Effect of Proton Pump Inhibitors on Colorectal Cancer


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Effect of PPIs on Fecal pH and Serum Gastrin
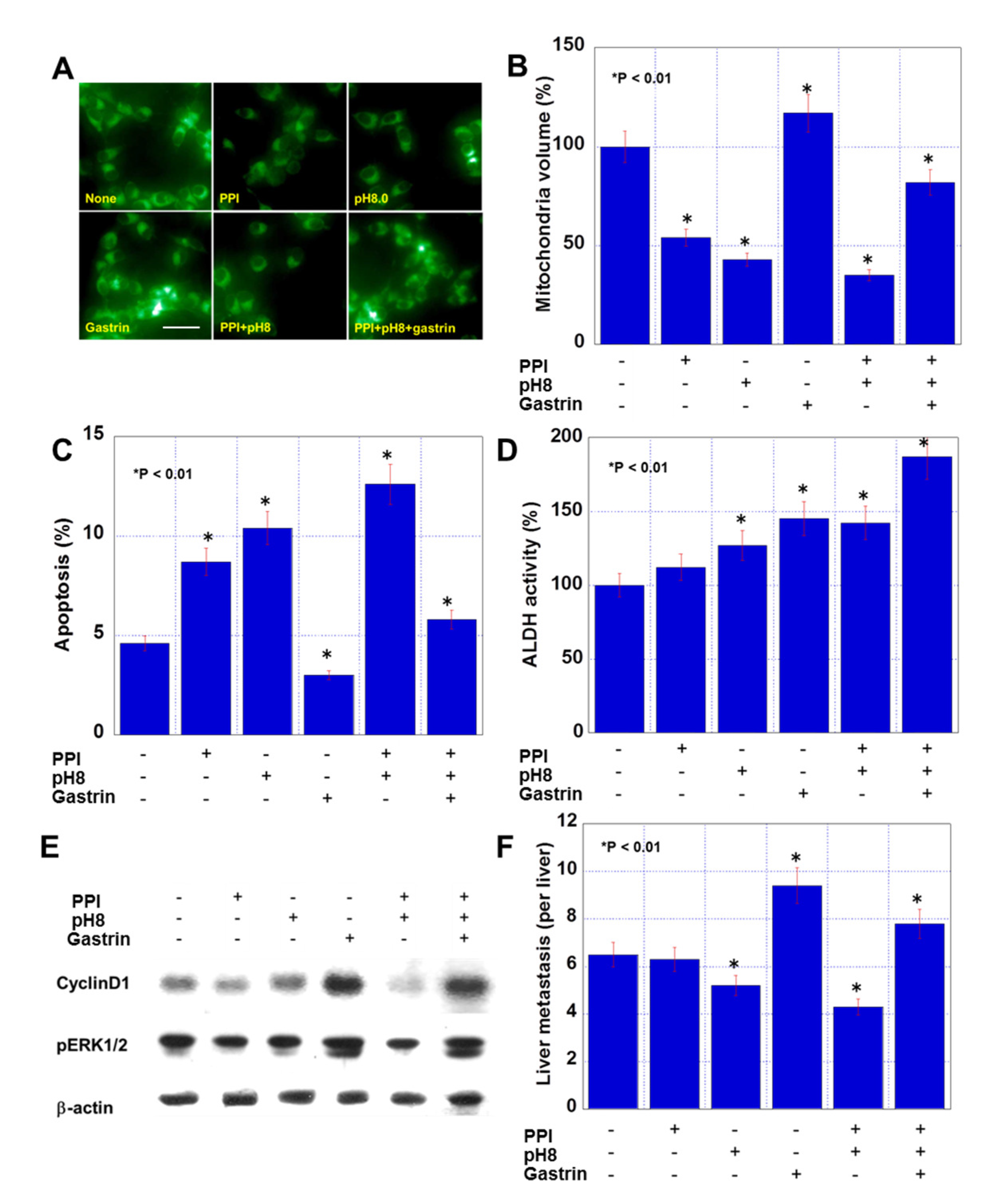
2.2. Effect of PPI, pH, and Gastrin on CMT Mouse Colon Cancer Cells
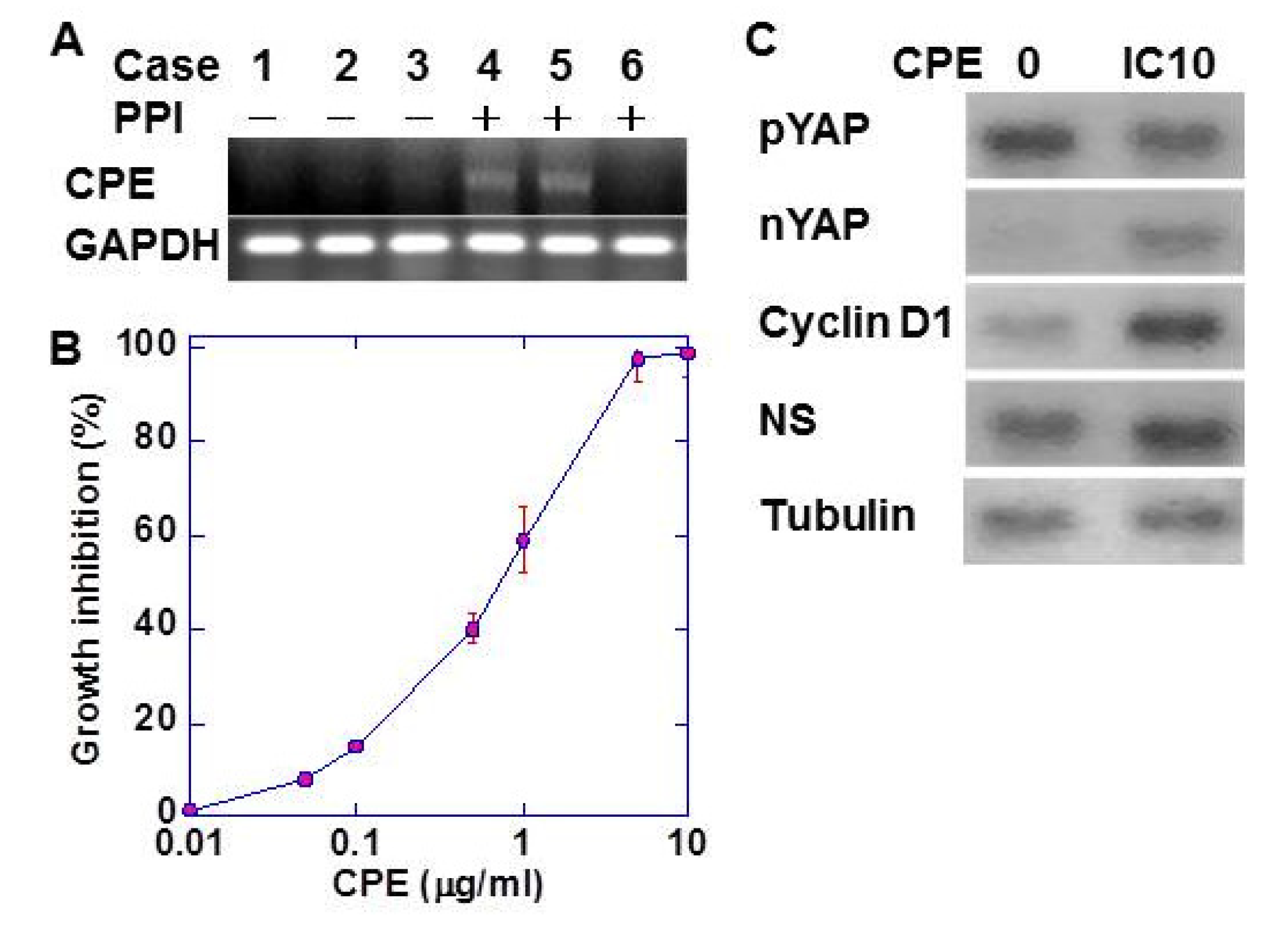
2.3. Effect of CPE on CMT93 Cells
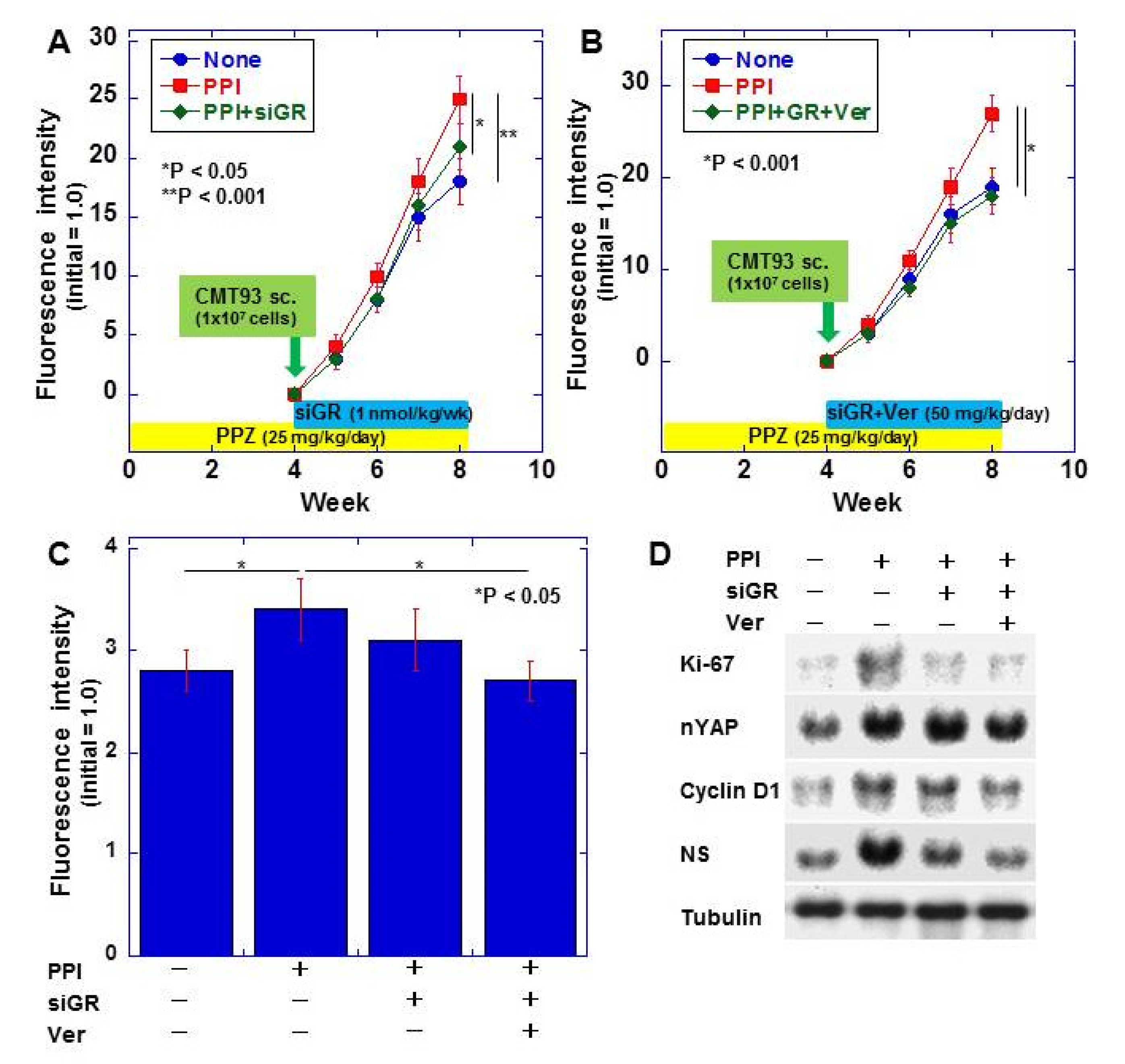
2.4. Effect of PPIs on CMT93 Mouse Tumors
3. Discussion
4. Materials and Methods
4.1. Cell Line
4.2. Assessment of Cell Growth and Apoptosis
4.3. Chamber Invasion Assay
4.4. Animals
4.5. Animal Models
4.6. In Vivo Imaging of Tumor
4.7. Stool pH
4.8. SiRNA
4.9. Reverse Transcription-Polymerase Chain Reaction (RT-PCR)
4.10. Western Blotting Analysis
4.11. Enzyme-Linked Immunosorbent Assay (ELISA)
4.12. Mitochondrial Volume
4.13. Aldehyde Dehydrogenase (ALDH) Activity
4.14. Surgical Specimens
4.15. Bacterial DNA Amplification
4.16. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fox, R.K.; Muniraj, T. Pharmacologic Therapies in Gastrointestinal Diseases. Med. Clin. N. Am. 2016, 100, 827–850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scarpignato, C.; Gatta, L.; Zullo, A.; Blandizzi, C.; the SIF-AIGO-FIMMG Group; Italian Society of Pharmacology; the Italian Association of Hospital Gastroenterologists; the Italian Federation of General Practitioners. Effective and safe proton pump inhibitor therapy in acid-related diseases—A position paper addressing benefits and potential harms of acid suppression. BMC Med. 2016, 14, 179. [Google Scholar] [CrossRef]
- Mori, S.; Kadochi, Y.; Luo, Y.; Fujiwara-Tani, R.; Nishiguchi, Y.; Kishi, S.; Fujii, K.; Ohmori, H.; Kuniyasu, H. Proton pump inhibitor induced collagen expression in colonocytes is associated with collagenous colitis. World J. Gastroenterol. 2017, 23, 1586–1593. [Google Scholar] [CrossRef] [PubMed]
