentrations at which Effects Observed | References |
|---|---|---|---|---|
| BPA | 3T3-L1 adipocytes |
| 1 & 100 nM | [87,88] |
| BPA | 3T3-F442A adipocytes |
| 100 μM 1 & 100 μM | [89] |
| TBT | Differentiating 3T3-L1 adipocytes |
| 50 nM | [90] |
| TBT | Differentiating mMSC adipocytes |
| 100 nM | [81] |
| PFOA | 3T3-L1 adipocytes |
| 200 μM | [91] |
| ppDDE | 3T3-L1 adipocytes |
| 2 & 20 μM | [92,93] |
| TPP | 3T3-L1 adipocytes |
| 0.1–50 μM 50 μM | [79] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Legler, J.; Zalko, D.; Jourdan, F.; Jacobs, M.; Fromenty, B.; Balaguer, P.; Bourguet, W.; Munic Kos, V.; Nadal, A.; Beausoleil, C.; et al. The GOLIATH Project: Towards an Internationally Harmonised Approach for Testing Metabolism Disrupting Compounds. Int. J. Mol. Sci. 2020, 21, 3480. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21103480
Legler J, Zalko D, Jourdan F, Jacobs M, Fromenty B, Balaguer P, Bourguet W, Munic Kos V, Nadal A, Beausoleil C, et al. The GOLIATH Project: Towards an Internationally Harmonised Approach for Testing Metabolism Disrupting Compounds. International Journal of Molecular Sciences. 2020; 21(10):3480. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21103480
Chicago/Turabian StyleLegler, Juliette, Daniel Zalko, Fabien Jourdan, Miriam Jacobs, Bernard Fromenty, Patrick Balaguer, William Bourguet, Vesna Munic Kos, Angel Nadal, Claire Beausoleil, and et al. 2020. "The GOLIATH Project: Towards an Internationally Harmonised Approach for Testing Metabolism Disrupting Compounds" International Journal of Molecular Sciences 21, no. 10: 3480. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21103480