Toxic Effects of Nonylphenol on Neonatal Testicular Development in Mouse Organ Culture

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Development of the Mouse Fetal Testes in In Vitro Culture System
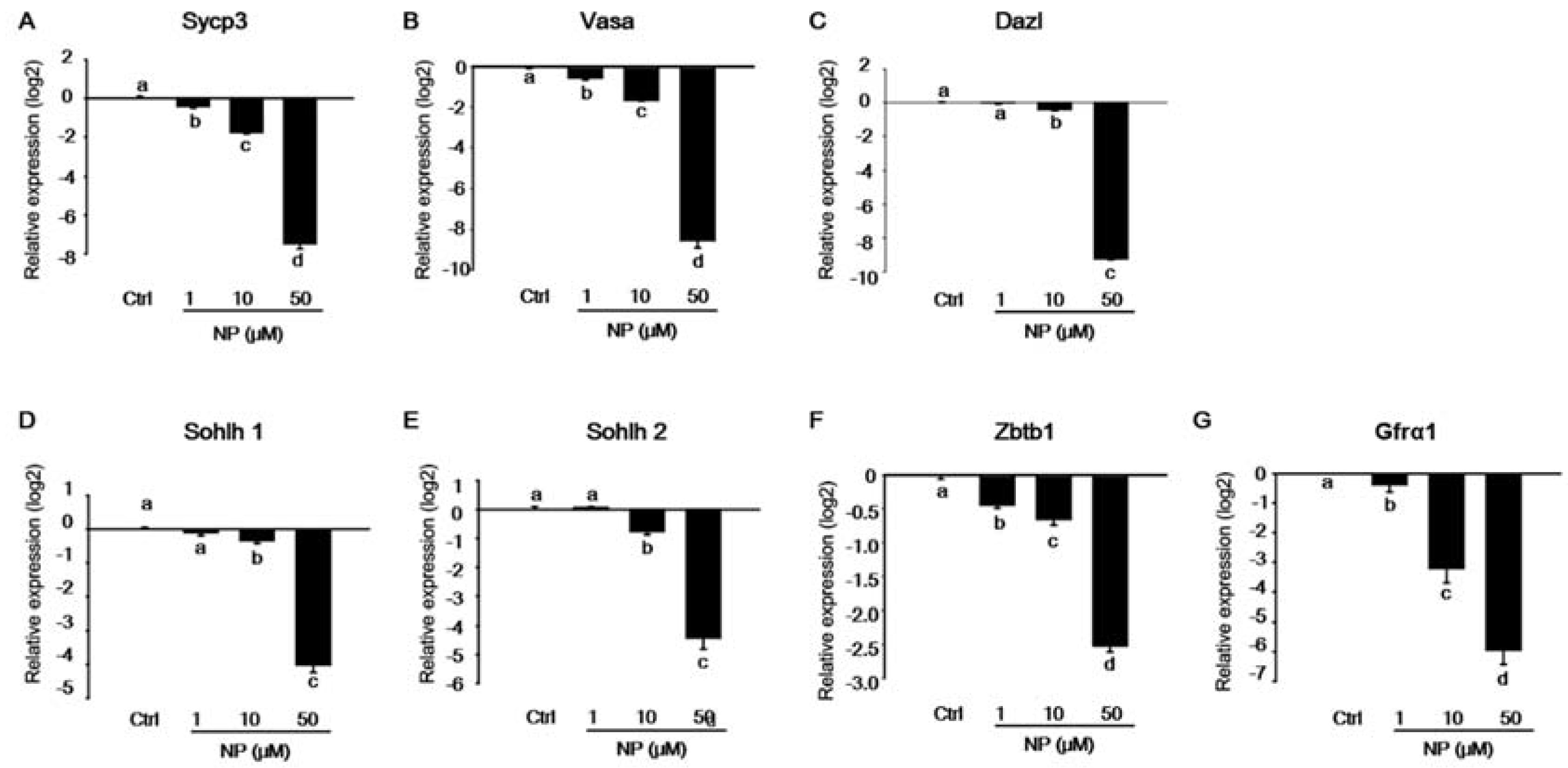
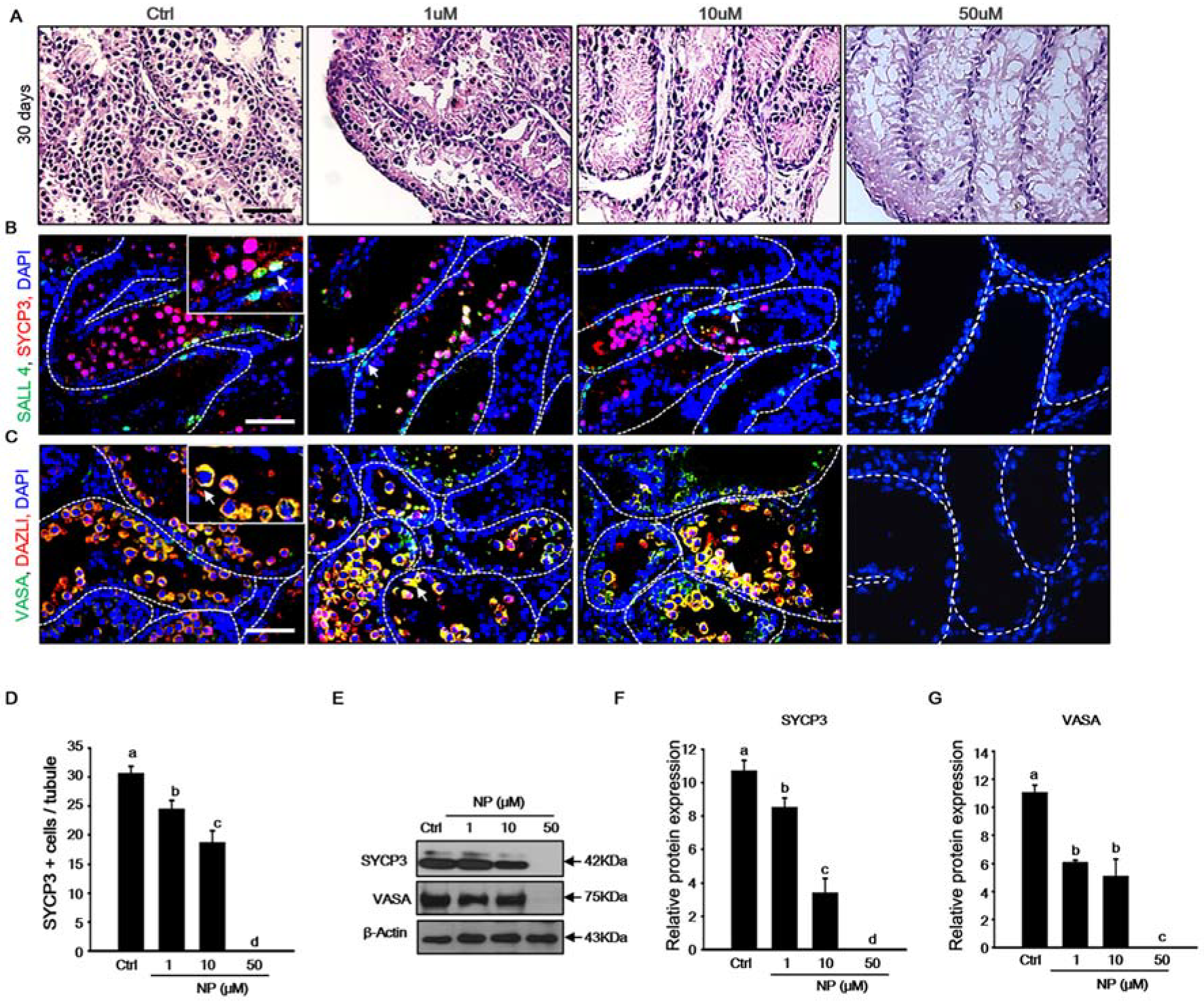
2.2. Effect of Nonylphenol on Germ Cells in MTFs
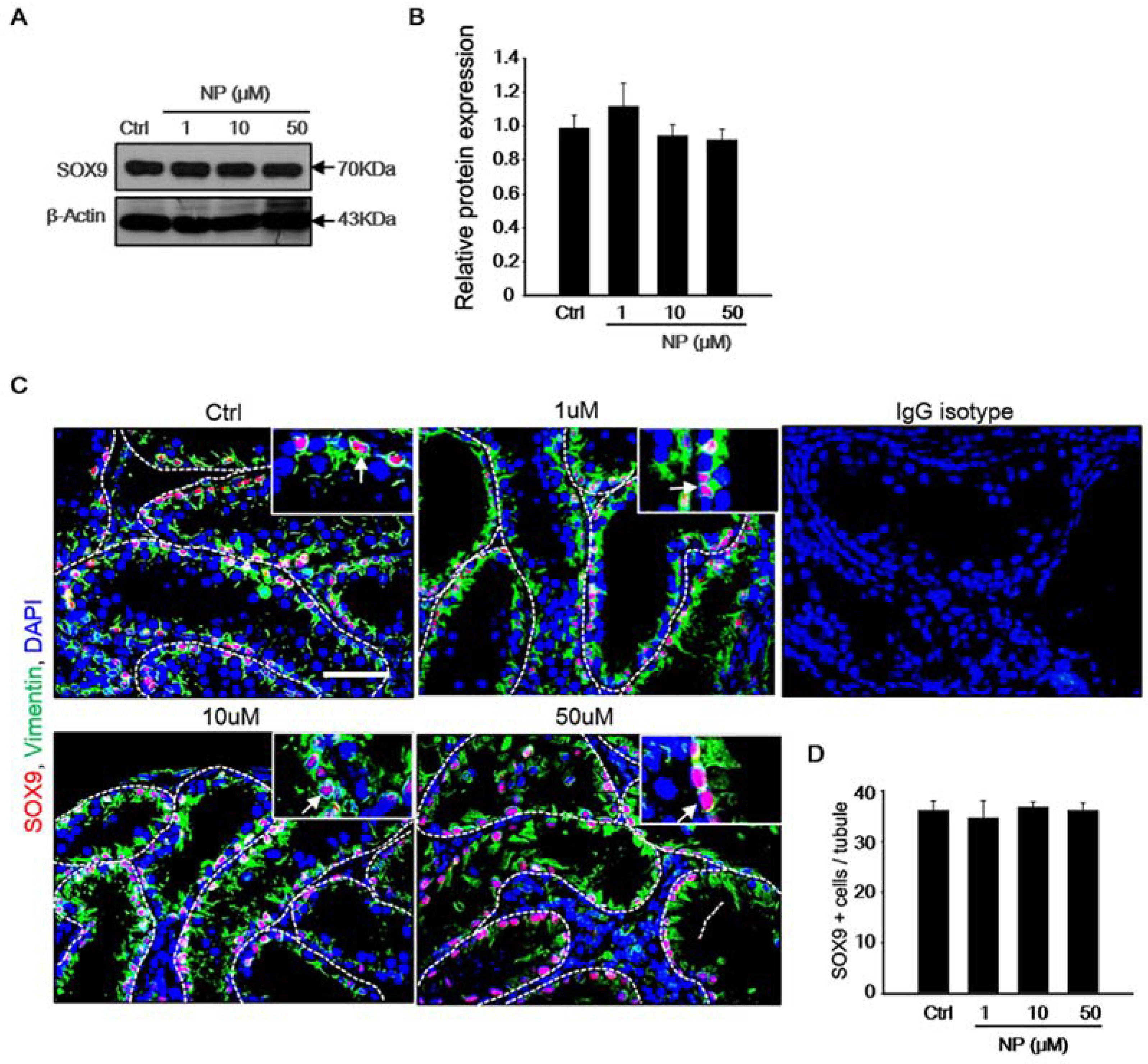
2.3. Effect of Nonylphenol on Sertoli Cells in MTF Culture
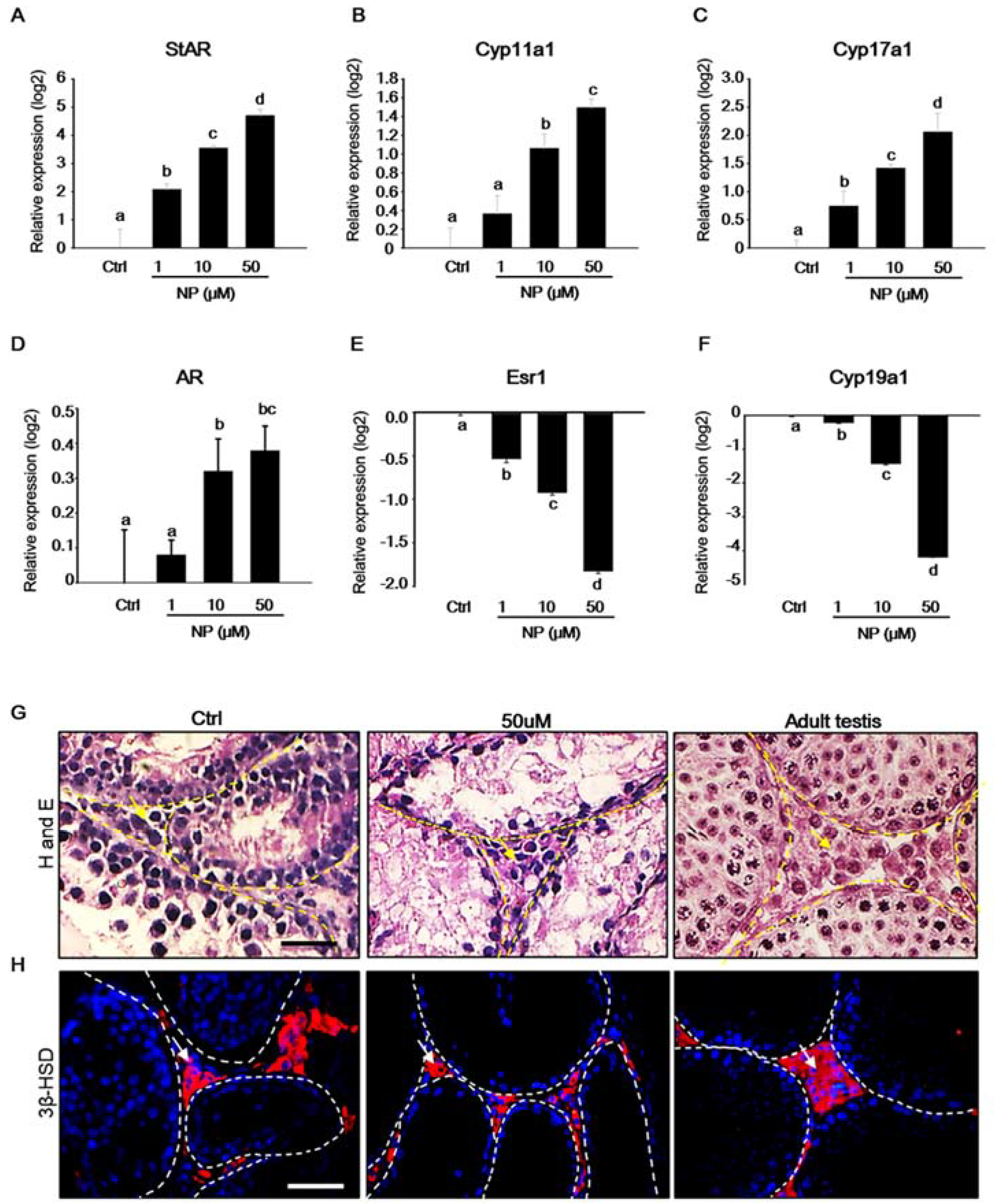
2.4. Effect of Nonylphenol on mRNA Expression of Steroidogenic Genes in MTFs
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Organ Culture Method and Test Compound
4.3. Histological Analysis and Immunostaining
4.4. RNA Isolation and Quantitative PCR
4.5. Immunoblotting
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Noorimotlagh, Z.; Haqhighi, N.J.; Ahmadimoghadam, M.; Rahim, F. An updated systematic review on the possible effect of nonylphenol on male fertility. Environ. Sci. Pollut. Int. 2017, 24, 3298–3314. [Google Scholar] [CrossRef] [PubMed]
- Miles-Richardson, S.R.; Pierens, S.L.; Nichols, K.M.; Kramer, V.J.; Snyder, E.M.; Snyder, S.A.; Render, J.A.; Fitzgerald, S.D.; Giesy, J.P. Effects of waterborne exposure to 4-nonylphenol and nonylphenol ethoxylate on secondary sex characteristics and gonads of fathead minnows (Pimephales promelas). Environ. Res. 1999, 80, 122–137. [Google Scholar] [CrossRef] [PubMed]
- Puy-Azurmendi, E.; Olivares, A.; Vallejo, A.; Ortiz-Zarragoitia, M.; Piña, B.; Zuloaga, O.; Cajaraville, M.P. Estrogenic effects of nonylphenol and octylphenol isomers in vitro by recombinant yeast assay (RYA) and in vivo with early life stages of zebrafish. Sci. Total Environ. 2014, 466, 1–10. [Google Scholar] [CrossRef]
- Jin, X.; Wang, Y.; Jin, W.; Rao, K.; Giesy, J.P.; Hollert, H.; Richardson, K.L.; Wang, Z. Ecological risk of nonylphenol in China surface waters based on reproductive fitness. Environ. Sci. Technol. 2014, 48, 1256–1262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cherry, N.; Labrèche, F.; Collins, J.; Tulandi, T. Occupational exposure to solvents and male infertility. Occup. Environ. Med. 2001, 58, 635–640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawaguchi, M.; Inoue, K.; Sakui, N.; Ito, R.; Izumi, S.; Makino, T.; Okanouchi, N.; Nakazawa, H. Stir bar sorptive extraction and thermal desorption-gas chromatography-mass spectrometry for the measurement of 4-nonylphenol and 4-tert-octylphenol in human biological samples. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2004, 799, 119–125. [Google Scholar] [CrossRef]