- Rao, C.; Patlolla, J.M.; Zhang, Y.; Li, Q.; Steele, V.E. Anti-carcinogenic properties of omeprazole against human colon cancer cells and azoxymethane-induced colonic aberrant crypt foci formation in rats. Int. J. Oncol. 2011, 40, 170–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, M.; Dhindsa, G.; Friedland, S.; Triadafilopoulos, G. Long-term use of proton pump inhibitors does not affect the frequency, growth, or histologic characteristics of colon adenomas. Aliment. Pharmacol. Ther. 2007, 26, 1051–1061. [Google Scholar] [CrossRef]
- Zeng, X.; Liu, L.; Zheng, M.; Sun, H.; Xiao, J.; Lu, T.; Huang, G.; Chen, P.; Zhang, J.; Zhu, F.; et al. Pantoprazole, an FDA-approved proton-pump inhibitor, suppresses colorectal cancer growth by targeting T-cell-originated protein kinase. Oncotarget 2016, 7, 22460–22473. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.-S.; Lee, H.J.; Park, J.-M.; Han, Y.-M.; Kangwan, N.; Oh, J.Y.; Lee, D.Y.; Hahm, K.B. Targeted molecular ablation of cancer stem cells for curing gastrointestinal cancers. Expert Rev. Gastroenterol. Hepatol. 2017, 11, 1059–1070. [Google Scholar] [CrossRef]
- Haastrup, P.F.; Thompson, W.; Søndergaard, J.; Jarbøl, D.E. Side Effects of Long-Term Proton Pump Inhibitor Use: A Review. Basic Clin. Pharmacol. Toxicol. 2018, 123, 114–121. [Google Scholar] [CrossRef] [Green Version]
- Waldum, H.L.; Sørdal, Ø.; Fossmark, R. Proton pump inhibitors (PPIs) may cause gastric cancer-clinical consequences. Scand. J. Gastroenterol. 2018, 53, 639–642. [Google Scholar] [CrossRef]
- Joshi, S.N.; Gardner, J.D. Gastrin and Colon Cancer: A Unifying Hypothesis. Dig. Dis. 1996, 14, 334–344. [Google Scholar] [CrossRef]
- Smith, J.P.; Stock, E.A.; Wotring, M.G.; McLaughlin, P.J.; Zagon, I.S. Characterization of the CCK-B/gastrin-like receptor in human colon cancer. Am. J. Physiol. Integr. Comp. Physiol. 1996, 271, R797–R805. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.M.; Watson, S. Gastrin and gastrin receptor activation: An early event in the adenoma-carcinoma sequence. Gut 2000, 47, 820–824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, P.; Walker, J.P.; Townsend, C.M.; Thompson, J.C. Role of gastrin and gastrin receptors on the growth of a transplantable mouse colon carcinoma (MC-26) in BALB/c mice. Cancer Res. 1986, 46, 1612–1616. [Google Scholar] [PubMed]
- Dial, M.S. Proton pump inhibitor use and enteric infections. Am. J. Gastroenterol. 2009, 104 (Suppl. S2), S10–S16. [Google Scholar] [PubMed]
- Villafuerte-Gálvez, J.A.; Kelly, C.P. Proton pump inhibitors and risk of Clostridium difficile infection. Curr. Opin. Gastroenterol. 2018, 34, 11–18. [Google Scholar] [CrossRef]
- Pant, C.; Madonia, P.; Minocha, A. Does PPI therapy predispose to Clostridium difficile infection? Nat. Rev. Gastroenterol. Hepatol. 2009, 6, 555–557. [Google Scholar] [CrossRef]
- Li, B.; Ma, H.; Wang, Z.; Lihong, L. When omeprazole met with asymptomatic Clostridium difficile colonization in a postoperative colon cancer patient. Medicine 2017, 96, 9089. [Google Scholar] [CrossRef]
- Hung, Y.-P.; Ko, W.-C.; Chou, P.-H.; Chen, Y.-H.; Lin, H.-J.; Liu, Y.-H.; Tsai, H.-W.; Lee, J.-C.; Tsai, P.-J. Proton-Pump Inhibitor Exposure AggravatesClostridium difficile—Associated Colitis: Evidence From a Mouse Model. J. Infect. Dis. 2015, 212, 654–663. [Google Scholar] [CrossRef] [Green Version]
- Nakashima, C.; Yamamoto, K.; Kishi, S.; Sasaki, T.; Ohmori, H.; Fujiwara-Tani, R.; Mori, S.; Kawahara, I.; Nishiguchi, Y.; Mori, T.; et al. Clostridium perfringens enterotoxin induces claudin-4 to activate YAP in oral squamous cell carcinomas. Oncotarget 2020, 11, 309–321. [Google Scholar] [CrossRef] [Green Version]
- Singh, P.; Owlia, A.; Varro, A.; Dai, B.; Rajaraman, S.; Wood, T. Gastrin gene expression is required for the proliferation and tumorigenicity of human colon cancer cells. Cancer Res. 1996, 56, 4111–4115. [Google Scholar]