- Tan, B.L.; Mohd, M.A. Analysis of selected pesticides and alkylphenols in human cord blood by gas chromatograph-mass spectrometer. Talanta 2003, 61, 385–391. [Google Scholar] [CrossRef]
- Shekhar, S.; Sood, S.; Showkat, S.; Lite, C.; Chandrasekhar, A.; Vairamani, M.; Barathi, S.; Santosh, W. Detection of phenolic endocrine disrupting chemicals (EDCs) from maternal blood plasma and amniotic fluid in Indian population. Gen. Comp. Endocrinol. 2017, 241, 100–107. [Google Scholar] [CrossRef]
- Chen, G.W.; Ding, W.H.; Ku, H.Y.; Chao, H.R.; Chen, H.Y.; Huang, M.C.; Wang, S.L. Alkylphenols in human milk and their relations to dietary habits in central Taiwan. Food Chem. Toxicol. 2010, 48, 1939–1944. [Google Scholar] [CrossRef]
- Zhou, F.; Zhang, L.; Liu, A.; Shen, Y.; Yuan, J.; Yu, X.; Feng, X.; Xu, Q.; Cheng, C. Measurement of phenolic environmental estrogens in human urine samples by HPLC-MS/MS and primary discussion the possible linkage with uterine leiomyoma. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2013, 938, 80–85. [Google Scholar] [CrossRef]
- Otaka, H.; Yasuhara, A.; Morita, M. Determination of bisphenol A and 4-nonylphenol in human milk using alkaline digestion and cleanup by solid-phase extraction. Anal. Sci. 2003, 19, 1663–1666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ademollo, N.; Ferrara, F.; Delise, M.; Fabietti, F.; Funari, E. Nonylphenol and Octylphenol in Human Breast Milk. Environ. Int. 2008, 34, 984–987. [Google Scholar] [CrossRef] [PubMed]
- Ko, D.S.; Lee, D.R.; Song, H.; Kim, J.H.; Lim, C.K. Octylphenol and nonylphenol affect decidualization of human endometrial stromal cells. Reprod. Toxicol. 2019, 89, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Oh, S.; Gye, M.C.; Shin, I. Comparative toxicological evaluation of nonylphenol and nonylphenol polyethoxylates using human keratinocytes. Drug Chem. Toxicol. 2018, 41, 486–491. [Google Scholar] [CrossRef]