- Smith, J.P.; Solomon, T.E. Effects of Gastrin, Proglumide, and Somatostatin on Growth of Human Colon Cancer. Gastroenterology 1988, 95, 1541–1548. [Google Scholar] [CrossRef]
- Smith, A.M.; Watson, S.A. Review article: Gastrin and colorectal cancer. Aliment. Pharmacol. Ther. 2000, 14, 1231–1247. [Google Scholar] [CrossRef] [PubMed]
- Song, L.-J.; Liu, R.-J.; Zeng, Z.; Alper, S.L.; Cui, H.-J.; Lu, Y.; Zheng, L.; Yan, Z.-W.; Fu, G.-H. Gastrin inhibits a novel, pathological colon cancer signaling pathway involving EGR1, AE2, and P-ERK. J. Mol. Med. 2012, 90, 707–718. [Google Scholar] [CrossRef] [PubMed]
- Huang, B.-P.; Lin, C.-H.; Chen, Y.-C.; Kao, S.-H. Expression of cholecystokinin receptors in colon cancer and the clinical correlation in Taiwan. Tumor Biol. 2015, 37, 4579–4584. [Google Scholar] [CrossRef]
- Beauchamp, R.D.; Townsend, C.M.; Singh, P.; Glass, E.J.; Thompson, J.C. Proglumide, A Gastrin Receptor Antagonist, Inhibits Growth of Colon Cancer and Enhances Survival in Mice. Ann. Surg. 1985, 202, 303–309. [Google Scholar] [CrossRef]
- Frattini, M.; Balestra, D.; Suardi, S.; Oggionni, M.; Alberici, P.; Radice, P.; Costa, A.; Daidone, M.G.; Leo, E.; Pilotti, S.; et al. Different Genetic Features Associated with Colon and Rectal Carcinogenesis. Clin. Cancer Res. 2004, 10, 4015–4021. [Google Scholar] [CrossRef] [Green Version]
- Nakata, H.; Wang, S.-L.; Chung, D.C.; Westwick, J.K.; Tillotson, L.G. Oncogenic ras induces gastrin gene expression in colon cancer. Gastroenterology 1998, 115, 1144–1153. [Google Scholar] [CrossRef]
- Hori, H.; Nakata, H.; Baba, H.; Genzo, I.; Yamada, H.; Chihara, K. Oncogenic ras induces Gastrin/CCKB receptor gene expression in human colon cancer cell lines LoVo and Colo320HSR. Gastroenterology 2003, 124, A133. [Google Scholar] [CrossRef] [Green Version]
- Jump, R.L.P.; Pultz, M.J.; Donskey, C.J. Vegetative Clostridium difficile Survives in Room Air on Moist Surfaces and in Gastric Contents with Reduced Acidity: A Potential Mechanism To Explain the Association between Proton Pump Inhibitors and C. difficile-Associated Diarrhea? Antimicrob. Agents Chemother. 2007, 51, 2883–2887. [Google Scholar] [CrossRef] [Green Version]