- Lukac, N.; Lukacova, J.; Pinto, B.; Knazicka, Z.; Tvrda, E.; Massanyi, P. The effect of nonylphenol on the motility and viability of bovine spermatozoa in vitro. J. Environ. Sci. Health 2013, 48, 973–979. [Google Scholar] [CrossRef]
- Uguz, C.; Varisli, O.; Agca, C.; Evans, T.; Agca, Y. In vitro effects of nonylphenol on motility, mitochondrial, acrosomal and chromatin integrity of ram and boar spermatozoa. Andrologia 2015, 47, 910–919. [Google Scholar]
- Uguz, C.; Varisli, O.; Agca, C.; Agca, Y. Effects of nonylphenol on motility and subcellular elements of epididymal rat sperm. Reprod. Toxicol. 2009, 28, 542–549. [Google Scholar] [CrossRef]
- De Jager, C.; Bornman, M.S.; van der Horst, G. The effect of p-nonylphenol, an environmental toxicant with oestrogenic properties, on fertility potential in adult male rats. Andrologia 1999, 31, 99–106. [Google Scholar] [CrossRef]
- Gong, Y.; Wu, J.; Huang, Y.; Shen, S.; Han, X. Nonylphenol induces apoptosis in rat testicular Sertoli cells via endoplasmic reticulum stress. Toxicol. Lett. 2009, 186, 84–95. [Google Scholar] [CrossRef]
- Gong, Y.; Han, X.D. Nonylphenol-induced oxidative stress and cytotoxicity in testicular Sertoli cells. Reprod. Toxicol. 2006, 22, 623–630. [Google Scholar] [CrossRef]
- Cardinali, M.; Maradonna, F.; Olivotto, I.; Bortoluzzi, G.; Mosconi, G.; Polzonetti-Magni, A.M.; Carnevali, O. Temporary impairment of reproduction in freshwater teleost exposed to nonylphenol. Reprod. Toxicol. 2004, 18, 597–604. [Google Scholar] [CrossRef] [PubMed]
- Duan, P.; Hu, C.; Butler, H.J.; Quan, C.; Chen, W.; Huang, W.; Tang, S.; Zhou, W.; Yuan, M.; Shi, Y.; et al. Effects of 4-nonylphenol on spermatogenesis and induction of testicular apoptosis through oxidative stress-related pathways. Reprod. Toxicol. 2016, 62, 27–38. [Google Scholar] [CrossRef] [Green Version]
- Sato, T.; Katagiri, K.; Gohbara, A.; Inoue, K.; Ogonuki, N.; Ogura, A.; Kubota, Y.; Ogawa, T. In vitro production of functional sperm in cultured neonatal mouse testes. Nature 2011, 471, 504–507. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, N.; Merry, G.E.; Inselman, A.L.; Sloper, D.T.; Del Valle, P.L.; Sato, T.; Ogawa, T.; Hansen, D.K. Evaluation of Culture Time and Media in an In Vitro Testis Organ Culture System. Birth. Defects. Res. 2017, 109, 465–474. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, N.; Sloper, D.; Del Valle, P.L. Evaluation of an in vitro mouse testis organ culture system for assessing male reproductive toxicity. Birth. Defects. Res. 2019, 111, 70–77. [Google Scholar] [CrossRef]
- Park, H.J.; Lee, W.Y.; Zhang, M.; Hong, K.H.; Park, C.; Kim, J.H.; Song, H. Evaluation of Resmethrin Toxicity to Neonatal Testes in Organ Culture. Toxicol. Sci. 2020, 173, 53–64. [Google Scholar] [CrossRef]
- Zhang, T.; Zhou, Y.; Li, L.; Zhao, Y.; De Felici, M.; Reiter, R.J.; Shen, W. Melatonin protects prepuberal testis from deleterious effects of bisphenol A or diethylhexyl phthalate by preserving H3K9 methylation. J. Pineal Res. 2018, 65, 12497. [Google Scholar] [CrossRef] [PubMed]
- Lopes, F.; Smith, R.; Nash, S.; Mitchell, R.T.; Spears, N. Irinotecan metabolite SN38 results in germ cell loss in the testis but not in the ovary of prepubertal mice. Mol. Hum. Reprod. 2016, 22, 745–755. [Google Scholar] [CrossRef]
- Drumond, A.L.; Meistrich, M.L.; Chiarini-Garcia, H. Spermatogonial morphology and kinetics during testis development in mice: A high-resolution light microscopy approach. Reproduction 2011, 142, 145–155. [Google Scholar] [CrossRef] [Green Version]
- Stanley, E.; Lin, C.Y.; Jin, S.; Liu, J.; Sottas, C.M.; Ge, R.; Zirkin, B.R.; Chen, H. Identification, proliferation, and differentiation of adult Leydig stem cells. Endocrinology 2012, 153, 5002–5010. [Google Scholar] [CrossRef] [Green Version]
- Smith, L.B.; Walker, W.H. The regulation of spermatogenesis by androgens. Semin. Cell. Dev. Biol. 2014, 30, 2–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ying, F.; Ding, C.; Ge, R.; Wang, X.; Li, F.; Zhang, Y.; Zeng, Q.; Yu, B.; Ji, R.; Han, X. Comparative evaluation of nonylphenol isomers on steroidogenesis of rat Leydig Cells. Toxicol. In Vitro 2012, 26, 1114–1121. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.J.; Wang, K.L.; Wang, S.W.; Hwang, G.S.; Mao, I.F.; Chen, M.L.; Wang, P.S. Differential effects of nonylphenol on testosterone secretion in rat Leydig cells. Toxicology 2010, 268, 1–7. [Google Scholar] [CrossRef]
- Chang, H.; Gao, F.; Guillou, F.; Taketo, M.M.; Huff, V.; Behringer, R.R. Wt1 negatively regulates beta-catenin signaling during testis development. Development 2008, 135, 1875–1885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bigsby, R.; Chapin, R.E.; Daston, G.P.; Davis, B.J.; Gorski, J.; Gray, L.E.; Howdeshell, K.L.; Zoeller, R.T.; vom Saal, F.S. Evaluating the effects of endocrine disruptors on endocrine function during development. Environ. Health. Perspect. 1999, 107, 613–618. [Google Scholar]