- Swietach, P.; Hulikova, A.; Patiar, S.; Vaughan-Jones, R.D.; Harris, A.L. Importance of Intracellular pH in Determining the Uptake and Efficacy of the Weakly Basic Chemotherapeutic Drug, Doxorubicin. PLoS ONE 2012, 7, e35949. [Google Scholar] [CrossRef]
- Hojo, M.; Asahara, T.; Nagahara, A.; Takeda, T.; Matsumoto, K.; Ueyama, H.; Matsumoto, K.; Asaoka, D.; Takahashi, T.; Nomoto, K.; et al. Gut Microbiota Composition Before and After Use of Proton Pump Inhibitors. Dig. Dis. Sci. 2018, 63, 2940–2949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biswal, S. Proton pump inhibitors and risk for Clostridium difficile associated diarrhea. Biomed. J. 2014, 37, 178. [Google Scholar] [CrossRef] [PubMed]
- Naito, Y.; Kashiwagi, K.; Takagi, T.; Andoh, A.; Inoue, R. Intestinal Dysbiosis Secondary to Proton-Pump Inhibitor Use. Digestion 2018, 97, 195–204. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Cresci, G.A.; Kirby, D.F. Proton Pump Inhibitors: Risks and Rewards and Emerging Consequences to the Gut Microbiome. Nutr. Clin. Pract. 2018, 33, 614–624. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Wang, Y.; Li, S.J. Lansoprazole induces apoptosis of breast cancer cells through inhibition of intracellular proton extrusion. Biochem. Biophys. Res. Commun. 2014, 448, 424–429. [Google Scholar] [CrossRef]
- Ihraiz, W.G.; Ahram, M.; Bardaweel, S.K. Proton pump inhibitors enhance chemosensitivity, promote apoptosis, and suppress migration of breast cancer cells. Acta Pharm. 2020, 70, 179–190. [Google Scholar] [CrossRef] [Green Version]
- Kuniyasu, H.; Yano, S.; Sasaki, T.; Sasahira, T.; Sone, S.; Ohmori, H. Colon cancer cell-derived high mobility group 1/amphoterin induces growth inhibition and apoptosis in macrophages. Am. J. Pathol. 2005, 166, 751–760. [Google Scholar] [CrossRef] [Green Version]