- Mikkilä, T.F.; Toppari, J.; Paranko, J. Effects of neonatal exposure to 4-tert-octylphenol, diethylstilbestrol, and flutamide on steroidogenesis in infantile rat testis. Toxicol. Sci. 2006, 91, 456–466. [Google Scholar] [CrossRef] [Green Version]
- Lee, P.C. Disruption of male reproductive tract development by administration of the xenoestrogen, nonylphenol, to male newborn rats. Endocrine 1998, 9, 105–111. [Google Scholar] [CrossRef]
- Huang, Q.; Bu, L.; Yang, T.; Yang, Y.; Huang, S.; Yang, J.; Guo, Y.; Liu, C. Effects of nonylphyenol administration on serum, liver and testis estrogen metabolism. Chemosphere 2019, 235, 543–549. [Google Scholar] [CrossRef]
- Duan, P.; Hu, C.; Butler, H.J.; Quan, C.; Chen, W.; Huang, W.; Tang, S.; Zhou, W.; Yuan, M.; Shi, Y.; et al. 4-Nonylphenol induces disruption of spermatogenesis associated with oxidative stress-related apoptosis by targeting p53-Bcl-2/Bax-Fas/FasL signaling. Environ. Toxicol. 2017, 32, 739–753. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Wang, F.; Gong, Y.; Li, D.; Sha, J.; Huang, X.; Han, X. Proteomic analysis of changes induced by nonylphenol in Sprague-Dawley rat Sertoli cells. Chem. Res. Toxicol. 2009, 22, 668–675. [Google Scholar] [CrossRef]
- Choi, M.S.; Park, H.J.; Oh, J.H.; Lee, E.H.; Park, S.M.; Yoon, S. Nonylphenol-induced apoptotic cell death in mouse TM4 Sertoli cells via the generation of reactive oxygen species and activation of the ERK signaling pathway. J. Appl. Toxicol. 2014, 34, 628–636. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Wang, X.; Wang, Y.; Zhang, L.; Xu, B.; Lv, L.; Cui, X.; Gao, F. Wt1 is involved in leydig cell steroid hormone biosynthesis by regulating paracrine factor expression in mice. Biol. Reprod. 2014, 90, 71. [Google Scholar] [CrossRef] [PubMed]
- Ohyama, K.; Ohta, M.; Hosaka, Y.Z.; Tanabe, Y.; Ohyama, T.; Yamano, Y. Expression of anti-Mullerian hormone and its type II receptor in germ cells of maturing rat testis. Endocr. J. 2015, 62, 997–1006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jambor, T.; Lukáčová, J.; Tvrdá, E.; Kňažická, Z.; Forgács, Z.; Lukáč, N. The Impact of 4-Nonylphenol on the Viability and Hormone Production of Mouse Leydig Cells. Folia Biol. 2016, 62, 34–39. [Google Scholar]
- Chang, Y.C.; Wang, K.L.; Hsu, R.L.; Jian, C.Y.; Hsia, S.M.; Wang, P.S. Effects of Nonylphenol on the Production of Testosterone by Rat Leydig Cells. Biol. Reprod. 2010, 83, 525. [Google Scholar] [CrossRef]
- Gong, Y.I.; Han, X.D. Effect of Nonylphenol on Steroidogenesis of Rat Leydig Cells. J. Environ. Sci. Health B 2006, 41, 705–715. [Google Scholar] [CrossRef]
- Kazemi, S.; Feizi, F.; Aghapour, F.; Joorsaraee, G.A.; Moghadamnia, A.A. Histopathology and Histomorphometric Investigation of Bisphenol A and Nonylphenol on the Male Rat Reproductive system. N. Am. J. Med. Sci. 2016, 8, 215–221. [Google Scholar]
- Bartke, A.; Williams, K.I.; Dalterio, S. Effects of estrogens on testicular testosterone production in vitro. Biol. Reprod. 1977, 17, 645–649. [Google Scholar] [CrossRef] [Green Version]
- Bonefeld-Jørgensen, E.C.; Long, M.; Hofmeister, M.V.; Vinggaard, A.M. Endocrine-disrupting Potential of Bisphenol A, Bisphenol A Dimethacrylate, 4-n-nonylphenol, and 4-n-octylphenol in Vitro. Environ. Health Perspect. 2007, 115, 69–76. [Google Scholar] [CrossRef] [Green Version]
- Guiguen, Y.; Fostier, A.; Piferrer, F.; Chang, C.F. Ovarian Aromatase and Estrogens: A Pivotal Role for Gonadal Sex Differentiation and Sex Change in Fish. Gen. Comp. Endocrinol. 2010, 165, 352–366. [Google Scholar] [CrossRef]
- Soares, A.; Guieysse, B.; Jefferson, B.; Cartmell, E.; Lester, J.N. Nonylphenol in the Environment: A Critical Review on Occurrence, Fate, Toxicity and Treatment in Wastewaters. Environ. Int. 2008, 34, 1033–1049. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.; Chen, Q.; He, N.; Diao, P.P.; Jia, L.X.; Duan, S.S. Effect of Environmentally-Relevant Concentrations of Nonylphenol on Sexual Differentiation in Zebrafish: A Multi-Generational Study. Sci. Rep. 2017, 7, 42907. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Liu, X.; Wang, H.; Wu, T.; Hu, X.; Qin, F.; Wang, Z. Expression of Two Cytochrome P450 Aromatase Genes Is Regulated by Endocrine Disrupting Chemicals in Rare Minnow Gobiocypris Rarus Juveniles. Comp. Biochem. Physiol. C. Toxicol. Pharmacol. 2010, 152, 313–320. [Google Scholar]