- Kuniyasu, H.; Oue, N.; Wakikawa, A.; Shigeishi, H.; Matsutani, N.; Kuraoka, K.; Ito, R.; Akashi, M.; Yasui, W. Expression of receptors for advanced glycation end-products (RAGE) is closely associated with the invasive and metastatic activity of gastric cancer. J. Pathol. 2002, 196, 163–170. [Google Scholar] [CrossRef]
- Sasaki, T.; Kitadai, Y.; Nakamura, T.; Kim, J.-S.; Tsan, R.Z.; Kuwai, T.; Langley, R.R.; Fan, M.; Kim, S.-J.; Fidler, I.J. Inhibition of Epidermal Growth Factor Receptor and Vascular Endothelial Growth Factor Receptor Phosphorylation on Tumor-Associated Endothelial Cells Leads to Treatment of Orthotopic Human Colon Cancer in Nude Mice. Neoplasia 2007, 9, 1066–1077. [Google Scholar] [CrossRef] [Green Version]
- Fidler, I.J. Orthotopic implantation of human colon carcinomas into nude mice provides a valuable model for the biology and therapy of metastasis. Cancer Metastasis Rev. 1991, 10, 229–243. [Google Scholar] [CrossRef]
- Luo, Y.; Kishi, S.; Sasaki, T.; Ohmori, H.; Fujiwara-Tani, R.; Mori, S.; Goto, K.; Nishiguchi, Y.; Mori, T.; Kawahara, I.; et al. Targeting claudin-4 enhances chemosensitivity in breast cancer. Cancer Sci. 2020, 111, 1840–1850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuwada, M.; Chihara, Y.; Luo, Y.; Li, X.; Nishiguchi, Y.; Fujiwara, R.; Sasaki, T.; Fujii, K.; Ohmori, H.; Fujimoto, K.; et al. Pro-chemotherapeutic effects of antibody against extracellular domain of claudin-4 in bladder cancer. Cancer Lett. 2015, 369, 212–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chihara, Y.; Fujimoto, K.; Kondo, H.; Moriwaka, Y.; Sasahira, T.; Hirao, Y.; Kuniyasu, H. Anti-Tumor Effects of Liposome-Encapsulated Titanium Dioxide in Nude Mice. Pathobiology 2007, 74, 353–358. [Google Scholar] [CrossRef] [PubMed]