- Christiansen, T.; Korsgaard, B.; Jespersen, Å. Induction of vitellogenin synthesis by. nonylphenol and 17β-estradiol and effects on the testicular structure in the eelpout Zoarces viviparus. Environ. Res. 1998, 46, 141–144. [Google Scholar] [CrossRef]
- Ma, H.; Yao, Y.; Wang, C.; Zhang, L.; Cheng, L.; Wang, Y.; Wang, T.; Liang, E.; Jia, H.; Ye, Q.; et al. Transcription Factor Activity of Estrogen Receptor α Activation Upon Nonylphenol or Bisphenol A Treatment Enhances the in Vitro Proliferation, Invasion, and Migration of Neuroblastoma Cells. Oncol. Targets Ther. 2016, 9, 3451–3463. [Google Scholar] [PubMed] [Green Version]
- Liu, X.; Nie, S.; Yu, Q.; Wang, X.; Huang, D.; Xie, M. Downregulation of Steroid Hormone Receptor Expression and Activation of Cell Signal Transduction Pathways Induced by a Chiral Nonylphenol Isomer in Mouse Sertoli TM4 Cells. Environ. Toxicol. 2017, 32, 469–476. [Google Scholar] [CrossRef]
- Duan, P.; Hu, C.; Quan, C.; Yu, T.; Huang, W.; Chen, W.; Tang, S.; Shi, Y.; Martin, F.L.; Yang, K. 4-Nonylphenol induces autophagy and attenuates mTOR-p70S6K/4EBP1 signaling by modulating AMPK activation in Sertoli cells. Toxicol. Lett. 2017, 267, 21–31. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Nie, S.; Huang, D.; Xie, M. Mitogen-activated protein kinase and Akt pathways are involved in 4-n-nonyphenol induced apoptosis in mouse Sertoli TM4 cells. Environ. Toxicol. Pharmacol. 2015, 39, 815–824. [Google Scholar] [CrossRef]
- Chapin, R.E.; Winton, T.; Nowland, W.; Danis, N.; Kumpf, S.; Johnson, K.; Coburn, A.; Stukenborg, J.B. Lost in translation: The search for an in vitro screen for spermatogenic toxicity. Birth Defects Res. B Dev. Reprod. Toxicol. 2016, 107, 225–242. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C (T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | Company | Catalog Number | Dilution |
|---|---|---|---|
| VASA | Abcam | ab13840 | 1:300 |
| SALL4 | Abcam | ab57577 | 1:300 |
| Sox9 | Abcam | Ab185230 | 1:200 |
| Vimentin | Santa Cruz Biotech | sc-373717 | 1:300 |
| SYCP3 | Abcam | ab15093 | 1:300 |
| DAZL | BIO-RAD | MCA2336 | 1:200 |
| 3βHSD | Santa Cruz Biotech | sc30820 | 1:200 |
| βActin | Santa Cruz Biotech | ab13840 | 1:1000 |
| Gene | Forward Primer | Reverse Primer |
|---|---|---|