- Kuniyasu, H.; Oue, N.; Sasahira, T.; Yi, L.; Moriwaka, Y.; Shimomoto, T.; Fujii, K.; Ohmori, H.; Yasui, W. Reg IV enhances peritoneal metastasis in gastric carcinomas. Cell Prolif. 2009, 42, 110–121. [Google Scholar] [CrossRef] [PubMed]
- Matsushima-Otsuka, S.; Fujiwara-Tani, R.; Sasaki, T.; Ohmori, H.; Nakashima, C.; Kishi, S.; Nishiguchi, Y.; Fujii, K.; Luo, Y.; Kuniyasu, H. Significance of intranuclear angiotensin-II type 2 receptor in oral squamous cell carcinoma. Oncotarget 2018, 9, 36561–36574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanabe, E.; Kitayoshi, M.; Fujii, K.; Ohmori, H.; Luo, Y.; Kadochi, Y.; Mori, S.; Fujiwara, R.; Nishiguchi, Y.; Sasaki, T.; et al. Fatty acids inhibit anticancer effects of 5-fluorouracil in mouse cancer cell lines. Oncol. Lett. 2017, 14, 681–686. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Labbe, R.G.; Ryu, S. Inhibitory Effects of Collagen on the PCR for Detection of Clostridium perfringens. Appl. Environ. Microbiol. 2000, 66, 1213–1215. [Google Scholar] [CrossRef] [Green Version]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sasaki, T.; Mori, S.; Kishi, S.; Fujiwara-Tani, R.; Ohmori, H.; Nishiguchi, Y.; Hojo, Y.; Kawahara, I.; Nakashima, C.; Fujii, K.; et al. Effect of Proton Pump Inhibitors on Colorectal Cancer. Int. J. Mol. Sci. 2020, 21, 3877. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21113877
Sasaki T, Mori S, Kishi S, Fujiwara-Tani R, Ohmori H, Nishiguchi Y, Hojo Y, Kawahara I, Nakashima C, Fujii K, et al. Effect of Proton Pump Inhibitors on Colorectal Cancer. International Journal of Molecular Sciences. 2020; 21(11):3877. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21113877
Chicago/Turabian StyleSasaki, Takamitsu, Shiori Mori, Shingo Kishi, Rina Fujiwara-Tani, Hitoshi Ohmori, Yukiko Nishiguchi, Yudai Hojo, Isao Kawahara, Chie Nakashima, Kiyomu Fujii, and et al. 2020. "Effect of Proton Pump Inhibitors on Colorectal Cancer" International Journal of Molecular Sciences 21, no. 11: 3877. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21113877