| Sycp3 | 5′-CAGATGCTTCGAGGGTGTG-3′ | 5′-AAGGTGGCTTCCCAGATTTC-3′ |
| Dazl | 5′-GTCGAAGGGCTATGGATTTG-3′ | 5′-ACGTGGCTGCACATGATAAG-3 |
| Vasa | 5′-CCGCATGGCTAGAAGAGATT-3 | 5′-TTCCTCGTGTCAACAGATGC-3 |
| Zbtb16 | 5′-CCACCTTCGCTCACATACAG-3′ | 5′-TTGCCACAGCCATTACACTC-3′ |
| Sohlh1 | 5′-CATCTGCTGTGTCTCGGGTA-3′ | 5′-GCTGGAAGACTCTGGCTCAC-3′ |
| Sohlh2 | 5′-TGAGACGAGAACGCATCAAG-3′ | 5′-CCTCTGTGATGTGGCTGAGA-3′ |
| AR | 5′-GGCGGTCCTTCACTAATGTC-3′ | 5′-GACAGGTGCCTCATCCTCAC -3′ |
| ERα | 5′-GCACAAGCGTCAGAGAGATG-3′ | 5′-AGGACAAGGCAGGGCTATTC-3′ |
| Cyp19α1 | 5′-TTGAGACGATTCCAGGTGAAG-3′ | 5′-ATTTCCACAAGGTGCCTGTC-3′ |
| Cyp17α1 | 5′-TCCAGCATTGGAGAGTTTGC-3′ | 5′-ATGAGATGGCTTCCTGTTGG-3′ |
| Cyp11α1 | 5′-GACAATGGTTGGCTAAACCTG-3′ | 5′-GGGTCCACGATGTAAACTGAC-3′ |
| Star | 5′-TGGGCATACTCAACAACCAG-3′ | 5′-GTCTACCACCACCTCCAAGC-3′ |
| Sod1 | 5′-GGGTTCCACGTCCATCAGTA-3′ | 5′-AGTCACATTGCCCAGGTCTC-3′ |
| Cat | 5′-GCAGATACCTGTGAACTGTC-3′ | 5′-GTAGAATGTCCGCACCTGAG-3′ |
| Gpx | 5′-TTCGGACACCAGGAGAATGG-3′ | 5′-TAAAGAGCGGGTGAGCCTTC-3 |
| Bax | 5′-GCTGACATGTTTGCTGATGG-3′ | 5′-GATCAGCTCGGGCACTTTAG-3′ |
| Bok | 5′-CTGCCCCTGGAGGACGCTTG-3′ | 5′-CCGTCACCACAGGCTCCGAC-3′ |
| Fas | 5′-CTGATCCTCATTCCCGTACC-3′ | 5′-ATCATTGGCACCTCTTCAGC -3′ |
| Traf3 | 5′-AGAGTGAGTTGAGTGCACACTT-3′ | 5′-TACCGCGGAGCTGGCCTCAT-3′ |
| Wt1 | 5′-ATCCCAGGCAGGAAAGTGTG-3′ | 5′-GTGCTGTCTTGGAAGTCGGA-3′ |
| Amh | 5′-CCTGGAGGAAGTGACATGG-3′ | 5′-CAGGGTAGAGCACCAGCAG-3′ |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, H.-J.; Zhang, M.; Lee, W.-Y.; Hong, K.-H.; Do, J.T.; Park, C.; Song, H. Toxic Effects of Nonylphenol on Neonatal Testicular Development in Mouse Organ Culture. Int. J. Mol. Sci. 2020, 21, 3491. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21103491
Park H-J, Zhang M, Lee W-Y, Hong K-H, Do JT, Park C, Song H. Toxic Effects of Nonylphenol on Neonatal Testicular Development in Mouse Organ Culture. International Journal of Molecular Sciences. 2020; 21(10):3491. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21103491
Chicago/Turabian StylePark, Hyun-Jung, Mingtian Zhang, Won-Young Lee, Kwon-Ho Hong, Jeong Tae Do, Chankyu Park, and Hyuk Song. 2020. "Toxic Effects of Nonylphenol on Neonatal Testicular Development in Mouse Organ Culture" International Journal of Molecular Sciences 21, no. 10: 3491. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21103491